The Arabidopsis altered in stress response2 is Impaired in Resistance to Root and Leaf Necrotrophic Fungal Pathogens


Abstract
:1. Introduction
2. Results
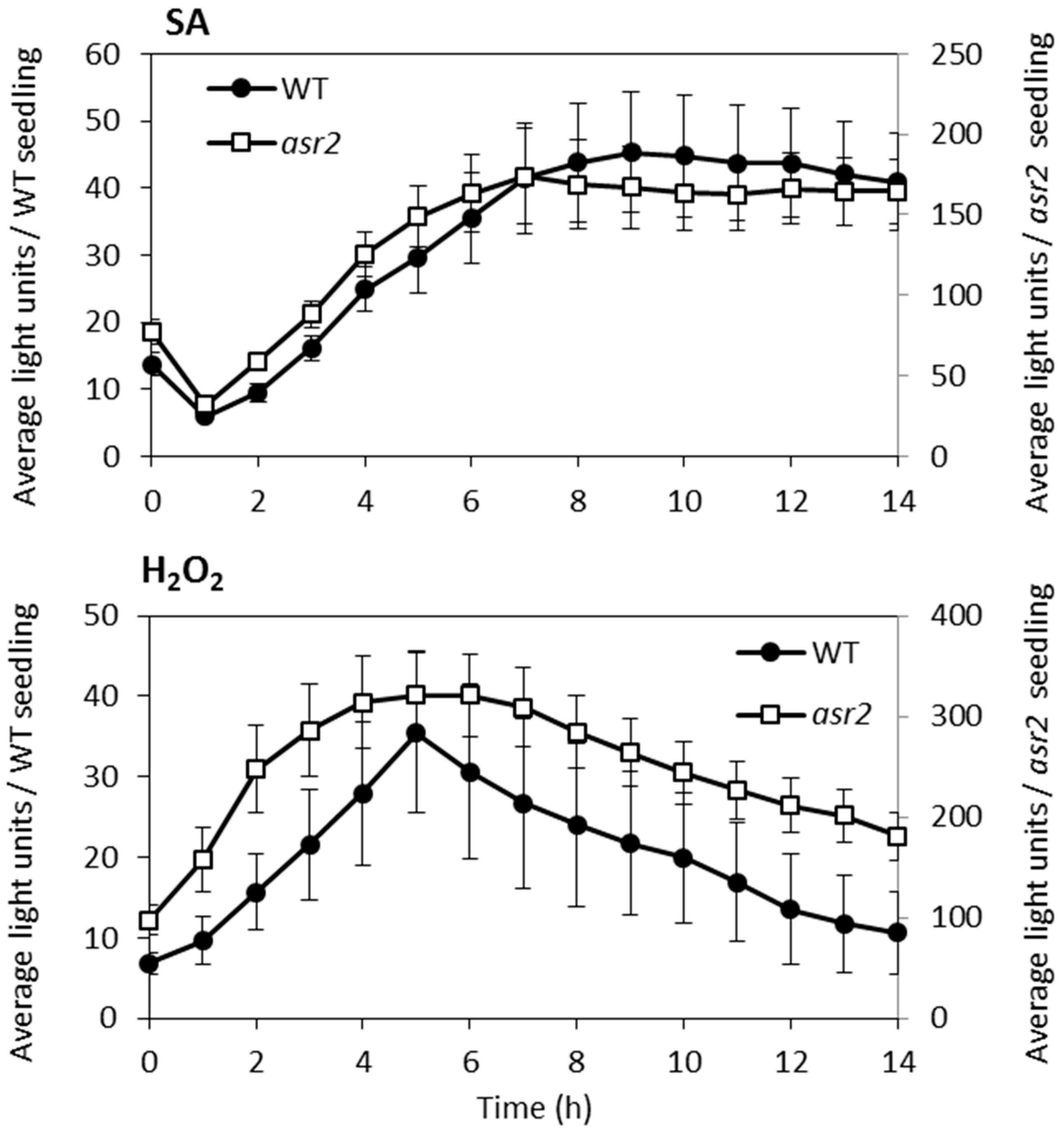
2.1. Identification of asr2
2.2. Basal Defence Gene Expression is Altered in asr2
