Structural Diversity and Function of Xyloglucan Sidechain Substituents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. XyG Structural Nomenclature and Presently Known Sidechain Diversity

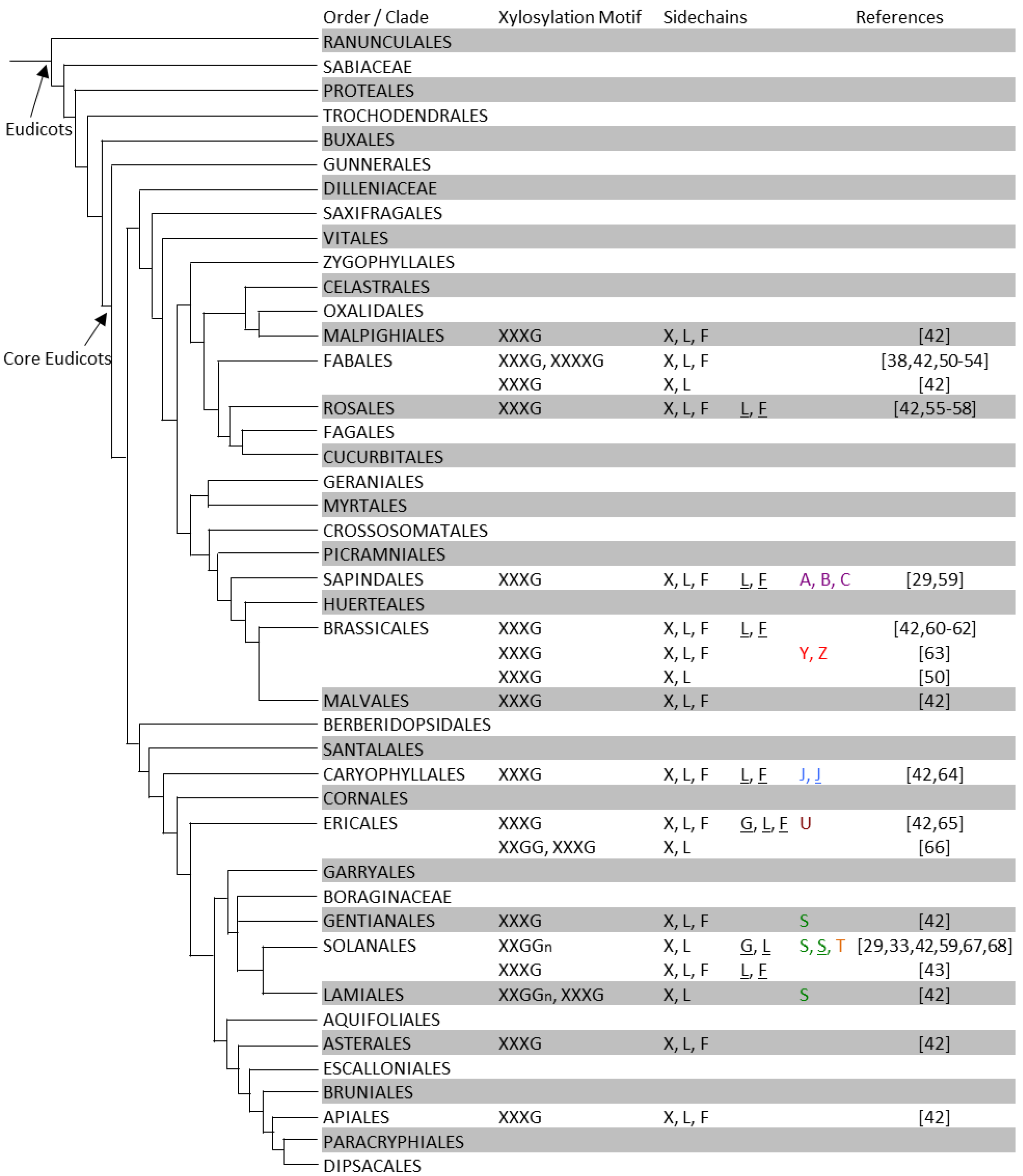
3. XyG Biodiversity


4. XyG Biosynthesis

5. Outlook
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carpita, N.C.; Gibeaut, D.M. Structural models of primary-cell walls in flowering plants—Consistency of molecular-structure with the physical-properties of the walls during growth. Plant J. 1993, 3, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.; Bauer, S.; Brininstool, G.; Facette, M.; Hamann, T.; Milne, J.; Osborne, E.; Paredez, A.; Persson, S.; Raab, T.; et al. Toward a systems approach to understanding plant-cell walls. Science 2004, 306, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Kooiman, P. The constitution of tamarindus-amyloid. Recl. Trav. Chim. Pay. B 1961, 80, 849–865. [Google Scholar] [CrossRef]
- Keegstra, K.; Talmadge, K.W.; Bauer, W.; Albersheim, P. The structure of plant cell walls III. A model of the walls of suspension-cultured sycamore cells based on the interconnections of the macromolecular components. Plant Physiol. 1973, 51, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Albersheim, P.; Darvill, A.; Roberts, K.; Sederoff, R.; Staehelin, A. Plant Cell Walls: From Chemistry to Biology; Garland Science, Taylor and Francis Group, LLC: New York, NY, USA, 2011; pp. 1–430. [Google Scholar]
- Hayashi, T. Xyloglucans in the primary-cell wall. Annu. Rev. Plant Phys. 1989, 40, 139–168. [Google Scholar] [CrossRef]
- Pauly, M.; Albersheim, P.; Darvill, A.; York, W.S. Molecular domains of the cellulose/xyloglucan network in the cell walls of higher plants. Plant J. 1999, 20, 629–639. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.; Roberts, K. Architecture of the primary cell wall. In The Cytoskeletal Basis of Plant Growth and Form; Academic Press: Waltham, MA, USA, 1991; pp. 109–129. [Google Scholar]
- Thompson, D.S. How do cell walls regulate plant growth? J. Exp. Bot. 2005, 56, 2275–2285. [Google Scholar] [CrossRef]
- Anderson, C.T.; Carroll, A.; Akhmetova, L.; Somerville, C. Real-time imaging of cellulose reorientation during cell wall expansion in Arabidopsis roots. Plant Physiol. 2010, 152, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Talbott, L.D.; Ray, P.M. Molecular size and separability features of pea cell wall polysaccharides Implications for models of primary wall structure. Plant Physiol. 1992, 98, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.-A.; Apperley, D.C.; Jarvis, M.C. Molecular rigidity in dry and hydrated onion cell walls. Plant Physiol. 1997, 115, 593–598. [Google Scholar] [PubMed]
- Takeda, T.; Furuta, Y.; Awano, T.; Mizuno, K.; Mitsuishi, Y.; Hayashi, T. Suppression and acceleration of cell elongation by integration of xyloglucans in pea stem segments. Proc. Natl. Acad. Sci. USA 2002, 99, 9055–9060. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, D.M.; Lerouxel, O.; Neumetzler, L.; Yamauchi, K.; Reinecke, A.; Freshour, G.; Zabotina, O.A.; Hahn, M.G.; Burgert, I.; Pauly, M.; et al. Disrupting two Arabidopsis thaliana xylosyltransferase genes results in plants deficient in xyloglucan, a major primary cell wall component. Plant Cell 2008, 20, 1519–1537. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.B.; Cosgrove, D.J. A revised architecture of primary cell walls based on biomechanical changes induced by substrate-specific endoglucanases. Plant Physiol. 2012, 158, 1933–1943. [Google Scholar] [CrossRef] [PubMed]
