Penium margaritaceum: A Unicellular Model Organism for Studying Plant Cell Wall Architecture and Dynamics

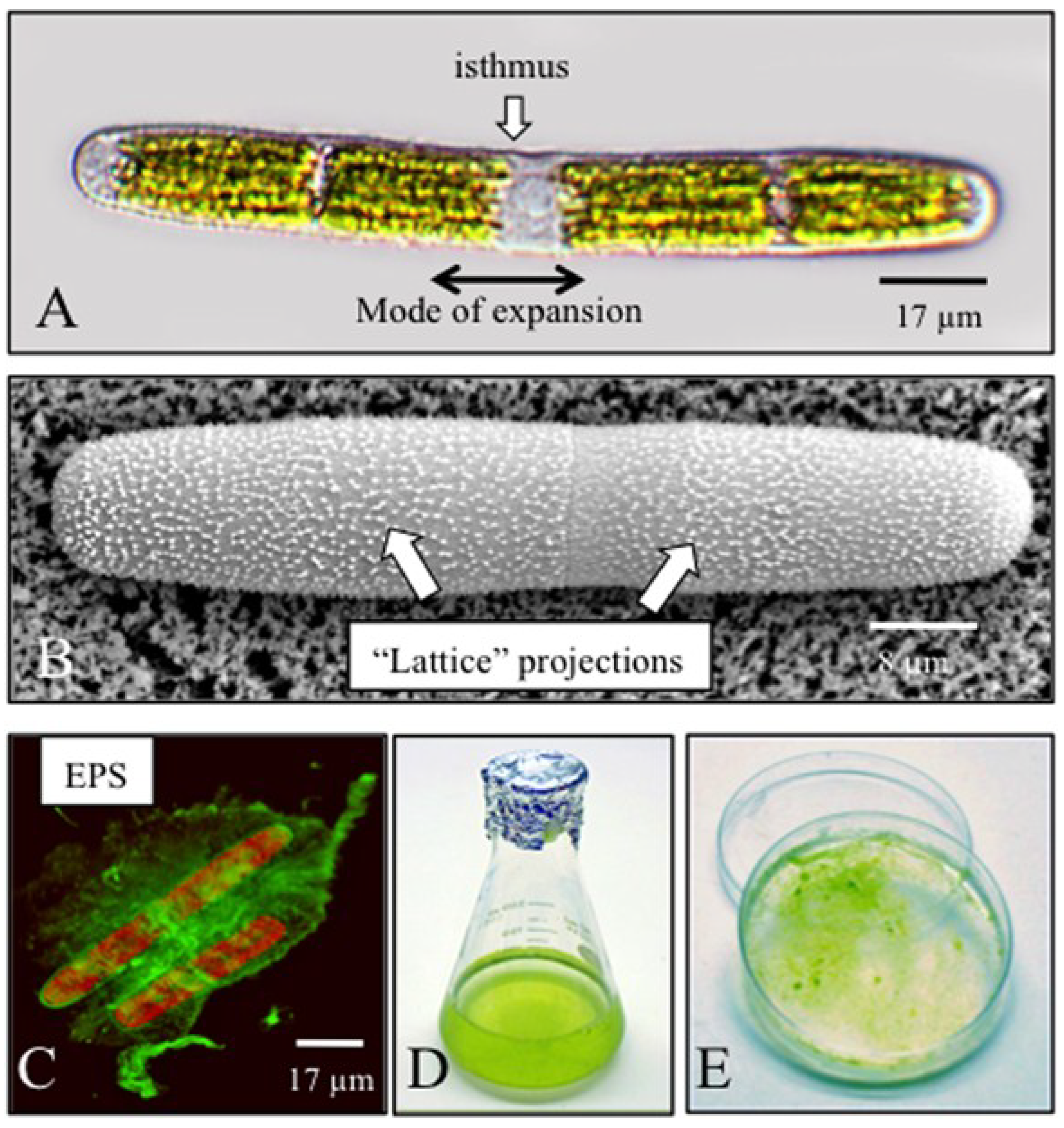{kind=link}
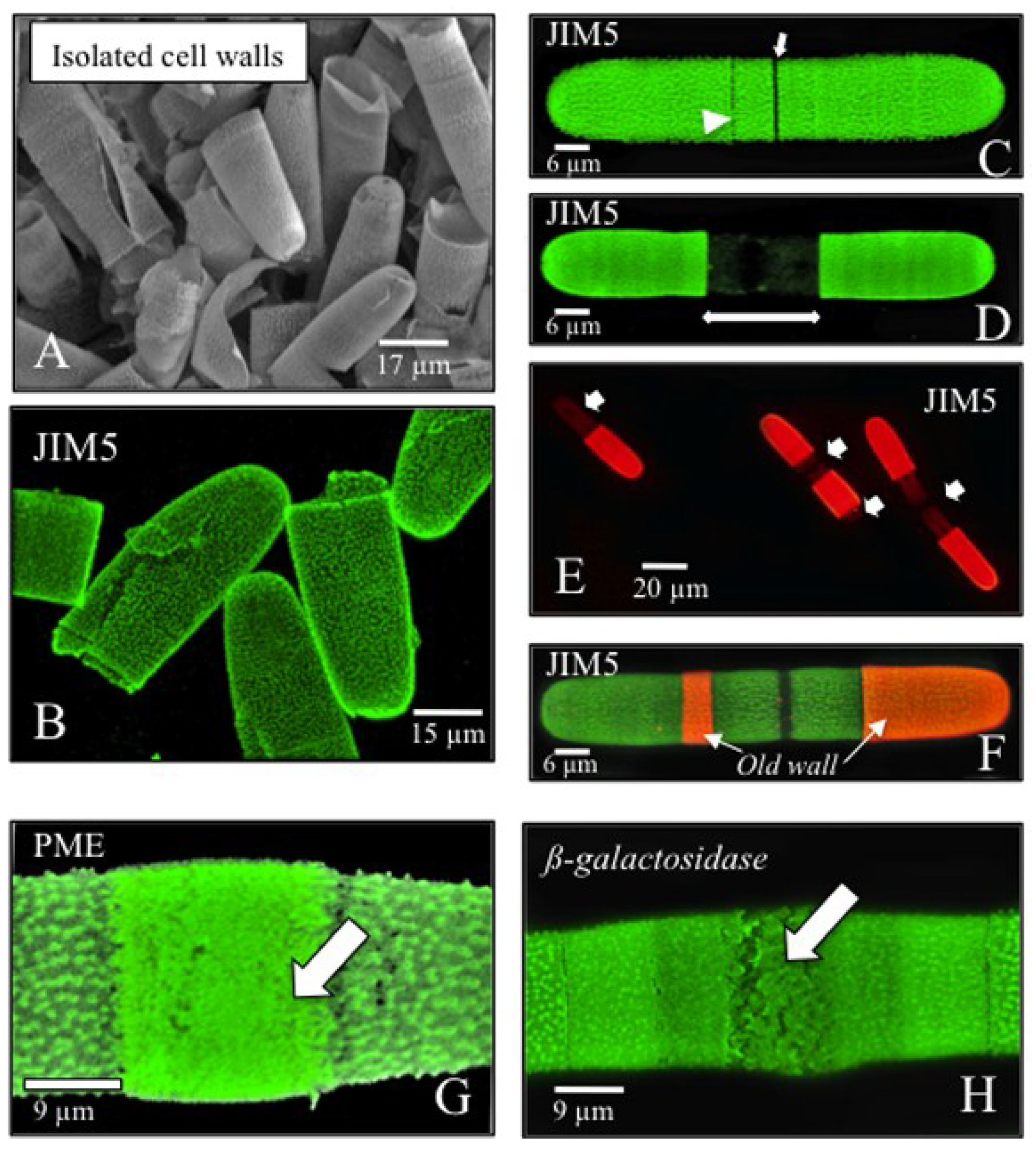{kind=link}
{kind=link}
{kind=link}
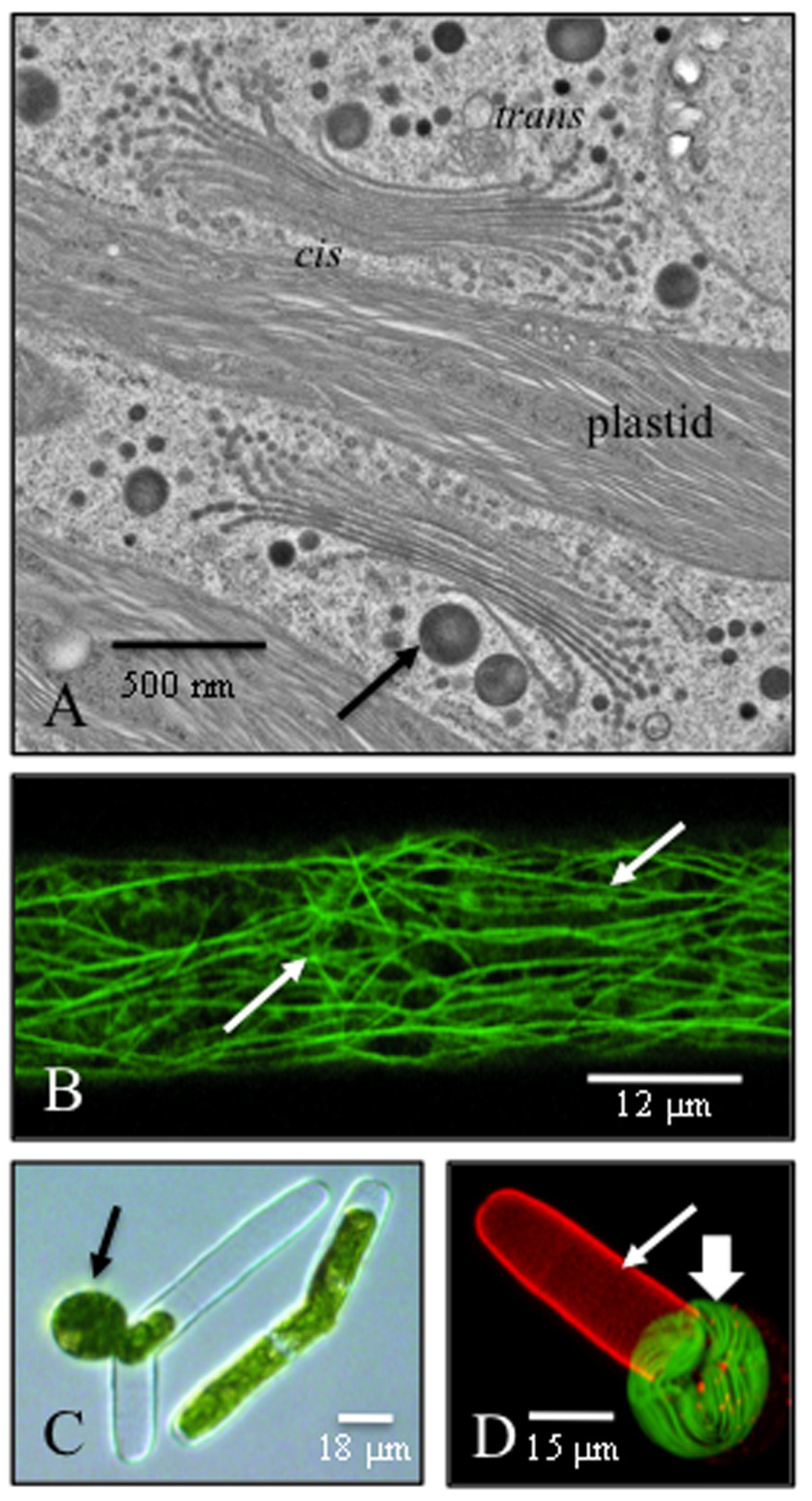{kind=link}
Abstract
:1. Introduction
2. General Biology and Growth Characteristics

3. The Cell Wall: Biochemistry, Labeling and Growth Monitoring
4. High Resolution Electron Microscopy Analyses



5. Experimental Analyses: Microarray or Single-Cell Approach
6. Intracellular Studies
7. Selective Dissolution of the Cell Wall and Protoplast Formation

8. Molecular Biology: Transformation Technology
9. The Future
Acknowledgements
Conflicts of Interest
References
- Harris, D.; Bulone, V.; Ding, S.-Y.; DeBolt, S. Tools for cellulose analysis in plant cell walls. Plant Physiol. 2010, 153, 420–426. [Google Scholar] [PubMed]
- Popper, Z. The plant cell wall. Methods and protocols. Ann. Bot. 2012, 109, vii–viii. [Google Scholar] [PubMed]
- Gilbert, H.J.; Knox, J.P.; Boraston, A.B. Advances in understanding the molecular basis of plant cell wall polysaccharide recognition by carbohydrate-binding modules. Curr. Opin. Struct. Biol. 2013, 23, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.L.; Fangel, J.U.; McCleary, B.; Ruzanski, C.; Rydahl, M.G.; Ralet, M.-C.; Farkas, V.; von Schantz, L.; Marcus, S.E.; Andersen, M.C.F.; et al. Versatile high-resolution oligosaccharide microarrays for plant glycobiology and cell wall research. J. Biol. Chem. 2012, 287, 39429–39438. [Google Scholar] [CrossRef] [PubMed]
- Pattathil, S.; Avci, U.; Baldwin, D.; Swennes, A.G.; McGill, J.A.; Popper, Z.; Bootten, T.; Albert, A.; Davis, R.H.; Chennareddy, C.; et al. A comprehensive toolkit of plant cell wall glycan-directed monoclonal antibodies. Plant Physiol. 2010, 153, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.J.; Marcus, S.E.; Knox, J.P. Cell wall biology: Perspectives from cell wall imaging. Mol. Plant 2011, 4, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, e204. [Google Scholar] [CrossRef] [PubMed]
