Whey: The Soil Bio-Community Enhancer That Selectively Controls Root-Knot Nematodes

 and
and 
Abstract
:1. Introduction
2. Results
2.1. Effects of Whey on the Microbial Communities
2.2. Effects of Whey on Enzyme Activities
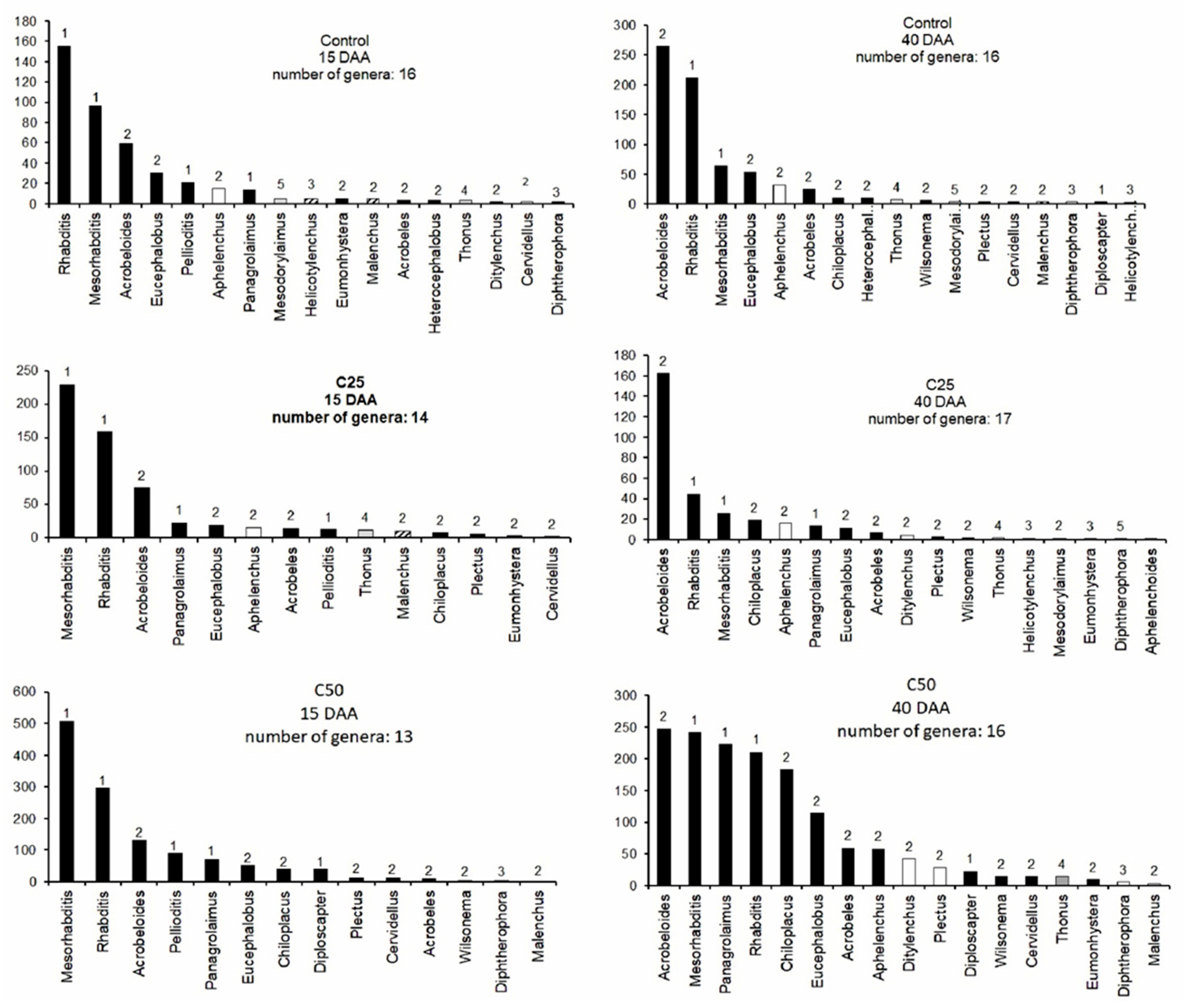
2.3. Effects of Whey on Nematode Bio-Communities
2.4. Paralysis Effect of Whey on the Plant Parasitic Nematode M. Javanica
2.5. Field Experiment
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Pot Bioassay
5.1.1. Phospholipid Fatty Acid (PLFA) Analysis
5.1.2. Enzyme Activity Assays
5.1.3. Soil Nematodes Extraction of and Community Analysis
5.2. Paralysis Bioassays Against Meloidogyne Javanica
5.3. Field-Efficacy Experiment against the Root Knot Nematodes
5.4. Statistical Analysis
5.4.1. Testing Effects of Whey as a Soil Amendment
5.4.2. Paralysis Effects of Whey on Meloidogyne javanica and Field-Efficacy Experiment
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| c-p | Colonizer-Persister |
| C25 | 3.125% (v/w) of whey in soil |
| C50 | 6.25% (v/w) of whey in soil |
| EC50/3d | half maximal Effective Concentration after 3 days |
References
- Gil-Pulido, Β.; Tarpey, Ε.; Almeida, Ε.L.; Finnegan, W.; Zhan, X.; Dobson, A.D.W.; O’Leary, N. Evaluation of dairy processing wastewater biotreatment in an IASBR system: Aeration rate impacts on performance and microbial ecology. Biotechnol. Rep. 2018, 19, e00263. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.; Prazeres, A.R.; Rivas, J. Cheese whey wastewater: Characterization and treatment. Sci. Total Environ. 2013, 445–446, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Grosu, L.; Fernandez, B.; Grigoraş, C.G.; Patriciu, O.I.; Grid-Alexa, I.C.; Nicuţă, D.; Ciobanu, D.; Gavrilă, L.; Fînaru, A.L. Valorization of whey from dairy industry for agricultural use as fertiliser: Effects on plant germination and growth. Environ. Eng. Manag. J. 2012, 11, 2203–2210. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Ferris, H.; Venette, R.C.; Scow, K.M. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralization function. Appl. Soil Ecol. 2004, 25, 19–35. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity interrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Monokrousos, N.; Papatheodorou, E.M.; Diamantopoulos, J.; Stamou, G.P. Soil quality variables in organically and conventionally cultivated field sites. Soil Biol. Biochem. 2006, 38, 1282–1289. [Google Scholar] [CrossRef]
- Commission Staff Working Document, Basic Substance Whey; European Commission Health and Food Safety Directorate-General: Brussel, Belgium, 2016.
- Demir, S.; Şensoy, S.E.; Ocak, E.; Tüfenkçi, S.; Durak, E.D.; Erdinç, C.; Ünsal, H. Effects of arbuscular mycorrhizal fungus, humic acid, and whey on wilt disease caused by Verticillium dahliae Kleb. in three solanaceous crops. Turk. J. Agric. For. 2015, 39, 300–309. [Google Scholar] [CrossRef]
- Machado, A.C.Z. Current nematode threats to Brazilian agriculture. Curr. Agric. Sci. Technol. 2014, 20, 26–35. [Google Scholar]
