
New Therapeutic Perspectives in Prostate Cancer: Patient-Derived Organoids and Patient-Derived Xenograft Models in Precision Medicine


Abstract
:1. Introduction
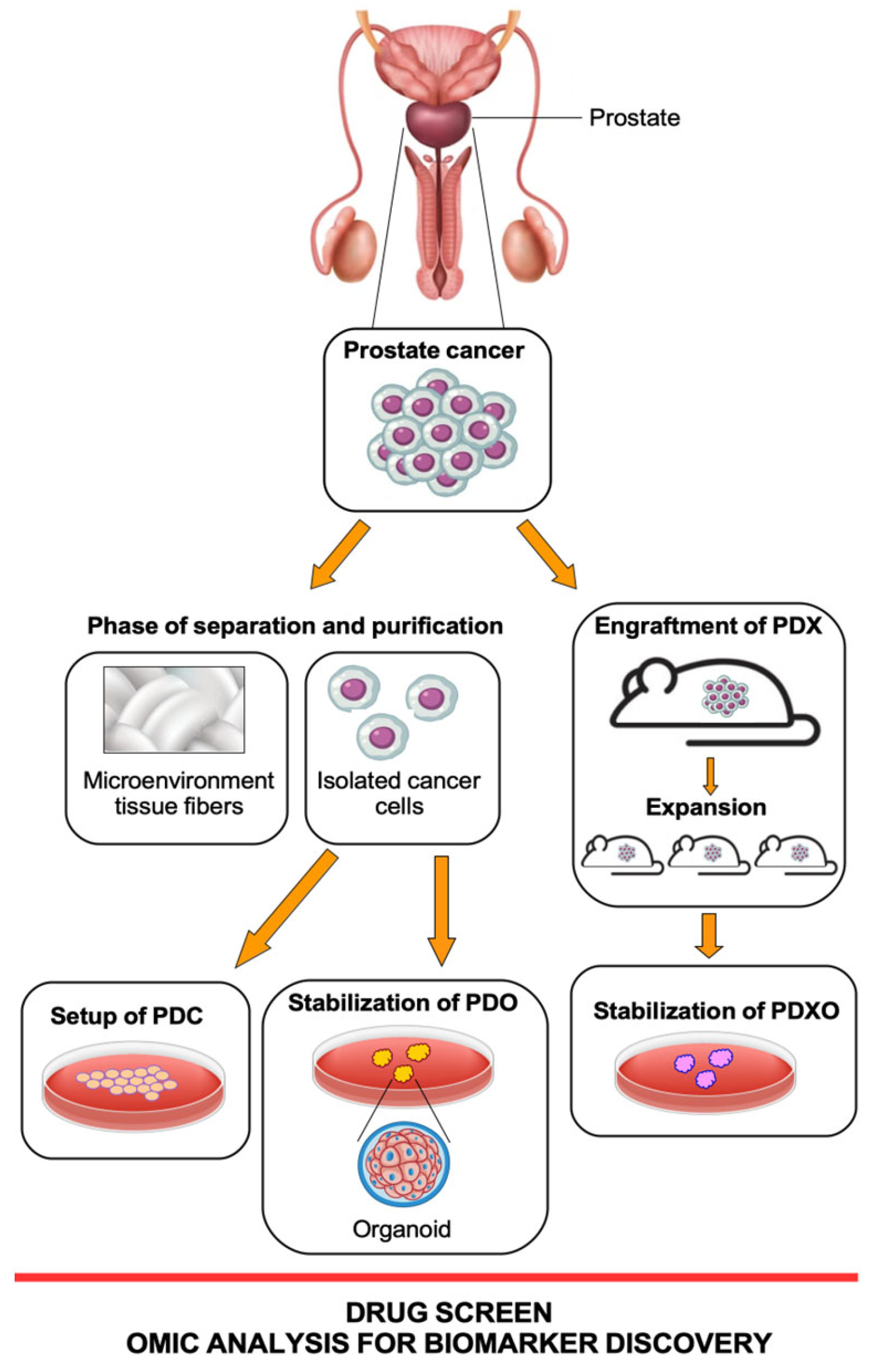
2. Patient-Derived Preclinical Models
2.1. Prostate Cancer PDX Models

{kind=link}
| Models | Benefits | Drawbacks |
|---|---|---|
| Mouse PDX-patient-derived Xenografts [12,18,19,25,26,27,28,29,34] | Possibility to develop the tumor in a physiological TME | Need for an animal house and high costs for maintaining the mice |
| Possibility to study tumor cell heterogeneity in vivo | Long time for engraftment experiments | |
| Crosstalk between factors of the murine immune system and the tumor | High failure rate in engraftment | |
| Possibility to study the response to therapies with in vivo parameters | ||
| PDO-patient-derived Organoids [20,36,37,38,39] | Limited costs for the formation and maintenance of organoids | Absence of a physiological TME |
| Formation of the organoids in a few days and possibility of amplification in more avatars in the first passages | After a few passages the organoids change the molecular characteristics of the tumor of origin | |
| Organoid ability to grow on scaffolds and mimic signaling as in physiological TME | ||
| Ability to reproduce the structure of the primary tumor tissue |
2.2. Prostate Cancer PDO Models
3. Current Application of PDO and PDX in PCa Cancer Research
4. Current Application of PDO and PDX in PCa Precision Medicine
5. Personalized Drug Screening
6. Drug Resistance
7. Biomarker Discovery
8. Application in Clinical Practice
9. General Challenges Addressed in the PDX and PDO Models and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Luining, W.I.; Cysouw, M.C.F.; Meijer, D.; Hendrikse, N.H.; Boellaard, R.; Vis, A.N.; Oprea-Lager, D.E. Targeting PSMA Revolutionizes the Role of Nuclear Medicine in Diagnosis and Treatment of Prostate Cancer. Cancers 2022, 14, 1169. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, S.; Moore, C.M.; Chiong, E.; Beltran, H.; Bristow, R.G.; Williams, S.G. Prostate Cancer. Lancet 2021, 398, 1075–1090. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, A.; Haskins, C.; Siddiqui, M.M.; Hussain, A.; D’Adamo, C. The Evolving Role of Diet in Prostate Cancer Risk and Progression. Curr. Opin. Oncol. 2019, 31, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi-Nejad, H.; Simmons, M.; Dakwar, G.; Dogra, V. Controversies in Transrectal Ultrasonography and Prostate Biopsy. Ultrasound Q. 2006, 22, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.J.; Pilcher, J.; Richenberg, J.; Patel, U.; Frauscher, F. Applications of transrectal ultrasound in prostate cancer. Br. J. Radiol. 