
Immune Response to SARS-CoV-2 Vaccines

 , and
, and 
Abstract
:1. Introduction

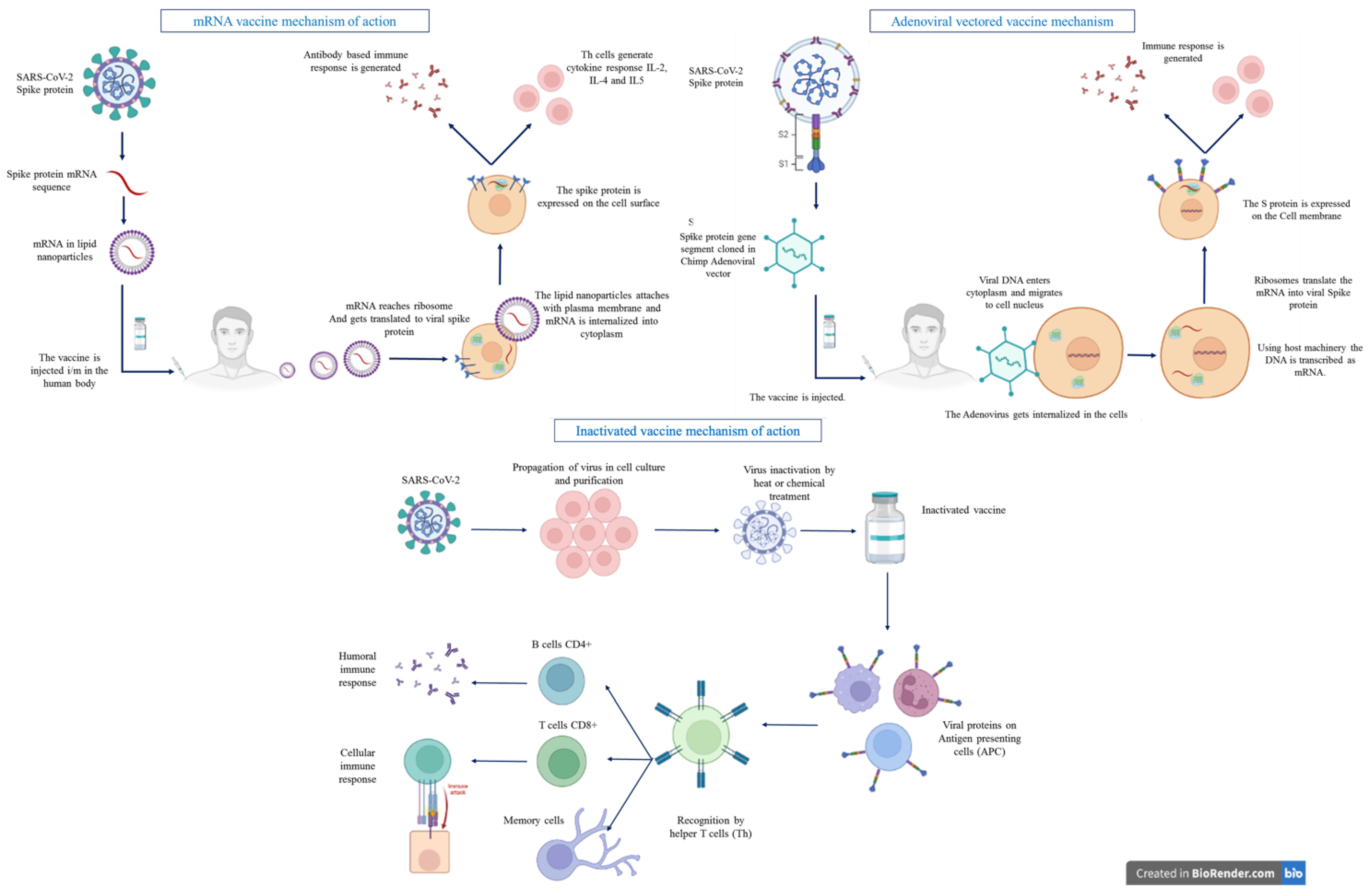{kind=link}
| Sr. no. | Vaccine | Efficacy | Side Effects |
|---|---|---|---|
| 1 | Pfizer BioNTech | N = 43,000 Overall efficacy 95% Preventing severe disease 87.5% [11] | Rare allergies and anaphylaxis |
| 2 | Moderna | N = 30,000 Overall efficacy 94.5% Preventing severe disease 100% [12] | Rare facial paralysis (Bell’s Palsy) |
| 3 | Astra—Zeneca Oxford | Overall efficacy 70% (64% after 1 dose) (70.4% after 2 doses) [13] | Rare thromboembolic events, rare cases of blood-clots, pulmonary embolism, thrombocytopenia |
| 4 | Janssen Johnson & Johnson | 72% in the USA 66% in Latin America 57% in South Africa [14] | Rare cases of blood clots, thrombocytopenia, Guillain-Barré Syndrome |
| 5 | Sputnik V | 91.6% [15] | No serious side effect |
| 6 | CoronaVac | COVID 19 Prevention: 65% Severe disease: 86–90% [16] | No serious side effect |
| 7 | Covaxin | COVID-19 prevention: 78% Severe disease: 93% Adults aged < 60 years: 79% Old Aged > 60 years: 68% [17] | Headache, fever, fatigue, muscle aches, nausea, pain, irritation, redness, and swelling at the site of the injection |
| 8 | Sinopharm BBIBP-CorV | COVID-19 prevention: 65% Severe disease: 86–90% [18,19] | No serious side effect |
| 9 | Epivac | Phase III clinical trials: 79% [20] | Data not available |
| 10 | MVC COVID-19 vaccine | Overall efficacy: 84% [21] | No vaccine-related Serious Adverse Reaction |
| 11 | Zifivax | Efficacy against COVID-19 Short-term: 81.4% Long term: 75.7% [22,23] | Injection site pain, headache, fatigue, fever, body ache, abdominal pain, nausea and vomiting, |
| 12 | CorbeVax | Efficacy: Phase II & III clinical data is pending [24] | Fever/Pyrexia, Headache, Fatigue, Arthralgia, Urticaria, Chills, Injection site swelling |
| 13 | Soberana 02 | First shot: 71% Second shot: 92.4% [25] | No serious side effect, fewer than 1% of participants in the phase III trial developed a fever |
2. Vaccine Development Approaches
3. Immune Response Generated by SARS-CoV-2 Infection
B and T Cell Immune Response
4. mRNA Vaccines
4.1. Mechanism of Action
4.2. B Cell and Antibody Response
4.3. T Cell Response
4.4. Pfizer–BioNTech and Moderna
4.4.1. Performance Evaluation of Pfizer–BioNTech Vaccine
4.4.2. Performance Evaluation of Moderna Vaccine
5. Viral Vector Vaccines
5.1. Astra-Zeneca Oxford
5.2. Johnson and Johnson (Janssen)
5.3. Sputnik V
6. Inactivated Vaccines
6.1. CoronaVac
6.2. BBIBP-CorV
6.3. WIBP-CorV (Sinopharm)
6.4. Covaxin
7. Subunit Vaccines
7.1. NVX-CoV2373
7.2. EpiVacCorona
7.3. MVC COVID-19 Vaccine
7.4. Zifivax
7.5. Corbevax
7.6. Soberana 02
8. COVID-19 Vaccine Adjuvants
8.1. B Defensing
8.2. Alum, Emulsion and Liposome
8.3. CpG Adjuvant
8.4. SARS-CoV-2 Variants and Their Effect on Vaccines
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Chi, W.; Su, J.; Ferrall, L.; Hung, C.; Wu, T. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 104. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, K. The success of SARS-CoV-2 vaccines and challenges ahead. Cell Host Microbe 2021, 29, 1111–1123. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Banu, S.; Singh, P.; Sowpati, D.; Mishra, R. SARS-CoV-2 genomics: An Indian perspective on sequencing viral variants. J. Biosci. 2021, 46, 22. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; Dorabawila, V.; Easton, D.; Bauer, U.; Kumar, J.; Hoen, R.; Hoefer, D.; Wu, M.; Lutterloh, E.; Conroy, M.; et al. COVID-19 Vaccine Effectiveness in New York State. N. Engl. J. Med. 2022, 386, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, E.; Ritchie, H.; Ortiz-Ospina, E.; Roser, M.; Hasell, J.; Appel, C.; Giattino, C.; Rodés-Guirao, L. A global database of COVID-19 vaccinations. Nat. Hum. Behav. 2021, 5, 947–953. [Google Scholar] [CrossRef]
