Lateral Transport of Organic and Inorganic Solutes
by

Emilie Aubry
1, 
Sylvie Dinant
1,
Françoise Vilaine
1,
Catherine Bellini
1,2 and
Rozenn Le Hir
1,* 1
Institut Jean-Pierre Bourgin, INRA, AgroParisTech, CNRS, Université Paris-Saclay, 78000 Versailles, France
2
Umeå Plant Science Centre, Department of Plant Physiology, Umeå University, 90183 Umeå, Sweden
*
Author to whom correspondence should be addressed.
Plants 2019, 8(1), 20; https://doi.org/10.3390/plants8010020
Submission received: 14 November 2018
/
Revised: 10 January 2019
/
Accepted: 11 January 2019
/
Published: 15 January 2019
(This article belongs to the Special Issue Plant Phloem Development)
Abstract
:Organic (e.g., sugars and amino acids) and inorganic (e.g., K+, Na+, PO42−, and SO42−) solutes are transported long-distance throughout plants. Lateral movement of these compounds between the xylem and the phloem, and vice versa, has also been reported in several plant species since the 1930s, and is believed to be important in the overall resource allocation. Studies of Arabidopsis thaliana have provided us with a better knowledge of the anatomical framework in which the lateral transport takes place, and have highlighted the role of specialized vascular and perivascular cells as an interface for solute exchanges. Important breakthroughs have also been made, mainly in Arabidopsis, in identifying some of the proteins involved in the cell-to-cell translocation of solutes, most notably a range of plasma membrane transporters that act in different cell types. Finally, in the future, state-of-art imaging techniques should help to better characterize the lateral transport of these compounds on a cellular level. This review brings the lateral transport of sugars and inorganic solutes back into focus and highlights its importance in terms of our overall understanding of plant resource allocation.
1. Introduction
In plants, organic (e.g., organic acids, sugars, amino acids) and inorganic (e.g., K+, Na+, NH4+, NO3−, SO42−, PO42−, Ca2+) solutes are dissolved in a water-based mixture (i.e., phloem and xylem sap) that is distributed throughout the plant. Understanding how these nutrients are transported between organs and how this transport is coordinated is of major interest for the long-term goal of improving resource allocation in plants and in fine plant biomass production. The solute movement between and within the different organs or tissues of a plant depends on the plant’s developmental stage and on the environmental conditions that the plant is facing. Nevertheless, depending on the type of compound, different transport systems act in a complementary way to allow the solutes to reach their targeted tissue and cell (i.e., long-distance versus short-distance transport, apoplasmic versus symplasmic pathway).
Over the long-distance, the transport of water and nutrients is achieved by the plant’s vascular system, which is composed of phloem and xylem tissues [1]. In addition, several compounds, including sugars [2], amino acids [2], minerals [3], ions [4], water [5], miRNA [6], transcription factors [6,7,8,9], hormones [10,11], secondary metabolites [12,13], and more complex molecules like monolignols [14,15], have been shown to be transported inside the different vascular cell types and between the vascular and the perivascular tissues (i.e., the endodermis, the pericycle, and vascular parenchyma cells). More specifically, when addressing the cell-to-cell movement of solutes (e.g., sugars, amino acids, and ions), authors refer equally to “solute exchange” [16], “radial solute exchange”, “radial transport” [2,5], or “lateral transport” [17]. Moreover, this cell-to-cell movement has in some cases been associated with transport between specific tissues in a preferential direction, such as “xylem-to-phloem transfer” or “phloem-to-xylem transfer” [2,18,19].
During the 1960s and 1970s, extensive literature addressing lateral transport in various species was published, and was last reviewed by van Bel in 1990 [2]. These studies unequivocally stress the importance of this process in overall plant resource allocation. Since then, the use of the model plant Arabidopsis thaliana has allowed significant progress to be made in the identification of the molecular actors involved in such processes, even if direct experimental proof of lateral transport in this species is scarce (Table 1). After presenting experimental evidence for the existence of lateral transport in various plant species, this review will detail the anatomical framework in which solute exchanges take place, as well as the molecular actors involved in the process, mostly identified in Arabidopsis thaliana. Of the various organic and inorganic solutes for which lateral transport has been shown (Table 1), this review will not address (or will only mention briefly) xylem-to-phloem transport of calcium, magnesium, sulfate, and nitrogen, because this has been reviewed elsewhere [20,21,22,23,24]. Instead, the review will focus on the transport of photosynthates and on the transport of potassium, sodium, and phosphate as inorganic ions.
2. Evidence for the Existence of Lateral Transport in Vascular Plants
Stout and Hoagland first reported the experimental existence of lateral transport at the end of the 1930s. Radioactive isotopes of potassium, sodium, and phosphorus were used to show that these ions were transported from xylem to phloem in geranium (Pelargonium zonale) and in willow (Salix lasiandra) [25]. Later, during the 1960s and 1970s, numerous articles were published that addressed the lateral transport of organic and inorganic solutes in both herbaceous and woody species (Table 1 and references therein). Most of these studies also used radio-labelled sugars, amino acids, or ions to support their conclusions. Interestingly, not all inorganic ions are equal with regard to lateral transport. According to Pate (1975), a solute should fulfil several physiological criteria in order to move freely within the plant, such as a low degree of involvement in organic linkage, rapid cycling through the leaves, and a high concentration in the vascular tissues [16]. Among the different inorganic ions, potassium (K+), phosphate (PO42−), and sulfate (SO42−) come closest to meeting all of these criteria. Sodium (Na+) is also transferred from xylem to phloem, but to a lesser extent than K+, while calcium ions fail to cross from xylem to phloem, even if perfused at a high concentration [26,27]. Regardless of the plant species, amino acids mainly move from xylem to phloem, while sugars move in both phloem-to-xylem and xylem-to-phloem directions (Table 1). For instance, Van Bel (1984) demonstrated in tomato plants (Lycopersicum esculentum) that the nutritional demands of young leaves were almost completely met by xylem-to-phloem transport of amino acids in the tomato stem [28]. In the legume species, Lupinus albus, it was shown that vegetative apices received 73% of their N and 14% of their C through the xylem [29], while in soybean (Glycine max), 6% of C and 51% of N enters the fruit via xylem-to-phloem transfer in the stem [30]. Phloem-to-xylem transfer of sugars has also been described in several species, including woody species (Table 1 and references therein). Moreover, bidirectional movement of water and ions (e.g., potassium, sodium, phosphorus, calcium, and magnesium) in both angiosperms and gymnosperms (Table 1 and references therein) was reported. Lateral transport of solutes is thus an important component of resource allocation, as has been shown in several species [31] (Table 1 and references therein). However, most of these studies lacked specific data about the anatomical framework in which lateral transport takes place. Over the past 15 years, studies performed on the model plant Arabidopsis thaliana have given us a better picture of the cell types involved and have highlighted the role of vascular and perivascular cells in this process.
3. Going In and Out of the Vascular System: The Role of Specialized Vascular and Perivascular Cells
In higher plants, the vascular system is composed of phloem and xylem tissues, which are organized in an organ-specific manner (Figure 1). Anatomically, the phloem tissue is composed of phloem parenchyma cells, companion cells, and sieve elements, and mainly accounts for the long-distance transport of sugars [49], amino acids [50], phytohormones [51], and nucleic acids [52] from source (carbon fixing) to sink (carbohydrate import dependent) organs. The xylem tissue is composed of xylem parenchyma cells, xylem fibers, and xylem vessels that provide structural support, as well as enable the transport of water [53], minerals [1], and phytohormones [51] from below- to above-ground organs. Additionally, layers of specialized parenchyma cells and perivascular cells surrounding the phloem and xylem tissues, such as the pericycle in roots or the bundle sheath in leaves, also constitute an important interface involved in the lateral distribution of solutes between vascular cell types, as well as from vascular tissue to the outer cell layers, and vice versa.
3.1. The Bundle Sheath and Transfer Cells in Leaves
In leaves, phloem and xylem tissues face the adaxial and the abaxial part of the lamina, respectively, and are organized into bundles encircled by a parenchymatous layer of cells known as the bundle sheath (BS) [54] (Figure 1A). In previous studies, the BS cells of C4 plants have received the most attention because of their specialized role in efficient CO2 fixation in the Calvin–Benson cycle [55], while the role of BS cells in C3 plant leaves has been much less widely explored. Nevertheless, several studies performed on Arabidopsis indicate an important role for BS cells in the lateral transport of organic and inorganic solutes and suggest that BS cells facing phloem and xylem may play distinct roles [56]. In particular, those that face the xylem may form a selective barrier, preventing excessive amounts of ions entering or leaving the xylem [57]. In addition, they might also act as a hydraulic regulatory barrier to reduce loss of water from the veins during abiotic stress conditions [58]. In minor veins of source leaves, structural evidence suggests that sugars will cross the BS cells (those facing the phloem tissue), preferably through plasmodesmata, to reach the phloem parenchyma cells before being loaded into the phloem sieve elements [59] (Figure 1A,B). In addition, an apoplasmic route is also likely for the transport of organic and inorganic solutes between BS cells and vascular tissues, as suggested by the identification of candidate plasma membrane transporters detailed below [56,60,61,62] (Figure 1B).
Besides the important role of the bundle sheath in the lateral transport of solutes, other vascular cell types are involved in cell-to-cell transport between the phloem and the xylem and within each of these tissues. For instance, transfer cells (TCs), which can differentiate from existing vascular or non-vascular cell types (e.g., bundle sheath, xylem or phloem parenchyma cells), and which are characterized by prominent plasma membrane ingrowths, have been suggested to be a site of intense transport activity between the different vascular cell types [63]. Most studied in the phloem of minor veins, TC development occurs quite early (after an average of 10 days of growth) in cotyledons as well as in young leaves of Arabidopsis thaliana plants grown in normal conditions [64]. In adult leaves, a basipetal gradient of wall ingrowth deposition that has parallels with the sink/source transition is observed [64]. Additionally, TCs can develop in response to changing environmental conditions, most probably as a result of an increased demand for solute transport [65]. It has been proposed, for example, that phloem parenchyma transfer cells (PP TCs), which are highly metabolically active, could fuel massive transmembrane transport, notably of sugar compounds, through the plasmodesmata as well as through active and/or passive transporter systems [66]. Finally, the role of PP TCs in controlling membrane transport capacity has been extended to their role as a physical barrier that prevents access of invading pathogens to sugar-rich sieve elements [67]. Even if progress has been made in our understanding of the role of TCs, the molecular actors involved in the transport of organic solutes (i.e., sugar and amino acids) between TCs and phloem companion cells still need to be identified. On the basis of the localization of members of the SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTERS (SWEET) family in the plasma membrane ingrowths of TCs [68,69], the existence of a facilitated transport of sugars should be further explored.
3.2. Vascular Parenchyma Cells, Fibers, and Rays in the Stem
In Arabidopsis thaliana, the floral stem (also referred to as the inflorescence stem) is composed of collateral vascular bundles of phloem, cambium, and xylem, separated from each other by interfascicular fibers (Figure 1C). In particular, fibers referred to as bast fibers or xylary fibers (depending on whether they belong to phloem or xylem tissue) are found in the phloem and xylem [70]. Fibers can also be found in the hypocotyl phloem and xylem during secondary growth [71]. Even if the two types of fibers (i.e., xylary fibers and interfascicular fibers) do not originate from the same vascular cambium (i.e., the interfascicular cambium for interfascicular fibers and the fascicular cambium for xylary fibers), both provide structural support for the plant, thanks to their thick secondary cell walls [72]. It has recently been proposed that Arabidopsis xylary fibers also provide some of the monolignols used for xylem vessel lignification [15]. Xylem parenchyma cells (also referred to as xylary parenchyma cells) are also involved in supplying monolignols to the developing xylem vessels [15]. However, it is not currently known exactly which transport pathway is involved in this type of process. Certain members of the ATP-binding cassette (ABC) transporter family that have been shown to transport monolignols could be good candidates [73,74]. Additionally, whether or not exchanges between parenchyma cells/xylary fibers and developing vascular conducting cells exist for compounds other than monolignols (e.g., sugars) or occur in other tissues (e.g., phloem) remains an open question. Furthermore, if sugar exchanges do exist, questions of whether these are mediated through plasmodesmata, or active and/or passive transport pathways, represent interesting issues that should be addressed (Figure 1D), especially in terms of xylem secondary cell wall formation, which requires a significant amount of sugar to sustain cell wall polysaccharide synthesis.