- Meier, H.; Reid, J. Reserve polysaccharides other than starch in higher plants. In Plant Carbohydrates I; Springer: Heidelberg, Germany, 1982; pp. 418–471. [Google Scholar]
- Rao, P.; Srivastava, H. Tamarind. In Industrial Gums, 2nd ed.; Academic Press: New York, NY, USA, 1973; pp. 369–411. [Google Scholar]
- Edwards, M.; Bowman, Y.; Dea, I.; Reid, J. A β-d-Galactosidase from nasturtium (Tropaeolum majus L.) cotyledons. Purification, properties, and demonstration that xyloglucan is the natural substrate. J. Biol. Chem. 1988, 263, 4333–4337. [Google Scholar] [PubMed]
- Mishra, A.; Malhotra, A.V. Tamarind xyloglucan: A polysaccharide with versatile application potential. J. Mater. Chem. 2009, 19, 8528–8536. [Google Scholar] [CrossRef]
- Mishra, M.; Khandare, J. Evaluation of tamarind seed polysaccharide as a biodegradable carrier for colon specific drug delivery. Int. J. Pharm. Pharm. Sci. 2011, 3, 139–142. [Google Scholar]
- Rolando, M.; Valente, C. Establishing the tolerability and performance of tamarind seed polysaccharide (TSP) in treating dry eye syndrome: Results of a clinical study. BMC Ophthalmol. 2007, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Ghelardi, E.; Tavanti, A.; Celandroni, F.; Lupetti, A.; Blandizzi, C.; Boldrini, E.; Campa, M.; Senesi, S. Effect of a novel mucoadhesive polysaccharide obtained from tamarind seeds on the intraocular penetration of gentamicin and ofloxacin in rabbits. J. Antimicrob. Chemother. 2000, 46, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Greffe, L.; Bessueille, L.; Bulone, V.; Brumer, H. Synthesis, preliminary characterization, and application of novel surfactants from highly branched xyloglucan oligosaccharides. Glycobiology 2005, 15, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Brumer, H., 3rd; Zhou, Q.; Baumann, M.J.; Carlsson, K.; Teeri, T.T. Activation of crystalline cellulose surfaces through the chemoenzymatic modification of xyloglucan. J. Am. Chem. Soc. 2004, 126, 5715–5721. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Gille, S.; Liu, L.; Mansoori, N.; de Souza, A.; Schultink, A.; Xiong, G. Hemicellulose biosynthesis. Planta 2013, 238, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Fry, S.C.; York, W.S.; Albersheim, P.; Darvill, A.; Hayashi, T.; Joseleau, J.P.; Kato, Y.; Lorences, E.P.; Maclachlan, G.A.; Mcneil, M.; et al. An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol. Plantarum 1993, 89, 1–3. [Google Scholar] [CrossRef]
- Tuomivaara, S.T.; Yaoi, K.; O’Neill, M.A.; York, W.S. Generation and structural validation of a library of diverse xyloglucan-derived oligosaccharides, including an update on xyloglucan nomenclature. Carbohydr. Res. 2014. [Google Scholar] [CrossRef]
- Kiefer, L.L.; York, W.S.; Darvill, A.G.; Albersheim, P. Xyloglucan isolated from suspension-cultured sycamore cell-walls is O-acetylated. Phytochemistry 1989, 28, 2105–2107. [Google Scholar] [CrossRef]