- Graham, B. Rapid COVID-19 vaccine development. Science 2020, 368, 945–946. [Google Scholar] [CrossRef]
- Cavanagh, D. Severe acute respiratory syndrome vaccine development: Experiences of vaccination against avian infectious bronchitis coronavirus. Avian Pathol. 2003, 32, 567–582. [Google Scholar] [CrossRef] [Green Version]
- Saif, L. Vaccines for COVID-19: Perspectives, prospects, and challenges based on candidate SARS, MERS, and animal coronavirus vaccines. Eur. Med. J. 2020, 10, 200324. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.; Tamin, A.; Soloff, A.; D’Aiuto, L.; Nwanegbo, E.; Robbins, P.; Bellini, W.; Barratt-Boyes, S.; Gambotto, A. Effects of a SARS-associated coronavirus vaccine in monkeys. Lancet 2003, 362, 1895–1896. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Okada, K.; Kenniston, T.; Raj, V.; AlHajri, M.; Farag, E.; AlHajri, F.; Osterhaus, A.; Haagmans, B.; Gambotto, A. Immunogenicity of an adenoviral-based Middle East Respiratory Syndrome coronavirus vaccine in BALB/c mice. Vaccine 2014, 32, 5975–5982. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.; El Sahly, H.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.; Rouphael, N.; Creech, C.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Janssen COVID-19 Vaccine. Available online: https://www.fda.gov/emergency-preparedness-and-response/coronavirus-disease-2019-covid-19/janssen-covid-19-vaccine (accessed on 23 May 2022).
- COVID-19 Vaccine AstraZeneca. Available online: https://www.ema.europa.eu/en/documents/product-information/covid-19-vaccine-astrazeneca-product-information-approved-chmp-29-january-2021-pending-endorsement_en.pdf (accessed on 23 May 2022).
- Sputnik V Vaccine. Available online: https://www.precisionvaccinations.com/vaccines/sputnik-v-vaccine#:~:text=The%20Sputnik%20V%20vaccine%20is,10*11%20particles%20per%20dose (accessed on 23 May 2022).
- Sinovac: COVID-19 Vaccine (Vero Cell Line) Inactivated. Available online: https://cdn.who.int/media/docs/default-source/immunization/sage/2021/april/4_sage29apr2021_sinovac.pdf (accessed on 23 May 2022).
- Evidence Assessment: Sinopharm/BBIBP COVID-19 Vaccine. Available online: https://cdn.who.int/media/docs/defaultsource/immunization/sage/2021/april/2_sage29apr2021_critical-evidence_sinopharm.pdf (accessed on 23 May 2022).
- Sinopharm (Vero Cell) Inactivated, COVID-19 Vaccine. Available online: https://www.who.int/publications/m/item/sinopharm-vero-cell---inactivated-covid-19-vaccine (accessed on 23 May 2022).
- COVAXIN: Factsheet for Recipients and Caregivers. Available online: https://www.bharatbiotech.com/images/covaxin/covaxin-factsheet.pdf (accessed on 23 May 2022).
- Precisionvaccinations. EpiVacCorona Vaccine. 2022. Available online: https://www.precisionvaccinations.com/vaccines/epivaccorona-vaccine#:~:text=The%20EpiVacCorona%20vaccine%20relies%20on,due%20to%20artificially%20synthesized%20peptides (accessed on 23 May 2022).
- MVC-COV1901 VACCINE PROFILE. Available online: https://cdn.who.int/media/docs/default-source/blue-print/developer_medigen_allen-lien.pdf (accessed on 23 May 2022).
- Dai, L.; Gao, L.; Tao, L.; Hadinegoro, S.; Erkin, M.; Ying, Z.; He, P.; Girsang, R.; Vergara, H.; Akram, J.; et al. Efficacy and Safety of the RBD-Dimer–Based COVID-19 Vaccine ZF2001 in Adults. N. Engl. J. Med. 2022, 386, 2097–2111. [Google Scholar] [CrossRef]
- Precisionvaccinations. Zifivax (ZF2001) COVID-19 Vaccine. 2022. Available online: https://www.precisionvaccinations.com/vaccines/zifivax-zf2001-covid-19-vaccine (accessed on 23 May 2022).
- Mediccreview. SOBERANA, Cuba’s COVID-19 Vaccine Candidates. Available online: https://mediccreview.org/soberana-cuba-covid-19-vaccine-candidates/ (accessed on 23 May 2022).
- Precisionvaccinations. CorbeVax COVID-19 Vaccine. 2022. Available online: https://www.precisionvaccinations.com/vaccines/corbevax-covid-19-vaccine (accessed on 23 May 2022).
