Identification of Leaf Promoters for Use in Transgenic Wheat

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construct Generation for Transient Expression Analysis in Nicotiana benthamiana Leaves
2.2. Construct Generation for Transient and Stable GUS Expression Analysis in Wheat
2.3. Construct Generation for Stable Expression of a Small FLAG-Tagged Protein in Wheat Leaves
2.4. Agrobacterium-Mediated Transient Expression in Nicotiana benthamiana Leaves
2.5. Transient Expression in Wheat Leaves
2.6. Histochemical GUS Assays
2.7. Production and Selection of Transgenic Wheat Plants
2.8. Plant Growth Conditions
2.9. RNA Isolation, cDNA Synthesis and qPCR
2.10. Protein Extraction and Western Blot Analysis for FLAG Protein
3. Results
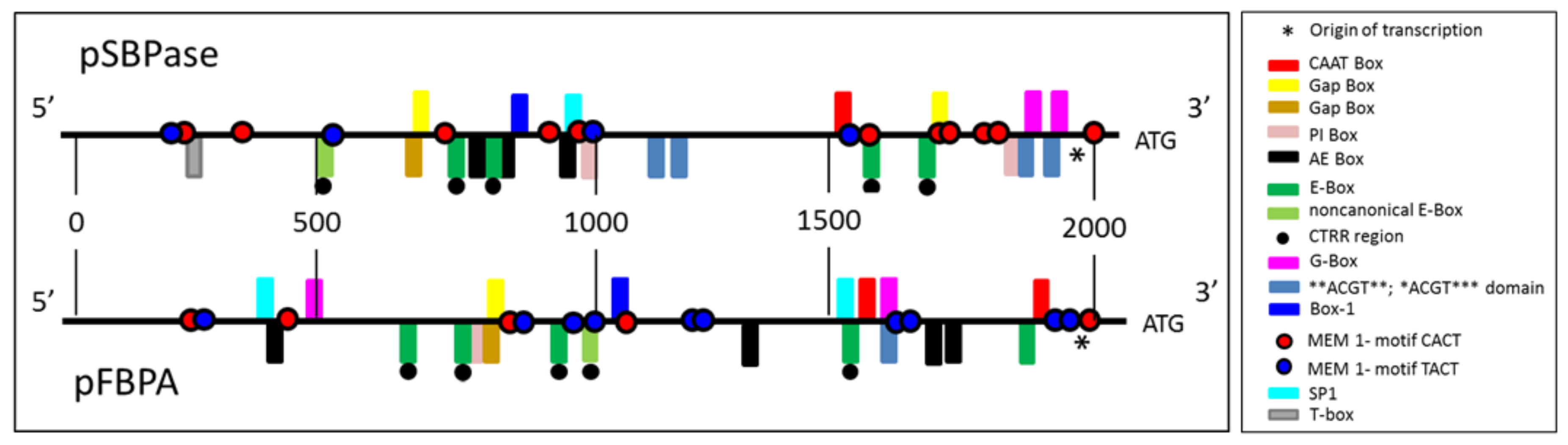
3.1. Identification and Analysis of Brachypodium distachyon SBPase and FBPA Promoters
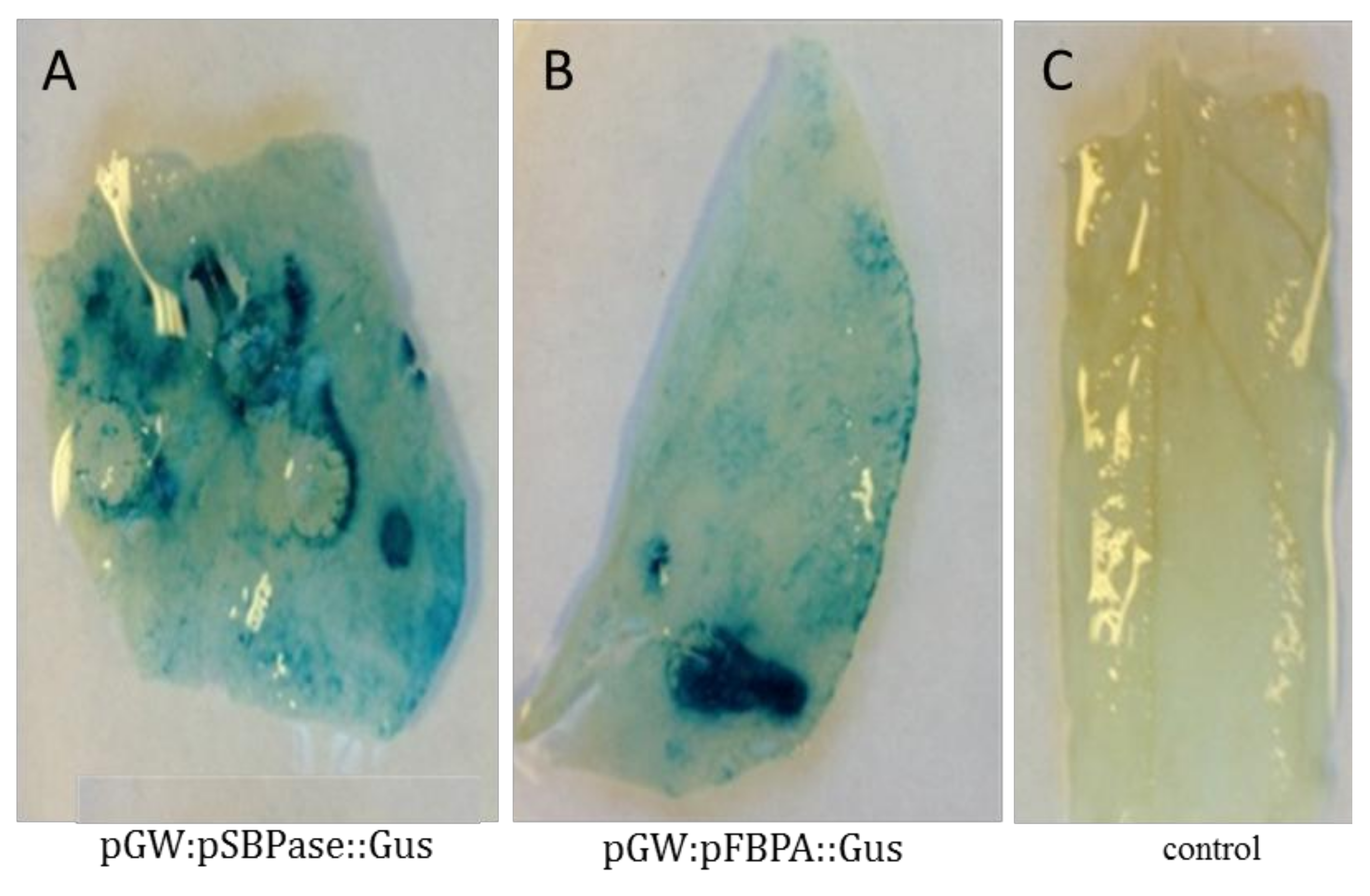
3.2. Agrobacterium-Mediated Transient Expression Analysis in Nicotiana benthamiana Leaves
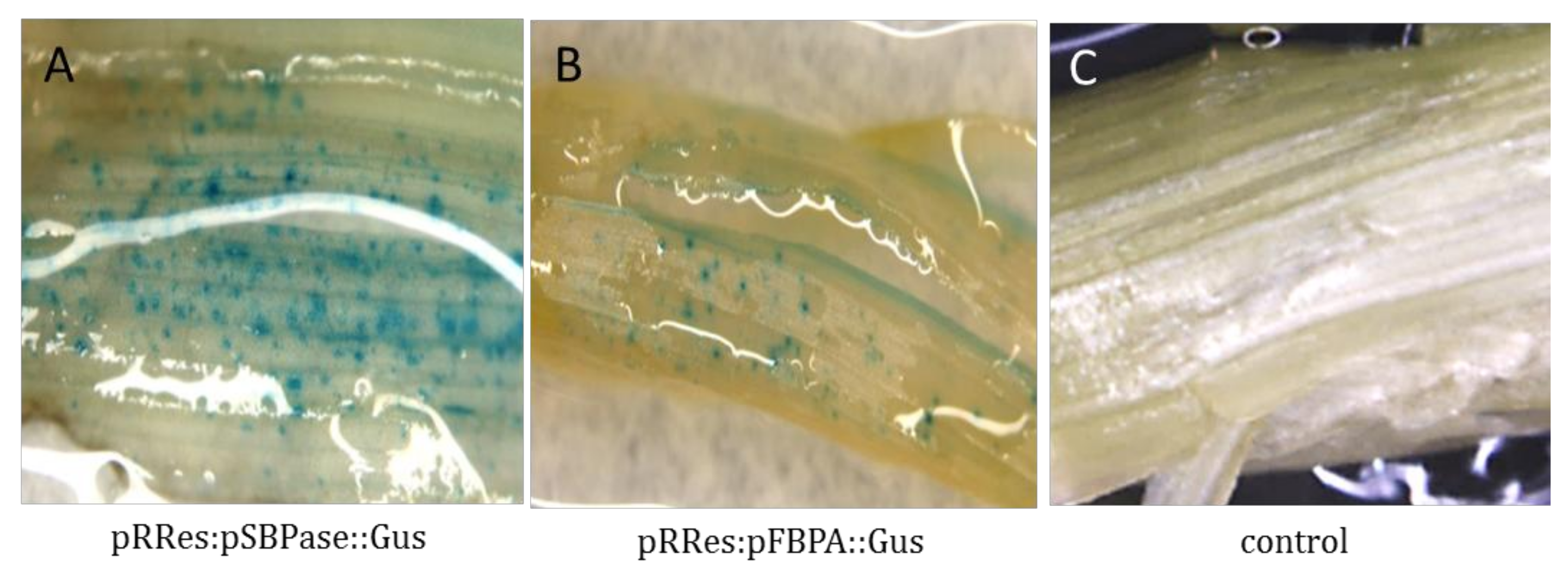
3.3. Transient Expression Analysis in Wheat
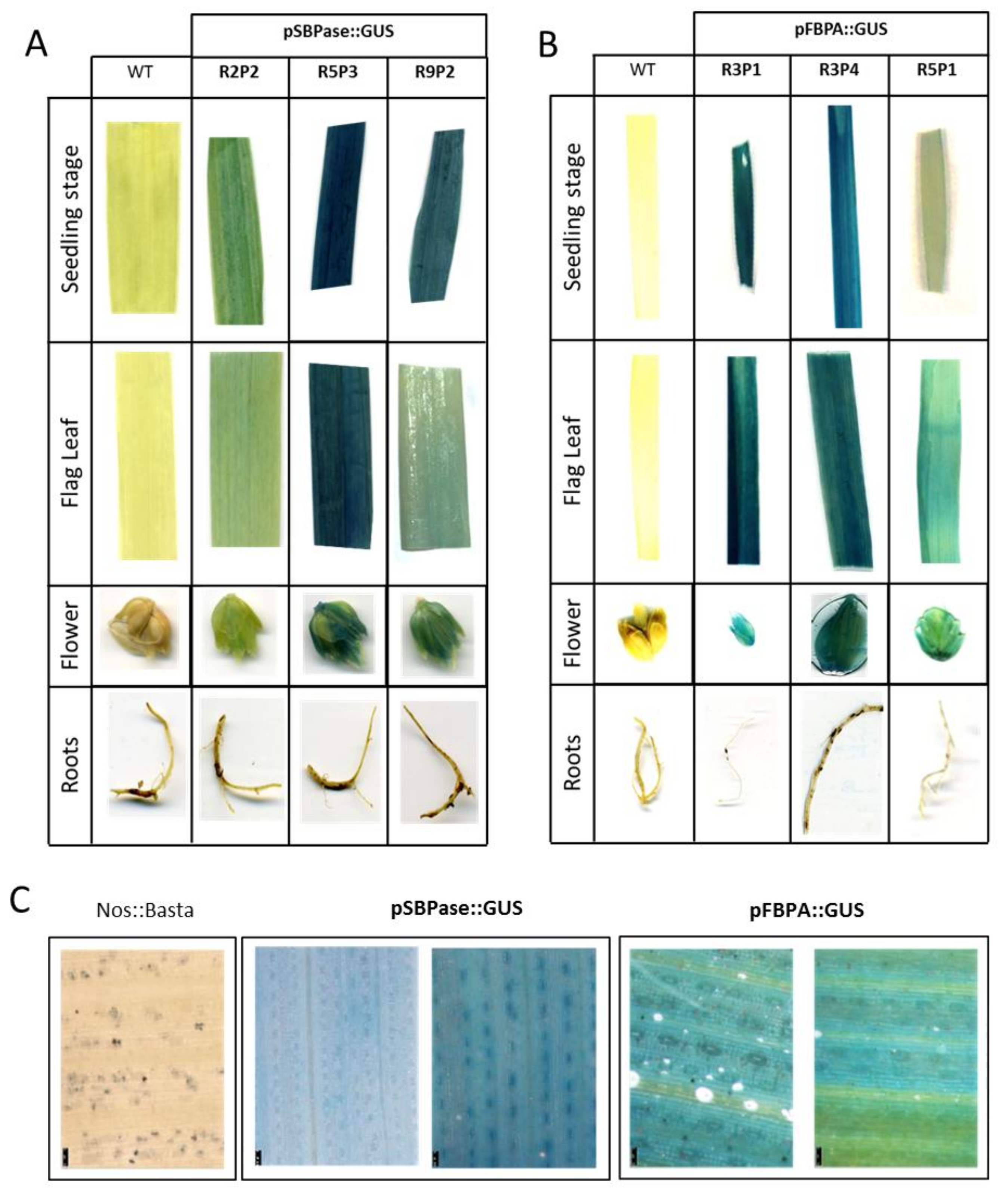
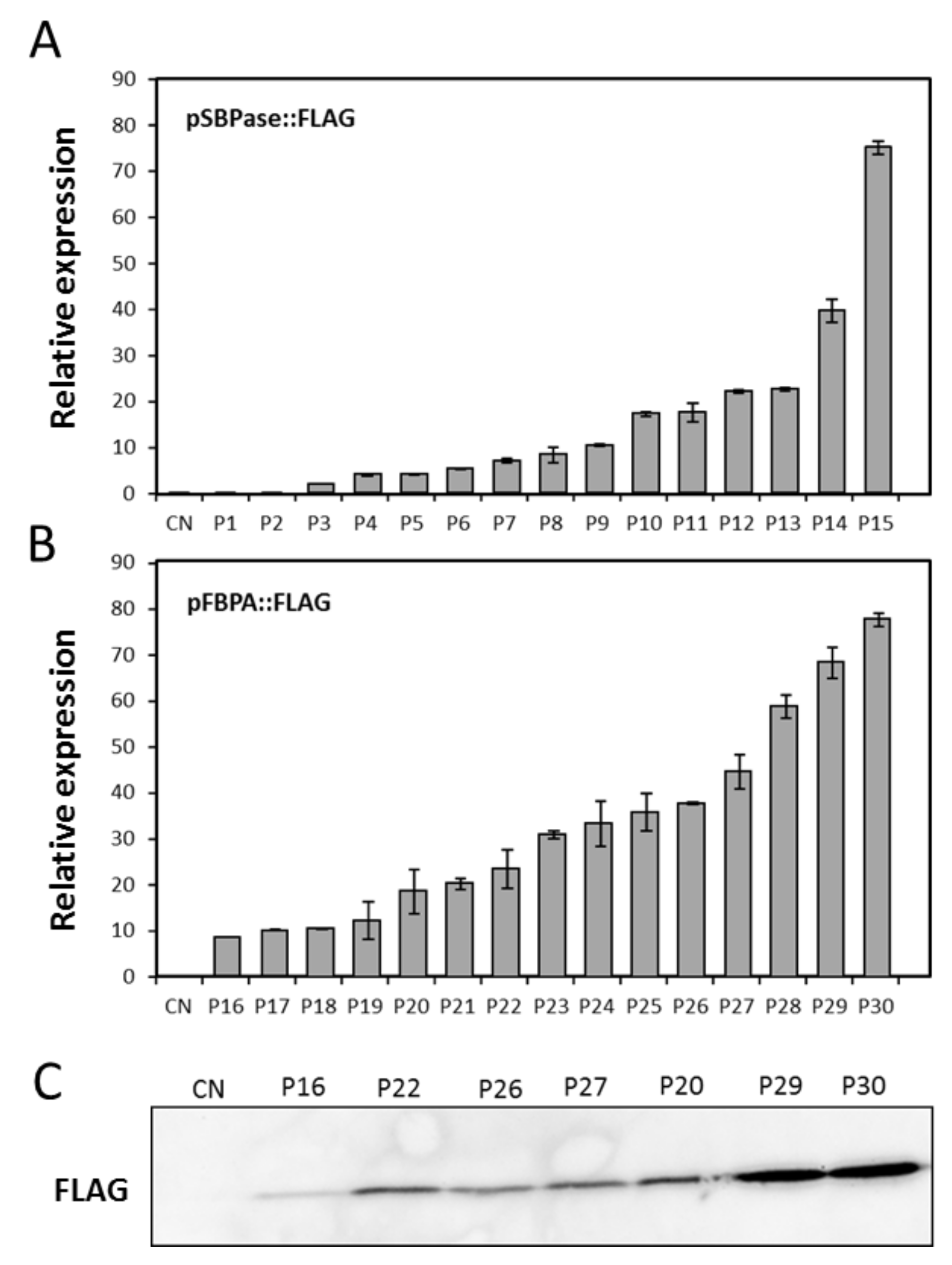
3.4. Stable Expression Analysis in Wheat
3.5. Evaluation of the SBPase or FBPA Promoters to Drive Expression of an Introduced Coding Sequence from Algae
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Braun, H.J.; Atlin, G.; Payne, T. Multi-location testing as a tool to identify plant response to global climate change. In Climate Change and Crop Production; Climate Change Series; Reynolds, M.P., Ed.; CABI: Surrey, UK, 2010; pp. 115–138. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J. The challenge of feeding 9–10 billion people equitably and sustainably. J. Agric. Sci. 2014, 152, S2–S8. [Google Scholar] [CrossRef]
- RSOL. Royal Society of London, Reaping the Benefits: Science and the Sustainable Intensification of Global Agriculture; Royal Society: London, UK, 2009. [Google Scholar]
- Davies, W.J.; Ribaut, J.M. Stress resilience in crop plants: Strategic thinking to address local food production problems. Food Energy Secur. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Le Quere, C.; Raupach, M.R.; Canadell, J.G.; Marland, G.; Bopp, L.; Ciais, P.; Conway, T.J.; Doney, S.C.; Feely, R.A.; Foster, P.; et al. Trends in the sources and sinks of carbon dioxide. Nat. Geosci. 2009, 2, 831–836. [Google Scholar] [CrossRef]
