
Molecular and Cytological Identification of Wheat-Thinopyrum intermedium Partial Amphiploid Line 92048 with Resistance to Stripe Rust and Fusarium Head Blight
Abstract
:1. Introduction
2. Results
2.1. ND-FISH Analysis of 92048 by Multiple Oligo Probes
2.2. Molecular Marker Analysis
2.3. Identification of Resistance to Stripe Rust and FHB
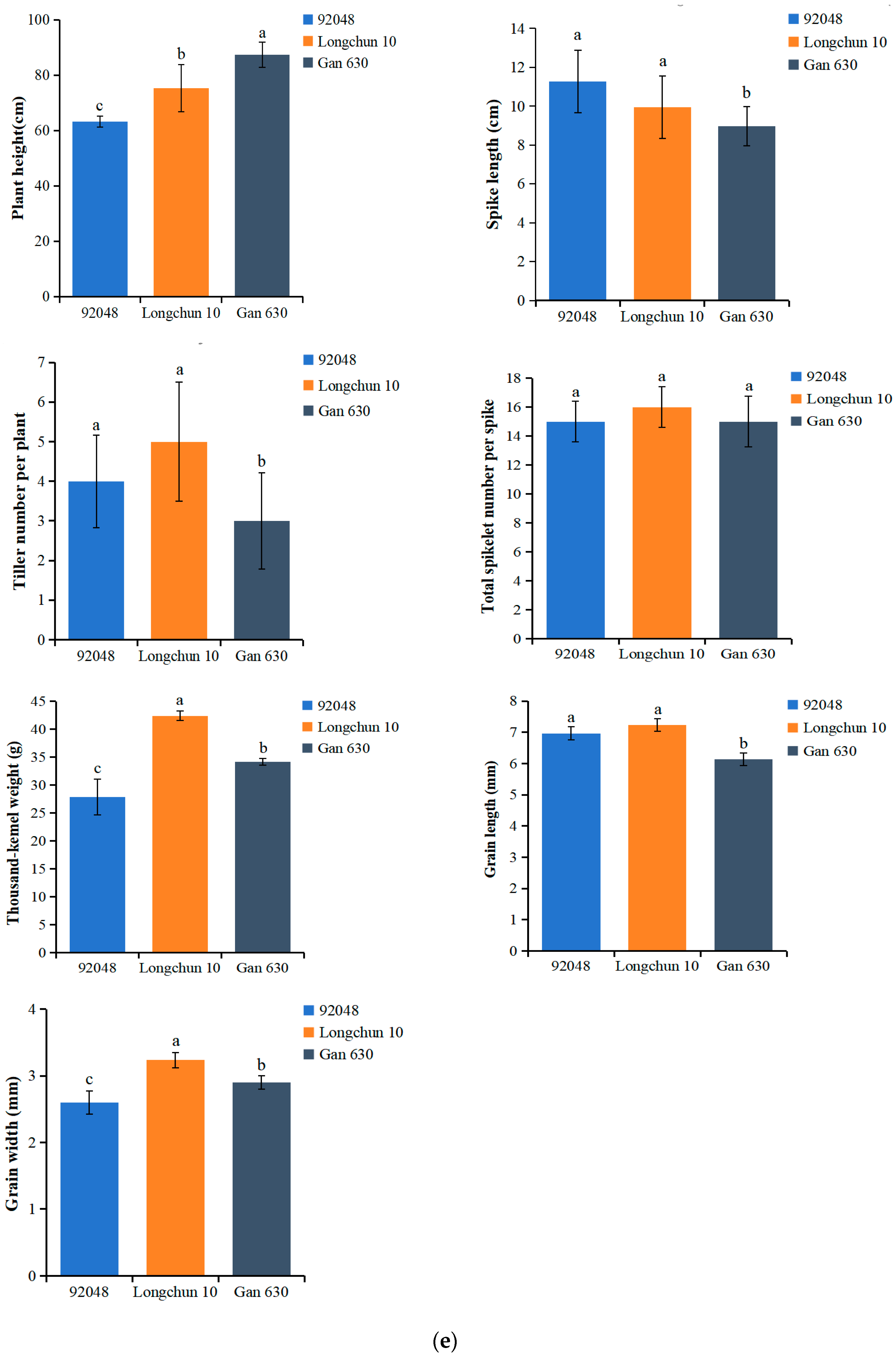
2.4. Assessment of Agronomic Performance
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Molecular Cytogenetic Analyses
4.3. Molecular Marker Analysis
4.4. Stripe Rust and FHB Resistance Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, S.K.; Xu, Z.R.; Xie, G.D.; Zhen, L.; Wang, L.E.; Guo, J.H.; Hou, P.; He, Z.H. The history of China’s food security in the past hundred years. J. Agric. 2018, 8, 186–192. [Google Scholar]
- He, Z.H.; Zhuang, Q.S.; Cheng, S.H.; Yu, Z.W.; Zhao, Z.D.; Liu, X. Wheat production and technology improvement in China. J. Agric. 2018, 8, 99–106. [Google Scholar]
- Zhang, X.J.; Han, D.J.; Zeng, Q.D.; Duan, Y.H.; Yuan, F.P.; Shi, J.D.; Wang, Q.L.; Wu, J.H.; Huang, L.; Kang, Z. Fine mapping of wheat stripe rust resistance gene Yr26 based on collinearity of wheat with Brachypodium distachyon and rice. PLoS ONE 2013, 8, e57885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L. Characteristics of wheat stripe rust and control measures. Anhui Agric. Sci. Bull. 2022, 28, 79–81. [Google Scholar]
- Niu, H.; Jiang, Y.M.; Niu, J.S. Research advances in the genetics and breeding of wheat (Triticum aestivum) resistance to fusarium head blight. J. Agric. Biotechnol. 2020, 28, 530–542. [Google Scholar]
- Liu, S.J.; Zhang, X.J.; Li, X.; Liu, C.; Bai, J.R.; Ren, Y.K.; Zheng, J.; Li, S.J.; Guo, H.J.; Mei, C.; et al. Development of specific STS markers for the sixth homologous group of Thinopyrum intermedium. Acta Pratacult. Sin. 2019, 28, 139–145. [Google Scholar]
- Mcintosh, R.; Yamazaki, Y.; Devos, K.M.; Dubcovsky, J.; Rogers, W.J.; Rogers, R. Catalogue of gene symbols for wheat. Annu. Wheat Newsl. 2003, 49, 246–282. [Google Scholar]
- Klymiuk, V.; Chawla, H.S.; Wiebe, K.; Ens, J.; Fatiukha, A.; Govta, L.; Fahima, T.; Ponzniak, C.J. Discovery of stripe rust resistance with incomplete dominance in wild emmer wheat using bulked segregant analysis sequencing. Commun. Biolog. 2022, 5, 826. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Zeng, S.M. Systems analysis of wheat stripe rust epidemics in China. Eur. J. Plant Pathol. 2008, 121, 425–438. [Google Scholar]