- Czub, M.; Weingartl, H.; Czub, S.; He, R.; Cao, J. Evaluation of modified vaccinia virus Ankara based recombinant SARS vaccine in ferrets. Vaccine 2005, 23, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Greenough, T.; Babcock, G.; Roberts, A.; Hernandez, H.; Thomas, W., Jr.; Coccia, J.; Graziano, R.; Srinivasan, M.; Lowy, I.; Finberg, R.; et al. Development and Characterization of a Severe Acute Respiratory Syndrome–Associated Coronavirus–Neutralizing Human Monoclonal Antibody That Provides Effective Immunoprophylaxis in Mice. J. Infect. Dis. 2005, 191, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Tripp, R.; Haynes, L.; Moore, D.; Anderson, B.; Tamin, A.; Harcourt, B.; Jones, L.; Yilla, M.; Babcock, G.; Greenough, T.; et al. Monoclonal antibodies to SARS-associated coronavirus (SARS-CoV): Identification of neutralizing and antibodies reactive to S, N, M and E viral proteins. J. Virol. Methods 2005, 128, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Thomas, W.; Guarner, J.; Lamirande, E.; Babcock, G.; Greenough, T.; Vogel, L.; Hayes, N.; Sullivan, J.; Zaki, S.; et al. Therapy with a Severe Acute Respiratory Syndrome–Associated Coronavirus–Neutralizing Human Monoclonal Antibody Reduces Disease Severity and Viral Burden in Golden Syrian Hamsters. J. Infect. Dis. 2006, 193, 685–692. [Google Scholar] [CrossRef]
- Jiang, S.; Lu, L.; Du, L. Development of SARS vaccines and therapeutics is still needed. Future Virol. 2013, 8, 1–2. [Google Scholar] [CrossRef]
- NHS. “SARS (Severe Acute Respiratory Syndrome)”. National Health Service. 2020. Available online: https://web.archive.org/web/20200309174230/https://www.nhs.uk/conditions/sars/ (accessed on 30 January 2022).
- Song, Z.; Xu, Y.; Bao, L.; Zhang, L.; Yu, P.; Qu, Y.; Zhu, H.; Zhao, W.; Han, Y.; Qin, C. From SARS to MERS, Thrusting Coronaviruses into the Spotlight. Viruses 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Butler, D. SARS veterans tackle coronavirus. Nature 2012, 490, 20. [Google Scholar] [CrossRef] [Green Version]
- Shehata, M.M.; Gomaa, M.R.; Ali, M.A.; Kayali, G. Middle East respiratory syndrome coronavirus: A comprehensive review. Front. Med. 2016, 10, 120–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modjarrad, K.; Roberts, C.C.; Mills, K.T.; Castellano, A.R.; Paolino, K.; Muthumani, K.; Reuschel, E.L.; Robb, M.L.; Racine, T.; Oh, M.D.; et al. Safety and immunogenicity of an anti-Middle East respiratory syndrome coronavirus DNA vaccine: A phase 1, open-label, single-arm, dose-escalation trial. Lancet Infect. Dis. 2019, 19, 1013–1022. [Google Scholar] [CrossRef] [Green Version]
- Yong, C.Y.; Ong, H.K.; Yeap, S.K.; Ho, K.L.; Tan, W.S. Recent Advances in the Vaccine Development Against Middle East Respiratory Syndrome-Coronavirus. Front. Microbiol. 2019, 10, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karikó, K.; Muramatsu, H.; Keller, J.M.; Weissman, D. Increased Erythropoiesis in Mice Injected with Submicrogram Quantities of Pseudouridine-containing mRNA Encoding Erythropoietin. Mol. Ther. 2012, 20, 948–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.A.; Reesor, E.K.G.; Xu, Y.; Zope, H.R.; Zetter, B.R.; Shi, J. Biomaterials for mRNA delivery. Biomater. Sci. 2015, 3, 1519–1533. [Google Scholar] [CrossRef] [Green Version]
- Sharma, O.; Sultan, A.A.; Ding, H.; Triggle, C.R. A Review of the Progress and Challenges of Developing a Vaccine for COVID-19. Front. Immunol. 2020, 11, 585354. [Google Scholar] [CrossRef]
- Bok, K.; Sitar, S.; Graham, B.S.; Mascola, J.R. Accelerated COVID-19 vaccine development: Milestones, lessons, and prospects. Immunity 2021, 54, 1636–1651. [Google Scholar] [CrossRef]
- Turner, J.S.; O’Halloran, J.A.; Kalaidina, E.; Kim, W.; Schmitz, A.J.; Zhou, J.Q.; Lei, T.; Thapa, M.; Chen, R.E.; Case, J.B.; et al. SARS-CoV-2 mRNA vaccines induce persistent human germinal centre responses. Nature 2021, 596, 109–113. [Google Scholar] [CrossRef]
- Le, T.T.; Cramer, J.P.; Chen, R.; Mayhew, S. Evolution of the COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 667–668. [Google Scholar] [CrossRef]
- Arbeitman, C.R.; Rojas, P.; Ojeda-May, P.; Garcia, M.E. The SARS-CoV-2 spike protein is vulnerable to moderate electric fields. Nat. Commun. 2021, 12, 5407. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Dutta, N.K.; Mazumdar, K.; Gordy, J.T.; Dutch, R.E. The Nucleocapsid Protein of SARS-CoV-2: A Target for Vaccine Development. J. Virol. 2020, 94, e00647-20. [Google Scholar] [CrossRef] [PubMed]
- Nikolaidis, M.; Markoulatos, P.; Van de Peer, Y.; Oliver, S.G.; Amoutzias, G.D.; Hepp, C. The neighborhood of the Spike gene is a hotspot for modular intertypic homologous and non-homologous recombination in Coronavirus genomes. Mol. Biol. Evol. 2021, 39, msab292. [Google Scholar] [CrossRef] [PubMed]
- Thanh, L.T.; Andreadakis, Z.; Kumar, A.; Gómez Román, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef]
- Diamond, M.S.; Pierson, T.C. The Challenges of Vaccine Development against a New Virus during a Pandemic. Cell Host Microbe 2020, 27, 699–703. [Google Scholar] [CrossRef]
- McLellan, J. Structure-based design of prefusion-stabilized SARS-CoV-2 spikes. Acta Crystallogr. Sect. A Found. Adv. 2020, 76, a209. [Google Scholar] [CrossRef]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; Veen, W.; Brüggen, M.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune response to SARS-CoV-2 and mechanisms of immunopathological changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, X.; Qu, M.; Aninwene, G.; Jucaud, V.; Moon, J.; Gu, Z.; Sun, W.; Khademhosseini, A. Engineering Antiviral Vaccines. ACS Nano 2020, 14, 12370–12389. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.; Herrler, G.; Wu, N.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; LiuXijun, R.; Shuai, H.; Sun, Z.; Zhao, Y.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and rapid transmission of SARS-CoV-2 in ferrets. Cell Host Microbe 2020, 27, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stertz, S.; Reichelt, M.; Spiegel, M.; Kuri, T.; Martinez-Sobrido, L.; Garcia-Sastre, A.; Weber, F.; Kochs, G. The intracellular sites of early replication and budding of SARS-coronavirus. Virology 2007, 361, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, K.L.; Kok, K.H.; Ng, M.H.; Poon, V.K.; Yuen, K.Y.; Zheng, B.J.; Jin, D.Y. Severe acute respiratory syndrome coronavirus M protein inhibits type I interferon production by impeding the formation of TRAF3.TANK.TBK1/IKKepsilon complex. J. Biol. Chem. 2009, 284, 16202–16209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shneider, A.; Kudriavtsev, A.; Vakhrusheva, A. Can melatonin reduce the severity of COVID-19 pandemic? Int. Rev. Immunol. 2020, 39, 153–162. [Google Scholar] [CrossRef]