2.3. ASR2 is Necessary for Resistance Against the Necrotrophic Fungal Pathogens Fusarium oxysporum and Alternaria brassicicola
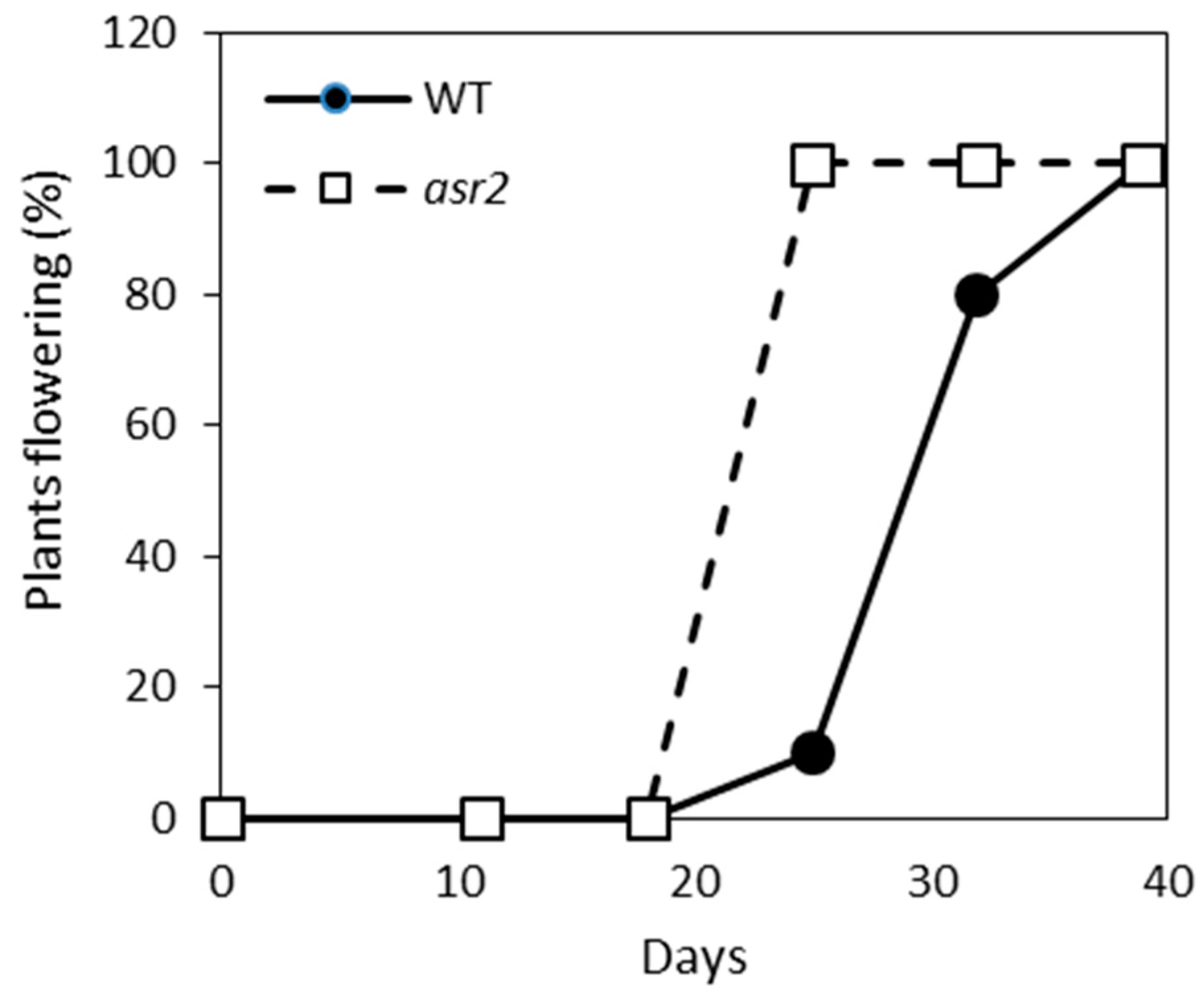
2.4. asr2 Has an Accelerated Flowering Phenotype
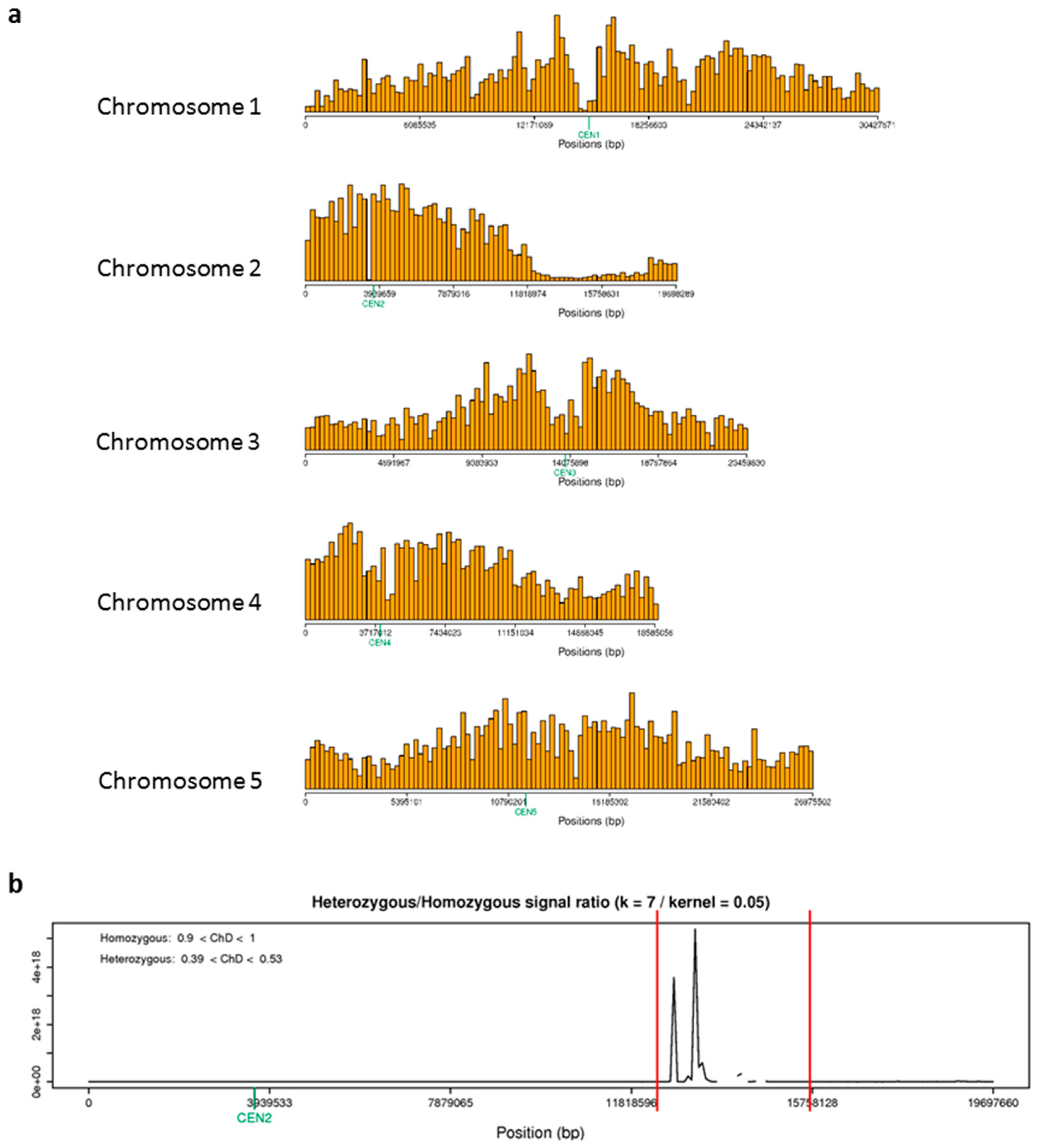
2.5. Fine Mapping Localises asr2 to Four Genes on Chromosome 2
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Bioluminescence and Luciferase Assays
4.3. Genetic and Mapping Studies
4.4. Pathogen and Flowering Time Assays
4.5. RNA Isolation and qRT-PCR
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/home/en (accessed on 19 November 2018).