- Han, D.J.; Wang, Q.L.; Zhang, L.; Wei, G.R.; Zeng, Q.D.; Zhao, J.; Wang, X.J.; Huang, L.L.; Kang, Z.S. Evaluation of resistance of current wheat cultivars to stripe rust in Northwest China, North China, and the Middle and Lower Reaches of Changjiang River epidemic area. Sci. Agric. Sin. 2010, 43, 2889–2896. [Google Scholar]
- Mcintosh, R.A.; Mu, J.; Han, D.J.; Kang, Z.S. Wheat stripe rust resistance gene Yr24/Yr26: A retrospective review. Crop J. 2018, 6, 321–329. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Zhang, H.Z.; Bai, B.; Li, J.; Huang, L.; Xu, Z.B.; Chen, Y.X.; Liu, X.; Cao, T.J.; Li, M.M.; et al. Current Status and Strategies for Utilization of Stripe Rust Resistance Genes in Wheat Breeding Program of China. Sci. Agric. Sin. 2024, 57, 34–51. [Google Scholar]
- Zhan, H.X.; Chang, Z.J.; Li, G.R.; Jia, J.Q.; Guo, H.J.; Zhang, X.J.; Li, X.; Qiao, L.Y.; Yang, Z.J. Cytogenetic and molecular identification of a wheat (Triticum aestivum)-Thinopyrum intermedium introgression line with resistance to stripe rust. J. Agric. Biotechnol. 2014, 22, 841–845. [Google Scholar]
- Song, J.; Ma, D.; Yin, J.; Yang, L.; He, Y.; Zhu, Z.; Tong, H.; Chen, L.; Zhu, G.; Liu, Y.; et al. Genome-Wide Characterization and Expression Profiling of Squamosa Promoter Binding Protein-Like (SBP) Transcription Factors in Wheat (Triticum aestivum L.). Agronomy 2019, 9, 527. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, H.; Li, C.L.; Han, D.J.; Chen, Y.F. Detection and Analysis of Stripe Rust Resistance Genes in Adult Period in Wheat Varieties (Lines). Mol. Plant Breed. 2022, 20, 5044–5054. [Google Scholar]
- Liu, Y.K.; Tong, H.W.; Zhu, Z.W.; Chen, L.; Zou, J.; Zhang, Y.Q.; Gao, C.B. Progress in Research on Mechanism of Resistance to Fusarium Head Blight in Wheat. Sci. Agric. Sin. 2016, 49, 1476–1488. [Google Scholar]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Sylvie, C.; Anita, B.B. Fine mapping Fhb1, a major gene controlling Fusarium head blight resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, P.A.; Somers, D.J.; Anita, B.B. Mapping of Fhb2 on chromosome 6BS: A gene controlling Fusarium head blight field resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Chen, P.D.; Gill, B.S. Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to Fusarium head blight disease of wheat. Theor. Appl. Genet. 2008, 117, 1155–1166. [Google Scholar] [CrossRef]
- Cainong, J.C.; Bockus, W.W.; Feng, Y.; Chen, P.; Qi, L.L.; Sehgal, S.K.; Danilova, T.V.; Koo, D.H.; Friebe, B.; Gill, B.S. Chromosome engineering, mapping, and transferring of resistance to Fusarium head blight disease from Elymus tsukushiensis into wheat. Theor. Appl. Genet. 2015, 128, 1019–1027. [Google Scholar] [CrossRef]