- Doblin, M.S.; Pettolino, F.; Bacic, A. Plant cell walls: The skeleton of the plant world. Funct. Plant. Biol. 2010, 37, 357–381. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Keegstra, K. Plant cell walls. Plant Physiol. 2010, 154, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Popper, Z.A.; Gurvan, M.; Hervé, C.; Domozych, D.S.; Willats, W.G.T.; Tuohy, M.; Kloareg, B.; Stengel, D.B. Evolution and diversity of plant cell walls: From algae to flowering plants. Ann. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef]
- Dhugga, K.S. Biosynthesis of non-cellulosic polysaccharides of plant cell walls. Phytochemistry 2012, 74, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, C.; Persson, S. Co-expression of cell-wall related genes: New tools and insights. Front. Plant. Sci. 2012, 3, e83. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Z.; Dixon, R.A. On-off switches for secondary cell wall biosynthesis. Mol. Plant 2012, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Liepman, A.H.; Wightman, R.; Geshi, N.; Turner, S.R.; Scheller, H.V. Arabidopsis—A powerful model system for plant cell wall research. Plant J. 2010, 61, 1107–1121. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.T.; Wallace, I.S.; Somerville, C.R. Metabolic click-labeling with a fucose analog reveals pectin delivery, architecture, and dynamics in Arabidopsis cell walls. Proc. Natl. Acad. Sci. USA 2012, 109, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Eberhard, S.; Pattathil, S.; Warder, C.; Glushka, J.; Yuan, C.; Hao, Z.; Zhu, X.; Avci, U.; Miller, J.S.; et al. An Arabidopsis cell wall proteoglycan consists of pectin and arabinoxylan covalently linked to an arabinogalactan protein. Plant Cell 2013, 25, 270–287. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, R.; Daisuke, T.; Fujino, T.; Itoh, T.; Nishitani, K. Cell wall dynamics in tobacco BY-2 cells. In Biotechnology in Agriculture and Technology, 2004 ed.; Nagata, T., Hasezawa, S., Inze, D., Eds.; Springer-Verlag: Berlin, Germany, 2004; Volume 53, pp. 217–230. [Google Scholar]