- York, W.S.; Oates, J.E.; van Halbeek, H.; Darvill, A.G.; Albersheim, P.; Tiller, P.R.; Dell, A. Location of the O-acetyl substituents on a nonasaccharide repeating unit of sycamore extracellular xyloglucan. Carbohydr. Res. 1988, 173, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Sims, I.M.; Munro, S.L.; Currie, G.; Craik, D.; Bacic, A. Structural characterisation of xyloglucan secreted by suspension-cultured cells of Nicotiana plumbaginifolia. Carbohydr. Res. 1996, 293, 147–172. [Google Scholar] [CrossRef] [PubMed]
- York, W.S.; Kolli, V.S.K.; Orlando, R.; Albersheim, P.; Darvill, A.G. The structures of arabinoxyloglucans produced by solanaceous plants. Carbohydr. Res. 1996, 285, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Cash, M.; Darvill, A.G.; York, W.S. NMR characterization of endogenously O-acetylated oligosaccharides isolated from tomato (Lycopersicon esculentum) xyloglucan. Carbohydr. Res. 2005, 340, 1818–1825. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Goto, C.; Numata, M.; Suzuki, T.; Nakagawa, Y.; Hoshino, T.; Uchiyama, T. O-acetylated xyloglucan in extracellular polysaccharides from cell-suspension cultures of Mentha. Phytochemistry 1996, 41, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Kamerling, J.P.; Schauer, R.; Shukla, A.K.; Stoll, S.; Halbeek, H.; Vliegenthart, J. Migration of O‐acetyl groups in N,O‐acetylneuraminic acids. Eur. J. Biochem. 1987, 162, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Kabel, M.A.; de Waard, P.; Schols, H.A.; Voragen, A.G. Location of O-acetyl substituents in xylo-oligosaccharides obtained from hydrothermally treated Eucalyptus wood. Carbohydr. Res. 2003, 338, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M. Development of Analytical Tools to Study Plant Cell Wall Xyloglucan; Shaker Verlag GmbH: Herzogenrath, Germany, 1999; pp. 1–252. [Google Scholar]
- Pauly, M.; Qin, Q.; Greene, H.; Albersheim, P.; Darvill, A.; York, W.S. Changes in the structure of xyloglucan during cell elongation. Planta 2001, 212, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Darvill, A.G.; Eberhard, S.; York, W.S.; O’Neill, M.A. Moss and liverwort xyloglucans contain galacturonic acid and are structurally distinct from the xyloglucans synthesized by hornworts and vascular plants. Glycobiology 2008, 18, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Popper, Z.A.; Fry, S.C. Primary cell wall composition of bryophytes and charophytes. Ann. Bot. 2003, 91, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.D.; Harholt, J.; Ulvskov, P.; Johansen, I.E.; Fangel, J.U.; Doblin, M.S.; Bacic, A.; Willats, W.G. Evidence for land plant cell wall biosynthetic mechanisms in charophyte green algae. Ann. Bot. 2014, 114, 1217–1236. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.; Jia, Z.; Pena, M.J.; Cash, M.; Harper, A.; Blackburn, A.R., 2nd; Darvill, A.; York, W.S. Structural analysis of xyloglucans in the primary cell walls of plants in the subclass Asteridae. Carbohydr. Res. 2005, 340, 1826–1840. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, E.R.; Moller, I.E.; Cassin, A.; Jones, D.F.; Koh, P.L.; Ratnayake, S.; Beahan, C.T.; Wilson, S.M.; Bacic, A.; Newbigin, E. In vitro grown pollen tubes of Nicotiana alata actively synthesise a fucosylated xyloglucan. PLoS One 2013, 8, e77140. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.S.; Harris, P.J. Structures of xyloglucans in primary cell walls of gymnosperms, monilophytes (ferns sensu lato) and lycophytes. Phytochemistry 2012, 79, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Bremer, B.; Bremer, K.; Chase, M.; Fay, M.; Reveal, J.; Soltis, D.; Soltis, P.; Stevens, P. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Kakegawa, K.; Edashige, Y.; Ishii, T. Xyloglucan from xylem-differentiating zones of Cryptomeria japonica. Phytochemistry 1998, 47, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.S.; Harris, P.J. Xyloglucans of monocotyledons have diverse structures. Mol. Plant 2009, 2, 943–965. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.A.; York, W.S. The composition and structure of plant primary cell walls. In The Plant Cell Wall; Blackwell Publishing: Oxford, UK, 2003; pp. 1–54. [Google Scholar]
- Gibeaut, D.M.; Pauly, M.; Bacic, A.; Fincher, G.B. Changes in cell wall polysaccharides in developing barley (Hordeum vulgare) coleoptiles. Planta 2005, 221, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Buckeridge, M.S. Seed cell wall storage polysaccharides: Models to understand cell wall biosynthesis and degradation. Plant Physiol. 2010, 154, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Vinueza, N.R.; Gallardo, V.A.; Klimek, J.F.; Carpita, N.C.; Kenttämaa, H.I. Analysis of xyloglucans by ambient chloride attachment ionization tandem mass spectrometry. Carbohydr. Polym. 2013, 98, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Huisman, M.; Weel, K.; Schols, H.; Voragen, A. Xyloglucan from soybean (Glycine max) meal is composed of XXXG-type building units. Carbohydr. Polym. 2000, 42, 185–191. [Google Scholar] [CrossRef]
- Alonso-Simón, A.; Neumetzler, L.; García-Angulo, P.; Encina, A.; Acebes, J.; Álvarez, J.; Hayashi, T. Plasticity of xyloglucan composition in bean (Phaseolus vulgaris)-cultured cells during habituation and dehabituation to lethal concentrations of dichlobenil. Mol. Plant 2010, 3, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Picout, D.R.; Ellis, P.R.; Ross-Murphy, S.B.; Reid, J. A novel xyloglucan from seeds of Afzelia africana Se. Pers.—Extraction, characterization, and conformational properties. Carbohydr. Res. 2005, 340, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Watt, D.; Brasch, D.; Larsen, D.; Melton, L. Isolation, characterisation, and NMR study of xyloglucan from enzymatically depectinised and non-depectinised apple pomace. Carbohydr. Polym. 1999, 39, 165–180. [Google Scholar] [CrossRef]
- Galvez-Lopez, D.; Laurens, F.; Quemener, B.; Lahaye, M. Variability of cell wall polysaccharides composition and hemicellulose enzymatic profile in an apple progeny. Int. J. Biol. Macromol. 2011, 49, 1104–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahaye, M.; Falourd, X.; Quemener, B.; Ralet, M.C.; Howad, W.; Dirlewanger, E.; Arús, P. Cell wall polysaccharide chemistry of peach genotypes with contrasted textures and other fruit traits. J. Agric. Food Chem. 2012, 60, 6594–6605. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Vigouroux, J.; Quémener, B.; Bonnin, E.; Lahaye, M. Novel and diverse fine structures in LiCl-DMSO extracted apple hemicelluloses. Carbohydr. Polym. 2014, 108, 46–57. [Google Scholar] [CrossRef] [PubMed]