- Wang, Y.W.; Feng, Y.G.; Liu, R.R.; Lu, C.T.; Cao, A.Z.; Zhang, R.Q. Introgression and Characterization of the Homologous Group 1 Chromosomes from Roegneria kamoji into Common Wheat. Curr. Biotechnol. 2021, 11, 567–573. [Google Scholar]
- Wang, H.G.; Sun, S.L.; Ge, W.Y.; Zhao, L.F.; Hou, B.Q.; Wang, K.; Lyu, Z.F.; Chen, L.Y.; Xu, S.S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus under lies Fusarium head blight resistance in wheat. Science 2020, 368, 5435. [Google Scholar] [CrossRef] [PubMed]
- Bonin, C.M.; Kolb, F.L. Resistance to Fusarium head blight and kernel damage in a winter wheat recombinant inbred line population. Crop Sci. 2009, 49, 1304–1312. [Google Scholar] [CrossRef]
- Cheng, W.H.; Yin, X.C.; Wu, D.L.; Xu, J.H.; Zhao, F.C.; Yang, Z.Y.; Shi, J.R. Progress in Biological Control of Fusarium head blight in Wheat. Jiangsu Agric. Sci. 2020, 48, 12–18. [Google Scholar]
- Li, L.L.; Yang, X.M.; Li, X.Q.; Dong, Y.C. Study and utilization of wild relatives of wheat in China. Rev. China Agric. Sci. Technol. 2000, 2, 73–76. [Google Scholar]
- Li, Z.S.; Rong, S.; Zhong, G.C.; Li, R.L. Studies on Distant Hybridization between Wheat and Elymus repens; Science Press: Beijing, China, 1985. [Google Scholar]
- Wang, Y.L. Molecular Cytogenetic Characterization of the Derivatives from Wheat-Thinopyrum intermedium-Psathyrostachys huashanica Hybrids with Resistance to Stripe Rust. Master’s Thesis, Sichuan Agricultural University, Yaan, China, 2015. [Google Scholar]
- Liu, J.; Chang, Z.J.; Zhang, X.J.; Yang, Z.J.; Li, X.; Jia, J.Q.; Zhan, H.X.; Guo, H.J.; Wang, J.M. Putative Thinopyrum intermedium-derived stripe rustresistance gene Yr50 maps on wheat chromosome arm 4BL. Theor. Appl. Genet. 2013, 126, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Friebe, B.; Jiang, J.M.; Gill, B.S.; Dyck, P.L. Radiation-induced nonhomoelogous wheat-Agropyron intermedium chromosomal translocations conferring resistance to leaf rust. Theor. Appl. Genet. 1993, 86, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Danilova, T.V.; Rouse, M.N.; Bowden, R.L.; Friebe, B.S.; Gill, B.; Pumphrey, M.O. Development and characterization of a compensating wheat-Thinopyrum intermedium robertsonian translocation with Sr44 resistance to stem rust (Ug99). Theor. Appl. Genet. 2013, 126, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.G.; Luo, H.Y.; Chang, Z.J.; Zhang, H.Y.; Zhang, M.; Ren, Z.L. Characterization and chromosomal location of Pm40 in common wheat: A new gene for resistance to fusarium head blight derived from Elytrigia Intermedium. Theor. Appl. Genet. 2009, 118, 1059–1064. [Google Scholar] [CrossRef]