- Sørensen, I.; Pettolino, F.A.; Bacic, A.; Ralph, J.; Lu, F.; O’Neill, M.A.; Fei, Z.; Rose, J.K.C.; Domozych, D.S.; Willats, W.G.T. The charophycean green algae provide insights into the early origins of plant cell walls. Plant J. 2011, 68, 201–211. [Google Scholar] [CrossRef]
- Sørensen, I.; Domozych, D.S.; Willats, W.G.T. How have plant cell walls evolved? Plant Physiol. 2010, 153, 366–372. [Google Scholar] [CrossRef]
- Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G.T. The cell walls of green algae: A journey through evolution and diversity. Front. Plant Sci. 2012, 3, e82. [Google Scholar] [CrossRef] [PubMed]
- Proseus, T.E.; Boyer, J.S. Calcium pectate chemistry causes growth to be stored in Chara corallina: A test of the pectate cycle. Plant Cell Environ. 2008, 31, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Lütz-Meindl, U. Pectin-like carbohydrates in the green alga Micrasterias characterized by cytochemical analysis and energy filtering TEM. J. Microsc. 2008, 231, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Lütz-Meindl, U.; Brosch-Salomon, S. Cell wall secretion in the green alga Micrasterias. J. Microsc. 2000, 198, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S.; Serfis, A.; Kiemle, S.N.; Gretz, M.R. The structure and biochemistry of charophycean cell walls: I. Pectins of Penium margaritaceum. Protoplasma 2007, 230, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S.; Sørensen, I.; Popper, Z.A.; Ochs, J.; Andreas, A.; Fangel, J.U.; Pielach, A.; Sacks, C.; Brechka, H.; Willats, W.G.T.; et al. Pectin metabolism and assembly in the cell gall of the Charophyte Green alga Penium margaritaceum. Plant Physiol. 2014, 165, 105–118. [Google Scholar] [CrossRef]