- Solomon, S. Climate Change 2007—The Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- Driever, S.M.; Simkin, A.J.; Alotaibi, S.; Fisk, S.J.; Madgwick, P.J.; Sparks, C.A.; Jones, H.D.; Lawson, T.; Parry, M.A.J.; Raines, C.A. Increased SBPase activity improves photosynthesis and grain yield in wheat grown in greenhouse conditions. Philos. Trans. R. Soc. B 2017, 372, 1730. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Florian, A.; Arrivault, S.; Stitt, M.; Fernie, A.R.; Bauwe, H. Glycine decarboxylase controls photosynthesis and plant growth. FEBS Lett. 2012, 586, 3692–3697. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; McAusland, L.; Lawson, T.; Raines, C.A. Over-expression of the RieskeFeS protein increases electron transport rates and biomass yield. Plant Physiol. 2017, 175, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Chida, H.; Nakazawa, A.; Akazaki, H.; Hirano, T.; Suruga, K.; Ogawa, M.; Satoh, T.; Kadokura, K.; Yamada, S.; Hakamata, W.; et al. Expression of the algal cytochrome c6 gene in Arabidopsis enhances photosynthesis and growth. Plant Cell Physiol. 2007, 48, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Lopez-Calcagno, P.E.; Davey, P.A.; Headland, L.R.; Lawson, T.; Timm, S.; Bauwe, H.; Raines, C.A. Simultaneous stimulation of sedoheptulose 1,7-bisphosphatase, fructose 1,6-bisphophate aldolase and the photorespiratory glycine decarboxylase H-protein increases CO2 assimilation, vegetative biomass and seed yield in Arabidopsis. Plant Biotechnol. J. 2017, 15, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; McAusland, L.; Headland, L.R.; Lawson, T.; Raines, C.A. Multigene manipulation of photosynthetic carbon assimilation increases co2 fixation and biomass yield in tobacco. J. Exp. Bot. 2015, 66, 4075–4090. [Google Scholar] [CrossRef] [PubMed]
- Odell, J.T.; Nagy, F.; Chua, N.H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 1985, 313, 810–812. [Google Scholar] [CrossRef] [PubMed]