- Deftereos, S.G.; Siasos, G.; Giannopoulos, G.; Vrachatis, D.A.; Angelidis, C.; Giotaki, S.G.; Gargalianos, P.; Giamarellou, H.; Gogos, C.; Daikos, G.; et al. The Greek study in the effects of colchicine in COVID-19 complications prevention (GRECCO-19 study): Rationale and study design. Hellenic J. Cardiol. 2020, 61, 42–45. [Google Scholar] [CrossRef]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVID-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost Agents 2020, 34, 327–331. [Google Scholar]
- Patel, A.; Bah, M.A.; Weiner, D.B. In Vivo Delivery of Nucleic Acid-Encoded Monoclonal Antibodies. BioDrugs 2020, 34, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, I.; Aleya, L.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Bungau, S. CD147-spike protein interaction in COVID-19: Get the ball rolling with a novel receptor and therapeutic target. Sci. Total Environ. 2022, 808, 152072. [Google Scholar] [CrossRef]
- Xiong, L.; Edwards, C.K.; Zhou, L. The biological function and clinical utilization of CD147 in human diseases: A review of the current scientific literature. Int. J. Mol. Sci. 2014, 15, 17411–17441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrich, H.; Pillat, M.M. CD147 as a target for COVID-19 treatment: Suggested effects of azithromycin and stem cell engagement. Stem Cell Rev. Rep. 2020, 16, 434–440. [Google Scholar] [CrossRef]
- Jansen, J.M.; Gerlach, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J. Clin. Virol. 2019, 119, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.X.; Liu, B.Z.; Deng, H.J.; Wu, G.C.; Deng, K.; Chen, Y.K.; Liao, P.; Qiu, J.F.; Lin, Y.; Cai, X.F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Karamloo, F.; König, R. SARS-CoV-2 immunogenicity at the crossroads. Allergy 2020, 75, 1822–1824. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A sequence homology and bioinformatic approach can predict candidate targets for immune responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680. [Google Scholar] [CrossRef]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary identification of potential vaccine targets for the COVID-19 coronavirus (SARS-CoV-2) based on SARS-CoV immunological studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [Green Version]
- Ng, O.W.; Chia, A.; Tan, A.T.; Jadi, R.S.; Leong, H.N.; Bertoletti, A.; Tan, Y.J. Memory T cell responses targeting the SARS coronavirus persist up to 11 years post-infection. Vaccine 2016, 34, 2008–2014. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; De Smedt, S.C.; Dewitte, H. Three decades of messenger RNA vaccine development. Nano Today 2019, 28, 100766. [Google Scholar] [CrossRef]
- Krammer, F. SARS-CoV-2 vaccines in development. Nature 2020, 586, 516–527. [Google Scholar] [CrossRef]
- Park, K.S.; Sun, X.; Aikins, M.E.; Moon, J.J. Non-viral COVID-19 vaccine delivery systems. Adv. Drug Deliv. Rev. 2021, 169, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the messenger: Advances in technologies for therapeutic mRNA delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef] [Green Version]
- CDC 2021. COVID-19 ACIP Vaccine Recommendations. Available online: https://www.cdc.gov/vaccines/hcp/acip-recs/vacc-specific/covid-19.html (accessed on 25 February 2022).
- EC European commission. Safe COVID-19 Vaccines for Europeans. Available online: https://ec.europa.eu/info/live-work-travel-eu/coronavirus-response/safe-covid-19-vaccines-europeans_en (accessed on 25 February 2022).
- Shimabukuro, T. Allergic reactions including anaphylaxis after receipt of the first dose of Pfizer-BioNTech COVID-19 vaccine—United States, December 14–23, 2020. Am. J. Transplant. 2021, 21, 1332. [Google Scholar] [CrossRef]
- Moghimi, S.M. Allergic reactions and anaphylaxis to LNP-based COVID-19 vaccines. Mol. Ther. 2021, 29, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Batty, C.; Heise, M.; Bachelder, E.; Ainslie, K. Vaccine formulations in clinical development for the prevention of severe acute respiratory syndrome coronavirus 2 infection. Adv. Drug Deliv. Rev. 2021, 169, 168–189. [Google Scholar] [CrossRef] [PubMed]
- Hajj, K.; Whitehead, K. Tools for translation: Non-viral materials for therapeutic mRNA delivery. Nat. Rev. Mater. 2017, 2, 17056. [Google Scholar] [CrossRef]
- Schlake, T.; Thess, A.; Fotin-Mleczek, M.; Kallen, K. Developing mRNA-vaccine technologies. RNA Biol. 2012, 9, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, K.; Li, R.; Zhang, L. mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection. Int. J. Mol. Sci. 2020, 21, 6582. [Google Scholar] [CrossRef]
- Kyriakidis, N.; López-Cortés, A.; González, E.; Grimaldos, A.; Prado, E. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Mesin, L.; Ersching, J.; Victora, G.D. Germinal Center B Cell Dynamics. Immunity 2016, 45, 471–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallusto, F.; Lanzavecchia, A.; Araki, K.; Ahmed, R. From Vaccines to Memory and Back. Immunity 2010, 33, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amanna, I.J.; Carlson, N.E.; Slifka, M.K. Duration of Humoral Immunity to Common Viral and Vaccine Antigens. N. Engl. J. Med. 2007, 357, 1903–1915. [Google Scholar] [CrossRef] [Green Version]
- Robbiani, D.; Gaebler, C.; Muecksch, F.; Lorenzi, J.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Laczkó, D.; Hogan, M.; Toulmin, S.; Hicks, P.; Lederer, K.; Gaudette, B.; Castaño, D.; Amanat, F.; Muramatsu, H.; Oguin, T.H.; et al. A Single Immunization with Nucleoside-Modified mRNA Vaccines Elicits Strong Cellular and Humoral Immune Responses against SARS-CoV-2 in Mice. Immunity 2020, 53, 724–732. [Google Scholar] [CrossRef]
- Lederer, K.; Castaño, D.; Gómez Atria, D.; Oguin, T.; Wang, S.; Manzoni, T.; Muramatsu, H.; Hogan, M.; Amanat, F.; Cherubin, P.; et al. SARS-CoV-2 mRNA Vaccines Foster Potent Antigen-Specific Germinal Center Responses Associated with Neutralizing Antibody Generation. Immunity 2020, 53, 1281–1295. [Google Scholar] [CrossRef]