- Jones, J.G.; Kleczewski, N.M.; Desaeger, J.; Meyer, S.L.; Johnson, G.C. Evaluation of nematicides for southern root knot nematode management in lima bean. Crop Prot. 2017, 96, 151–157. [Google Scholar] [CrossRef]
- Lagos, S.; Perruchon, C.; Katsoula, A.; Karpouzas, D.G. Isolation and characterization of soil bacteria able to rapidly degrade the organophosphorus nematicide fosthiazate. Lett. Appl. Microbiol. 2019, 68, 149–155. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation (EC) No 1107/2009 of the European 515 Parliament and of the Council of 21 October 2009 concerning the placing of plant 516 protection products on the market and repealing Council Directives 79/117/EEC 517 and 91/414/EEC. Off. J. Eur. Union 2009, 309, 1–50. [Google Scholar]
- Villaverde, J.J.; Sandín-España, P.; Sevilla-Morán, B.; López-Goti, C.; Alonso-Prados, J.L. Biopesticides from Natural Products: Current Development, Legislative Framework, and Future Trends. Bioresources 2016, 11, 5618–5640. [Google Scholar] [CrossRef]
- Spence, K.O.; Lewis, E.E. Biopesticides with complex modes of action: Direct and indirect effects of DiTera® on Meloidogyne incognita. Nematology 2010, 12, 835–846. [Google Scholar] [CrossRef]
- Bohnenstengel, F.I.; Wray, V.; Witte, L.; Srivastava, R.P.; Proksch, P. Insecticidal meliacarpins (C-Seco limonoids) from Melia azedarach. Phytochemistry 1999, 50, 977–982. [Google Scholar] [CrossRef]
- Kraal, P.; Klaas, G.J.N.; Kaal, J.; Tietema, A. Carbon respiration and nitrogen dynamics in Corsican pine litter amended with aluminium and tannins. Soil Biol. Biochem. 2009, 41, 2318–2327. [Google Scholar] [CrossRef]
- Papatheodorou, E.M.; Kordatos, H.; Kouseras, T.; Monokrousos, N.; Menkissoglu-Spiroudi, U.; Diamantopoulos, J.; Stamou, G.P.; Argyropoulou, M.D. Differential responses of structural and functional aspects of soil microbes and nematodes to abiotic and biotic modifications of the soil environment. Appl. Soil Ecol. 2012, 61, 26–33. [Google Scholar] [CrossRef]
- Sanaullah, M.; Blagodatskaya, E.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Drought effects on microbial biomass and enzyme activities in the rhizosphere of grasses depend on plant community composition. Appl. Soil Ecol. 2011, 48, 38–44. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Verhoeven, R. Response of soil microfauna to organic fertilization in sandy virgin soils of coastal dunes. Biol. Fertil. Soils 2001, 34, 390–396. [Google Scholar] [CrossRef]
- Meidute, S.; Demoling, F.; Bååth, E. Antagonistic and synergistic effects of fungal and bacterial growth in soil after adding different carbon and nitrogen sources. Soil Biol. Biochem. 2008, 40, 2334–2343. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Esperschütz, J.; Gattinger, A.; Mäder, P.; Schloter, M.; Fließbach, A. Response of soil microbial biomass and community structures to conventional and organic farming systems under identical crop rotations. FEMS Microbiol. Ecol. 2007, 61, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Simpson, M.J. Temperature and substrate controls on microbial phospholipid fatty acid composition during incubation of grassland soils contrasting in organic matter quality. Soil Biol. Biochem. 2009, 41, 804–812. [Google Scholar] [CrossRef]