- Exposito-Rodriguez, M.; Laissue, P.P.; Lopez-Calcagno, P.E.; Mullineaux, P.M.; Raines, C.A.; Simkin, A.J. Development of pGEMINI, a plant gateway destination vector allowing the simultaneous integration of two cDNA via a single LR-clonase reaction. Plants 2017, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Richins, R.D.; Scholthof, H.B.; Shepherd, R.J. Sequence of figwort mosaic-virus DNA (Caulimovirus group). Nucleic Acids Res. 1987, 15, 8451–8466. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.C.; Simkin, A.J.; Janssen, B.J.; Templeton, K.R.; Loucas, H.M.; Simons, J.L.; Karunairetnam, S.; Gleave, A.P.; Clark, D.G.; Klee, H.J. The decreased apical dominance1/petunia hybrida carotenoid cleavage dioxygenase8 gene affects branch production and plays a role in leaf senescence, root growth, and flower development. Plant Cell 2005, 17, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Underwood, B.A.; Auldridge, M.; Loucas, H.M.; Shibuya, K.; Schmelz, E.; Clark, D.G.; Klee, H.J. Circadian regulation of the phccd1 carotenoid cleavage dioxygenase controls emission of beta-ionone, a fragrance volatile of petunia flowers. Plant Physiol. 2004, 136, 3504–3514. [Google Scholar] [CrossRef] [PubMed]
- De Borne Dorlhac, F.; Vincentz, M.; Chupeau, Y.; Vaucheret, H. Co-suppression of nitrate reductase host genes and transgenes in transgenic tobacco plants. Mol. Gen. Genet. 1994, 243, 613–621. [Google Scholar] [CrossRef]
- Palauqui, J.C.; Elmayan, T.; De Borne, F.D.; Crete, P.; Charles, C.; Vaucheret, H. Frequencies, timing, and spatial patterns of co-suppression of nitrate reductase and nitrite reductase in transgenic tobacco plants. Plant Physiol. 1996, 112, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Dupree, P.; Pwee, K.H.; Gray, J.C. Expression of photosynthesis gene-promoter fusions in leaf epidermal-cells of transgenic tobacco plants. Plant J. 1991, 1, 115–120. [Google Scholar] [CrossRef]
- Xu, M.; Bernat, G.; Singh, A.; Mi, H.; Rogner, M.; Pakrasi, H.B.; Ogawa, T. Properties of mutants of Synechocystis sp. Strain PCC 6803 lacking inorganic carbon sequestration systems. Plant Cell Physiol. 2008, 49, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, M.; Chen, H.C.; Simkin, A.J.; Romer, S.; Shipton, C.A.; Drake, R.; Schuch, W.; Bramley, P.M. Upregulation of two ripening-related genes from a nonclimacteric plant (pepper) in a transgenic climacteric plant (tomato). Plant J. 1998, 13, 351–361. [Google Scholar] [CrossRef]
- Manac’h, N.; Kuntz, M. Stress induction of a nuclear gene encoding for a plastid protein is mediated by photo-oxidative events. Plant Physiol. Biochem. 1999, 37, 859–868. [Google Scholar] [CrossRef]