- Corbett, K.; Edwards, D.; Leist, S.; Abiona, O.; Boyoglu-Barnum, S.; Gillespie, R.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef]
- Vogel, A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.; Kranz, L.M.; Walzer, K.C.; Hein, S.; Güler, A.; et al. A prefusion SARS-CoV-2 spike RNA vaccine is highly immunogenic and prevents lung infection in non-human primates. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rauch, S.; Roth, N.; Schwendt, K.; Fotin-Mleczek, M.; Mueller, S.; Petsch, B. mRNA-based SARS-CoV-2 vaccine candidate CVnCoV induces high levels of virus-neutralising antibodies and mediates protection in rodents. NPJ Vaccines 2021, 6, 57. [Google Scholar] [CrossRef]
- Zhang, N.; Li, X.; Deng, Y.; Zhao, H.; Huang, Y.; Yang, G.; Huang, W.; Gao, P.; Zhou, C.; Zhang, R.; et al. A Thermostable mRNA Vaccine against COVID-19. Cell 2020, 182, 1271–1283. [Google Scholar] [CrossRef]
- Tai, W.; Zhang, X.; Drelich, A.; Shi, J.; Hsu, J.C.; Luchsinger, L.; Hillyer, C.D.; Tseng, C.-T.K.; Jiang, S.; Du, L. A Novel Receptor-Binding Domain (RBD)-Based MRNA Vaccine against SARS-CoV-2. Cell Res. 2020, 30, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fontela, C.; Dowling, W.; Funnell, S.; Gsell, P.; Riveros-Balta, A.; Albrecht, R.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Lu, G.; Tan, S.; Xia, J.; Xiong, H.; Yu, X.; Qi, Q.; Yu, X.; Li, L.; Yu, H.; et al. A COVID-19 mRNA vaccine encoding SARS-CoV-2 virus-like particles induces a strong antiviral-like immune response in mice. Cell Res. 2020, 30, 936–939. [Google Scholar] [CrossRef] [PubMed]
- Oberhardt, V.; Luxenburger, H.; Kemming, J.; Schulien, I.; Ciminski, K.; Giese, S.; Csernalabics, B.; Lang-Meli, J.; Janowska, I.; Staniek, J.; et al. Rapid and stable mobilization of CD8+ T cells by SARS-CoV-2 mRNA vaccine. Nature 2021, 597, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.; Porter, F.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Morphology, genome organization, replication, and pathogenesis of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Coronavirus Dis. 2020. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Cox, R.J.; Brokstad, K.A. Not just antibodies: B cells and T cells mediate immunity to COVID-19. Nat. Rev. Immunol. 2020, 20, 581–582. [Google Scholar] [CrossRef]
- Bettini, E.; Locci, M. SARS-CoV-2 mRNA Vaccines: Immunological Mechanism and Beyond. Vaccines 2021, 9, 147. [Google Scholar] [CrossRef]
- Mascellino, M.; Di Timoteo, F.; De Angelis, M.; Oliva, A. Overview of the Main Anti-SARS-CoV-2 Vaccines: Mechanism of Action, Efficacy and Safety. Infect. Drug Resist. 2021, 14, 3459–3476. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.; Mateus, J.; Kato, Y.; Hastie, K.; Yu, E.; Faliti, C.; Grifoni, A.; Ramirez, S.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Ren, L.; Yang, J.; Guo, L.; Feng, L.; Ma, C.; Wang, X.; Leng, Z.; Tong, X.; Zhou, W.; et al. Seroprevalence and humoral immune durability of anti-SARS-CoV-2 antibodies in Wuhan, China: A longitudinal, population-level, cross-sectional study. Lancet. 2021, 397, 1075–1084. [Google Scholar] [CrossRef]
- McNeil, M.M.; DeStefano, F. Vaccine-associated hypersensitivity. J. Allergy Clin. Immunol. 2018, 141, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- FDA 2021a. Vaccine Pfizer Data Sheet. Fact Sheet for Healthcare Providers Administrating Vaccines (Vaccination Providers) Emergency Use Authorization (EUA) of Pfizer/BioNTech COVID-19 Vaccine to Prevent Coronavirus Disease 2019 (COVID-19). 2021. Available online: https://www.fda.gov/media/144413/download (accessed on 6 February 2022).
- FDA 2021b. Vaccine Moderna Data Sheet. Fact Sheet for Healthcare Providers Administrating (EUA) of Vaccines (Vaccination Providers) Emergency Use Authorization of Moderna COVID-19 Vaccine to Prevent Coronavirus Disease 2019 (COVID-19). 2020. Available online: https://www.fda.gov/media/144637/download (accessed on 6 February 2022).
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Pai, M.; Schull, M.; Razak, F.; Maltsev, A.; Miller, K.J.; Razak, F.; Schull, M.; Schwartz, B.; Stall, N.M.; Steiner, R.; et al. Vaccine–induced prothrombotic immune thrombocytopenia (VIPIT) following AstraZeneca COVID-19 vaccination. Sci. Briefs 2020. Available online: http://covid19-sciencetable.ca/science-briefs (accessed on 13 June 2022).
- Greinacher, A.; Thiele, T.; Warkentin, T.; Weisser, K.; Kyrle, P.; Eichinger, S. Thrombotic Thrombocytopenia after ChAdOx1 nCov-19 Vaccination. N. Engl. J. Med. 2021, 384, 2092–2101. [Google Scholar] [CrossRef]
- Vilches, T.; Zhang, K.; Van Exan, R.; Langley, J.; Moghadas, S. Projecting the impact of a two-dose COVID-19 vaccination campaign in Ontario, Canada. Vaccine 2021, 39, 2360–2365. [Google Scholar] [CrossRef]
- Emary, K.R.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 VOC 202012/01 (B.1.1.7), an exploratory analysis of a randomised controlled trial. Lancet 2021, 397, 1351–1362. [Google Scholar] [CrossRef]
- Knoll, M.; Wonodi, C. Oxford–AstraZeneca COVID-19 vaccine efficacy. Lancet 2021, 397, 72–74. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against COVID-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Livingston, E.H.; Malani, P.N.; Creech, C.B. The Johnson & Johnson Vaccine for COVID-19. JAMA 2021, 325, 1575. [Google Scholar] [CrossRef] [PubMed]
- Shay, D.K.; Gee, J.; Su, J.R.; Myers, T.R.; Marquez, P.; Liu, R.; Zhang, B.; Licata, C.; Clark, T.A.; Shimabukuro, T.T. Safety Monitoring of the Janssen (Johnson & Johnson) COVID-19 Vaccine—United States, March–April 2021. MMWR 2021, 70, 680–684. [Google Scholar] [CrossRef]
- Novavax and Janssen Report Positive Phase III Results for COVID-19 Vaccine Candidates. Clin. OMICs 2021, 8, 10. [CrossRef]
- National Institute of Allergy and Infectious Disease (NIAID) and Pharmaceutical Janssen Companies from the U.S. Department of Health and Human Services’. Janssen Investigational COVID-19 Vaccine: Interim Analysis of Phase 3 Clinical Data Released. 2021. Available online: https://www.nih.gov/news-events/news-releases/janssen-investigational-covid-19-vaccine-interim-analysis-phase-3-clinical-data-released (accessed on 6 February 2022).
- Vaccine Janssen Data Sheet 2021. John Galloway (King’s College London). 2021. Available online: https://www.janssenlabels.com/emergency-use-authorization/Janssen+COVID-19+Vaccine-HCP-fact-sheet.pdf (accessed on 6 February 2022).