- Ntalli, N.; Monokrousos, N.; Rumbos, C.; Kontea, D.; Zioga, D.; Argyropoulou, D.M.; Menkissoglu-Spiroudi, U.; Tsiropoulos, G.N. Greenhouse biofumigation with Melia azedarach controls Meloidogyne spp. and enhances soil biological activity. J. Pest Sci. 2018, 91, 29–40. [Google Scholar]
- Chodak, M.; Niklińska, M. Effect of texture and tree species on microbial properties of mine soils. Appl. Soil Ecol. 2010, 46, 268–275. [Google Scholar] [CrossRef]
- Izquierdo, I.; Caravaca, F.; Alguacil, M.M.; Hernández, G.; Roldán, A. Use of microbiological indicators for evaluating success in soil restoration after revegetation of a mining area under subtropical conditions. Appl. Soil Ecol. 2005, 30, 3–10. [Google Scholar] [CrossRef]
- Monokrousos, N.; Charalampidis, G.; Boutsis, G.; Sousanidou, V.; Papatheodorou, E.M.; Argyropoulou, M.D. Plant-induced differentiation of soil variables and nematode community structure in a Mediterranean serpentine ecosystem. Soil Res. 2014, 52, 593–603. [Google Scholar] [CrossRef]
- Visser, S.; Parkinson, D. Soil biological criteria as indicators of soil quality: Soil microorganisms. Am. J. Altern. Agric. 1992, 7, 33–37. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Nannipieri, P.; Pedrazzini, F.; Arcara, P.G.; Piovanelli, C. Changes in amino acids, enzyme activities, and biomasses during soil microbial growth. Soil Sci. 1979, 127, 26–34. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, X.; Deng, S.; Dong, X.; Song, A.; Yao, J.; Fang, W.; Chen, F. The Effects of Fungicide, Soil Fumigant, Bio-Organic Fertilizer and Their Combined Application on Chrysanthemum Fusarium Wilt Controlling, Soil Enzyme Activities and Microbial Properties. Molecules 2016, 21, 526. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Dong, Y.; Wan, Y.; Hu, G.; Wang, Y. Effects of Different Fertilizing Treatments on Contents of Soil Nutrients and Soil Enzyme Activity. Soils 2010, 42, 249–255. [Google Scholar]
- Tsiafouli, M.A.; Argyropoulou, M.D.; Stamou, G.P.; Sgardelis, S.P. Is duration of organic management reflected on nematode communities of cultivated soils? Belg. J. Zool 2007, 137, 165–175. [Google Scholar]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; de Ruiter, P.C.; van der Putten, W.H.; Birkhofer, K.; Hemerik, L.; de Vries, F.T.; Bardgett, R.D.; Brady, M.V.; et al. Intensive agriculture reduces soil biodiversity across Europe. Glob. Chang. Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef]
- Ntalli, N.; Zioga, D.; Argyropoulou, D.M.; Papatheodorou, M.E.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Anise, parsley and rocket as nematicidal soil amendments and their impact on non-target soil organisms. Appl. Soil Ecol. 2019, 143, 17–25. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Ferris, H.; Matute, M.M. Structural and functional succession in the nematode fauna of a soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Steel, H.; Buchan, D.; Bert, W.; De Neve, S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Sci. Rep. 2016, 6, 32862. [Google Scholar] [CrossRef]