- York, W.S.; Impallomeni, G.; Hisamatsu, M.; Albersheim, P.; Darvill, A.G. Eleven newly characterized xyloglucan oligoglycosyl alditols: The specific effects of sidechain structure and location on 1H-NMR chemical shifts. Carbohydr. Res. 1995, 267, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Louvet, R.; Rayon, C.; Domon, J.-M.; Rusterucci, C.; Fournet, F.; Leaustic, A.; Crépeau, M.-J.; Ralet, M.-C.; Rihouey, C.; Bardor, M. Major changes in the cell wall during silique development in Arabidopsis thaliana. Phytochemistry 2011, 72, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lerouxel, O.; Choo, T.S.; Seveno, M.; Usadel, B.; Faye, L.; Lerouge, P.; Pauly, M. Rapid structural phenotyping of plant cell wall mutants by enzymatic oligosaccharide fingerprinting. Plant Physiol. 2002, 130, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Crombie, H.J.; Chengappa, S.; Hellyer, A.; Reid, J.S. A xyloglucan oligosaccharide-active, transglycosylating β-d-glucosidase from the cotyledons of nasturtium (Tropaeolum majus L.) seedlings—Purification, properties and characterization of a cDNA clone. Plant J. 1998, 15, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Kong, Y.; York, W.S.; O’Neill, M.A. A galacturonic acid-containing xyloglucan is involved in Arabidopsis root hair tip growth. Plant Cell 2012, 24, 4511–4524. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Joshi, M.; Jha, B. Oligosaccharide mass profiling of nutritionally important Salicornia brachiata, an extreme halophyte. Carbohydr. Polym. 2013, 92, 1942–1945. [Google Scholar] [CrossRef] [PubMed]
- Hilz, H.; de Jong, L.E.; Kabel, M.A.; Verhoef, R.; Schols, H.A.; Voragen, A.G. Bilberry xyloglucan—Novel building blocks containing β-xylose within a complex structure. Carbohydr. Res. 2007, 342, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Aboughe-Angone, S.; Nguema-Ona, E.; Ghosh, P.; Lerouge, P.; Ishii, T.; Ray, B.; Driouich, A. Cell wall carbohydrates from fruit pulp of Argania spinosa: Structural analysis of pectin and xyloglucan polysaccharides. Carbohydr. Res. 2008, 343, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Assor, C.; Quemener, B.; Vigouroux, J.; Lahaye, M. Fractionation and structural characterization of LiCl-DMSO soluble hemicelluloses from tomato. Carbohydr. Polym. 2013, 94, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Qin, Q.; Darvill, A.G.; York, W.S. Structure of the xyloglucan produced by suspension-cultured tomato cells. Carbohydr. Res. 2003, 338, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Schultink, A.; Cheng, K.; Park, Y.B.; Cosgrove, D.J.; Pauly, M. The identification of two arabinosyltransferases from tomato reveals functional equivalency of xyloglucan side chain substituents. Plant Physiol. 2013, 163, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Hantus, S.; Pauly, M.; Darvill, A.G.; Albersheim, P.; York, W.S. Structural characterization of novel L-galactose-containing oligosaccharide subunits of jojoba seed xyloglucans. Carbohydr. Res. 1997, 304, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Selvendran, R.R.; Ryden, P. Isolation and analysis of plant cell walls. In Methods in Plant Biochemistry; Dey, P.M., Ed.; Academic Press: New York, NY, USA, 1990; pp. 549–579. [Google Scholar]