- Sørensen, I.; Fei, Z.; Andreas, A.; Willats, W.G.T.; Domozych, D.S.; Rose, J.K.C. Stable transformation and reverse genetic analysis of Penium margaritaceum: A platform for studies of Charophycean Green Algae, the immediate ancestors of land plants. Plant J. 2014, 77, 339–351. [Google Scholar] [CrossRef]
- Krejci, M.R.; Wasserman, B.; Finney, L.; McNulty, I.; Legnini, D. Selectivity in biomineralization of barium and strontium. J. Struc. Biol. 2011, 176, 192–202. [Google Scholar] [CrossRef]
- Domozych, D.S.; Lambiasse, L.; Kiemle, S.N.; Gretz, M.R. Cell-wall development and bipolar growth in the desmid Penium margaritaceum (Zygnematophyceae, Streptophyta): Asymmetry in a symmetric world. J. Phycol. 2009, 45, 879–893. [Google Scholar] [CrossRef]
- Url, T.; Höftberger, M.; Meindl, U. Microtubule-microfilament-controlled nuclear migration in the desmid Euastrum oblongum. J. Phycol. 2004, 28, 537–544. [Google Scholar] [CrossRef]
- Domozych, D.S.; Kort, S.; Benton, S.; Yu, T. The extracellular polymeric substance of the green alga Penium margaritaceum and its role in biofilm formation. Biofilms 2005, 2, 129–144. [Google Scholar] [CrossRef]
- Nichols, H.W. Growth media-freshwater. In Handbook of Phycological Methods: Culture Methods and Growth Measurements, 1st ed.; Stein, J.R., Ed.; Cambridge University Press: New York, NY, USA, 1973; Volume 1, pp. 39–78. [Google Scholar]
- Clausen, M.H.; Willats, W.G.T.; Knox, J.P. Synthetic methyl hexagalacturonate hapten inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7. Carbohydr. Res. 2003, 338, 1797–1800. [Google Scholar] [CrossRef] [PubMed]
- Liners, F.; Letesson, J.J.; Didembourg, C.; van Cutsem, P. Monoclonal antibodies against pectin: Recognition of a conformation induced by calcium. Plant Physiol. 1989, 91, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Liners, F.; Thibault, J.F.; van Cutsem, P. Influence of the degree of polymerization of oligogalacturonates and of esterification pattern of pectin on their recognition by monoclonal antibodies. Plant Physiol. 