- Friebe, B.; Qi, L.L.; Wilson, D.L.; Chang, Z.J.; Seifers, D.L.; Martin, T.J.; Fritz, A.K.; Gill, B.S. Wheat-Thinopyrum intermedium recombinants resistant to wheat streak mosaic virus and Triticum mosaic virus. Crop Sci. 2009, 49, 1221–1226. [Google Scholar] [CrossRef]
- Xin, Z.Y.; Xu, H.J.; Chen, X.; Li, Z.S.; Zhou, G.H.; Qian, Y.X.; Cheng, Z.M.; LARKIN, P.J.; BANKS, P.; APPELS, R.; et al. Development of common wheat germplasm resistant to barley yellow dwarf virus by biotecnology. Sci. China (Ser. B) 1991, 21, 36–42. [Google Scholar]
- Liu, C.; Han, R.; Wang, X.L.; Gong, W.P.; Cheng, D.G.; Cao, X.Y.; Liu, A.F.; Li, H.S.; Liu, J.J. Research progress of wheat wild hybridization, disease resistance genes transfer and utilization. Sci. Agric. Sin. 2020, 53, 1287–1308. [Google Scholar]
- Long, D. Molecular Cytogenetic Analysis of Wheat and Triticum durum-Lophopyrumn elogatum Amphidiploid Alien Chromosome Lines. Master’s Thesis, Sichuan Agricultural University, Yaan, China, 2015. [Google Scholar]
- Luo, X.J.; Li, X.; Qiao, L.Y.; Guo, H.J.; Jia, J.Q.; Zhang, S.W.; Mei, C.; Chang, Z.J.; Zhang, X.J. Research Progress of Wheat Genetic Improvement Derived from Elytrigia. J. Shanxi Agric. Sci. 2019, 47, 482–485, 490. [Google Scholar]
- Luo, X.J.; Qiao, L.Y.; Li, X.; Guo, H.J.; Yan, X.T.; Zhang, S.W.; Chang, L.F.; Yan, J.L.; Chang, Z.J.; Zhang, X.J. Molecular Cytogenetic Identification of Triticum aestivum-Thinopyrum intermedium Substitution Line CH188. J. Shanxi Agric. Sci. 2020, 48, 317–320. [Google Scholar]
- Cui, L.; Ren, Y.K.; Murray, T.D.; Yan, W.Z.; Guo, Q.; Niu, Y.Q.; Sun, Y.; Li, H.J. Development of Perennial Wheat Through Hybridization Between Wheat and Wheatgrasses: A Review. Engineering 2018, 4, 507–513. [Google Scholar] [CrossRef]
- Yu, Z.H.; Wang, H.J.; Xu, Y.F.; Li, Y.S.; Lang, T.; Yang, Z.J.; Li, G.R. Characterization of chromosomal rearrangement in new wheat-Thinopyrum intermedium addition lines carrying Thinopyrum-specific grain hardness genes. Agronomy 2019, 9, 18. [Google Scholar] [CrossRef]
- Yu, Z.H.; Wang, H.J.; Yang, E.N.; Li, G.R.; Yang, Z.J. Precise identification of chromosome constitution and rearrangements in wheat-Thinopyrum intermedium derivatives by ND-FISH and Oligo-FISH painting. Plants 2022, 11, 2109. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J. Development and Validation of a STS Marker Set Specific to Thinopyrum intermedium Genome. Master’s Thesis, Shanxi University, Taiyuan, China, 2020. [Google Scholar]
- Peterson, R.; Campbell, A.; Hannah, A. Diagrammatic scale for estimating rust intensity of leaves and stem of cereals. Can. J. Res. Sect. C 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Zhang, B.; Li, J.X.; Wang, Z.; Shi, L.C.; Li, J.B. Evaluation of resistance to Fusarium head blight and investigation of agronomic traits in 54 wheat cultivars (lines). Plant Prot. 2023, 49, 276–283. [Google Scholar]