- Salinas, K.A.; Edenborn, S.L.; Sexstone, A.J.; Kotcon, J.B. Bacterial preferences of the bacterivorous soil nematode Cephalobus brevicauda (Cephalobidae): Effect of bacterial type and size. Pedobiologia 2007, 51, 55–64. [Google Scholar] [CrossRef]
- Yu, L.; Yan, X.; Ye, C.; Zhao, H.; Chen, X.; Hu, F.; Li, H. Bacterial respiration and growth rates affect the feeding preferences, brood size and lifespan of Caenorhabditis elegans. PLoS ONE 2015, 10, e0134401. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, J.; Chen, Y.; Chen, X.; Chen, L.; Li, H.; Zhang, X.X.; Sun, B. Nematode grazing promotes bacterial community dynamics in soil at the aggregate level. ISME J. 2017, 11, 2705–2717. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yu, L.; Xu, J.; Yan, X.; Li, H.; Whalen, J.K.; Hu, F. Bacterial traits and quality contribute to the diet choice and survival of bacterial-feeding nematodes. Soil Biol. Biochem. 2017, 115, 467–474. [Google Scholar] [CrossRef]
- Siddiqi, M.R. Tylenchida: Parasites of Plants and Insects; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Boulton, A.M.; Jaffee, B.A.; Scow, K.M. Effects of a common harvester ant (Messor andrei) on richness and abundance of soil biota. Appl. Soil Ecol. 2003, 23, 257–265. [Google Scholar] [CrossRef]
- Monokrousos, N.; Boutsis, G.; Diamantopoulos, J.D. Development of soil chemical and biological properties in the initial stages of post-mining deposition sites. Environ. Monit. Assess. 2014, 186, 9065–9074. [Google Scholar] [CrossRef]
- Ugarte, C.M.; Zaborski, E.R.; Wander, M.M. Nematode indicators as integrative measures of soil condition in organic cropping systems. Soil Biol. Biochem. 2013, 64, 103–113. [Google Scholar] [CrossRef]
- Neher, D.A. Role of nematodes in soil health and their use as indicators. J. Nematol. 2001, 33, 161. [Google Scholar]
- Zeng, S.; Brown, S.; Przemeck, S.M.C.; Simpson, H.V. Milk and milk components reduce the motility of Ostertagia circumcincta larvae in vitro. N. Z. Vet. J. 2003, 51, 174–178. [Google Scholar] [CrossRef]
- Marchand, P.A. Basic substances: An opportunity for approval of low-concern substances under EU pesticide regulation. Pest Manag. Sci. 2015, 71, 1197–1200. [Google Scholar] [CrossRef]
- Karpouzas, D.G.; Karanasios, E.; Menkissoglu-Spiroudi, U. Enhanced microbial degradation of Cadusafos in potato monoculture: Demonstration and characterization. Chemosphere 2004, 56, 549–559. [Google Scholar] [CrossRef]
- Papadopoulou, E.S.; Karpouzas, D.G.; Menkissoglu-Spiroudi, U. Extraction parameters significantly influence and the quantity and the profile of PLFAs extracted from soil. Microb. Ecol. 2011, 62, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Spyrou, I.M.; Karpouzas, D.G.; Menkissoglu-Spiroudi, U. Do botanical pesticides alter the structure of the soil microbial community? Microb. Ecol. 2009, 58, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Findlay, R.H. Determination of microbial community structure using phospholipid fatty acid profiles. In Molecular Microbial Ecology Manual, 2nd ed.; Kowalchuk, G.A., Bruijn, F.J.D., Head, I.M., Akkermans, A.D.L., Elsas, J.D.V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 983–1004. [Google Scholar]