- Jones, I.; Roy, P. Sputnik V COVID-19 vaccine candidate appears safe and effective. Lancet 2021, 397, 642–643. [Google Scholar] [CrossRef]
- Logunov, D.; Dolzhikova, I.; Shcheblyakov, D.; Tukhvatulin, A.; Zubkova, O.; Dzharullaeva, A.; Kovyrshina, A.; Lubenets, N.; Grousova, D.; Erokhova, A.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Afshar, S.; Masoudi, M.; Gholizadeh, P.; Asgharzadeh, M.; Ganbarov, K.; Köse, Ş.; Yousefi, M.; Kafil, H. SARS-CoV-2 (COVID-19) vaccines structure, mechanisms and effectiveness: A review. Int. J. Biol. Macromol. 2021, 188, 740–750. [Google Scholar] [CrossRef]
- Grigoryan, L.; Pulendran, B. The immunology of SARS-CoV-2 infections and vaccines. Semin. Immunol. 2020, 50, 101422. [Google Scholar] [CrossRef]
- Speiser, D.; Bachmann, M. COVID-19: Mechanisms of Vaccination and Immunity. Vaccines 2020, 8, 404. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Talukder, A.; Kalita, C.; Neog, N.; Goswami, C.; Sarma, M.; Hazarika, I. A comparative analysis on the safety and efficacy of Covaxin versus other vaccines against COVID-19: A review. Z. Für Nat. C 2022. [Google Scholar] [CrossRef] [PubMed]
- Ella, R.; Reddy, S.; Blackwelder, W.; Potdar, V.; Yadav, P.; Sarangi, V.; Aileni, V.; Kanungo, S.; Rai, S.; Reddy, P.; et al. Efficacy, safety, and lot-to-lot immunogenicity of an inactivated SARS-CoV-2 vaccine (BBV152): Interim results of a randomised, double-blind, controlled, phase 3 trial. Lancet 2021, 398, 2173–2184. [Google Scholar] [CrossRef]
- Sadarangani, M.; Marchant, A.; Kollmann, T. Immunological mechanisms of vaccine-induced protection against COVID-19 in humans. Nat. Rev. Immunol. 2021, 21, 475–484. [Google Scholar] [CrossRef]
- Peng, X.; Cheng, J.; Gong, H.; Yuan, M.; Zhao, X.; Li, Z.; Wei, D. Advances in the design and development of SARS-CoV-2 vaccines. Mil. Med. Res. 2021, 8, 67. [Google Scholar] [CrossRef]
- Ryzhikov, A.; Ryzhikov, E.; Bogryantseva, M.; Danilenko, E.; Imatdinov, I.; Nechaeva, E.; Pyankov, O.; Pyankova, O.; Susloparov, I.; Taranov, O.; et al. Immunogenicity and protectivity of the peptide candidate vaccine against SARS-CoV-2. Ann. Russ. Acad. Med. Sci. 2021, 76, 5–19. [Google Scholar] [CrossRef]
- Hsieh, S.; Liu, W.; Huang, Y.; Lin, Y.; Hsieh, E.; Lian, W.; Chen, C.; Janssen, R.; Tai, I.; Chang, S. Safety and Immunogenicity of a Recombinant Stabilized Prefusion SARS-CoV-2 Spike Protein Vaccine (MVC-COV1901) Adjuvanted With CpG 1018 and Aluminum Hydroxide in Healthy Adults: A Phase 1, Dose-Escalation Study. SSRN Electron. J. 2021, 38, 100989. [Google Scholar]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef]
- Mallapaty, S. Iran hopes to defeat COVID with home-grown crop of vaccines. Nature 2021, 596, 475. [Google Scholar] [CrossRef]
- Sasaki, S.; Okuda, K. The use of conventional immunologic adjuvants in DNA vaccine preparations. In DNA Vaccines; Humana Press: Totowa, NJ, USA, 2000; pp. 241–249. [Google Scholar] [CrossRef]
- Waqas, M.; Haider, A.; Rehman, A.; Qasim, M.; Umar, A.; Sufyan, M.; Akram, H.; Mir, A.; Razzaq, R.; Rasool, D.; et al. Immunoinformatics and Molecular Docking Studies Predicted Potential Multiepitope-Based Peptide Vaccine and Novel Compounds against Novel SARS-CoV-2 through Virtual Screening. BioMed Res. Int. 2021, 2021, 1596834. [Google Scholar] [CrossRef]
- Tian, J.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef]
- Richmond, P.; Hatchuel, L.; Dong, M.; Ma, B.; Hu, B.; Smolenov, I.; Li, P.; Liang, P.; Han, H.; Liang, J.; et al. Safety and immunogenicity of S-Trimer (SCB-2019), a protein subunit vaccine candidate for COVID-19 in healthy adults: A phase 1, randomised, double-blind, placebo-controlled trial. Lancet 2021, 397, 682–694. [Google Scholar] [CrossRef]
- Kalita, P.; Padhi, A.; Zhang, K.; Tripathi, T. Design of a peptide-based subunit vaccine against novel coronavirus SARS-CoV-2. Microb. Pathog. 2020, 145, 104236. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Jakhar, R.; Sehrawat, N. Designing spike protein (S-Protein) based multi-epitope peptide vaccine against SARS COVID-19 by immunoinformatics. Heliyon 2020, 6, e05528. [Google Scholar] [CrossRef]
- Behmard, E.; Soleymani, B.; Najafi, A.; Barzegari, E. Immunoinformatic design of a COVID-19 subunit vaccine using entire structural immunogenic epitopes of SARS-CoV-2. Sci. Rep. 2020. [Google Scholar] [CrossRef]
- Machado, L.; Ottolini, B. An Evolutionary History of Defensins: A Role for Copy Number Variation in Maximizing Host Innate and Adaptive Immune Responses. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, D.; Chertov, O.; Lubkowski, J. The Structure of Human β-Defensin-1. J. Biol. Chem. 2001, 276, 39021–39026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, L.; Mburu, Y.; Mathers, A.; Fluharty, E.; Larregina, A.; Ferris, R.; Falo, L. Human Beta-Defensin 3 Induces Maturation of Human Langerhans Cell–Like Dendritic Cells: An Antimicrobial Peptide that Functions as an Endogenous Adjuvant. J. Investig. Dermatol. 2013, 133, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Schutte, B.; McCray, P. β-Defensins in Lung Host Defense. Annu. Rev. Physiol. 2002, 64, 709–748. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, Y.; Jang, S.; Jang, Y. Human β-defensin 2 plays a regulatory role in innate antiviral immunity and is capable of potentiating the induction of antigen-specific immunity. Virol. J. 2018, 15, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wørzner, K.; Sheward, D.; Schmidt, S.; Hanke, L.; Zimmermann, J.; McInerney, G.; Karlsson Hedestam, G.; Murrell, B.; Christensen, D.; Pedersen, G. Adjuvanted SARS-CoV-2 spike protein elicits neutralizing antibodies and CD4 T cell responses after a single immunization in mice. EBioMedicine 2021, 63, 103197. [Google Scholar] [CrossRef] [PubMed]