2012, 85, S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Sivaraman, A.; Bhat, K.R.S. Screening and Detection of Prostate Cancer-Review of Literature and Current Perspective. Indian J. Surg. Oncol. 2017, 8, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Wilt, T.J.; Jones, K.M.; Barry, M.J.; Andriole, G.L.; Culkin, D.; Wheeler, T.; Aronson, W.J.; Brawer, M.K. Follow-up of Prostatectomy versus Observation for Early Prostate Cancer. N. Engl. J. Med. 2017, 377, 132–142. [Google Scholar] [CrossRef]
- Loriot, Y.; Massard, C.; Fizazi, K. Recent developments in treatments targeting castration-resistant prostate cancer bone metastases. Ann. Oncol. 2012, 23, 1085–1094. [Google Scholar] [CrossRef]
- Nuhn, P.; De Bono, J.S.; Fizazi, K.; Freedland, S.J.; Grilli, M.; Kantoff, P.W.; Sonpavde, G.; Sternberg, C.N.; Yegnasubramanian, S.; Antonarakis, E.S. Update on Systemic Prostate Cancer Therapies: Management of Metastatic Castration-Resistant Prostate Cancer in the Era of Precision Oncology. Eur. Urol. 2019, 75, 88–99. [Google Scholar] [CrossRef]
- Mateo, J.; McKay, R.; Abida, W.; Aggarwal, R.; Alumkal, J.; Alva, A.; Feng, F.; Gao, X.; Graff, J.; Hussain, M.; et al. Accelerating Precision Medicine in Metastatic Prostate Cancer. Nat. Cancer 2020, 1, 1041–1053. [Google Scholar] [CrossRef]
- Sobel, R.E.; Sadar, M.D. Cell lines used in prostate cancer research: A compendium of old and new lines—Part 1. J. Urol. 2005, 173, 342–359. [Google Scholar] [CrossRef] [PubMed]
- Risbridger, G.P.; Lawrence, M.G.; Taylor, R.A. PDX: Moving Beyond Drug Screening to Versatile Models for Research Discovery. J. Endocr. Soc. 2020, 4, bvaa132. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Brehm, M.A.; Garcia-Martinez, J.V.; Greiner, D.L. Humanized Mice for Immune System Investigation: Progress, Promise and Challenges. Nat. Rev. Immunol. 2012, 12, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Toivanen, R.; Berman, D.M.; Wang, H.; Pedersen, J.; Frydenberg, M.; Meeker, A.K.; Ellem, S.J.; Risbridger, G.P.; Taylor, R.A. Brief Report: A Bioassay to Identify Primary Human Prostate Cancer Repopulating Cells. Stem Cells 2011, 29, 1310–1314. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Wang, Y.; Lin, D.; Wang, Y. Patient-derived xenograft models of neuroendocrine prostate cancer. Cancer Lett. 2022, 525, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Beshiri, M.; Agarwal, S.; Yin, J.J.; Kelly, K. Prostate Organoids: Emerging Experimental Tools for Translational Research. J. Clin. Investig. 2023, 133, e169616. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, K.K.; Cattaneo, C.M.; Weeber, F.; Chalabi, M.; van de Haar, J.; Fanchi, L.F.; Slagter, M.; van der Velden, D.L.; Kaing, S.; Kelderman, S.; et al. Generation of Tumor-Reactive T Cells by Co-culture of Peripheral Blood Lymphocytes and Tumor Organoids. Cell 2018, 174, 1586–1598.e12. [Google Scholar] [CrossRef] [PubMed]