- Rousidou, C.; Papadopoulou, E.S.; Kortsinidou, M.; Giannakou, I.O.; Singh, B.K.; Menkissoglu-Spiroudi, U.; Karpouzas, D.G. Biopesticides: Harmful or harmless to ammonia oxidizing microorganisms? The case of a Paecilomyces lilacinus-based nematicide. Soil Biol. Biochem. 2013, 67, 98–105. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties; Weaver, R.W., Angles, J.S., Bottomley, P.S., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. [Google Scholar]
- S’Jacob, J.J.; van Bezooijen, J. A Manual for Practical Work in Nematology; Department of Nematology, Wageningen Agricultural University: Wageningen, The Netherlands, 1984. [Google Scholar]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Hussey, R.S.; Barker, K.R. Comparison of methods of collecting inocula of Meloidogyne spp. including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Ntalli, N.G.; Menkissoglu-Spiroudi, U.; Giannakou, I. Nematicidal activity of powder and extracts of Melia azedarach fruits against Meloidogyne incognita. Ann. Appl. Biol. 2010, 156, 309–317. [Google Scholar] [CrossRef]
- Anderson, M.J. PERMANOVA_2Factor: A FORTRAN Computer Program for Permutational Multivariate Analysis of Variance (for Any Two-Factor ANOVA design) Using Permutation Tests; Department of Statistics, University of Auckland: Auckland, New Zealand, 2004. [Google Scholar]
- Bongers, T. De nematoden van Nederland; Pirola: Schoorl, The Netherlands, 1994. [Google Scholar]
- Puntener, W. Manual for field trials. In Plant Protection, 2nd ed.; Ciba Geigy Limited: Basle, Switzerland, 1981; 205p. [Google Scholar]
- Seefeldt, S.S.; Jensen, J.E.; Fuerst, E.P. Log-logistic analysis of herbicide dose response relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Exposure | EC50 (% v/v) | Std. Error | CI95% |
|---|---|---|---|
| 1 | 4.9 | 0.602 | 3.6–6.1 |
| 3 | 3.2 | 0.137 | 2.9–3.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntalli, N.; Tsiafouli, M.A.; Tzani, K.; Mavridi, O.; Oplos, C.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Whey: The Soil Bio-Community Enhancer That Selectively Controls Root-Knot Nematodes. Plants 2019, 8, 445. https://doi.org/10.3390/plants8110445
Ntalli N, Tsiafouli MA, Tzani K, Mavridi O, Oplos C, Menkissoglu-Spiroudi U, Monokrousos N. Whey: The Soil Bio-Community Enhancer That Selectively Controls Root-Knot Nematodes. Plants. 2019; 8(11):445. https://doi.org/10.3390/plants8110445
Chicago/Turabian StyleNtalli, Nikoletta, Maria A. Tsiafouli, Kaliopi Tzani, Olga Mavridi, Chrisostomos Oplos, Urania Menkissoglu-Spiroudi, and Nikolaos Monokrousos. 2019. "Whey: The Soil Bio-Community Enhancer That Selectively Controls Root-Knot Nematodes" Plants 8, no. 11: 445. https://doi.org/10.3390/plants8110445