- Haun, B.; Lai, C.; Williams, C.; Wong, T.; Lieberman, M.; Pessaint, L.; Andersen, H.; Lehrer, A. CoVaccine HT™ Adjuvant Potentiates Robust Immune Responses to Recombinant SARS-CoV-2 Spike S1 Immunization. Front. Immunol. 2020, 11, 599587. [Google Scholar] [CrossRef] [PubMed]
- Bonam, S.; Partidos, C.; Halmuthur, S.; Muller, S. An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Klinman, D. Immunotherapeutic uses of CpG oligodeoxynucleotides. Nat. Rev. Immunol. 2004, 4, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Klinman, D. Adjuvant Activity of CpG Oligodeoxynucleotides. Int. Rev. Immunol. 2006, 25, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Oberemok, V.; Laikova, K.; Yurchenko, K.; Marochkin, N.; Fomochkina, I.; Kubyshkin, A. SARS-CoV-2 will constantly sweep its tracks: A vaccine containing CpG motifs in ‘lasso’ for the multi-faced virus. Inflamm. Res. 2020, 69, 801–812. [Google Scholar] [CrossRef]
- Martin, M.; VanInsberghe, D.; Koelle, K. Insights from SARS-CoV-2 sequences. Science 2021, 371, 466–467. [Google Scholar] [CrossRef]
- Di Caro, A.; Cunha, F.; Petrosillo, N.; Beeching, N.; Ergonul, O.; Petersen, E.; Koopmans, M. Severe acute respiratory syndrome coronavirus 2 escape mutants and protective immunity from natural infections or immunizations. Clin. Microbiol. Infect. 2021, 27, 823–826. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.; Bhattacharya, M.; Agoramoorthy, G.; Lee, S. Evolution, Mode of Transmission, and Mutational Landscape of Newly Emerging SARS-CoV-2 Variants. mBio 2021, 12, e01140-21. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A. Present variants of concern and variants of interest of severe acute respiratory syndrome coronavirus 2: Their significant mutations in S-glycoprotein, infectivity, re-infectivity, immune escape and vaccines activity. Rev. Med. Virol. 2021, 32, e2270. [Google Scholar] [CrossRef]
- Mohapatra, R.; Tiwari, R.; Sarangi, A.; Islam, M.; Chakraborty, C.; Dhama, K. Omicron (B.1.1.529) variant of SARS-CoV-2: Concerns, challenges, and recent updates. J. Med. Virol. 2022, 94, 2336–2342. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A. Emerging mutations in the SARS-CoV-2 variants and their role in antibody escape to small molecule-based therapeutic resistance. Curr. Opin. Pharmacol. 2022, 62, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.; Bhattacharya, M.; Lee, S. A Detailed Overview of Immune Escape, Antibody Escape, Partial Vaccine Escape of SARS-CoV-2 and Their Emerging Variants With Escape Mutations. Front. Immunol. 2022, 13, 801522. [Google Scholar] [CrossRef]
- Moore, J.; Offit, P. SARS-CoV-2 Vaccines and the Growing Threat of Viral Variants. JAMA 2021, 325, 821. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Chatterjee, S.; Sharma, A.; Agoramoorthy, G.; Chakraborty, C. D614G mutation and SARS-CoV-2: Impact on S-protein structure, function, infectivity, and immunity. Appl. Microbiol. Biotechnol. 2021, 105, 9035–9045. [Google Scholar] [CrossRef]
- Chakraborty, C.; Saha, A.; Sharma, A.; Bhattacharya, M.; Lee, S.; Agoramoorthy, G. D614G mutation eventuates in all VOI and VOC in SARS-CoV-2: Is it part of the positive selection pioneered by Darwin? Mol. Ther. Nucleic Acids 2021, 26, 237–241. [Google Scholar] [CrossRef]
- Collier, D.; De Marco, A.; Ferreira, I.; Meng, B.; Datir, R.; Walls, A.; Kemp, S.; Bassi, J.; Pinto, D.; Silacci-Fregni, C.; et al. Sensitivity of SARS-CoV-2 B.1.1.7 to mRNA vaccine-elicited antibodies. Nature 2021, 593, 136–141. [Google Scholar] [CrossRef]
- Wang, P.; Nair, M.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.; et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef]
- Geurts van Kessel, C.; Geers, D.; Schmitz, K.; Mykytyn, A.; Lamers, M.; Bogers, S.; Scherbeijn, S.; Gommers, L.; Sablerolles, R.; Nieuwkoop, N.; et al. Divergent SARS-CoV-2 Omicron–reactive T and B cell responses in COVID-19 vaccine recipients. Sci. Immunol. 2022, 7, eabo2202. [Google Scholar] [CrossRef] [PubMed]
- More Than 12.2 Billion Shots Given: COVID-19 Tracker. Available online: https://www.bloomberg.com/graphics/covid-vaccine-tracker-global-distribution/ (accessed on 24 May 2022).
- Ritchie, H.; Mathieu, E.; Rodés-Guirao, L.; Appel, C.; Giattino, C.; Ortiz-Ospina, E.; Hasell, J.; Macdonald, B.; Beltekian, D.; Roser, M. Coronavirus Pandemic (COVID-19). Our World Data. 2020. Available online: https://ourworldindata.org/covid-vaccinations (accessed on 24 May 2022).
- CDC: New Omicron Subvariant Takes Over as Dominant Coronavirus Strain. Available online: //www.usnews.com/news/national-news/articles/2022-05-24/cdc-new-omicron-subvariant-ba-2-12-1-takes-over-as-dominant-coronavirus-strain (accessed on 24 May 2022).