Like in the Arabidopsis floral stem, in ligneous species, the woody stem is composed of xylem vessels and xylary fibers, but it also contains an additional cell type known as a ray. Rays, which are rarely found in the Arabidopsis floral stem and hypocotyl [71,75], are living cells responsible for the lateral transport of nutrients across the wood and for storage of carbohydrates [72]. Given their role, ray cells might thus be compared to the xylary parenchyma cells found in the Arabidopsis floral stem. In particular, it has been shown that during wood xylem development, there is an increasing requirement of sugars (i.e., glucose, fructose, and phosphate-sugars) that can be used for sustaining secondary cell wall formation [76]. Regarding the route taken by these sugars, they are most probably unloaded from the sieve elements into the wood rays and then reach the developing xylem vessels through a combination of symplasmic and apoplasmic pathways [77,78].
3.3. The Vascular Parenchyma Cells, Pericycle, and Endodermis in the Root
In a sink organ such as the Arabidopsis root, the young stele (composed of phloem, xylem, and vascular parenchyma cells, also known as stelar parenchyma) is surrounded by the pericycle and the endodermis (Figure 1E) [79]. As the roots get older, the pericycle cells start to divide and contribute, along with the dividing procambium, to the generation of the vascular cambium [80].
The term “stelar parenchyma” encompasses both the phloem parenchyma cells and the xylem parenchyma cells. In general, little attention has been given to the exact role of the root phloem parenchyma cells, while the root xylem parenchyma cells have received the most attention, mainly because of their role in ion/metal transport through the root. Nevertheless, experimental proof of lateral transport of organic and/or inorganic solutes between the phloem and the xylem through the vascular parenchyma cells in the roots is still lacking. Interestingly, a pioneering study of onion root (Allium cepa), which established a map of plasmodesmata distribution between the different vascular root cell types, identified several plasmodesmata at the interface between phloem–stelar and parenchyma–xylem vessels, suggesting a possible “direct” symplasmic pathway (i.e., without passing through the pericycle) for lateral transport (Figure 1F) [81].
The study conducted by Ma et al. (2001) also highlighted the fact that the pericycle would also play a significant role in the symplasmic transport of ions and sugars, both along the radial path and in a tangential direction across the radial walls (Figure 1F) [81]. However, direct proof of plasmodesmata-mediated transport of ions still needs to be provided. Later, a more refined analysis of the young root in the model plant Arabidopsis showed the existence of two types of pericycle cells; namely xylem-pole pericycle cells and phloem-pole pericycle cells (Figure 1E,F) [82]. Xylem-pole pericycle cells have been shown to be involved in lateral root formation as well as in the apoplasmic xylem loading of boron [83,84], while phloem-pole pericycle cells are specifically involved in the unloading of molecules from the phloem sieve elements [85]. In a recent study, Ross-Elliott et al. (2018), using a combination of imaging and mathematical modelling, accurately proved the existence of a novel type of plasmodesmata named funnel plasmodesmata, which are specifically responsible for connecting the phloem-pole pericycle cells to the protophloem sieve elements at the Arabidopsis root tip (Figure 1F). Ross-Elliott et al. suggest that sugars unloaded from the protophloem sieve elements move through the funnel plasmodesmata and are then able to access all other cell types through the “regular” plasmodesmata system (Figure 1F). Other molecules, such as macromolecules, are also able to pass through the funnel plasmodesmata, but are subsequently retained in the phloem-pole pericycle cells [85]. These results represent an important advance in our understanding of how these different cell types are connected, but also raise the question of whether funnel plasmodesmata exist in sink organs other than roots, and thus represent a potential sink-specific mode for sugar unloading.
The role of the endodermis as a bidirectional barrier that controls the solutes’ access to the vascular cylinder, as well as prevents nutrients from leaking out, is also quite well established [86,87]. Recently, it has been shown that, as in the pericycle, two endodermal cell types co-exist in roots and each cell type responds differently to nutrients and hormones [88]. It is also suggested that these cells display different uptake and sensing potential [88]. These findings add an extra level of complexity to the role of the endodermis in the lateral transport of solutes and raise the long-standing question regarding the coordination of lateral transport of solutes between the endodermis and the pericycle cells.
4. Molecular Actors Involved in Lateral Transport of Solutes
In the context of lateral transport, determining whether a solute follows an active or a facilitated transport pathway depends mostly on its relative concentration in the different tissue layers that it crosses. Thus, if a solute (e.g., sucrose) is more concentrated in the phloem sap than it is in the xylem sap, it is logical to assume that its phloem-to-xylem transfer will follow the concentration gradient through plasmodesmata and/or the action of facilitators. Conversely, xylem-to-phloem transfer of the solute would need an active transport step in order to overcome the concentration gradient.
Sugar gradients have been shown at the tissue level in Vicia faba cotyledons; in the Ricinus communis hypocotyl; in the Populus tremula wood forming region [76,89,90]; and, very recently, at the microscopic level in different Barley (Hordeum vulgare) and Arabidopsis organs [91]. In contrast, almost no direct experimental proof of inorganic gradients exists [3,92,93]. Nonetheless, over the last 15 years, numerous plasma membrane solute transporters that could be involved in the intercellular transport of solutes have been characterized. These are expressed either in the leaf, in the root, or in the stem vascular and perivascular cells of dicotyledonous species (Table 2). Interestingly, both active and passive transporters (i.e., facilitators) acting mostly at the interface between vascular parenchyma cells and conducting cells have been identified (Table 2).
4.1. Lateral Transport of Inorganic Solutes from Xylem-To-Phloem
Inorganic solutes can be divided into anions (i.e., nitrite, nitrate, chloride, sulfate, and phosphate) and cations (i.e., ammonium, potassium, sodium, calcium, and magnesium). The lateral transport of inorganic nitrogen (i.e., nitrite, nitrate, and ammonium) has been the most studied (for review, [23]), and some of the molecular actors have been characterized (Table 2). Other inorganic ions are also most probably transported from xylem to phloem through an apoplasmic and/or coupled trans-cellular pathway. Nonetheless, movement of inorganic ions through plasmodesmata, for example, between the pericycle and protoxylem in a sink organ such as the onion root (Allium cepa) [81], cannot be excluded. In this section, we will more specifically address transporters expressed in vascular and perivascular cells that could be good candidates for the lateral transport of potassium, sodium, and phosphate, because a substantial cycling of these ions has been shown [16,133].
4.1.1. Potassium and Sodium
Potassium (K+) is the most abundant cation and is essential for many physiological processes [134]. Sodium (Na+), in contrast, has a dual role, being beneficial for plants at low concentrations, but becoming toxic as its concentration rises [134]. Sodium and potassium are often exchangeable, mainly because of their chemical similarities. The lateral transfer of Na and K, along with their long-distance transport, must thus be tightly coordinated to control the balance between the two. Both cations are taken up by the root system through numerous plasma membrane-localized transporters that are mostly expressed in the xylem parenchyma cells, such as the high-affinity potassium and sodium transporter 1 (HKT1), the cation/H+ exchanger (CHX14 and CHX21), the shaker-like outward channel (SKOR), and the Na+/H+ antiporter (SOS1) (Table 2 and references therein) [135]. Once loaded into the xylem sap through an active and/or a passive transport system [136], potassium and sodium are transported through the stem and unloaded at the target organs (e.g., the leaf) through the action of the high affinity potassium/sodium symporter (HKT1) (Table 2 and Figure 2). In addition, phloem/xylem sap analysis using radioactive isotopes of potassium and sodium showed that both cations are also partially transferred from the xylem to the phloem to be sent back to the root [25,31,137]. This process has been studied in particular in Ricinus communis [31]. In this species, more than 30% of the total Na+ taken up in the root is actually recycled back to the root after a xylem-to-phloem transfer. Additionally, half of the K+ loaded into the xylem sap is recycled back via the phloem to be incorporated into the roots. Even if this general flow model emphasizes the role of the lateral transfer between xylem and phloem for both cations in this species [31], the question of whether or not the unloading of potassium and sodium from the root phloem occurs through plasmodesmata needs to be further addressed.
Peuke [31] also noted that, in terms of proportion, the general flow model for K+ is quite similar to that of the N, suggesting a strong interaction between the two compounds. In tomato, the K+ concentration in the xylem sap has been shown to influence the rate of amino acid uptake, with a low K+ concentration stimulating amino acid uptake, and vice versa (Table 2 and references therein). More broadly, it has been shown that each time a substrate is taken up through a proton symport, K+ ions are released into the transpiration stream [138]. Regarding the translocation stream, analysis of mutants deficient in the expression of the AKT2/3 potassium channel localized in the phloem parenchyma cells in the Arabidopsis floral stem (Figure 2) showed that the retrieval mechanisms of sugars along the phloem tissue are also intimately linked to the transport of K+ ions via the action of AKT2/3 [139]. Subsequent studies identified that K+ ions moving in the phloem sap can be assimilated into an energy store that can be used to overcome local energy limitations generated by the action of the K+ and sugar proton-coupled transporters [140,141].
As AKT2/3 is a channel that is responsible for the facilitated diffusion of potassium, its mode of action implies the presence of a potassium gradient between the phloem companion cell/sieve element complex and the surrounding tissues that can be used locally to assure the retrieval and subsequent reloading of sugar by and from the transport phloem [141]. However, the exact nature of the transporters involved in such a process is still not known with certainty.
Sodium ions can also be loaded into the root xylem vessel via SOS1, a xylem parenchyma-localized antiporter, which mediates Na+/H+ exchanges [108]. Interestingly, SOS1 is also located in the xylem parenchyma cells of the floral stem and leaves [108], suggesting that this transporter could work both to load sodium into the xylem vessel and to retrieve it along the transpiration stream (Figure 2). Once in the transpiration stream, it has been suggested that sodium could be laterally transferred from xylem to phloem, likely via the action of the HKT1 transporter. This transfer has been suggested to be biologically relevant in order to prevent ion overaccumulation in the shoot of Arabidopsis or cotton in the context of high salinity [142,143]. However, Davenport et al. [144] later challenged the role of HKT1 in this process and, therefore, the very existence of sodium recirculation to the root through the phloem has been questioned and remains under debate [145]. One way to address this question would be to estimate the relative Na+ fluxes in the phloem and xylem sap. While in Ricinus communis, the recirculation of sodium has been estimated to be around 30%, similar experiments in monocotyledons suggest that lateral transfer of sodium would only account for 5–7% of the total sodium uptake [145]. Without questioning the existence of sodium recirculation, these discrepancies point out the need to carefully estimate phloem and xylem flow rates according to the plant species and environmental conditions.
4.1.2. Phosphate
Phosphorus (P) is a macronutrient essential for cellular processes such as energy production, redox reactions, photosynthesis, and phosphorylation/dephosphorylation reactions [146]. Phosphorus enters the plant root system in the form of inorganic P (Pi), such as PO43−, H2PO4−, or HPO42−, through an energized process involving H+/Pi co-transport in order to overcome the negative membrane potential [146]. Pi is then transported through the different root tissues through the action of, at least, the AtPHO1 and AtPHO1;H1 transporters, which are expressed in the endodermis and the pericycle, respectively, before being loaded into the xylem sap for root-to-shoot transport [102,103]. Additionally, under long-term Pi starvation, Pi is remobilized from the old leaves toward the sink tissues (e.g., the growing root or the seeds). This remobilization from shoot-to-root requires the action of the phloem-localized PHT1;5 transporter [104,147]. Although phosphorus was the first anion for which lateral transport was shown [25], it is still not known with certainty which transporters are involved in Pi recycling from the xylem to the phloem, although the antiporters PHO1 and PHT1;5 could be involved in such a process (Figure 2). In addition, it has been suggested that in rice, Pi might also be transported by members of the SULTR transporter family [148]. As most phosphate transporters are conserved between monocots and dicots, assessing the role of SULTR transporters in the lateral transfer of Pi may also be relevant. To improve our limited knowledge of lateral transport of Pi, one of the first steps will thus be to carefully identify the vascular cell types in which the plasmalemmal Pi transporters are expressed. Once this is achieved, genetic tools and Pi flow modelling could then be combined to help to better define the contribution of long-distance versus lateral Pi transport.