- Simkin, A.J.; Gaffe, J.; Alcaraz, J.P.; Carde, J.P.; Bramley, P.M.; Fraser, P.D.; Kuntz, M. Fibrillin influence on plastid ultrastructure and pigment content in tomato fruit. Phytochemistry 2007, 68, 1545–1556. [Google Scholar] [CrossRef] [PubMed]
- Muller-Rober, B.; Ellenberg, J.; Provart, N.; Willmitzer, L.; Busch, H.; Becker, D.; Dietrich, P.; Hoth, S.; Hedrich, R. Cloning and electrophysiological analysis of kst1, an inward rectifying k+ channel expressed in potato guard cells. EMBO J. 1995, 14, 2409–2416. [Google Scholar] [PubMed]
- Kelly, G.; Moshelion, M.; David-Schwartz, R.; Halperin, O.; Wallach, R.; Attia, Z.; Belausov, E.; Granot, D. Hexokinase mediates stomatal closure. Plant J. 2013, 75, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Lawson, T.; Simkin, A.J.; Kelly, G.; Granot, D. Mesophyll photosynthesis and guard cell metabolism impacts on stomatal behaviour. New Phytol. 2014, 203, 1064–1081. [Google Scholar] [CrossRef] [PubMed]
- Baumlein, H.; Nagy, I.; Villarroel, R.; Inze, D.; Wobus, U. Cis-analysis of a seed protein gene promoter—The conservative RY repeat CATGCATG within the legumin box is essential for tissue-specific expression of a legumin gene. Plant J. 1992, 2, 233–239. [Google Scholar] [PubMed]
- Thilmony, R.; Guttman, M.E.; Lin, J.W.; Blechl, A.E. The wheat HMW-glutenin 1Dy10 gene promoter controls endosperm expression in Brachypodium distachyon. GM Crops Food 2014, 5, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Cui, C.J.; Chen, L.; Sun, Y.L.; Wang, F.F.; Hussain, J.; Li, Y.; Wang, C.; Wang, C.; Chen, M.J.; et al. Isolation and characterization of an endosperm-specific promoter from wheat (Triticum aestivum L.). Z. Naturforsch. C 2012, 67, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Lamacchia, C.; Shewry, P.R.; Di Fonzo, N.; Forsyth, J.L.; Harris, N.; Lazzeri, P.A.; Napier, J.A.; Halford, N.G.; Barcelo, P. Endosperm-specific activity of a storage protein gene promoter in transgenic wheat seed. J. Exp. Bot. 2001, 52, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; McCarthy, J.; Petiard, V.; Tanksley, S.; Lin, C. Oleosin Genes and Promoters from Coffee. U.S. Patent WO2007005928A2, 11 January 2007. [Google Scholar]