- Tentler, J.J.; Tan, A.C.; Weekes, C.D.; Jimeno, A.; Leong, S.; Pitts, T.M.; Arcaroli, J.J.; Messersmith, W.A.; Eckhardt, S.G. Patient-derived tumour xenografts as models for oncology drug development. Nat. Rev. Clin. Oncol. 2012, 9, 338–350. [Google Scholar] [CrossRef]
- Cassidy, J.W.; Caldas, C.; Bruna, A. Maintaining Tumor Heterogeneity in Patient-Derived Tumor Xenografts. Cancer Res. 2015, 75, 2963–2968. [Google Scholar] [CrossRef]
- Beshiri, M.L.; Tice, C.M.; Tran, C.; Nguyen, H.M.; Sowalsky, A.G.; Agarwal, S.; Jansson, K.H.; Yang, Q.; McGowen, K.M.; Yin, J.; et al. A PDX/Organoid Biobank of Advanced Prostate Cancers Captures Genomic and Phenotypic Heterogeneity for Disease Modeling and Therapeutic Screening. Clin. Cancer Res. 2018, 24, 4332–4345. [Google Scholar] [CrossRef]
- Shi, C.; Chen, X.; Tan, D. Development of Patient-Derived Xenograft Models of Prostate Cancer for Maintaining Tumor Heterogeneity. Transl. Androl. Urol. 2019, 8, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Mattie, M.; Christensen, A.; Chang, M.S.; Yeh, W.; Said, S.; Shostak, Y.; Capo, L.; Verlinsky, A.; An, Z.; Joseph, I.; et al. Molecular Characterization of Patient-Derived Human Pancreatic Tumor Xenograft Models for Preclinical and Translational Development of Cancer Therapeutics. Neoplasia 2013, 15, 1138–1150. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Gao, S.; Liu, R.; Shen, J.; Shi, X.; Bai, S.; Wang, H.; Zheng, K.; Shao, Z.; Liang, C.; et al. Oncological and Genetic Factors Impacting PDX Model Construction with NSG Mice in Pancreatic Cancer. FASEB J. 2019, 33, 873–884. [Google Scholar] [CrossRef]
- Blomme, A.; Van Simaeys, G.; Doumont, G.; Costanza, B.; Bellier, J.; Otaka, Y.; Sherer, F.; Lovinfosse, P.; Boutry, S.; Palacios, A.P.; et al. Murine Stroma Adopts a Human-like Metabolic Phenotype in the PDX Model of Colorectal Cancer and Liver Metastases. Oncogene 2018, 37, 1237–1250. [Google Scholar] [CrossRef]
- Stripecke, R.; Münz, C.; Schuringa, J.J.; Bissig, K.-D.; Soper, B.; Meeham, T.; Yao, L.-C.; Di Santo, J.P.; Brehm, M.; Rodriguez, E.; et al. Innovations, Challenges, and Minimal Information for Standardization of Humanized Mice. EMBO Mol. Med. 2020, 12, e8662. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Vaeteewoottacharn, K.; Kariya, R. Application of Highly Immunocompromised Mice for the Establishment of Patient-Derived Xenograft (PDX) Models. Cells 2019, 8, 889. [Google Scholar] [CrossRef]