4.2. Lateral Transport of Sugars in the Vascular System
Experimental proof for the lateral transport of sugars has been provided for both the phloem-to-xylem and the xylem-to-phloem direction (Table 1). This transport is thought to primarily act in carbon transport and recirculatization via the phloem or the xylem. At a more local scale, the leakage-retrieval mechanism of sugars (recruiting both plasmodesmata or facilitators for leakage and energized transporters for retrieval) that occurs along the transport phloem (for review, [17]) can also be regarded as the lateral transport process.
4.2.1. Leakage-Retrieval Process along the Transport Phloem
The first evidence for a leakage-retrieval process along the transport phloem was obtained in experiments performed on the Phaseolus vulgaris stem [42,149,150,151,152]. This was demonstrated by the combined use of 11C-labelled substrate, electron microscopy techniques, and application of p-chloromercuribenzene sulphonic acid (PCMBS), which blocks active sugar loading into the phloem [42,149,150,151,152]. In Phaseolus vulgaris, passive leakage of photosynthates from the phloem sieve elements was shown to occur at a rate of 4% cm−1. The lost sugars were then assumed to be continuously reloaded by active transport [152]. In Arabidopsis, the sucrose/H+ symporter SUC2, located at the plasma membrane of the companion cells, has been shown to play a role in retrieving leaked sucrose along the transport phloem in addition to having a well-known role in loading sucrose into the phloem sieve elements [45] (Figure 2). Another part of the leaked sucrose is used by the lateral sinks, such as the cambium and possibly the xylem, for growth and tissue maintenance [17]. However, the exact proportion of sucrose involved in this leakage-retrieval cycle, as well as the molecular actors involved, remain difficult to determine. Nevertheless, observation of a partial complementation of the suc2 mutant line by expression of the SUC2 gene under control of the minor vein-specific galactinol synthase promoter from melon (CmGAS1) in this mutant suggests that the retrieval mechanism is not negligeable [153]. It is worth mentioning that this type of leakage-retrieval process most likely depends on the plant species, developmental stage, and growth conditions [154].
4.2.2. Phloem-To-Xylem and Xylem-To-Phloem Transport of Sugars in the Stem
In tree species, a proposed important role of the lateral transport of sugars is the prevention and/or repair of xylem embolism and/or cavitation. Transport of sugars between vascular cell types has been demonstrated during the xylem refilling process that occurs following an embolism [155]. It has been hypothesized that, during such a phenomenon, movement of water and sugars stored in the phloem towards the xylem vessels could be used to repair the embolism [155]. Additionally, as suggested for the walnut tree (Juglans regia), cell-to-cell transport of sugars between ray cells and xylem vessels, most likely mediated by a SUC2/SUT1 ortholog, could also account for the embolism repair [113] (Figure 2). Furthermore, it has been hypothetized that the starch stored in ray cell xylem parenchyma cells would depolymerize during xylem refilling and that the sugars produced during this phase are then loaded into cavitated conduits. In turn, the reduced level of starch in the xylem parenchyma cells would result in the cells becoming strong sinks, with the consequent unloading of sugars and water from the phloem directed to the refilling conduits [155]. The difference in sugar concentration between the two compartments/tissues would imply preferential use of plasmodesmata or a facilitated sugar transport system to sustain such a movement. Here, the sugar efflux transporters of the SWEET family could be good candidates [156].
In the Arabidopsis floral stem, the formation of xylem fibers and vessels and the subsequent extra-thickening of their secondary cell walls also appears to rely on lateral movement of sugars [121]. The existence of a sugar gradient between xylary parenchyma cells and xylem vessels has been suggested in a study of the double mutant sweet11sweet12 [121]. SWEET11 and SWEET12 are encoded for facilitators that transport sugars along the concentration gradient without any energized process [68], and are expressed in the phloem and xylem parenchyma cells of the floral stem [121] (Figure 2). The sweet11sweet12 double mutant was found to exhibit defects during development of the floral stem vascular system, as well as a modified xylem secondary cell wall composition [121]. Modification of the phloem cell wall composition has also been shown in the double mutant line using synchrotron-based Fourier-transformed infrared spectroscopy (FTIR) [157], suggesting that facilitated sugar transport between vascular parenchyma cells and developing conducting cells (i.e., xylem vessels, xylary fibers, and phloem sieve elements) is required to sustain normal cell wall formation. However, whether this movement takes place in the phloem-to-xylem direction or more locally between vascular parenchyma cells and conducting cells needs to be further addressed.
Moreover, it is not yet known if a similar transport mode exists in other plant species, such as ligneous species, in particular during wood formation, which requires a high amount of sugars for the synthesis of cell wall polysaccharides and lignin. The recent release of the aspen transcriptome profiling datasets, with a resolution of a few microns in the wood forming region, has enabled analysis of the expression pattern of SWEET genes during xylem formation [78,158]. Analyses have shown that several SWEET genes are also expressed in this region, supporting the possible participation of PtSWEET11/12 in the sugar exchange between rays and xylem vessels [78]. Interestingly, in aspen, it has been shown that once in the apoplasm of xylem tissue, sucrose can be delivered to the developing vessels through the action of the PttSUT3 (sucrose/H+ symporter) to support the secondary cell wall formation [77]. In addition, increased levels of hexoses and hexose phosphates, as well as UDP–glucose, in cells undergoing secondary cell wall formation have been identified across the wood forming region in poplar. This confirms the higher need for sugars in these cells and the existence of a sugar gradient along the xylem area undergoing secondary cell wall maturation and cell death [76]. A similar role for the SUT1 transporter has also been hypothesized in Solanaceous species (tomato, potato, and tobacco) because of its presence in both the phloem and the xylem parenchyma cells [119]. Further studies of members of the SUC/SUT and SWEET family will be needed to refine their possible role in the lateral transport of sugars (Figure 2).
4.2.3. Lateral Transport of Sugar in the Leaf
Even if there is no evidence for lateral transport of sugars between the vascular tissues in leaves, previous studies have identified interesting candidates that could be involved in the cell-to-cell transport of organic solutes between leaf bundle sheath and vascular tissues. Cui et al. (2014), using ChIP-chip experiments, identified several targets of the SHORT-ROOT (SHR) and/or SCARECROW-LIKE23 (SCL23) transcription factors, which specify the fate of bundle sheath cells in Arabidopsis leaves. Among them, some are involved in sugar transport, including the hexose transporters PMT5 and STP1 and the sucrose transporter SUC1. Interestingly, additional targets identified in this study are known to be involved in nitrogen (NFP6.3, NPF4.6, PROT3, UMAMIT1, and CAT6), potassium (KUP1 and KT12), magnesium (MGT9/MRS2-2 and MGT6/MRS2-4), and phosphate transport (PHT1;3) [159]. Furthermore, by characterizing the SHORT-ROOT/SCARECROW/SCARECROW-LIKE23 (SHR/SCR/SCL23) regulatory module, Cui et al. (2014) showed that in leaf major veins, SCR is preferentially expressed in BS cells facing the phloem, while SCL23 is expressed in BS cells facing the xylem. This suggests that, depending on their closest cell neighbors, BS cells could fulfil specific roles [159].
Additionally, cell-specific transcriptomic studies in Arabidopsis allowed the identification of differentially expressed genes in the leaf bundle sheath cells [61,62]. In particular, they identified genes coding for sugar transporters (i.e., AtSWEET10, AtSUC8), as well as amino acid/organic acid transporters (i.e., AtGLR3.6, AtALMT6).
Altogether, these works suggest the existence of a fine regulation of lateral solute transport mediated by an active and/or a passive transport system, and provide evidence that the lateral transport also integrates oriented flow of organic and inorganic solutes, either towards the adaxial or the abaxial side of the lamina.
5. State-Of-The-Art Biophysical Tools for Monitoring in Situ Lateral Transport of Solutes
A better understanding of lateral transport of solutes in vascular tissues requires correct description of the types of connections that exist between the vascular cells. This is particularly important for anatomically highly complex organs such as the stem. In the 1980s to 1990s, this issue was addressed using transmission electron microscopy, which allows plasmodesmata frequency maps between cells to be determined and, therefore, symplasmic continuity or discontinuity to be identified [160,161]. In addition, in rice leaf blades, combined use of fluorescent markers and confocal scanning laser microscopy has enabled identification of a symplasmic phloem-loading pathway between xylem parenchyma cells and phloem sieve elements [162].
In addition, Liesche et al. (2012) developed the three-dimensional photoactivation microscopy technique in order to quantify plasmodesmata-mediated cell wall permeability between different cell types with cage fluorescein as tracer, enabling them to quantify the real-time mobility between cells in angiosperms and gymnosperms [163,164]. As it is now possible to synthetize conjugated-molecules, such as fluorescent phytohormones [165,166], which are mobile between different cell types, and hydrophilic gold nanoparticles, which can be transported in the vascular tissues [167], the development of conjugated sugars might also be used as a more direct approach to trace sugars at the tissue or cell level. In line with these techniques, the fluorescent coumarine glucoside esculin, which mimics sucrose translocation [168], is a promising tool for exploring lateral transport. The use of infrared spectroscopy (FTIR) combined with state-of-the-art statistical modeling has also been proven effective for quantitatively imaging the sucrose content at the microscopic level in barley leaf, stem, and seeds, as well as in Arabidopsis hypocotyls [91]. This breakthrough technology thus represents a great opportunity to better characterize the lateral transport of metabolites in the future.
Lateral exchanges of sugars and amino acids might also be monitored at a higher spatial resolution using isotope tracers such as 15N and 13C/11C and cryo-secondary ion mass spectroscopy. This technique has already been successfully used to demonstrate lateral exchanges of magnesium, potassium, and calcium between vascular tissues in the french bean (Phaseolus vulgaris) stem [3]. This type of mineral ion movement has also been observed in planta with the real-time radioisotope imaging system (RRIS) [169]. This technique also allowed the authors to follow the temporal evolution of 14C-labelled metabolites over 24 h in the different organs at the whole plant level [169]. Finally, the use of new imaging techniques in plant sciences, such as position emission tomography (PET), which exploits positron-emitting radioactive metabolite analogues, will provide exciting opportunities to improve our understanding of solute flows within the vascular system [170]. State-of-the-art techniques such as these will likely be required to better understand the lateral movement of a wide range of compounds including sugars and inorganic ions.
6. Concluding Remarks and Future Prospects
Since the existence of lateral movement of inorganic solutes was first reported in the 1930s, significant progress has been made in identifying several of the molecular actors involved in organic and inorganic solute exchanges. It has been discovered that the lateral transport of sugars is controlled by coordinated symplasmic and apoplasmic pathways, which constitutes a new paradigm that still needs to be confirmed for the lateral transfer of other solutes. In this scheme, another important step will be to better understand the environmental factors regulating the relative contribution of both pathways, as well as to take into account that their contributions probably vary between organs within a plant species and between plant species. Thus, more knowledge needs to be acquired at the cell level (e.g., plasmodesmata density, localization of the solute transporters) before the relative contributions of both pathways to the lateral transport can be determined. In addition, the use of cell-specific and single-cell technology combined with next generation sequencing would help to identify additional transporter families that could be involved in lateral movement [60,171]. For instance, the use of bundle sheath (e.g., SHR and SCR23), xylem parenchyma (e.g., PRX47), or phloem parenchyma (e.g., GIGANTEA) specific promoters combined with the translatome tibosome affinity purification (TRAP) technology, developed by Mustroph et al. [60], could help to provide a better understanding of the specificity of these cell types, as well as to go further in the identification of transporters involved in solute loading and unloading between vascular and perivascular cells.
Overall, addressing these questions will help to better comprehend how the carbon, ion, and nitrogen allocation and the interaction between them are coordinated throughout the plant body. This knowledge could finally be used to achieve a complete model of the resource economy in plants and thus to improve crop yield.
Author Contributions
All authors contributed to the writing and editing of this review article.