- Pauly, M.; Andersen, L.N.; Kauppinen, S.; Kofod, L.V.; York, W.S.; Albersheim, P.; Darvill, A. A xyloglucan-specific endo-β-1,4-glucanase from Aspergillus aculeatus: Expression cloning in yeast, purification and characterization of the recombinant enzyme. Glycobiology 1999, 9, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Obel, N.; Erben, V.; Schwarz, T.; Kühnel, S.; Fodor, A.; Pauly, M. Microanalysis of plant cell wall polysaccharides. Mol. Plant 2009, 2, 922–932. [Google Scholar] [PubMed]
- Watanabe, T.; Shida, M.; Murayama, T.; Furuyama, Y.; Nakajima, T.; Matsuda, K.; Kainuma, K. Xyloglucan in cell walls of rice hull. Carbohydr. Res. 1984, 129, 229–242. [Google Scholar] [CrossRef]
- Günl, M.; Neumetzler, L.; Kraemer, F.; de Souza, A.; Schultink, A.; Pena, M.; York, W.S.; Pauly, M. AXY8 encodes an α-fucosidase, underscoring the importance of apoplastic metabolism on the fine structure of Arabidopsis cell wall polysaccharides. Plant Cell 2011, 23, 4025–4040. [Google Scholar] [CrossRef] [PubMed]
- Marcus, S.E.; Verhertbruggen, Y.; Herve, C.; Ordaz-Ortiz, J.J.; Farkas, V.; Pedersen, H.L.; Willats, W.G.T.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, e60. [Google Scholar] [CrossRef]
- Brennan, M.; Harris, P.J. Distribution of fucosylated xyloglucans among the walls of different cell types in monocotyledons determined by immunofluorescence microscopy. Mol. Plant 2011, 4, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Lerouxel, O.; Cavalier, D.M.; Liepman, A.H.; Keegstra, K. Biosynthesis of plant cell wall polysaccharides—A complex process. Curr. Opin. Plant Biol. 2006, 9, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.-R.; Pauly, M. Glycosyltransferases and cell wall biosynthesis: Novel players and insights. Curr. Opin. Plant Biol. 2004, 7, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Cocuron, J.C.; Lerouxel, O.; Drakakaki, G.; Alonso, A.P.; Liepman, A.H.; Keegstra, K.; Raikhel, N.; Wilkerson, C.G. A gene from the cellulose synthase-like C family encodes a β-1,4 glucan synthase. Proc. Natl. Acad. Sci. USA 2007, 104, 8550–8555. [Google Scholar] [CrossRef] [PubMed]
- Faik, A.; Price, N.J.; Raikhel, N.V.; Keegstra, K. An Arabidopsis gene encoding an α-xylosyltransferase involved in xyloglucan biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 7797–7802. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, D.M.; Keegstra, K. Two xyloglucan xylosyltransferases catalyze the addition of multiple xylosyl residues to cellohexaose. J. Biol. Chem. 2006, 281, 34197–34207. [Google Scholar] [CrossRef] [PubMed]
- Ade, C.P.; Bemm, F.; Dickson, J.M.; Walter, C.; Harris, P.J. Family 34 glycosyltransferase (GT34) genes and proteins in Pinus radiata (radiata pine) and Pinus taeda (loblolly pine). Plant J. 2014, 78, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Zabotina, O.A.; van de Ven, W.T.G.; Freshour, G.; Drakakaki, G.; Cavalier, D.; Mouille, G.; Hahn, M.G.; Keegstra, K.; Raikhel, N.V. Arabidopsis XXT5 gene encodes a putative α-1,6-xylosyltransferase that is involved in xyloglucan biosynthesis. Plant J. 2008, 56, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Vuttipongchaikij, S.; Brocklehurst, D.; Steele-King, C.; Ashford, D.A.; Gomez, L.D.; McQueen-Mason, S.J. Arabidopsis GT34 family contains five xyloglucan α-1,6-xylosyltransferases. New Phytol. 2012, 195, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Zabotina, O.A.; Avci, U.; Cavalier, D.; Pattathil, S.; Chou, Y.H.; Eberhard, S.; Danhof, L.; Keegstra, K.; Hahn, M.G. Mutations in multiple XXT genes of Arabidopsis reveal the complexity of xyloglucan biosynthesis. Plant Physiol. 2012, 159, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, S.; Ng, S.; Zhang, B.; Zhou, Y.; Whelan, J.; Wu, P.; Shou, H. Mutation in xyloglucan 6-xylosytransferase results in abnormal root hair development in Oryza sativa. J. Exp. Bot. 2014. [Google Scholar] [PubMed]