- Christensen, A.H.; Sharrock, R.A.; Quail, P.H. Maize polyubiquitin genes—Structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 1992, 18, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Mcelroy, D.; Zhang, W.G.; Cao, J.; Wu, R. Isolation of an efficient actin promoter for use in rice transformation. Plant Cell 1990, 2, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Dasgupta, I. Downstream promoter sequence of an Indian isolate of Rice tungro bacilliform virus alters tissue-specific expression in host rice and acts differentially in heterologous system. Plant Mol. Biol. 2007, 65, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyyapakrasi, M.; Peng, J.Y.; Elmer, J.S.; Laco, G.; Shen, P.; Kaniewska, M.B.; Kononowicz, H.; Wen, F.J.; Hodges, T.K.; Beachy, R.N. Specificity of a promoter from the Rice tungro bacilliform virus for expression in phloem tissues. Plant J. 1993, 4, 71–79. [Google Scholar] [CrossRef]
- Mukherjee, S.; Stasolla, C.; Brule-Babel, A.; Ayele, B.T. Isolation and characterization of rubisco small subunit gene promoter from common wheat (Triticum aestivum L.). Plant Signal. Behav. 2015, 10, e989033. [Google Scholar] [CrossRef] [PubMed]
- Sparks, C.A.; Castleden, C.K.; West, J.; Habash, D.Z.; Madgwick, P.J.; Paul, M.J.; Noctor, G.; Harrison, J.; Wu, R.; Wilkinson, J.; et al. Potential for manipulating carbon metabolism in wheat. Ann. Appl. Biol. 2001, 138, 33–45. [Google Scholar] [CrossRef]
- Peremarti, A.; Twyman, R.M.; Gomez-Galera, S.; Naqvi, S.; Farre, G.; Sabalza, M.; Miralpeix, B.; Dashevskaya, S.; Yuan, D.; Ramessar, K.; et al. Promoter diversity in multigene transformation. Plant Mol. Biol. 2010, 73, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, K.; Suzuki, N.; Iwamae, T.; Inui, M.; Yukawa, H. Increased fructose 1,6-bisphosphate aldolase in plastids enhances growth and photosynthesis of tobacco plants. J. Exp. Bot. 2012, 63, 3001–3009. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, S.; Lawson, T.; Zakhleniuk, O.V.; Lloyd, J.C.; Raines, C.A.; Fryer, M. Increased sedoheptulose-1,7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 2005, 138, 451–460. [Google Scholar] [CrossRef] [PubMed]
- The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. Available online: https://www.nature.com/articles/nature08747.pdf (accessed on 1 March 2018).