- Moons, A. Regulatory and functional interactions of plant growth regulators and plant glutathione s-transferases (GSTs). Vitam. Horm. 2005, 72, 155–202. [Google Scholar] [PubMed]
- Dixon, D.P.; Edwards, R. Glutathione transferases. Arabidopsis Book 2010, 8, e0131. [Google Scholar] [CrossRef] [PubMed]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Frova, C. The plant glutathione transferase gene family: Genomic structure, functions, expression and evolution. Physiol. Plant. 2003, 119, 469–479. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Carrie, C.; Andersson, C.R.; Sivasithamparam, K.; Whelan, J.; Singh, K.B. Differential gene expression and subcellular targeting of Arabidopsis glutathione s-transferase F8 is achieved through alternative transcription start sites. J. Biol. Chem. 2007, 282, 28915–28928. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.; Huang, S.; Thatcher, L.F.; Foley, R.C.; Anderson, C.R.; Carroll, A.J.; Millar, A.H.; Singh, K.B. Mitochondrial complex ii has a key role in mitochondrial-derived reactive oxygen species influence on plant stress gene regulation and defense. Proc. Natl. Acad. Sci. USA 2011, 108, 10768–10773. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Kamphuis, L.G.; Hane, J.K.; Onate-Sanchez, L.; Singh, K.B. The Arabidopsis KH-domain rna-binding protein ESR1 functions in components of jasmonate signalling, unlinking growth restraint and resistance to stress. PLoS ONE 2015, 10, e0126978. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.E.; Thomas, V.; Truman, B.; Lilley, K.; Mansfield, J.; Grant, M. Specific changes in the Arabidopsis proteome in response to bacterial challenge: Differentiating basal and R-gene mediated resistance. Phytochemistry 2004, 65, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Chao, G.; Singh, K.B. The promoter of a H2O2-inducible, Arabidopsis glutathione s-transferase gene contains closely linked OBF- and OBP1-binding sites. Plant J. 1996, 10, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Foley, R.C.; Sappl, P.G.; Perl-Treves, R.; Millar, A.H.; Singh, K.B. Desensitization of GSTF8 induction by a prior chemical treatment is long lasting and operates in a tissue-dependent manner. Plant Physiol. 2006, 142, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Perl-Treves, R.; Foley, R.C.; Chen, W.; Singh, K.B. Early induction of the Arabidopsis GSTF8 promoter by specific strains of the fungal pathogen Rhizoctonia solani. Mol. Plant Microbe Interact. 2004, 17, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Sappl, P.G.; Carroll, A.J.; Clifton, R.; Lister, R.; Whelan, J.; Harvey Millar, A.; Singh, K.B. The arabidopsis glutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J. 2009, 58, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Sappl, P.G.; Oñate-Sánchez, L.; Singh, K.B.; Millar, A.H. Proteomic analysis of glutathione s-transferases of Arabidopsis thaliana reveals differential salicylic acid-induced expression of the plant-specific phi and tau classes. Plant Mol. Biol. 2004, 54, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Singh, K.B. The auxin, hydrogen peroxide and salicylic acid induced expression of the Arabidopsis GST6 promoter is mediated in part by an ocs element. Plant J. 1999, 19, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thatcher, L.F.; Foley, R.; Casarotto, H.J.; Gao, L.-L.; Kamphuis, L.G.; Melser, S.; Singh, K.B. The Arabidopsis RNA polymerase II carboxyl terminal domain (CTD) phosphatase-like1 (CPL1) is a biotic stress susceptibility gene. Sci. Rep. 2018, 8, 13454. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G.N. Plant diseases caused by fungi. In Plant Pathology, 5th ed.; Agrios, G.N., Ed.; Academic Press: San Diego, CA, USA, 2005; Chapter Eleven; pp. 385–614. [Google Scholar]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Dong, X. Making sense of hormone crosstalk during plant immune responses. Cell Host Microbe 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.; Eggermont, K.; Penninckx, I.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed]
- van Schie, C.C.N.; Takken, F.L.W. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Anderson, J.P.; Singh, K.B. Plant defence responses: What have we learnt from arabidopsis? Funct. Plant Biol. 2005, 32, 1–19. [Google Scholar] [CrossRef]
- Austin, R.S.; Vidaurre, D.; Stamatiou, G.; Breit, R.; Provart, N.J.; Bonetta, D.; Zhang, J.; Fung, P.; Gong, Y.; Wang, P.W.; et al. Next-generation mapping of Arabidopsis genes. Plant J. 2011, 67, 715–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manna, S. An overview of pentatricopeptide repeat proteins and their applications. Biochimie 2015, 113, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koussevitzky, S.; Nott, A.; Mockler, T.C.; Hong, F.; Sachetto-Martins, G.; Surpin, M.; Lim, J.; Mittler, R.; Chory, J. Signals from chloroplasts converge to regulate nuclear gene expression. Science 2007, 316, 715–719. [Google Scholar] [CrossRef]