Funding
Research in the laboratories of the authors was supported by grants from the Institut National de la Recherche Agronomique (INRA); from the CNRS; and from a French State grant (LabEx Saclay Plant Sciences-SPS, ANR-10-LABX-0040-SPS), managed by the French National Research Agency under an “Investments for the Future” program (ANR-11-IDEX-0003-02). EA is supported by a grant from SPS.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.R.; Helariutta, Y.; He, X.Q.; Fukuda, H.; Kang, J.; Brady, S.M.; et al. The Plant Vascular System: Evolution, Development and Functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef] [PubMed]
- Van Bel, A.J.E. Xylem-phloem exchange via the rays: The undervalued route of transport. J. Exp. Bot. 1990, 41, 631–644. [Google Scholar] [CrossRef]
- Metzner, R.; Schneider, H.U.; Breuer, U.; Thorpe, M.R.; Schurr, U.; Schroeder, W.H. Tracing Cationic Nutrients from Xylem into Stem Tissue of French Bean by Stable Isotope Tracers and Cryo-Secondary Ion Mass Spectrometry. Plant Physiol. 2010, 152, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Ogata, G. Radial Transport of Sodium and Chloride into Tomato Root Xylem. Plant Physiol. 1972, 50, 64–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfautsch, S.; Renard, J.; Tjoelker, M.G.; Salih, A. Phloem as capacitor: Radial transfer of water into xylem of tree stems occurs via symplastic transport in ray parenchyma. Plant Physiol. 2015, 167, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Sevilem, I.; Miyashima, S.; Helariutta, Y. Cell-to-cell communication via plasmodesmata in vascular plants. Cell Adhes. Migr. 2013, 7, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Colinas, J.; Wang, J.Y.; Mace, D.; Ohler, U.; Benfey, P.N. Transcriptional and posttranscriptional regulation of transcription factor expression in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2006, 103, 6055–6060. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Ahmad, M.; Rim, Y.; Lucas, W.J.; Kim, J.-Y. Evolutionary and molecular analysis of Dof transcription factors identified a conserved motif for intercellular protein trafficking. New Phytol. 2013, 198, 1250–1260. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, X.; Lee, J.-Y.; Lee, J.-Y. Cell-to-Cell Movement of Two Interacting AT-Hook Factors in Arabidopsis Root Vascular Tissue Patterning. Plant Cell 2013, 25, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benková, E.; Mähönen, A.P.; Helariutta, Y. A mutually inhibitory interaction between auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef]
- Thorpe, M.R.; Ferrieri, A.P.; Herth, M.M.; Ferrieri, R.A. 11C-imaging: Methyl jasmonate moves in both phloem and xylem, promotes transport of jasmonate, and of photoassimilate even after proton transport is decoupled. Planta 2007, 226, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Yazaki, K. Accumulation and membrane transport of plant alkaloids. Curr. Pharm. Biotechnol. 2007, 8, 244–252. [Google Scholar] [CrossRef]
- Yazaki, K.; Sugiyama, A.; Morita, M.; Shitan, N. Secondary transport as an efficient membrane transport mechanism for plant secondary metabolites. Phytochem. Rev. 2008, 7, 513–524. [Google Scholar] [CrossRef]
- Smith, R.A.; Schuetz, M.; Roach, M.; Mansfield, S.D.; Ellis, B.; Samuels, L. Neighboring Parenchyma Cells Contribute to Arabidopsis Xylem Lignification, while Lignification of Interfascicular Fibers Is Cell Autonomous. Plant Cell 2013, 25, 3988–3999. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.A.; Schuetz, M.; Karlen, S.D.; Bird, D.; Tokunaga, N.; Sato, Y.; Mansfield, S.D.; Ralph, J.; Samuels, A.L. Defining the Diverse Cell Populations Contributing to Lignification in Arabidopsis thaliana Stems. Plant Physiol. 2017, 174, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Pate, J.S. Exchange of Solutes between Phloem and Xylem and Circulation in the Whole Plant; Zimmermann, M.H., Milburn, J.A., Eds.; Transport; Springer: Berlin/Heidelberg, Germany, 1975; ISBN 978-3-642-66163-1. [Google Scholar]
- De Schepper, V.; De Swaef, T.; Bauweraerts, I.; Steppe, K. Phloem transport: A review of mechanisms and controls. J. Exp. Bot. 2013, 64, 4839–4850. [Google Scholar] [CrossRef]
- Zhang, L.; Tan, Q.; Lee, R.; Trethewy, A.; Lee, Y.H.; Tegeder, M. Altered xylem-phloem transfer of amino acids affects metabolism and leads to increased seed yield and oil content in Arabidopsis. Plant Cell Online 2010, 22, 3603–3620. [Google Scholar] [CrossRef]
- White, P.J. Long-distance Transport in the Xylem and Phloem. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 49–70. ISBN 9780123849052. [Google Scholar]
- González-Fontes, A.; Navarro-Gochicoa, M.T.; Ceacero, C.J.; Herrera-Rodríguez, M.B.; Camacho-Cristóbal, J.J.; Rexach, J. Understanding Calcium Transport and Signaling, and Its Use Efficiency in Vascular Plants; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128112946. [Google Scholar]
- Chen, Z.C.; Peng, W.T.; Li, J.; Liao, H. Functional dissection and transport mechanism of magnesium in plants. Semin. Cell Dev. Biol. 2018, 74, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Gigolashvili, T.; Kopriva, S. Transporters in plant sulfur metabolism. Front. Plant Sci. 2014, 5, 1–16. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Hammes, U.Z. The way out and in: Phloem loading and unloading of amino acids. Curr. Opin. Plant Biol. 2018, 43, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Stout, P.R.; Hoagland, D.R. Upward and lateral movement of salt in certain plants as indicated by radioactive isotopes of potassium, sodium, and phosphorus absorbed by roots. Am. J. Bot. 1939, 26, 320–324. [Google Scholar] [CrossRef]
- Peel, A.J. The movement of ions from the xylem solution into the sieve tubes of willow. J. Exp. Bot. 1963, 14, 438–447. [Google Scholar] [CrossRef]
- Hoad, G.V.; Peel, A.J. Studies on the movement of solutes between the sieve tubes and surrounding tissues: I. Interference between solutes and rate of translocation measurements. J. Exp. Bot. 1965, 16, 742–758. [Google Scholar] [CrossRef]
- Van Bel, A.J.E. Quantification of the xylem-to-phloem transfer of amino acids by use of inulin [14C]carboxylic acid as xylem transport marker. Plant Sci. Lett. 1984, 35, 81–85. [Google Scholar] [CrossRef]
- Pate, J.S.; Layzell, D.B.; McNeil, D.L. Modeling the transport and utilization of carbon and nitrogen in a nodulated legume. Plant Physiol. 1979, 63, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Layzell, D.B.; Larue, T.A. Modeling C and N transport to developing soybean fruits. Plant Physiol. 1982, 70, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Peuke, A.D. Correlations in concentrations, xylem and phloem flows, and partitioning of elements and ions in intact plants. A summary and statistical re-evaluation of modelling experiments in Ricinus communis. J. Exp. Bot. 2010, 61, 635–655. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, T.C.; Dickson, R.E.; Larson, P.R. Comparative Distribution and Metabolism of Xylem-Borne Amino Compounds and Sucrose in Shoots of Populus deltoides. Plant Physiol. 1985, 77, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, P.J.; Pate, J.S. Selectivity in xylem to phloem transfer of amino acids in fruiting shoots of white lupin (Lupinus albus L.). Planta 1975. [Google Scholar] [CrossRef]
- Pate, J.S.; Sharkey, P.J.; Lewis, O.A.M. Xylem to phloem transfer of solutes in fruiting shoots of legumes, studied by a phloem bleeding technique. Planta 1975, 122, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Hardy, P.J.; Possingham, J.V. Studies on translocation of metabolites in the xylem of grapevine shoots. J. Exp. Bot. 1969, 20, 325–335. [Google Scholar] [CrossRef]
- Ford, J.; Peel, A.J. The movement of sugars into the sieve elements of bark strips of willow. J. Exp. Bot. 1967, 18, 607–619. [Google Scholar] [CrossRef]
- Gardner, D.C.J.; Peel, A.J. Metabolism and transport of 14C-labelled glutamic and aspartic acids in the phloem of willow. Phytochemistry 1971, 10, 2385–2387. [Google Scholar] [CrossRef]
- Swanson, C.A.; Shishiny, E.D.H. Translocation of sugars in the concord grape. Plant Physiol. 1958, 33–37. [Google Scholar] [CrossRef]
- Nelson, C.D.; Gorham, P.R. Translocation of radioactive sugars in the stems of soybean seedlings. Can. J. Bot. 1957, 35, 703–713. [Google Scholar] [CrossRef]
- Peel, A.J. Demonstration of Solute Movement from the Extracambial Tissues into the Xylem Stream in Willow. J. Exp. Bot. 1967, 18, 600–606. [Google Scholar] [CrossRef]
- Biddulph, S.F.; Biddulph, O.; Cory, R. Visual indications of S35 and P32 translocation in the phloem. Am. J. Bot. 1956, 45, 648–652. [Google Scholar] [CrossRef]
- Minchin, P.E.H.; Thorpe, M.R. Measurement of unloading and reloading of photo-assimilate within the stem of bean. J. Exp. Bot. 1987, 38, 211–220. [Google Scholar] [CrossRef]
- Mason, T.; Maskell, E. Studies on the transport of carbohydrates in the cotton plant: II. The factors determining the rate and the direction of movement of sugars. Ann. Bot. 1928, 42, 571–636. [Google Scholar] [CrossRef]
- Zimmermann, M.H. Translocation of organic substances in trees. III. The removal of sugars from the sieve tubes in the white ash (Fraxinus americana L.). Plant Physiol. 1957, 33, 213–217. [Google Scholar] [CrossRef]
- Gould, N.; Thorpe, M.R.; Pritchard, J.; Christeller, J.T.; Williams, L.E.; Roeb, G.; Schurr, U.; Minchin, P.E.H. AtSUC2 has a role for sucrose retrieval along the phloem pathway: Evidence from carbon-11 tracer studies. Plant Sci. Int. J. Exp. Plant Biol. 2012, 188, 97. [Google Scholar] [CrossRef] [PubMed]
- Geßler, A.; Weber, P.; Schneider, S.; Rennenberg, H. Bidirectional exchange of amino compounds between phloem and xylem during long-distance transport in Norway spruce trees (Picea abies [L.] Karst). J. Exp. Bot. 2003, 54, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Wolf, O.; Jeschke, W.D. Modeling of Sodium and Potassium Flows via Phloem and Xylem in the Shoot of Salt-stressed Barley. J. Plant Physiol. 1987, 128, 371–386. [Google Scholar] [CrossRef]
- Jeschke, W.D.; Pate, J.S. Cation and chloride partitioning through xylem and phloem within the whole plant of Ricinus communis L. under conditions of salt stress. J. Exp. Bot. 1991, 42, 1105–1116. [Google Scholar] [CrossRef]
- Dinant, S.; Lemoine, R. The phloem pathway: New issues and old debates. C. R. Biol. 2010, 333, 307–319. [Google Scholar] [CrossRef]
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef]
- Lacombe, B.; Achard, P. Long-distance transport of phytohormones through the plant vascular system. Curr. Opin. Plant Biol. 2016, 34, 1–8. [Google Scholar] [CrossRef]
- Spiegelman, Z.; Golan, G.; Wolf, S. Don’t kill the messenger: Long-distance trafficking of mRNA molecules. Plant Sci. 2013, 213, 1–8. [Google Scholar] [CrossRef]