- Draper, J.; Mur, L.A.J.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; Routledge, A.P.M. Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Kurose, T.; Hino, T.; Tanaka, K.; Kawamukai, M.; Niwa, Y.; Toyooka, K.; Matsuoka, K.; Jinbo, T.; Kimura, T. Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 2007, 104, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Goodin, M.M.; Zaitlin, D.; Naidu, R.A.; Lommel, S.A. Nicotiana benthamiana: Its history and future as a model for plant–pathogen interactions. Mol. Plant Microbe Interact. 2008, 21, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, L.; Szittya, G.; Silhavy, D.; Burgyan, J. Molecular mechanism of RNA silencing suppression mediated by p19 protein of tombusviruses. EMBO J. 2004, 23, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Kapila, J.; De Rycke, R.; van Montagui, M.; Angenon, G. An agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci. 1997, 122, 101–108. [Google Scholar] [CrossRef]
- Wydro, M.; Kozubek, E.; Lehmann, P. Optimization of transient agrobacterium-mediated gene expression system in leaves of Nicotiana benthamiana. Acta Biochim. Pol. 2006, 53, 289–298. [Google Scholar] [PubMed]
- Jones, H.D.; Doherty, A.; Sparks, C.A. Transient transformation of plants. Methods Mol. Biol. 2009, 513, 131–152. [Google Scholar] [PubMed]
- Rasco-Gaunt, S.; Barcelo, P. Immature inflorescence culture of cereals. A highly responsive system for regeneration and transformation. Methods Mol. Biol. 1999, 111, 71–81. [Google Scholar] [PubMed]
- Sparks, C.A.; Jones, H.D. Genetic transformation of wheat via particle bombardment. Methods Mol. Biol. 2014, 1099, 201–218. [Google Scholar] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions—Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher-plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [PubMed]
- Christensen, A.H.; Quail, P.H. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgenic Res. 1996, 5, 213–218. [Google Scholar] [CrossRef]
- Lopez-Calcagno, P.E.; Abuzaid, A.O.; Lawson, T.; Raines, C.A. Arabidopsis CP12 mutants have reduced levels of phosphoribulokinase and impaired function of the Calvin-Benson cycle. J. Exp. Bot. 2017, 68, 2285–2298. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.P.; Willingham, N.M.; Lloyd, J.C.; Raines, C.A. Reduced sedoheptulose-1,7-bisphosphatase levels in transgenic tobacco lead to decreased photosynthetic capacity and altered carbohydrate accumulation. Planta 1998, 204, 27–36. [Google Scholar] [CrossRef]
- Willingham, N.M.; Lloyd, J.C.; Raines, C.A. Molecular cloning of the Arabidopsis thaliana sedoheptulose-1,7-biphosphatase gene and expression studies in wheat and Arabidopsis thaliana. Plant Mol. Biol. 1994, 26, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Matzke, A.J.; Matzke, M.A. Position effects and epigenetic silencing of plant transgenes. Curr. Opin. Plant Biol. 1998, 1, 142–148. [Google Scholar] [CrossRef]
- Kooter, J.M.; Matzke, M.A.; Meyer, P. Listening to the silent genes: Transgene silencing, gene regulation and pathogen control. Trends Plant Sci. 1999, 4, 340–347. [Google Scholar] [CrossRef]
- Selker, E.U. Gene silencing: Repeats that count. Cell 1999, 97, 157–160. [Google Scholar] [CrossRef]
- Peach, C.; Velten, J. Transgene expression variability (position effect) of CAT and GUS reporter genes driven by linked divergent T-DNA promoters. Plant Mol. Biol. 1991, 17, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, S.; Chakravarthy, S.; Pental, D.; Burma, P.K. Analysis of promoter activity in transgenic plants by normalizing expression with a reference gene: Anomalies due to the influence of the test promoter on the reference promoter. J. Biosci. 2009, 34, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, V.A.; Moscone, E.A.; Papp, I.; Michalowski, S.; Phelan, T.; Spiker, S.; Neuhuber, F.; Matzke, M.; Matzke, A.J.M. Molecular and cytogenetic analysis of stably and unstably expressed transgene loci in tobacco. Plant Cell 1997, 9, 1251–1264. [Google Scholar] [CrossRef] [PubMed]
- Molina, C.; Grotewold, E. Genome wide analysis of Arabidopsis core promoters. BMC Genom. 2005, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusnetsov, V.; Landsberger, M.; Meurer, J.; Oelmuller, R. The assembly of the CAAT-box binding complex at a photosynthesis gene promoter is regulated by light, cytokinin, and the stage of the plastids. J. Biol. Chem. 1999, 274, 36009–36014. [Google Scholar] [CrossRef] [PubMed]
- Akyildiz, M.; Gowik, U.; Engelmann, S.; Koczor, M.; Streubel, M.; Westhoff, P. Evolution and function of a cis-regulatory module for mesophyll-specific gene expression in the C4 dicot Flaveria trinervia. Plant Cell 2007, 19, 3391–3402. [Google Scholar] [CrossRef] [PubMed]
- Gowik, U.; Schulze, S.; Saladie, M.; Rolland, V.; Tanz, S.K.; Westhoff, P.; Ludwig, M. A mem1-like motif directs mesophyll cell-specific expression of the gene encoding the C4 carbonic anhydrase in Flaveria. J. Exp. Bot. 2017, 68, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.C.; Raines, C.; John, U.P.; Dyer, T.A.A. The chloroplast FBPase gene of wheat: Structure and expression of the promoter in photosynthetic and meristematic cells of transgenic tobacco plants. Mol. Gen. Genet. 1991, 225, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Raines, C.; Llloyd, J.C.; Willingham, N.M.; Potts, S.; Dyer, T.A.A. cDNA and gene sequences of wheat chloroplast sedoheptulose-1,7-bisphosphatase reveal homology with fructose-1,6-bisphosphatases. FEBS J. 1992, 205, 1053–1059. [Google Scholar] [CrossRef]