- Li, J.B.; Lang, T.; Li, B.; Yu, Z.H.; Wang, H.J.; Li, G.R.; Yang, E.N.; Yang, Z.J. Introduction of Thinopyrum intermedium ssp. trichophorum chromosomes to wheat by trigeneric hybridization involving Triticum, secale and Thinopyrum genera. Planta 2017, 245, 1121–1135. [Google Scholar] [CrossRef]
- Chen, W.Q.; Wu, L.R.; Liu, T.G.; Xu, S.C.; Jin, S.L.; Peng, Y.L.; Wang, B.T. Race dynamics, diversity, and virulence evolution in Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust in China from 2003 to 2007. Plant Dis. 2009, 93, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Wan, A.M.; Zhao, Z.H.; Chen, X.M.; He, Z.H.; Jin, S.L.; Jia, Q.Z.; Yao, G.; Yang, J.X.; Wang, B.T.; Li, G.B.; et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.G.; Liu, S.B.; Qi, Z.J.; Kong, F.J.; Gao, J.R. Application Studies of Elytrigia Intermedium in Hereditary Improvement of Wheat. J. Shandong Agric. Univ. 2000, 31, 333–336. [Google Scholar]
- Yan, X.Q.; Zhu, W.; Meng, Z.L. Occurrence and Comprehensive Control of Wheat Scab. Mod. Agric. Sci. Technol. 2019, 24, 86–88. [Google Scholar]
- Thomas, H.M.; Morgan, W.G.; Humphreys, M.W. Designing grasses with a future-combining the attributes of lolium and festuca. Euphytica 2003, 133, 19–26. [Google Scholar] [CrossRef]
- Raina, S.N.; Rani, V. GISH technology in plant genome research. Methods Cell Sci. 2001, 23, 83–104. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Jia, G.Y.; Tan, W.J.; Tian, R.; Zheng, X.C.; Feng, J.M.; Luo, X.Q.; Si, B.F.; Li, X.; Huang, K.B.; et al. Genome-wide QTL mapping for stripe rust resistance in spring wheat line PI 660122 using the Wheat 15K SNP array. Front. Plant Sci. 2023, 14, 1232897. [Google Scholar] [CrossRef]
- Singh, R.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.Y.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-espino, J. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef]
- An, D.G.; Xu, H.X.; Xu, Y.F. Enhancement of wheat distant hybridization germplasm. Chin. J. Eco-Agric. 2011, 19, 1011–1019. [Google Scholar] [CrossRef]
- Zhao, J.H.; Shang, Y.F.; Wang, S.J.; Yang, C.L.; Lu, X.B.; Sun, H.W. Identification of Mult-resistance of 152 Wheat Varieties (Strains) in Huang Huai Region. J. Triticeae Crops 2007, 6, 1123–1127. [Google Scholar]
- Li, G.R.; Chen, Q.H.; Jiang, W.X.; Zhang, A.H.; Yang, E.N.; Yang, Z.J. Molecular and Cytogenetic Identification of Wheat-Thinopyrum intermedium Double Substitution Line-Derived Progenies for Stripe Rust Resistance. Plants 2022, 12, 28. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef]