- Kim, H.K.; Park, J.; Hwang, I. Investigating water transport through the xylem network in vascular plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef] [Green Version]
- Leegood, R.C. Roles of the bundle sheath cells in leaves of C3 plants. J. Exp. Bot. 2008, 59, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.T. Walking the C 4 pathway: Past, present, and future. J. Exp. Bot. 2017, 68, 4057–4066. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Hao, Y.; Kong, D. SCARECROW Has a SHORT-ROOT-Independent Role in Modulating the Sugar Response. Plant Physiol. 2012, 158, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Sade, N.; Shatil-Cohen, A.; Attia, Z.; Maurel, C.; Boursiac, Y.; Kelly, G.; Granot, D.; Yaaran, A.; Lerner, S.; Moshelion, M. The Role of Plasma Membrane Aquaporins in Regulating the Bundle Sheath-Mesophyll Continuum and Leaf Hydraulics. Plant Physiol. 2014, 166, 1609–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shatil-Cohen, A.; Attia, Z.; Moshelion, M. Bundle-sheath cell regulation of xylem-mesophyll water transport via aquaporins under drought stress: A target of xylem-borne ABA? Plant J. 2011, 67, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Haritatos, E.; Medville, R.; Turgeon, R. Minor vein structure and sugar transport in Arabidopsis thaliana. Planta 2000, 211, 105–111. [Google Scholar] [CrossRef]
- Mustroph, A.; Zanetti, M.E.; Jang, C.J.H.; Holtan, H.E.; Repetti, P.P.; Galbraith, D.W.; Girke, T.; Bailey-Serres, J. Profiling translatomes of discrete cell populations resolves altered cellular priorities during hypoxia in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18843–18848. [Google Scholar] [CrossRef] [Green Version]
- Wigoda, N.; Pasmanik-Chor, M.; Yang, T.; Yu, L.; Moshelion, M.; Moran, N. Differential gene expression and transport functionality in the bundle sheath versus mesophyll—A potential role in leaf mineral homeostasis. J. Exp. Bot. 2017, 68, 3179–3190. [Google Scholar] [CrossRef] [PubMed]
- Aubry, S.; Smith-Unna, R.D.; Boursnell, C.M.; Kopriva, S.; Hibberd, J.M. Transcript residency on ribosomes reveals a key role for the Arabidopsis thaliana bundle sheath in sulfur and glucosinolate metabolism. Plant J. 2014, 78, 659–673. [Google Scholar] [CrossRef]
- Gunning, B.E.S.; Pate, J.S. “Transfer cells” Plant cells with wall ingrowths, specialized in relation to short distance transport of solutes-Their occurrence, structure, and development. Protoplasma 1969, 68, 107–133. [Google Scholar] [CrossRef]
- Wu, Y.; Hou, J.; Yu, F.; Nguyen, S.T.T.; McCurdy, D.W. Transcript Profiling Identifies NAC-Domain Genes Involved in Regulating Wall Ingrowth Deposition in Phloem Parenchyma Transfer Cells of Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W.I. Associations between the acclimation of phloem-cell wall ingrowths in minor veins and maximal photosynthesis rate. Front. Plant Sci. 2014, 15, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Arun Chinnappa, K.S.; Nguyen, T.T.S.; Hou, J.; Wu, Y.; McCurdy, D.W. Phloem parenchyma transfer cells in Arabidopsis—An experimental system to identify transcriptional regulators of wall ingrowth formation. Front. Plant Sci. 2013, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.T.T.; McCurdy, D.W. Wall ingrowth deposition in phloem parenchyma transfer cells in Arabidopsis: Heteroblastic variations and a potential role in pathogen defence. Plant Signal. Behav. 2017, 12, e1338226. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Qu, X.-Q.; Hou, B.-H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef]
- Cayla, T.; Le Hir, R.; Dinant, S. Live-cell imaging of fluorescently-tagged phloem proteins with confocal microscopy. In Phloem: Methods and Protocols; Liesche, J., Ed.; Springer Nature: Basel, Swizterland, 2019; in press. [Google Scholar]
- Strabala, T.J.; MacMillan, C.P. The Arabidopsis wood model-the case for the inflorescence stem. Plant Sci. 2013, 210, 193–205. [Google Scholar] [CrossRef]
- Chaffey, N.; Cholewa, E.; Regan, S.; Sundberg, B. Secondary xylem development in Arabidopsis: A model for wood formation. Physiol. Plant. 2002, 114, 594–600. [Google Scholar] [CrossRef]
- Schuetz, M.; Smith, R.; Ellis, B. Xylem tissue specification, patterning, and differentiation mechanisms. J. Exp. Bot. 2012, 64, 11–31. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.-C.; Liu, C.-J. ATP-binding cassette-like transporters are involved in the transport of lignin precursors across plasma and vacuolar membranes. Proc. Natl. Acad. Sci. USA 2010. [Google Scholar] [CrossRef]
- Alejandro, S.; Lee, Y.; Tohge, T.; Sudre, D.; Osorio, S.; Park, J.; Bovet, L.; Lee, Y.; Geldner, N.; Fernie, A.R.; et al. AtABCG29 is a monolignol transporter involved in lignin biosynthesis. Curr. Biol. 2012, 22, 1207–1212. [Google Scholar] [CrossRef]
- Mazur, E.; Kurczynska, E.U. Rays, intrusive growth, and storied cambium in the inflorescence stems of Arabidopsis thaliana (L.) Heynh. Protoplasma 2012, 249, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Roach, M.; Arrivault, S.; Mahboubi, A.; Krohn, N.; Sulpice, R.; Stitt, M.; Niittylä, T. Spatially resolved metabolic analysis reveals a central role for transcriptional control in carbon allocation to wood. J. Exp. Bot. 2017, 68, 3529–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahboubi, A.; Ratke, C.; Gorzsás, A.; Kumar, M.; Mellerowicz, E.J.; Niittyla, T. Aspen SUCROSE TRANSPORTER3 Allocates Carbon into Wood Fibers. Plant Physiol. 2013, 163, 1729–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahboubi, A.; Niittylä, T. Sucrose transport and carbon fluxes during wood formation. Physiol. Plant. 2018. [Google Scholar] [CrossRef] [PubMed]
- Van Norman, J.M. Asymmetry and cell polarity in root development. Dev. Biol. 2016, 419, 165–174. [Google Scholar] [CrossRef]
- Nieminen, K.; Blomster, T.; Helariutta, Y.; Mähönen, A.P. Vascular Cambium Development. Arab B 2015, 13, e0177. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Peterson, C.A. Frequencies of plasmodesmata in Allium cepa L. roots: Implications for solute transport pathways. J. Exp. Bot. 2001, 52, 1051–1061. [Google Scholar] [CrossRef]
- Beeckman, T.; De Smet, I. Pericycle. Curr. Biol. 2014, 24, 378–379. [Google Scholar] [CrossRef]
- Parizot, B.; Roberts, I.; Raes, J.; Beeckman, T.; de Smet, I. In silico analyses of pericycle cell populations reinforce their relation with associated vasculature in Arabidopsis. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1479–1488. [Google Scholar] [CrossRef] [Green Version]
- Takano, J.; Noguchi, K.; Yasumori, M.; Kobayashi, M.; Gajdos, Z.; Miwa, K.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. Arabidopsis boron transporter for xylem loading. Nature 2002, 420, 337–340. [Google Scholar] [CrossRef]
- Ross-Elliott, T.J.; Jensen, K.H.; Haaning, K.S.; Wager, B.M.; Knoblauch, J.; Howell, A.H.; Mullendore, D.L.; Monteith, A.G.; Paultre, D.; Yan, D.; et al. Phloem unloading in Arabidopsis roots is convective and regulated by the phloem-pole pericycle. Elife 2017, 6, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Doblas, V.G.; Geldner, N.; Barberon, M. The endodermis, a tightly controlled barrier for nutrients. Curr. Opin. Plant Biol. 2017, 39, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M. The endodermis as a checkpoint for nutrients. New Phytol. 2017, 213, 1604–1610. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.G.; Naseer, S.; Ursache, R.; Wybouw, B.; Smet, W.; De Rybel, B.; Vermeer, J.E.M.; Geldner, N. Diffusible repression of cytokinin signalling produces endodermal symmetry and passage cells. Nature 2018, 555, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Borisjuk, L.; Walenta, S.; Rolletschek, H.; Mueller-Klieser, W.; Wobus, U.; Weber, H. Spatial analysis of plant metabolism: Sucrose imaging within Vicia faba cotyledons reveals specific developmental patterns. Plant J. 2002, 29, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verscht, J.; Tomos, D.; Komor, E. Sugar concentrations along and across the Ricinus communis L. hypocotyl measured by single cell sampling analysis. Planta 2006, 224, 1303–1314. [Google Scholar] [CrossRef]
- Gündel, A.; Rolletschek, H.; Wagner, S.; Muszynska, A.; Borisjuk, L. Micro imaging displays the sucrose landscape within and along its allocation pathways. Plant Physiol. 2018, 49. [Google Scholar] [CrossRef]
- Metzner, R.; Thorpe, M.R.; Breuer, U.; Blümler, P.; Schurr, U.; Schneider, H.U.; Schroeder, W.H. Contrasting dynamics of water and mineral nutrients in stems shown by stable isotope tracers and cryo-SIMS. Plant Cell Environ. 2010, 33, 1393–1407. [Google Scholar] [CrossRef]
- Metzner, R.; Schneider, H.U.; Breuer, U.; Schroeder, W.H. Imaging Nutrient Distributions in Plant Tissue Using Time-of-Flight Secondary Ion Mass Spectrometry and Scanning Electron Microscopy. Plant Physiol. 2008, 147, 1774–1787. [Google Scholar] [CrossRef] [Green Version]
- Deeken, R.; Sanders, C.; Ache, P.; Hedrich, R. Developmental and light-dependent regulation of a phloem-localised K+ channel of Arabidopsis thaliana. Plant J. 2000, 23, 285–290. [Google Scholar] [CrossRef]
- Marten, I.; Hoth, S.; Deeken, R.; Ache, P.; Ketchum, K.A.; Hoshi, T.; Hedrich, R. AKT3, a phloem-localized K+ channel, is blocked by protons. Proc. Natl. Acad. Sci. USA 1999, 96, 7581–7586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmenero-Flores, J.M.; Martínez, G.; Gamba, G.; Vázquez, N.; Iglesias, D.J.; Brumós, J.; Talón, M. Identification and functional characterization of cation-chloride cotransporters in plants. Plant J. 2007, 50, 278–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, P.; Motes, C.M.; Park, S.; Hirschi, K.D. CHX14 is a plasma membrane K-efflux transporter that regulates K+redistribution in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 2223–2238. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.; Evans, A.R.; Newbury, H.J.; Pritchard, J. Functional analysis of CHX21: A putative sodium transporter in Arabidopsis. J. Exp. Bot. 2006, 57, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Sunarpi; Horie, T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.; Leung, H.; Hattori, K.; et al. Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na+ unloading from xylem vessels to xylem parenchyma cells. Plant J. 2005, 44, 928–938. [Google Scholar] [CrossRef] [Green Version]
- An, D.; Chen, J.G.; Gao, Y.Q.; Li, X.; Chao, Z.F.; Chen, Z.R.; Li, Q.Q.; Han, M.L.; Wang, Y.L.; Wang, Y.F.; et al. AtHKT1 drives adaptation of Arabidopsis thaliana to salinity by reducing floral sodium content. PLoS Genet. 2017, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-H.; Kuo, H.-F.; Canivenc, G.; Lin, C.-S.; Lepetit, M.; Hsu, P.-K.; Tillard, P.; Lin, H.-L.; Wang, Y.-Y.; Tsai, C.-B.; et al. Mutation of the Arabidopsis NRT1.5 Nitrate Transporter Causes Defective Root-to-Shoot Nitrate Transport. Plant Cell Online 2008, 20, 2514–2528. [Google Scholar] [CrossRef]