- van Wees, S.C.M.; Chang, H.-S.; Zhu, T.; Glazebrook, J. Characterization of the early response of Arabidopsis to Alternaria brassicicola infection using expression profiling. Plant Physiol. 2003, 132, 606–617. [Google Scholar] [CrossRef] [PubMed]
- McGrath, K.C.; Dombrecht, B.; Manners, J.M.; Schenk, P.M.; Edgar, C.I.; Maclean, D.J.; Scheible, W.R.; Udvardi, M.K.; Kazan, K. Repressor- and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 2005, 139, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Kidd, B.N.; Edgar, C.I.; Kumar, K.K.; Aitken, E.A.; Schenk, P.M.; Manners, J.M.; Kazan, K. The mediator complex subunit PFT1 is a key regulator of jasmonate-dependent defense in Arabidopsis. Plant Cell 2009, 21, 2237–2252. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, L.F.; Kidd, B.N.; Kazan, K. Belowground defence strategies against Fusarium oxysporum. In Belowground Defence Strategies in Plants; Vos, C.M.F., Kazan, K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Part II; pp. 71–98. [Google Scholar]
- Thatcher, L.F.; Manners, J.M.; Kazan, K. Fusarium oxysporum hijacks coi1-mediated jasmonate signaling to promote disease development in Arabidopsis. Plant J. 2009, 58, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Fallath, T.; Kidd, B.N.; Stiller, J.; Davoine, C.; Bjorklund, S.; Manners, J.M.; Kazan, K.; Schenk, P.M. Mediator18 and Mediator20 confer susceptibility to Fusarium oxysporum in Arabidopsis thaliana. PLoS ONE 2017, 12, e0176022. [Google Scholar] [CrossRef] [PubMed]
- Kidd, B.N.; Aitken, E.A.; Schenk, P.M.; Manners, J.M.; Kazan, K. Plant mediator: Mediating the jasmonate response. Plant Signal. Behav. 2010, 5, 718–720. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.; Rusu, A.; Stiller, J.; Powell, J.; Manners, J.M.; Kazan, K. Investigating the association between flowering time and defense in the Arabidopsis thaliana-Fusarium oxysporum interaction. PLoS ONE 2015, 10, e0127699. [Google Scholar] [CrossRef] [PubMed]
- Rissel, D.; Heym, P.; Thor, K.; Brandt, W.; Wessjohann, L.; Peiter, E. No silver bullet—Canonical poly(adp-ribose) polymerases (parps) are no universal factors of abiotic and biotic stress resistance of Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Block, M.D.; Verduyn, C.; Brouwer, D.D.; Cornelissen, M. Poly(adp-ribose) polymerase in plants affects energy homeostasis, cell death and stress tolerance. Plant J. 2005, 41, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Liu, C.; de Oliveira, M.V.V.; Intorne, A.C.; Li, B.; Babilonia, K.; de Souza Filho, G.A.; Shan, L.; He, P. Protein poly(adp-ribosyl)ation regulates arabidopsis immune gene expression and defense responses. PLoS Genet. 2015, 11, e1004936. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, J.; An, L.; Doerge, R.W.; Chen, Z.J.; Grau, C.R.; Meng, J.; Osborn, T.C. Analysis of gene expression profiles in response to Sclerotinia sclerotiorum in Brassica napus. Planta 2007, 227, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chrom. | Position | Ref. Base | SNP Base | Depth | DC | Accession | Strand | Ref. Codon | SNP Codon | AA Change |
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 13358132 | C | T | 55 | 0.91 | AT2G31320 | - | GGA | GAA | G- > E |
| 2 | 13389698 | C | T | 57 | 0.98 | AT2G31400 | - | GAG | AAG | E- > K |
| 2 | 13612057 | C | T | 41 | 0.92 | AT2G31990 | - | GCT | ACT | A- > T |
| 2 | 14057731 | C | T | 46 | 0.91 | AT2G33170 | - | GGA | GAA | G- > E |
| 2 | 14829201 | C | T | 20 | 0.85 | AT2G35170 | + | CAA | TAA | Q- > * |
| 2 | 15025500 | C | T | 32 | 0.93 | AT2G35740 | - | GGT | AGT | G- > S |
| 2 | 15288977 | C | T | 49 | 0.96 | AT2G36420 | + | TCT | TTT | S- > F |
| 2 | 15451219 | G | A | 41 | 0.95 | AT2G36835 | - | CCG | CTG | P- > L |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thatcher, L.F.; Singh, K.B. The Arabidopsis altered in stress response2 is Impaired in Resistance to Root and Leaf Necrotrophic Fungal Pathogens. Plants 2019, 8, 60. https://doi.org/10.3390/plants8030060
Thatcher LF, Singh KB. The Arabidopsis altered in stress response2 is Impaired in Resistance to Root and Leaf Necrotrophic Fungal Pathogens. Plants. 2019; 8(3):60. https://doi.org/10.3390/plants8030060
Chicago/Turabian StyleThatcher, Louise F., and Karam B. Singh. 2019. "The Arabidopsis altered in stress response2 is Impaired in Resistance to Root and Leaf Necrotrophic Fungal Pathogens" Plants 8, no. 3: 60. https://doi.org/10.3390/plants8030060