- Tang, Z.X.; Yang, Z.J.; Fu, S.L. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Xi, W.; Tang, Z.; Tang, S.; Yang, Z.; Luo, J.; Fu, S. New ND-FISH-positive oligo probes for identifying Thinopyrum chromosomes in wheat backgrounds. Int. J. Mol. Sci. 2019, 20, 2031. [Google Scholar] [CrossRef]
- Cui, Y.; Xing, P.Y.; Qi, X.L.; Bao, Y.G.; Wang, H.G.; Richard, R.; Wang, C.; Li, X.F. Characterization of chromosome constitution in three wheat-Thinopyrum intermedium amphiploids revealed frequent rearrangement of alien and wheat chromosomes. BMC Plant Biol. 2021, 21, 129. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material Name | Infection Types (ITs) | Degree of Severity (DS) | Judged Resistances or Susceptibilities |
|---|---|---|---|
| 92048 | 1 | 5% | HR |
| Longchun 10 | 8 | 90% | HS |
| Gan 630 | 8 | 90% | HS |
| Stripe Rust | FHB | |||||
|---|---|---|---|---|---|---|
| Material Name | Infection Types (ITs) | Degree of Severity (DS) | Judged Resistances or Susceptibilities | Infection Types (ITs) | Degree of Severity (DS) | Judged Resistances or Susceptibilities |
| 92048 | 1 | 5% | HR | 1 | 5.88% | HR |
| Longchun 10 | 8 | 90% | HS | 3 | 56.25% | MS |
| Gan 630 | 8 | 90% | HS | 4 | 100% | HS |
| Probe | Oligonucleotide Sequence | 5′-Modification | References |
|---|---|---|---|
| Oligo-pSc119.2-1 | 5′CCGTTTTGTGGACTATTACTCACCGCTT TGG GGTCCCATAGCTAT 3′ | 5′-Cy5 | [57] |
| Oligo-pTa535-1 | 5′AAAAACTTGACGCACGTCACGTACAAATTGGACAAACTCTTTCGGAGTATCAGGGTTTC 3′ | 5′-Tamra | [57] |
| Oligo-B11 | 5′TCCGCTCACCTTGATGACAACATCAGG TGGAATTCCGTTCGAGGG 3′ | 5′-FAM | [58] |
| Oligo-pDb12H | 5′TCAGAATTTTTAGGATAGCAGAAGTAT TCG AAATACCCAGATTGCTACAG 3′ | 5′-FAM | [39] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; He, Y.; Feng, X.; Huang, M.; Huang, K.; Li, X.; Yang, S.; Ren, Y. Molecular and Cytological Identification of Wheat-Thinopyrum intermedium Partial Amphiploid Line 92048 with Resistance to Stripe Rust and Fusarium Head Blight. Plants 2024, 13, 1198. https://doi.org/10.3390/plants13091198
Luo X, He Y, Feng X, Huang M, Huang K, Li X, Yang S, Ren Y. Molecular and Cytological Identification of Wheat-Thinopyrum intermedium Partial Amphiploid Line 92048 with Resistance to Stripe Rust and Fusarium Head Blight. Plants. 2024; 13(9):1198. https://doi.org/10.3390/plants13091198
Chicago/Turabian StyleLuo, Xiaoqin, Yuanjiang He, Xianli Feng, Min Huang, Kebing Huang, Xin Li, Suizhuang Yang, and Yong Ren. 2024. "Molecular and Cytological Identification of Wheat-Thinopyrum intermedium Partial Amphiploid Line 92048 with Resistance to Stripe Rust and Fusarium Head Blight" Plants 13, no. 9: 1198. https://doi.org/10.3390/plants13091198