- Hamburger, D.; Rezzonico, E.; Somerville, C.; Poirier, Y. Identification and Characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. Society 2002, 14, 889–902. [Google Scholar] [CrossRef]
- Stefanovic, A.; Ribot, C.; Rouached, H.; Wang, Y.; Chong, J.; Belbahri, L.; Delessert, S.; Poirier, Y. Members of the PHO1 gene family show limited functional redundancy in phosphate transfer to the shoot, and are regulated by phosphate deficiency via distinct pathways. Plant J. 2007, 50, 982–994. [Google Scholar] [CrossRef] [Green Version]
- Mudge, S.R.; Rae, A.L.; Diatloff, E.; Smith, F.W. Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis. Plant J. 2002, 31, 341–353. [Google Scholar] [CrossRef]
- Inoue, Y.; Kobae, Y.; Omoto, E.; Tanaka, A.; Banba, M.; Takai, S.; Tamura, Y.; Hirose, A.; Komatsu, K.; Otagaki, S.; et al. The soybean mycorrhiza-inducible phosphate transporter gene, GmPT7, also shows localized expression at the tips of vein endings of senescent leaves. Plant Cell Physiol. 2014, 55, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Gaymard, F.; Pilot, G.; Lacombe, B.; Bouchez, D.; Bruneau, D.; Boucherez, J.; Michaux-Ferrière, N.; Thibaud, J.B.; Sentenac, H. Identification and disruption of a plant shaker-like outward channel involved in K+release into the xylem sap. Cell 1998, 94, 647–655. [Google Scholar] [CrossRef]
- Cubero-Font, P.; Maierhofer, T.; Jaslan, J.; Rosales, M.A.; Espartero, J.; Díaz-Rueda, P.; Müller, H.M.; Hürter, A.L.; AL-Rasheid, K.A.S.; Marten, I.; et al. Silent S-Type Anion Channel Subunit SLAH1 Gates SLAH3 Open for Chloride Root-to-Shoot Translocation. Curr. Biol. 2016, 26, 2213–2220. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Quintero, F.J.; Pardo, J.M.; Zhu, J.K. The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+transport in plants. Plant Cell 2002, 14, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Hayashi, N.; Yamaya, T.; Takahashi, H. Root-to-shoot transport of sulfate in Arabidopsis. Evidence for the role of SULTR3;5 as a component of low-affinity sulfate transport system in the root vasculature. Plant Physiol. 2004, 136, 4198–4204. [Google Scholar] [CrossRef]
- Dürr, J.; Bücking, H.; Mult, S.; Wildhagen, H.; Palme, K.; Rennenberg, H.; Ditengou, F.; Herschbach, C. Seasonal and cell type specific expression of sulfate transporters in the phloem of Populus reveals tree specific characteristics for SO42-storage and mobilization. Plant Mol. Biol. 2010, 72, 499–517. [Google Scholar] [CrossRef]
- Shakya, R.; Sturm, A. Characterization of source- and sink-specific sucrose/H+ symporters from carrot. Plant-Physiology 1998, 118, 1473–1480. [Google Scholar] [CrossRef]
- Decourteix, M.; Alves, G.; Brunel, N.; Ameglio, T.; Guilliot, A.; Lemoine, R.; Petel, G.; Sakr, S. JrSUT1, a putative xylem sucrose transporter, could mediate sucrose influx into xylem parenchyma cells and be up-regulated by freeze–thaw cycles over the autumn–winter period in walnut tree (Juglans regia L.). Plant Cell Environ. 2006, 29, 36–47. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Tay, K.H.C.; Tsai, C.-J.; Harding, S.A. The sucrose transporter family in Populus: The importance of a tonoplast PtaSUT4 to biomass and carbon partitioning. Plant J. 2011, 65, 757–770. [Google Scholar] [CrossRef]
- Truernit, E.; Sauer, N. The promoter of the Arabidopsis thaliana SUC2 sucrose-H+ symporter gene directs expression of B-glucuronidase to the phloem: Evidence for phloem loading and unloading by SUC. Planta 1995, 196, 564–570. [Google Scholar] [CrossRef]
- Stadler, R.; Sauer, N. The Arabidopsis thaliana AtSUC2 gene is specifically expressed in companion cells. Bot. Acta 1996, 109, 299–306. [Google Scholar] [CrossRef]
- Meyer, S.; Melzer, M.; Truernit, E.; Hummer, C.; Besenbeck, R.; Stadler, R.; Sauer, N. AtSUC3, a gene encoding a new Arabidopsis sucrose transporter, is expressed in cells adjacent to the vascular tissue and in a carpel cell layer. Plant J. 2000, 24, 869–882. [Google Scholar] [CrossRef]
- Meyer, S.; Lauterbach, C.; Niedermeier, M.; Barth, I.; Sjolund, R.D.; Sauer, N. Wounding enhances expression of AtSUC3, a sucrose transporter from Arabidopsis sieve elements and sink tissues. Plant Physiol. 2004, 134, 684–693. [Google Scholar] [CrossRef]
- Schmitt, B.; Stadler, R.; Sauer, N. Immunolocalization of solanaceous SUT1 proteins in companion cells and xylem parenchyma: New perspectives for phloem loading and transport. Plant Physiol. 2008, 148, 187–199. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Yang, C.; Tian, Z.; Li, J. AtSWEET4, a hexose facilitator, mediates sugar transport to axial sinks and affects plant development. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Le Hir, R.; Spinner, L.; Klemens, P.A.W.; Chakraborti, D.; De Marco, F.; Vilaine, F.; Wolff, N.; Lemoine, R.; Porcheron, B.; Géry, C.; et al. Disruption of the Sugar Transporters AtSWEET11 and AtSWEET12 Affects Vascular Development and Freezing Tolerance in Arabidopsis. Mol. Plant 2015, 8, 1687–1990. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Zhang, H.; Zhang, Q.; Zhai, H.; Liu, Q.; He, S. The Plasma Membrane-Localized Sucrose Transporter IbSWEET10 Contributes to the Resistance of Sweet Potato to Fusarium oxysporum. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Okumoto, S.; Schmidt, R.; Tegeder, M.; Fischer, W.N.; Rentsch, D.; Frommer, W.B.; Koch, W. High affinity amino acid transporters specifically expressed in xylem parenchyma and developing seeds of Arabidopsis. J. Biol. Chem. 2002, 277, 45338–45346. [Google Scholar] [CrossRef]
- Mayer, M.; Ludewig, U. Role of AMT1;1 in NH4+ acquisition in Arabidopsis thaliana. Plant Biol. 2006, 8, 522–528. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; von Wirén, N. A Critical Role of AMT2;1 in Root-To-Shoot Translocation of Ammonium in Arabidopsis. Mol. Plant 2017, 10, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Grallath, S. The AtProT Family. Compatible Solute Transporters with Similar Substrate Specificity But Differential Expression Patterns. Plant Physiol. 2005, 137, 117–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taochy, C.; Gaillard, I.; Ipotesi, E.; Oomen, R.; Leonhardt, N.; Zimmermann, S.; Peltier, J.B.; Szponarski, W.; Simonneau, T.; Sentenac, H.; et al. The Arabidopsis root stele transporter NPF2.3 contributes to nitrate translocation to shoots under salt stress. Plant J. 2015, 83, 466–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.C.; Lin, C.S.; Hsia, A.P.; Su, R.C.; Lin, H.L.; Tsay, Y.F. Mutation of a nitrate transporter, AtNRT1:4, results in a reduced petiole nitrate content and altered leaf development. Plant Cell Physiol. 2004, 45, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.A.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A.A. Nitrate Transport, Sensing, and Responses in Plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef]
- Li, J.-Y.; Fu, Y.-L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.-Z.; Zhang, Y.; Li, H.-M.; Huang, J.; et al. The Arabidopsis Nitrate Transporter NRT1.8 Functions in Nitrate Removal from the Xylem Sap and Mediates Cadmium Tolerance. Plant Cell Online 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-Y.; Tsay, Y.-F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef]
- Hsu, P.-K.; Tsay, Y.-F. Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef]
- Zhong, W.J.; Kaiser, W.; Kohler, J.; Bauer-Ruckdeschel, H.; Komor, E. Phloem loading of inorganic cations and anions by the seedling of Ricinus communis L. J. Plant Physiol. 1998, 152, 328–335. [Google Scholar] [CrossRef]
- Adams, E.; Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Zeppel, M.J.B.; Anderegg, W.R.L.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef] [Green Version]
- Amtmann, A.; Armengaud, P.; Volkov, V. Potassium Nutrition and Salt Stress; Wiley Online Library: Hoboken, NJ, USA, 2018; Volume 15, ISBN 9781119312994. [Google Scholar]
- Jeschke, W.D.; Pate, J.S.; Atkins, C.A. Partitioning of K+, Na+, Mg++, and Ca++ through Xylem and Phloem to Component Organs of Nodulated White Lupin under Mild Salinity. J. Plant Physiol. 1987, 128, 77–93. [Google Scholar] [CrossRef]
- Van Bel, A.J.E.; Van Erven, A.J. Potassium co-transport and antiport during the uptake of sucrose and glumatic acid from the xylem cells. Plant Sci. Lett. 1979, 15, 285–291. [Google Scholar] [CrossRef]
- Deeken, R.; Geiger, D.; Fromm, J.; Koroleva, O.; Ache, P.; Langenfeld-Heyser, R.; Sauer, N.; May, S.T.; Hedrich, R. Loss of the AKT2/3 potassium channel affects sugar loading into the phloem of Arabidopsis. Planta 2002, 216, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Gajdanowicz, P.; Michard, E.; Sandmann, M.; Rocha, M.; Corrêa, L.G.G.; Ramirez-Aguilar, S.J.; Gomez-Porras, J.L.; Gonzalez, W.; Thibaud, J.-B.; van Dongen, J.T.; et al. Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc. Natl. Acad. Sci. USA 2011, 108, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, I.; Gomez-Porras, J.L.; Riedelsberger, J. The potassium battery: A mobile energy source for transport processes in plant vascular tissues. New Phytol. 2017, 216, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Berthomieu, P.; Conéjéro, G.; Nublat, A.; Brackenbury, W.J.; Lambert, C.; Savio, C.; Uozumi, N.; Oiki, S.; Yamada, K.; Cellier, F.; et al. Functional analysis of AtHKT1 in Arabidopsis shows that Na+recirculation by the phloem is crucial for salt tolerance. EMBO J. 2003, 22, 2004–2014. [Google Scholar] [CrossRef]
- Kong, X.; Luo, Z.; Dong, H.; Eneji, A.E.; Li, W. Effects of non-uniform root zone salinity on water use, Na+recirculation, and Na+and H+flux in cotton. J. Exp. Bot. 2012, 63, 2105–2116. [Google Scholar] [CrossRef]
- Davenport, R.J.; Muñoz-Mayor, A.; Jha, D.; Essah, P.A.; Rus, A.; Tester, M. The Na+ transporter AtHKT1;1 controls retrieval of Na + from the xylem in Arabidopsis. Plant Cell Environ. 2007, 30, 497–507. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Ahmad, I.; Patishtan, J. Regulation of Na+ fluxes in plants. Front. Plant Sci. 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Srivastava, S.; Upadhyay, M.; Srivastava, A.; Abdelrahman, M.; Suprasanna, P.; Tran, L.-S. Cellular and Subcellular Phosphate Transport Machinery in Plants. Int. J. Mol. Sci. 2018, 19, 1914. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, V.K.; Jain, A.; Poling, M.D.; Lewis, A.J.; Raghothama, K.G.; Smith, A.P. Arabidopsis Pht1;5 Mobilizes Phosphate between Source and Sink Organs and Influences the Interaction between Phosphate Homeostasis and Ethylene Signaling. Plant Physiol. 2011, 156, 1149–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Deng, M.; Xu, J.; Zhu, X.; Mao, C. Molecular mechanism of phosphate signaling in plants. Semin. Cell Dev. Biol. 2017, 74, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.M.; Patrick, J.W.; Offler, C.E. The cellular pathway of radial transfer of photosynthates in stems of Phaseolus vulgaris L.: Effects of cellular plasmolysis and p-chloromercuribenzene sulphonic acid. Ann. Bot. 1987, 59, 635–642. [Google Scholar] [CrossRef]