- Hartley, G.E.; Zelm, M.C.v.; Robinson, M.J. The benefit of boosters: Diversity and inclusion in the COVID-19 memory response. Immunol. Cell Biol. 2022, 100, 15–17. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/imcb.12511 (accessed on 24 May 2022). [CrossRef] [PubMed]

| Sr. No. | Vaccine | Composition | Antigen Used | Dose | Storage Conditions |
|---|---|---|---|---|---|
| mRNA vaccine | |||||
| 1 | Pfizer BioNTech | Lipid nanoparticle (LNP) coupled mRNA 0.43 mg ALC-0315 = (4-hydroxybutyl) azanediyl) bis (hexane-6, 1-diyl) bis(2-hexyldecanoate), 0.05 mg ALC-0159 = 2-[(polyethylene glycol)-2000]-N,N ditetradecylacetamide, 0.09 mg 1,2-Distearoyl-sn-glycero-3-phosphocholine (DSPC), 0.2 mg Cholesterol, 0.01 mg Potassium dihydrogen phosphate, 0.07 mg Disodium hydrogen phosphate dihydrate pH 7–8 [11]. | Spike (S) protein gene full length | Two shots with a gap of 21 days | −70 °C (long term) 2–8 °C up to 5 days |
| 2 | Moderna | LNP encapsulated mRNA. The core of LNPs contains mRNA, an ionizable cationic lipid, a neutral lipid and water. SM-102 (heptadecan-9-yl 8-((2-hydroxyethyl) (6-oxo-6-(undecyloxy) hexyl) amino) octanoate}, PEG2000-DMG = 1-monomethoxypolyethyleneglycol-2,3-dimyristylglycerol with polyethylene glycol of average molecular weight 2000, 1,2-Distearoyl-sn-glycero-3 phosphocholine (DSPC), Cholesterol, Tris (tromethamine) pH 7–8 [12]. | Spike (S) protein gene full length | Two shots with a gap of 28 days | −20 °C (long term) 2–8 °C up to 30 days |
| Viral vectored vaccine | |||||
| 3 | Astra—Zeneca Oxford | Adenovirus: Viral vector (Replication deficient) encoding the SARS-CoV-2 Spike glycoprotein (ChAdOx1-S) not less than 2.5 × 108 infectious units (Inf.U). Origin: Chimpanzee Adenovirus, L-Histidine, L-Histidine hydrochloride monohydrate, Magnesium chloride hexahydrate, Polysorbate 80 (E 433), Ethanol, Sucrose, Sodium chloride, Disodium edetate (dihydrate), Water for injections [13]. | Spike (S) glycoprotein in the trimeric pre-fusion conformation | Two shots with a gap of 12 weeks | 2–7 °C up to 6 months |
| 4 | Janssen Johnson & Johnson | Adenovirus: Viral vector (Replication incompetent) expressing the SARS-CoV-2 spike protein, Origin: Human Adenovirus serotype 26, Citric acid monohydrate, trisodium citrate dihydrate, ethanol, 2-hydroxypropyl-β-cyclodextrin (HBCD), polysorbate-80, sodium chloride [14]. | Spike (S) glycoprotein | Only one shot | 2–8 °C up to 3 months |
| 5 | Sputnik V | Two Recombinant Adenovirus - Human Adenovirus serotype 26 - Human Adenovirus serotype 5 Containing the SARS-CoV-2 S protein gene, in the amount of (1.0 ± 0.5) × 1011 particles per 0.5 mL dose. Tris-(hydroxymethyl)-aminomethane, Sodium chloride, Sucrose, Magnesium chloride hexahydrate, Disodium EDTA dihydrate, Polysorbate 80, Ethanol, and Water [15]. | Unmodified full-length S-protein | Two shots with a gap of 21 days | −18 °C (liquid form) −2–8 °C (dry form) |
| Inactivated vaccines | |||||
| 6 | CoronaVac | Inactivated SARS-CoV-2 Virus (CZ02 strain) propagated on Vero cell line. 600SU per 0.5 mL. Adjuvant: Aluminum hydroxide Disodium hydrogen phosphate dodecahydrate, Sodium dihydrogen phosphate monohydrate, Sodium chloride [16]. | Beta-propiolactone inactivated SARS-CoV-2 whole virus | Two shots with a gap of 14 days | 2–8 °C |
| 7 | Covaxin | Inactivated SARS-CoV-2 Virus (NIV-2020-770 strain) 6 µg of whole-virion inactivated SARS-CoV-2 Adjuvant: Aluminum hydroxide gel (250 µg) TLR 7/8 agonist (imidazoquinolinone) 15 µg, 2-phenoxyethanol 2.5 mg, Phosphate buffer saline up to 0.5 mL [17]. | Whole-virion inactivated SARS-CoV-2 antigen (Beta-propiolactone) | Two shots with a gap of 4 weeks | 2–8 °C |
| 8 | Sinopharm BBIBP-CorV | Inactivated SARS-CoV-2 Virus (WIV04 strain) propagated on Vero cell line. 600SU per 0.5 mL. Adjuvant: Aluminum hydroxide Disodium hydrogen phosphate, Sodium dihydrogen phosphate monohydrate, Sodium chloride [18]. | Beta-propiolactone inactivated SARS-CoV-2 whole virus (1:4000 vol/vol at 2 to 8 °C) for 48 h | Two shots with a gap of 21–28 days | 2–8 °C up to 6 months (No freezing) |
| Subunit vaccine | |||||
| 9 | Epivac | Chemically synthesized peptide antigens of SARS-CoV-2 Spike protein.
Adjuvant: Aluminium Hydroxide [20]. | Chemically synthesized peptide antigens of SARS-CoV-2 Spike protein | Two shots with a gap of 21–28 days | 2–8 °C |
| 10 | MVC COVID-19 vaccine | CHO cell derived spike protein (Subunit) Liquid, S-2P protein (15 mcg) + CpG 1018 (750 mcg) + Al(OH)3 (375 mcg) in 0.5 mL [21]. | S-2P protein | Two shots with a gap of 4 weeks | 2–8 °C |
| 11 | Zifivax | ZifiVax ZF2001 (ZF-UZ-VAC-2001) is a protein subunit vaccine using a dimeric form of the receptor-binding domain (RBD) as the antigen. 25 μg or 50 μg per 0·5 mL in a vial. Adjuvant: Aluminum hydroxide [22]. | SARS-CoV-2 RBD antigen (residues 319–537, accession no. YP_009724390) Two copies in tandem repeat dimeric form. Manufactured in the CHOZN CHO K1 cell line | Three shots of 25 μg with a gap of 30 days | 2–8 °C |
| 12 | CorbeVax | Spike receptor-binding domain (RBD) Adjuvant: Alhydroge (Alum) and CpG 1018 [24]. | SARS-CoV-2 Spike protein RBD N1C1 | Two shots with a gap of 28 days | 2–8 °C |
| 13 | Soberana 02 | Conjugate vaccine in which the virus antigen, the receptor-binding domain (RBD), is chemically bound to the tetanus toxoid [25]. | SARS-CoV-2 Spike protein RBD | Two shots with a gap of 28 days | 2–8 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellamkonda, N.; Lambe, U.P.; Sawant, S.; Nandi, S.S.; Chakraborty, C.; Shukla, D. Immune Response to SARS-CoV-2 Vaccines. Biomedicines 2022, 10, 1464. https://doi.org/10.3390/biomedicines10071464
Bellamkonda N, Lambe UP, Sawant S, Nandi SS, Chakraborty C, Shukla D. Immune Response to SARS-CoV-2 Vaccines. Biomedicines. 2022; 10(7):1464. https://doi.org/10.3390/biomedicines10071464
Chicago/Turabian StyleBellamkonda, Navya, Upendra Pradeep Lambe, Sonali Sawant, Shyam Sundar Nandi, Chiranjib Chakraborty, and Deepak Shukla. 2022. "Immune Response to SARS-CoV-2 Vaccines" Biomedicines 10, no. 7: 1464. https://doi.org/10.3390/biomedicines10071464
APA StyleBellamkonda, N., Lambe, U. P., Sawant, S., Nandi, S. S., Chakraborty, C., & Shukla, D. (2022). Immune Response to SARS-CoV-2 Vaccines. Biomedicines, 10(7), 1464. https://doi.org/10.3390/biomedicines10071464