1992, 99, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- Willats, W.G.T.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.J.; Benen, J.; Christensen, T.M.; Visser, J.; Voragen, A.; Mikkelsen, J.D.; Knox, J.P. Analysis of pectic epitopes recognised by hybridoma and phage display monoclonal antibodies using defined oligosaccharides, polysaccharides, and enzymatic degradation. Carbohydr. Res. 2000, 327, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S.; Brechka, H.; Britton, A.; Toso, M. Cell wall growth and modulation dynamics in a model unicellular green alga—Penium margaritaceum: Live cell labeling with monoclonal antibodies. J. Bot. 2011, 2011, e632165. [Google Scholar]
- Domozych, D.S.; Sørensen, I.; Sacks, C.; Brechka, H.; Andreas, A.; Fangel, J.U.; Rose, J.K.C.; Willats, W.G.T.; Popper, Z.A. Disruption of the microtubule network alters cellulose deposition and causes major changes in pectin distribution in the cell wall of the green alga, Penium margaritaceum. J. Exp. Bot. 2014, 65, 465–479. [Google Scholar] [CrossRef] [PubMed]
- Nebenführ, A.; Staehelin, L.A. Mobile factories: Golgi dynamics in plant cells. Trends Plant Sci. 2001, 6, 160–167. [Google Scholar] [CrossRef] [PubMed]
- DaSilva, L.L.; Snapp, E.L.; Denecke, J.; Lippincott-Schwartz, J.; Hawes, C.; Brandizzi, F. Endoplasmic reticulum export sites and Golgi bodies behave as single mobile secretory units in plant cells. Plant Cell 2004, 16, 1753–1771. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S. Exopolymer production by the green alga Penium margaritaceum: Implications for biofilm residency. Int. J. Plant Sci. 2007, 168, 763–774. [Google Scholar] [CrossRef]
- Ochs, J.; LaRue, T.; Tinaz, B.; Yongue, C.; Domozych, D.S. The cortical cytoskeletal network and cell-wall dynamics in the unicellular charophycean green alga Penium margaritaceum. Ann. Bot. 2014. [CrossRef]