- Hayes, P.M.; Offler, C.E.; Patrick, J.W. Cellular structures, plasma membrane surface areas and plasmodesmatal frequencies of the stem of Phaseolus vulgaris L. in relation to radial photosynthate transfer. Ann. Bot. 1985, 56, 125–138. [Google Scholar] [CrossRef]
- Minchin, P.E.H.; Thorpe, M.R. Apoplastic phloem unloading in the stem of bean. J. Exp. Bot. 1984, 35, 538–550. [Google Scholar] [CrossRef]
- Minchin, P.E.H.; Ryan, K.G.; Thorpe, M.R. Further evidence of apoplastic unloading into the stem of bean: Identification of the phloem buffering pool. J. Exp. Bot. 1984, 35, 1744–1753. [Google Scholar] [CrossRef]
- Srivastava, A.C.; Ganesan, S.; Ismail, I.O.; Ayre, B.G. Functional characterization of the Arabidopsis AtSUC2 sucrose/H+ symporter by tissue-specific complementation reveals an essential role in phloem loading but not in long-distance transport. Plant Physiol. 2008, 148, 200–211. [Google Scholar] [CrossRef]
- Hafke, J.B.; van Amerongen, J.-K.; Kelling, F.; Furch, A.C.U.; Gaupels, F.; van Bel, A.J.E. Thermodynamic battle for photosynthate acquisition between sieve tubes and adjoining parenchyma in transport phloem. Plant Physiol. 2005, 138, 1527–1537. [Google Scholar] [CrossRef]
- Nardini, A.; Lo Gullo, M.A.; Salleo, S. Refilling embolized xylem conduits: Is it a matter of phloem unloading? Plant Sci. 2011, 180, 604–611. [Google Scholar] [CrossRef]
- Hacke, U.G.; Laur, J. Xylem refilling—A question of sugar transporters and pH? Plant Cell Environ. 2016, 39, 2347–2349. [Google Scholar] [CrossRef] [PubMed]
- Dinant, S.; Wolff, N.; De Marco, F.; Vilaine, F.; Gissot, L.; Aubry, E.; Sandt, C.; Bellini, C.; Le Hir, R. Synchrotron FTIR and Raman spectroscopy provide unique spectral fingerprints for Arabidopsis floral stem vascular tissues. BioRXiv 2018. [Google Scholar] [CrossRef] [PubMed]
- Sundell, D.; Street, N.R.; Kumar, M.; Mellerowicz, E.J.; Kucukoglu, M.; Johnsson, C.; Kumar, V.; Mannapperuma, C.; Delhomme, N.; Nilsson, O.; et al. AspWood: High-Spatial-Resolution Transcriptome Profiles Reveal Uncharacterized Modularity of Wood Formation in Populus tremula. Plant Cell 2017, 29, 1585–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Kong, D.; Liu, X.; Hao, Y. SCARECROW, SCR-LIKE 23 and SHORT-ROOT control bundle sheath cell fate and function in Arabidopsis thaliana. Plant J. 2014, 78, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Botha, C.E.J.; van Bel, A.J.E. Quantification of symplastic continuity as visualised by plasmodesmograms: Diagnostic value for phloem-loading pathways. Planta 1992, 187, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.G. Ultrastructure, plasmodesmatal frequency, and solute concentration in green areas of variegated Coleus blumei Benth. leaves. Planta 1986, 169, 141–152. [Google Scholar] [CrossRef]
- Botha, C.E.J.; Aoki, N.; Scofield, G.N.; Liu, L.; Furbank, R.T.; White, R.G. A xylem sap retrieval pathway in rice leaf blades: Evidence of a role for endocytosis? J. Exp. Bot. 2008, 59, 2945–2954. [Google Scholar] [CrossRef]
- Liesche, J.; Schulz, A. Quantification of plant cell coupling with three-dimensional photoactivation microscopy. J. Microsc. 2012, 247, 2–9. [Google Scholar] [CrossRef]
- Liesche, J.; Schulz, A. In vivo quantification of cell coupling in plants with different phloem-loading strategies. Plant Physiol. 2012, 159, 355–365. [Google Scholar] [CrossRef]
- Shani, E.; Weinstain, R.; Zhang, Y.; Castillejo, C.; Kaiserli, E.; Chory, J.; Tsien, R.Y.; Estelle, M. Gibberellins accumulate in the elongating endodermal cells of Arabidopsis root. Proc. Natl. Acad. Sci. USA 2013. [Google Scholar] [CrossRef]
- Irani, N.G.; Di Rubbo, S.; Mylle, E.; Van den Begin, J.; Schneider-Pizoń, J.; Hniliková, J.; Šíša, M.; Buyst, D.; Vilarrasa-Blasi, J.; Szatmári, A.-M.; et al. Fluorescent castasterone reveals BRI1 signaling from the plasma membrane. Nat. Chem. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, H.K.; Ryu, J.; Ahn, S.; Lee, S.J.; Hwang, I. Functional water flow pathways and hydraulic regulation in the xylem network of Arabidopsis. Plant Cell Physiol. 2015, 56, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Knox, K.; Paterlini, A.; Thomson, S.; Oparka, K.J. The coumarin glucoside, esculin, reveals rapid changes in phloem-transport velocity in response to environmental cues. Plant Physiol. 2018, 44. [Google Scholar] [CrossRef] [PubMed]
- Sugita, R.; Kobayashi, N.I.; Hirose, A.; Saito, T.; Iwata, R.; Tanoi, K.; Nakanishi, T.M. Visualization of uptake of mineral elements and the dynamics of photosynthates in arabidopsis by a newly developed Real-Time Radioisotope Imaging System (RRIS). Plant Cell Physiol. 2016, 57, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Hubeau, M.; Steppe, K. Plant-PET Scans: In Vivo Mapping of Xylem and Phloem Functioning. Trends Plant Sci. 2015, 20, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Shulse, C.N.; Cole, B.J.; Turco, G.M.; Zhu, Y.; Brady, S.M.; Dickel, D.E. High-throughput single-cell transcriptome profiling of plant cell types. bioRxiv 2018, 1–19. [Google Scholar] [CrossRef]
Figure 1.
Organic and inorganic solutes take several paths to enter and exit the plant vascular system. This scheme is based on the Arabidopsis thaliana anatomy. (A,C,E) Schematic representation of a source organ (source leaf) (A), a transport organ (floral stem) (C), and a sink organ (root) (E). (B,D,F) Schematic representation of the possible transport pathways taken by the organic and inorganic solutes between the different cell types in each organ ((B): leaf; (D): floral stem; and (F): root). 1. Loading of carbohydrates, organic acids, and amino acids in the sieve tubes. 2. Water flow between xylem and phloem. 3. Lateral transfer of amino acids from xylem to phloem. 4. N metabolism and N remobilization. 5. Leakage and retrieval of carbohydrates, amino acids, and ions between the phloem and the surroundings tissues. 6. Unloading of carbohydrates, organic acids, amino acids, and ions for the supply of metabolic precursors for cell division and expansion. 7. Uptake, efflux, and influx of inorganic solutes (e.g., NO3−, PO43−, K+, SO42−) and nitrogen to the xylem. 8. Absorption and flow of water to the xylem. 9. Unloading of carbohydrates, organic acids, and amino acids that will later be used as precursors for cell division and expansion. The light blue, pale yellow, pink, and green arrows represent the water, inorganic solutes, nitrogen, and sugar movement, respectively. The circles represent the transporter-mediated movement of organic and inorganic solutes. BS: bundle sheath; CC: companion cell; En: endodermis; PD: plasmodesmata; Ph: phloem; pP: phloem parenchyma cell; PPP: phloem–pole pericycle; Px: protoxylem; MC: mesophyll cell; Mx: metaxylem; SE: sieve element; xV: xylem vessel; xP: xylem parenchyma cell; XPP: xylem-pole pericycle.
Figure 1.
Organic and inorganic solutes take several paths to enter and exit the plant vascular system. This scheme is based on the Arabidopsis thaliana anatomy. (A,C,E) Schematic representation of a source organ (source leaf) (A), a transport organ (floral stem) (C), and a sink organ (root) (E). (B,D,F) Schematic representation of the possible transport pathways taken by the organic and inorganic solutes between the different cell types in each organ ((B): leaf; (D): floral stem; and (F): root). 1. Loading of carbohydrates, organic acids, and amino acids in the sieve tubes. 2. Water flow between xylem and phloem. 3. Lateral transfer of amino acids from xylem to phloem. 4. N metabolism and N remobilization. 5. Leakage and retrieval of carbohydrates, amino acids, and ions between the phloem and the surroundings tissues. 6. Unloading of carbohydrates, organic acids, amino acids, and ions for the supply of metabolic precursors for cell division and expansion. 7. Uptake, efflux, and influx of inorganic solutes (e.g., NO3−, PO43−, K+, SO42−) and nitrogen to the xylem. 8. Absorption and flow of water to the xylem. 9. Unloading of carbohydrates, organic acids, and amino acids that will later be used as precursors for cell division and expansion. The light blue, pale yellow, pink, and green arrows represent the water, inorganic solutes, nitrogen, and sugar movement, respectively. The circles represent the transporter-mediated movement of organic and inorganic solutes. BS: bundle sheath; CC: companion cell; En: endodermis; PD: plasmodesmata; Ph: phloem; pP: phloem parenchyma cell; PPP: phloem–pole pericycle; Px: protoxylem; MC: mesophyll cell; Mx: metaxylem; SE: sieve element; xV: xylem vessel; xP: xylem parenchyma cell; XPP: xylem-pole pericycle.

Figure 2.
Model for sugar and ion transport in the Arabidopsis floral stem. The upper panel shows a vascular bundle transversal section stained with alcian blue/safranin O. The primary cell walls appear in different shades of orange and the secondary cell walls appear in red. The lower panel is a sketch showing a longitudinal view of the different cell types present in a vascular bundle. The white bridges between two cells represent the plasmodesmata. The model was based on current knowledge of the spatial distribution of sugar and ion transporters in the Arabidopsis floral stem. Known locations of members of the sugar and ion transporters in other Arabidopsis organ or in woody stem have been added as putative locations in Arabidopsis floral stem (question marks). pPC: phloem parenchyma cell; CC: companion cell; SE: sieve element; SUC: SUCROSE TRANSPORTER; SWEET: SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTERS; xV: xylem vessel; xF: xylary fiber; xPC: xylem parenchyma cell.
Figure 2.
Model for sugar and ion transport in the Arabidopsis floral stem. The upper panel shows a vascular bundle transversal section stained with alcian blue/safranin O. The primary cell walls appear in different shades of orange and the secondary cell walls appear in red. The lower panel is a sketch showing a longitudinal view of the different cell types present in a vascular bundle. The white bridges between two cells represent the plasmodesmata. The model was based on current knowledge of the spatial distribution of sugar and ion transporters in the Arabidopsis floral stem. Known locations of members of the sugar and ion transporters in other Arabidopsis organ or in woody stem have been added as putative locations in Arabidopsis floral stem (question marks). pPC: phloem parenchyma cell; CC: companion cell; SE: sieve element; SUC: SUCROSE TRANSPORTER; SWEET: SUGAR WILL EVENTUALLY BE EXPORTED TRANSPORTERS; xV: xylem vessel; xF: xylary fiber; xPC: xylem parenchyma cell.


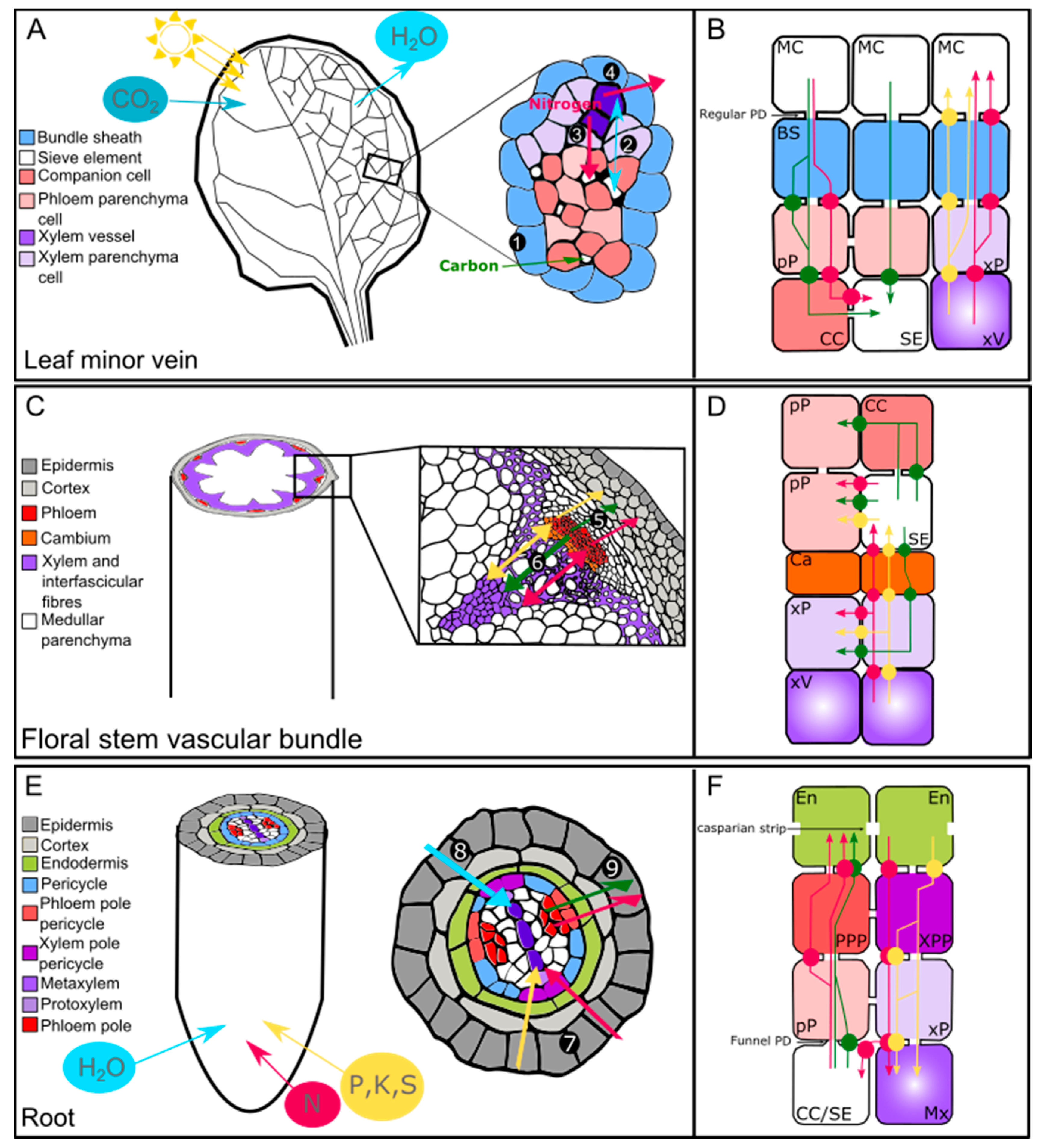{kind=link}
{kind=link}
Table 1.