- Madson, M.; Dunand, C.; Li, X.; Verma, R.; Vanzin, G.F.; Caplan, J.; Shoue, D.A.; Carpita, N.C.; Reiter, W.D. The MUR3 gene of Arabidopsis encodes a xyloglucan galactosyltransferase that is evolutionarily related to animal exostosins. Plant Cell 2003, 15, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.K.; Schultink, A.; Keegstra, K.; Wilkerson, C.G.; Pauly, M. RNA-Seq Analysis of developing nasturtium seeds (Tropaeolum majus): Identification and characterization of an additional galactosyltransferase involved in xyloglucan biosynthesis. Mol. Plant 2012, 5, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Lopes, F.J.F.; Pauly, M.; Brommonshenkel, S.H.; Lau, E.Y.; Diola, V.; Passos, J.L.; Loureiro, M.E. The EgMUR3 xyloglucan galactosyltransferase from Eucalyptus grandis complements the mur3 cell wall phenotype in Arabidopsis thaliana. Tree Genet. Genomes 2010, 6, 745–756. [Google Scholar] [CrossRef]
- Perrin, R.M.; DeRocher, A.E.; Bar-Peled, M.; Zeng, W.Q.; Norambuena, L.; Orellana, A.; Raikhel, N.V.; Keegstra, K. Xyloglucan fucosyltransferase, an enzyme involved in plant cell wall biosynthesis. Science 1999, 284, 1976–1979. [Google Scholar] [CrossRef] [PubMed]
- Vanzin, G.F.; Madson, M.; Carpita, N.C.; Raikhel, N.V.; Keegstra, K.; Reiter, W.D. The mur2 mutant of Arabidopsis thaliana lacks fucosylated xyloglucan because of a lesion in fucosyltransferase AtFUT1. Proc. Natl. Acad. Sci. USA 2002, 99, 3340–3345. [Google Scholar] [CrossRef] [PubMed]
- Gille, S.; de Souza, A.; Xiong, G.; Benz, M.; Cheng, K.; Schultink, A.; Reca, I.B.; Pauly, M. O-acetylation of Arabidopsis hemicellulose xyloglucan requires AXY4 or AXY4L, proteins with a TBL and DUF231 domain. Plant Cell 2011, 23, 4041–4053. [Google Scholar] [CrossRef] [PubMed]
- Günl, M.; Pauly, M. AXY3 encodes a α-xylosidase that impacts the structure and accessibility of the hemicellulose xyloglucan in Arabidopsis plant cell walls. Planta 2011, 233, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Pardo, B.; Gianzo, C.; Guitian, E.; Revilla, G.; Zarra, I. Lack of α-xylosidase activity in Arabidopsis alters xyloglucan composition and results in growth defects. Plant Physiol. 2010, 154, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Gianzo, C.; Iglesias, N.; Guitian, E.; Revilla, G.; Zarra, I. AtBGAL10 is the main xyloglucan beta-galactosidase in Arabidopsis, and its absence results in unusual xyloglucan subunits and growth defects. Plant Physiol. 2012, 158, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