- Ambrose, J.C.; Cyr, R.J. Mitotic spindle organization by the preprophase band. Mol. Plant 2008, 6, 950–960. [Google Scholar] [CrossRef]
- Rasmussen, C.G.; Wright, A.J.; Müller, S. The role of the cytoskeleton and associated proteins in determination of the plant cell division plane. Plant J. 2013, 75, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.; Boue, S.; Belmonte, J.C.I. Dedifferentiation, transdifferentiation and reprogramming: Three routes to regeneration. Nat. Rev. Mol. Cell Biol. 2011, 12, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Eeckhaut, T.; Lakshmanan, P.S.; Deryckere, D.; van Bockstaele, E.; van Huylenbroek, J. Progress in plant protoplast research. Planta 2013, 238, 991–1003. [Google Scholar] [CrossRef]
- Davey, M.R.; Anthony, P.; Patel, D.; Power, J.B. Plant protoplasts: Isolation, culture and plant regeneration. In Plant Cell Culture: Essential Methods, 1st ed.; Davey, M.R., Anthony, P., Eds.; Wiley and Sons: New York, NY, USA, 2010; pp. 153–173. [Google Scholar]
- Hori, K.; Maruyama, F.; Fujisawa, T.; Togashi, T.; Yamamoto, N.; Seo, M.; Sato, S.; Yamada, T.; Mori, H.; Tajima, N.; et al. Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation. Nat. Commun. 2014, 5, e3978. [Google Scholar]
- Timme, R.E.; Bachvaroff, T.S.; Delwiche, C.F. Broad phylogenomic sampling and the sister lineage of land plants. PLoS One 2012, 7, e29696. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domozych, D.S. Penium margaritaceum: A Unicellular Model Organism for Studying Plant Cell Wall Architecture and Dynamics. Plants 2014, 3, 543-558. https://doi.org/10.3390/plants3040543
Domozych DS. Penium margaritaceum: A Unicellular Model Organism for Studying Plant Cell Wall Architecture and Dynamics. Plants. 2014; 3(4):543-558. https://doi.org/10.3390/plants3040543
Chicago/Turabian StyleDomozych, David S. 2014. "Penium margaritaceum: A Unicellular Model Organism for Studying Plant Cell Wall Architecture and Dynamics" Plants 3, no. 4: 543-558. https://doi.org/10.3390/plants3040543