Experimental evidence for lateral transport of solutes in angiosperms and gymnosperms.
| Lateral Process | Compound | Method Used | Species | Reference |
|---|---|---|---|---|
| Xylem-to-phloem | Sucrose | 14C-sucrose | Populus deltoids | [32] |
| Glumatic acid, aspartic acid | 14C-labeled amino acid | Populus deltoids | [32] | |
| Valine, asparagine, threonine, serine, citrulline, glutamine | 14C-labeled amino acid | Lupinus albus | [33] | |
| Asparagine | 14C-asparagine | Spartium junceum | [34] | |
| Glucose | 14C-glucose | Vitis vinifera | [35] | |
| Inulin, valine, aminobutyric acid | 14C-labeled compounds | Lycopersicum esculentum | [28] | |
| Carbon and nitrogen | C/N ratio, sap analysis, and modeling | Glycine max | [30] | |
| Potassium and sodium | Sap analysis | Salix viminalis | [26] | |
| Potassium, sodium, and phosphorus | Radioactive isotopes | Pelargonium zonale Salix lasiandra | [25] | |
| Sucrose, glucose, and fructose | 14C-labeled sugars | Salix viminalis | [36] | |
| Glutamic acid, aspartic acid | 14C-labeled amino acid | Salix viminalis | [37] | |
| Phloem-to-xylem | Photosynthates | Physiological analysis | Ricinus communis | [31] |
| Photosynthates | 14CO2 | Vitis labruscana | [38] | |
| Glucose, fructose, and sucrose | 14C-labeled sugars | Glycine max | [39] | |
| Sucrose | 14CO2 | Salix viminalis | [40] | |
| Phosphorus and sulfur | 35S and 32P tracking experiments | Phaseolus vulgaris | [41] | |
| Leakage-retrieval process along the phloem | Photosynthates | 11CO2 | Phaseolus vulgaris | [42] |
| Sucrose and hexoses | Ringing experiment | Gossypium sp. | [43] | |
| Photosynthates | Defoliation experiment | Fraxinus americana | [44] | |
| Sucrose | 11CO2 tracking experiment | Arabidopsis thaliana | [45] | |
| Bidirectional exchange | Glutamine and asparagine | 14C- and 15N-labeled compounds | Picea abies | [46] |
| Potassium and sodium | Sap analysis | Lupinus albus | [47] | |
| Potassium | Sap analysis and modeling | Ricinus communis | [31] | |
| Potassium | Sap analysis | Hordeum vulgare | [47] | |
| Calcium and magnesium | Sap analysis | Ricinus communis | [48] | |
| Water | Fluorescent marker | Eucalyptus saligna | [5] |
Table 2.
Plasma membrane solute transporters showing a localization in vascular or perivascular cells in dicotyledonous species.
Table 2.
Plasma membrane solute transporters showing a localization in vascular or perivascular cells in dicotyledonous species.
| Gene Name | Gene Locus or Accession n° | Function | Organ | Expression Domain | Method Used | Species/[Reference] |
|---|---|---|---|---|---|---|
| Ion transport | ||||||
| AKT2/3 | At4g22200 | Potassium/sodium channel | Leaf, stem | Phloem and xylem parenchyma | Promoter GUS fusion | At/[94,95] |
| CCC | At1g30450 | Cation–chloride cotransporter | Leaf, root | Xylem parenchyma | Promoter GUS fusion | At/[96] |
| CHX14 | At1g06970 | Potassium efflux transporter | Root | Xylem parenchyma | Promoter GUS fusion | At/[97] |
| CHX21 | At2g31910 | Sodium transporter | Root | Endodermis | Protein immunolocalization | At/[98] |
| HKT1 | At4g10310 | Sodium/Potassium symporter | Leaf, root and stem | Phloem/xylem parenchyma, endodermis | Immuno-electron microscopy and promoter GUS fusion | At/[99,100] |
| NRT1.5/NPF7.3 | At1g32450 | Potassium and nitrate transporter | Root | Pericycle | In situ hybridization | At/[101] |
| PHO1 | At3g23430 | Inorganic phosphate exporter | Root | Stele, xylem-pole, endodermis | Promoter GUS fusion | At/[102] |
| PHO1;H1 | At1g68740 | Inorganic phosphate transporter | Leaf, root | Vascular cylinder, pericycle | Promoter GUS fusion | At/[103] |
| PHT1;5 | At2g32830 | Inorganic phosphate transporter | Leaf | Phloem | Promoter GUS fusion | At/[104] |
| GmPT7 | FJ814695 | Phosphate transporter | Leaf, root | Vascular cylinder | Promoter GUS fusion | Gm/[105] |
| SKOR | At3g02850 | Potassium efflux transporter | Root | Xylem parenchyma, pericycle | Promoter GUS fusion | At/[106] |
| SLAH1-3 | At1g62280; At4g27970; At5g24030 | Anion channel | Root | Xylem-pole pericycle | Promoter GUS and GFP fusion | At/[107] |
| SOS1 | At2g01980 | Sodium antiporter | Leaf, stem, root | Xylem parenchyma, pericycle | Promoter GUS fusion | At/[108] |
| SULTR2;1 | At5g10180 | Sulfate transporter | Leaf, root | Xylem parenchyma, phloem, pericycle | Promoter GUS and GFP fusion | At/[109] |
| SULTR2;2 | At1g77990 | Sulfate transporter | Leaf, root | Bundle sheath, phloem parenchyma | Promoter GUS and GFP fusion | At/[109] |
| SULTR3;5 | At5g19600 | Sulfate transporter | Root | Xylem parenchyma, pericycle | Promoter GFP fusion | At/[110] |
| PtaSULTR1;1 | DQ906929 | Sulfate transporter | Stem | Phloem companion cells, cambium | In situ hybridization | Pta/[111] |
| PtaSULTR3;3a | DQ906924 | Sulfate transporter | Leaf, stem | Companion cells, xylem parenchyma, rays | In situ hybridization | Pta/[111] |
| Sugar transport | ||||||
| DcSUT2 | Y16768 | Sucrose transporter | Root | Xylem and phloem parenchyma | Northern blot | Dc/[112] |
| JrSUT1 | AY504969 | Sucrose transporter | Stem | Xylem parenchyma | In situ hybridization | Jr/[113] |
| PtaSUT3 | POPTR_0019s11560 | Sucrose transporter | Stem | Xylem vessel, fiber | In situ hybridization | Pta/[114] |
| PttSUT3 | POPTR_0019s11560 | Sucrose transporter | Stem | Cambium, xylem vessels | Gene expression by qPCR | Ptt/[77] |
| SUC2 | At1g22710 | Sucrose transporter | Leaf | Phloem companion cells | Promoter GUS/GFP fusion | At/[115,116] |
| SUC3 | At2g02860 | Sucrose transporter | Leaf, stem | Phloem companion cells | Protein immunolocalization and promoter GFP fusion | At/[117,118] |
| SUT1 | X82276 (Nt); X82275 (Sly); X69165 (St) | Sucrose transporter | Leaf | Xylem parenchyma | Protein immunolocalization | St, Nt, Sly/[119] |
| SWEET4 | At3g28007 | Hexose facilitator | Root | Stele | Promoter GUS fusion | At/[120] |
| SWEET11 | At3g48740 | Sucrose and hexose facilitator | Leaf, stem | Phloem transfer cells, phloem/xylem parenchyma | GFP fusion protein | At/[68,121] |
| SWEET12 | At5g23660 | Sucrose and hexose facilitator | Leaf, stem | Phloem transfer cells, phloem/xylem parenchyma | GFP fusion protein | At/[68,121] |
| IbSWEET10 | Sucrose transporter | Root | Stele | Promoter GUS fusion | Ib/[122] | |
| Organic and inorganic nitrogen | ||||||
| AAP2 | At5g09220 | Amino acid transporter | Leaf, stem | Phloem companion cells | Promoter GUS and GFP fusion protein | At/[18] |
| AAP6 | At5g49630 | Amino acid transporter | Leaf | Xylem parenchyma | Promoter GUS | At/[123] |
| AMT1;1 | At4g13510 | Ammonium transporter | Root | Pericycle | GFP fusion protein | At/[124] |
| AMT2;1 | At2g38290 | Ammonium transporter | Root | Pericycle | Promoter GFP | At/[125] |
| AtProT1 | At2g39890 | Proline transporter | Leaf, root | Phloem and phloem parenchyma | Promoter GFP | At/[126] |
| NPF2.3 | At3g45680 | Nitrate transporter | Root | Pericycle | Promoter GUS and GFP fusion protein | At/[127] |
| NRT1.4/NPF6.2 | At2g26690 | Nitrate transporter | Leaf | Vascular system | In situ hybridization | At/[128,129] |
| NRT1.8/NPF7.2 | At4g21680 | Nitrate transporter | Root | Xylem parenchyma | Promoter GUS and in situ hybridization | At/[130] |
| NRT1.9/NPF2.9 | At1g18880 | Nitrate transporter | Root | Phloem companion cells | Promoter GUS and GFP fusion protein | At/[131] |
| NRT1.11/NPF1.2 | At1g52190 | Nitrate transporter | Leaf | Phloem companion cells | Promoter GUS and GFP fusion protein | At/[132] |
| NRT1.12/NPF1.1 | At3g16180 | Nitrate transporter | Leaf | Phloem companion cells | Promoter GUS and GFP fusion protein | At/[132] |
At: Arabidopsis thaliana; Bv: Beta vulgaris; Dc: Daucus carota; Gm: Glycine max; Ib: Ipomoea batatas; Jr: Juglans regia; Nt: Nicotiana tabacum; Vv: Vitis vinifera; Pta: Populus tremula x Populus alba; Ptt: Populus tremula x Populus tremuloïdes; Sly: Solanum lycopersicum; St: Solanum tuberosum.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aubry, E.; Dinant, S.; Vilaine, F.; Bellini, C.; Le Hir, R. Lateral Transport of Organic and Inorganic Solutes. Plants 2019, 8, 20. https://doi.org/10.3390/plants8010020
AMA Style
Aubry E, Dinant S, Vilaine F, Bellini C, Le Hir R. Lateral Transport of Organic and Inorganic Solutes. Plants. 2019; 8(1):20. https://doi.org/10.3390/plants8010020
Chicago/Turabian StyleAubry, Emilie, Sylvie Dinant, Françoise Vilaine, Catherine Bellini, and Rozenn Le Hir. 2019. "Lateral Transport of Organic and Inorganic Solutes" Plants 8, no. 1: 20. https://doi.org/10.3390/plants8010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.