- Kwon, H.B.; Park, S.C.; Peng, H.P.; Goodman, H.M.; Dewdney, J.; Shih, M.C. Identification of a light-responsive region of the nuclear gene encoding the b subunit of chloroplast glyceraldehyde 3-phosphate dehydrogenase from Arabidopsis thaliana. Plant Physiol. 1994, 105, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Conley, T.R.; Park, S.C.; Kwon, H.B.; Peng, H.P.; Shih, M.C. Characterization of cis-acting elements in light regulation of the nuclear gene encoding the A subunit of chloroplast isozymes of glyceraldehyde-3-phosphate dehydrogenase from Arabidopsis thaliana. Mol. Cell. Biol. 1994, 14, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kwon, H.B.; Shih, M.C. Cis-acting elements essential for light regulation of the nuclear gene encoding the A subunit of chloroplast glyceraldehyde 3-phosphate dehydrogenase in Arabidopsis thaliana. Plant Physiol. 1996, 112, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Choudhury, S.R.; Singh, S.K.; Das, K.P. Functional analysis of light-regulated promoter region of AtPolλ gene. Planta 2012, 235, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Baum, K.; Wienand, U.; Meier, I. Reduction of G-box binding factor DNA binding activity, but not g-box binding factor abundance, causes the downregulation of RBCS2 expression during early tomato fruit development. FEBS Lett. 1999, 454, 95–99. [Google Scholar] [CrossRef]
- Menkens, A.E.; Schindler, U.; Cashmore, A.R. The G-box: A ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem. Sci. 1995, 20, 506–510. [Google Scholar] [CrossRef]
- Williams, M.E.; Foster, R.; Chua, N.H. Sequences flanking the hexameric G-box core CACGTG affect the specificity of protein binding. Plant Cell 1992, 4, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Ishige, F.; Takaichi, M.; Foster, R.; Chua, N.H.; Oeda, K. A G-box motif (GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 1999, 18, 443–448. [Google Scholar] [CrossRef]
- De Boer, K.; Tilleman, S.; Pauwels, L.; Vanden Bossche, R.; De Sutter, V.; Vanderhaeghen, R.; Hilson, P.; Hamill, J.D.; Goossens, A. APETALA2/ETHYLENE RESPONSE FACTOR and basic helix-loop-helix tobacco transcription factors cooperatively mediate jasmonate-elicited nicotine biosynthesis. Plant J. Cell Mol. Biol. 2011, 66, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [PubMed]
- Ezer, D.; Shepherd, S.J.K.; Brestovitsky, A.; Dickinson, P.; Cortijo, S.; Charoensawan, V.; Box, M.S.; Biswas, S.; Jaeger, K.E.; Wigge, P.A. The g-box transcriptional regulatory code in Arabidopsis. Plant Physiol. 2017, 175, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Michael, T.P.; McClung, C.R. Enhancer trapping reveals widespread circadian clock transcriptional control in Arabidopsis. Plant Physiol. 2003, 132, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Hudson, M.E.; Quail, P.H. Identification of promoter motifs involved in the network of phytochrome a-regulated gene expression by combined analysis of genomic sequence and microarray data. Plant Physiol. 2003, 133, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, J.F.; Huq, E.; Quail, P.H. Direct targeting of light signals to a promoter element-bound transcription factor. Science 2000, 288, 859–863. [Google Scholar] [PubMed]
- Darlington, T.K.; Wager-Smith, K.; Ceriani, M.F.; Staknis, D.; Gekakis, N.; Steeves, T.D.; Weitz, C.J.; Takahashi, J.S.; Kay, S.A. Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim. Science 1998, 280, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, F.C.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the clock protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Kawagoe, Y.; Murai, N. Four distinct nuclear proteins recognize in vitro the proximal promoter of the bean seed storage protein beta-phaseolin gene conferring spatial and temporal control. Plant J. 1992, 2, 927–936. [Google Scholar] [PubMed]
- Chatthai, M.; Forward, B.S.; Yevtushenko, D.; Stefanov, I.; Osuska, L.; Osusky, M.; Misra, S. 2S storage protein gene of Douglas-fir: Characterization and activity of promoter in transgenic tobacco seeds. Plant Physiol. Biochem. 2004, 42, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Qian, T.Z.; Caillet, V.; Michoux, F.; Ben Amor, M.; Lin, C.W.; Tanksley, S.; McCarthy, J. Oleosin gene family of Coffea canephora: Quantitative expression analysis of five oleosin genes in developing and germinating coffee grain. J. Plant Physiol. 2006, 163, 691–708. [Google Scholar] [CrossRef] [PubMed]
- Munoz, E.; Brewer, M.; Baler, R. Modulation of BMAL/CLOCK/E-BOX complex activity by a CT-rich cis-acting element. Mol. Cell. Endocrinol. 2006, 252, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Li, X.; Li, K.W.; Liu, H.T.; Lin, C.T. Multiple bHLH proteins form heterodimers to mediate cry2-dependent regulation of flowering-time in Arabidopsis. PLoS Genet. 2013, 9, e1003861. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.P.; Patel, S.P.; McCarthy, J.J.; Rabchevsky, A.G.; Goldhamer, D.J.; Esser, K.A. A non-canonical E-box within the MyoD core enhancer is necessary for circadian expression in skeletal muscle. Nucleic Acids Res. 2012, 40, 3419–3430. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.Y.; Ichida, H.; Matsui, M.; Obokata, J.; Sakurai, T.; Satou, M.; Seki, M.; Shinozaki, K.; Abe, T. Identification of plant promoter constituents by analysis of local distribution of short sequences. BMC Genom. 2007, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.K.; Li, L.S.; Xu, L.N.; Zhou, J.; Zhuang, H.H.; Gong, X.D.; Wang, M.X.; Sun, S.S.M.; Zhuge, Q. Isolation and functional analysis of the poplar RBcS gene promoter. Plant Mol. Biol. Rep. 2013, 31, 120–127. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, F.T.; Jia, D.M.; Li, J.T.; Zhang, Y.M.; Jia, C.G.; Wang, D.P.; Pan, H.Y. Cloning and functional analysis of the promoter of a stress-inducible gene (ZmRXO1) in maize. Plant Mol. Biol. Rep. 2015, 33, 200–208. [Google Scholar] [CrossRef]