- Zablackis, E.; York, W.S.; Pauly, M.; Hantus, S.; Reiter, W.D.; Chapple, C.C.; Albersheim, P.; Darvill, A. Substitution of l-fucose by l-galactose in cell walls of Arabidopsis mur1. Science 1996, 272, 1808–1810. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Eberhard, S.; Albersheim, P.; Darvill, A.; York, W.S. Effects of the mur1 mutation on xyloglucans produced by suspension-cultured Arabidopsis thaliana cells. Planta 2001, 214, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Hisamatsu, M.; Impallomeni, G.; York, W.S.; Albersheim, P.; Darvill, A.G. A new undecasaccharide subunit of xyloglucans with two α-L-fucosyl residues. Carbohydr. Res. 1991, 211, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, K. Construction and restructuring of the cellulose-xyloglucan framework in the apoplast as mediated by the xyloglucan-related protein family—A hypothetical scheme. J. Plant Res. 1998, 111, 159–166. [Google Scholar] [CrossRef]
- Tabuchi, A.; Mori, H.; Kamisaka, S.; Hoson, T. A new type of endo-xyloglucan transferase devoted to xyloglucan hydrolysis in the cell wall of azuki bean epicotyls. Plant Cell Physiol. 2001, 42, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Nishitani, K.; Tominaga, R. Endoxyloglucan transferase, a novel class of glycosyltransferase that catalyzes transfer of a segment of xyloglucan molecule to another xyloglucan molecule. J. Biol. Chem. 1992, 267, 21058–21064. [Google Scholar] [PubMed]
- Fry, S.C.; Smith, R.C.; Renwick, K.F.; Martin, D.J.; Hodge, S.K.; Matthews, K.J. Xyloglucan endotransglycosylase, a new wall-loosening enzyme-activity from plants. Biochem. J. 1992, 282, 821–828. [Google Scholar] [PubMed]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH family of enzymes involved in xyloglucan endotransglucosylation and endohydrolysis: Current perspectives and a new unifying nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed]
- Eklof, J.M.; Brumer, H. The XTH gene family: An update on enzyme structure, function, and phylogeny in xyloglucan remodeling. Plant Physiol. 2010, 153, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Martinez-Vilchez, I.M.; Verbelen, J.-P.; Miller, J.G.; Fry, S.C. In vivo colocalization of xyloglucan endotransglycosylase activity and its donor substrate in the elongation zone of Arabidopsis roots. Plant Cell 2000, 12, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, N.; Abelenda, J.A.; Rodiño, M.; Sampedro, J.; Revilla, G.; Zarra, I. Apoplastic glycosidases active against xyloglucan oligosaccharides of Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Dick-Pérez, M.; Zhang, Y.; Hayes, J.; Salazar, A.; Zabotina, O.A.; Hong, M. Structure and interactions of plant cell-wall polysaccharides by two-and three-dimensional magic-angle-spinning solid-state NMR. Biochemistry 2011, 50, 989–1000. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schultink, A.; Liu, L.; Zhu, L.; Pauly, M. Structural Diversity and Function of Xyloglucan Sidechain Substituents. Plants 2014, 3, 526-542. https://doi.org/10.3390/plants3040526
Schultink A, Liu L, Zhu L, Pauly M. Structural Diversity and Function of Xyloglucan Sidechain Substituents. Plants. 2014; 3(4):526-542. https://doi.org/10.3390/plants3040526
Chicago/Turabian StyleSchultink, Alex, Lifeng Liu, Lei Zhu, and Markus Pauly. 2014. "Structural Diversity and Function of Xyloglucan Sidechain Substituents" Plants 3, no. 4: 526-542. https://doi.org/10.3390/plants3040526
APA StyleSchultink, A., Liu, L., Zhu, L., & Pauly, M. (2014). Structural Diversity and Function of Xyloglucan Sidechain Substituents. Plants, 3(4), 526-542. https://doi.org/10.3390/plants3040526