- Chan, C.S.; Guo, L.; Shih, M.C. Promoter analysis of the nuclear gene encoding the chloroplast glyceraldehyde-3-phosphate dehydrogenase b subunit of Arabidopsis thaliana. Plant Mol. Biol. 2001, 46, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Huang, X.H.; Li, L.L.; Deng, G.; Cheng, H.; Rong, X.F.; Li, J.B.; Cheng, S.Y. Molecular cloning and characterization of GbDXS and GbGGPPS gene promoters from Ginkgo biloba. Genet. Mol. Res. 2013, 12, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Morishima, A. Identification of preferred binding sites of a light-inducible DNA-binding factor (MNF1) within 5′-upstream sequence of C4-type phosphoenolpyruvate carboxylase gene in maize. Plant Mol. Biol. 1998, 38, 633–646. [Google Scholar] [CrossRef] [PubMed]
- Plesch, G.; Ehrhardt, T.; Mueller-Roeber, B. Involvement of TAAAG elements suggests a role for Dof transcription factors in guard cell-specific gene expression. Plant J. Cell Mol. Boil. 2001, 28, 455–464. [Google Scholar] [CrossRef]
- Lam, E.; Benfey, P.N.; Gilmartin, P.M.; Fang, R.X.; Chua, N.H. Site-specific mutations alter in vitro factor binding and change promoter expression pattern in transgenic plants. Proc. Natl. Acad. Sci. USA 1989, 86, 7890–7894. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Element | Signal Sequence | pSBPase Position | pFBPA Position | Functional Description | Reference |
|---|---|---|---|---|---|
| Activation Element (AE)-box | AGAAAC(A/T)(A/T) | 790, 859, 995 | 419, 1323, 1698, 1734 | The AE Box and Gap Box act together and are essential components of light responsive elements | [74,75] |
| ACGT-Box | **ACGT** *ACGT*** | 1214, 1289, 1852, 1934 | 1584 | Responsive to light, UV, drought and ABA | [88] |
| Box-1 | TTTCAAA TTTGAAA | 876 | 1163 | Light responsive element | [89,90] |
| CATT motif | GCATTC CCAAT CAAAT | 1554 | 1559 1892 | Part of a light responsive element. Evolutionarily conserved in eukaryotic organisms, including fungi, plants, and mammals | [67] |
| MEM1-motif YACT | CACT TACT | 241, 340, 755, 949, 1004, 1314, 1319, 1586, 1724, 1729, 1751, 1756, 2070 229, 593, 1082, 1569 | 205, 424, 857, 1176, 1260, 1988 226, 881, 982, 1046, 1243, 1250, 1600, 1666 1951, 1959 | CACT key component of the Mesophyll Expression Module (MEM) 1 found in the cis-regulatory element in the phosphoenolpyruvate carboxylase promoter of the C4 dicot Flaveria trinervia | [68] |
| E-Box | CANNTG | 769, 848, 1587, 1703 | 687, 775, 954, 1536, 1840 | An integral part of the circadian clock’s transcription–translation feedback loop | [86,87] |
| Non-canonical E-Box | CAGCTT | 515 | 1006 | An integral part of the circadian clock’s transcription–translation feedback loop | [93] |
| GAG-Motif | AGAGAGT | 146 | Light responsive element identified in the rbcS promoter from Poplar | [95] | |
| Gap-Box | ATGAA(G/A)A | 724, 1709 | 830 | Identified in the GapB promoter. Deletion of these repeats abolished light induction completely | [73] |
| Gap-Box | CAAATGAA(G/A)A | 721 | 827 | GapA promoter contains three sequences. Deletion of just one results in a six-fold decrease in light induction | [74] |
| G-Box | CACGTA CACGTG CACGTC GTACGTG TACGTG CACATGG CCACGTAA | 1852, 1935 | 1585 598 1584 | The G-box has been identified in the promoters of circadian-regulated genes in plants and is important for phytochrome-regulated transcriptional induction. Confers high-level constitutive expression in dicot and monocot plants | [76,77,78,79,80,81,82,83,84] |
| TAAAGSTKST1 | TAAAG | 941, 954 | 129 | Target site for trans-acting StDof1 protein controlling guard cell-specific gene expression; KST1 gene encodes a K+ influx channel of guard cells | [100] |
| PI-Box | GTGATCAC GTGATCAG GTGATCAA TTGATCAC | 1036 | 804 1847 | Identified in the GapB promoter. Mutation resulted in a reduction in light-activated gene transcription | [97] |
| T-Box | ACTTTG | 242 | Identified in the GapB promoter. Mutation results in a reduction in light-activated gene transcription | [97,98] | |
| SP1 | CC(G/A)CCC | 1013 | 1510 | Light responsive element identified in Zea mays | [98,99] |
| TGACG-motif | TGACG | 1288, 1742 | Cis-acting regulatory element involved in the MeJA-responsiveness. Mutation of the motif in the 35S promoter causes a 50% drop in expression in tobacco leaves | [101] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alotaibi, S.S.; Sparks, C.A.; Parry, M.A.J.; Simkin, A.J.; Raines, C.A. Identification of Leaf Promoters for Use in Transgenic Wheat. Plants 2018, 7, 27. https://doi.org/10.3390/plants7020027
Alotaibi SS, Sparks CA, Parry MAJ, Simkin AJ, Raines CA. Identification of Leaf Promoters for Use in Transgenic Wheat. Plants. 2018; 7(2):27. https://doi.org/10.3390/plants7020027
Chicago/Turabian StyleAlotaibi, Saqer S., Caroline A. Sparks, Martin A. J. Parry, Andrew J. Simkin, and Christine A. Raines. 2018. "Identification of Leaf Promoters for Use in Transgenic Wheat" Plants 7, no. 2: 27. https://doi.org/10.3390/plants7020027