- Nguyen, L.C.; Naulaerts, S.; Bruna, A.; Ghislat, G.; Ballester, P.J. Predicting Cancer Drug Response In Vivo by Learning an Optimal Feature Selection of Tumour Molecular Profiles. Biomedicines 2021, 9, 1319. [Google Scholar] [CrossRef]
- Gao, H.; Korn, J.M.; Ferretti, S.; Monahan, J.E.; Wang, Y.; Singh, M.; Zhang, C.; Schnell, C.; Yang, G.; Zhang, Y.; et al. High-Throughput Screening Using Patient-Derived Tumor Xenografts to Predict Clinical Trial Drug Response. Nat. Med. 2015, 21, 1318–1325. [Google Scholar] [CrossRef]
- Pompili, L.; Porru, M.; Caruso, C.; Biroccio, A.; Leonetti, C. Patient-Derived Xenografts: A Relevant Preclinical Model for Drug Development. J. Exp. Clin. Cancer Res. 2016, 35, 189. [Google Scholar] [CrossRef]
- Ledford, H. US Cancer Institute to Overhaul Tumour Cell Lines. Nature 2016, 530, 391. [Google Scholar] [CrossRef]
- Zhao, Y.; Shuen, T.W.H.; Toh, T.B.; Chan, X.Y.; Liu, M.; Tan, S.Y.; Fan, Y.; Yang, H.; Lyer, S.G.; Bonney, G.K.; et al. Development of a New Patient-Derived Xenograft Humanized Mouse Model to Study Human-Specific Tumour Microenvironment and Immunotherapy. Gut 2018, 67, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Buqué, A.; Galluzzi, L. Modeling Tumor Immunology and Immunotherapy in Mice. Trends Cancer 2018, 4, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, U.; Ha, G.; Tseng, Y.-Y.; Greenwald, N.F.; Oh, C.; Shih, J.; McFarland, J.M.; Wong, B.; Boehm, J.S.; Beroukhim, R.; et al. Patient-Derived Xenografts Undergo Mouse-Specific Tumor Evolution. Nat. Genet. 2017, 49, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, N.; Yang, J.; Shepherd, P.D.A.; Li-Ning-Tapia, E.M.; Labanca, E.; Manyam, G.C.; Ravoori, M.K.; Kundra, V.; Araujo, J.C.; Efstathiou, E.; et al. The MD Anderson Prostate Cancer Patient-Derived Xenograft Series (MDA PCa PDX) Captures the Molecular Landscape of Prostate Cancer and Facilitates Marker-Driven Therapy Development. Clin. Cancer Res. 2020, 26, 4933–4946. [Google Scholar] [CrossRef]
- Béraud, C.; Bidan, N.; Lassalle, M.; Lang, H.; Lindner, V.; Krucker, C.; Masliah-Planchon, J.; Potiron, E.; Lluel, P.; Massfelder, T.; et al. A New Tumorgraft Panel to Accelerate Precision Medicine in Prostate Cancer. Front. Oncol. 2023, 13, 1130048. [Google Scholar] [CrossRef] [PubMed]
- Cacciatore, A.; Albino, D.; Catapano, C.V.; Carbone, G.M. Preclinical Models of Neuroendocrine Prostate Cancer. Curr. Protoc. 2023, 3, e742. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Vela, I.; Sboner, A.; Iaquinta, P.J.; Karthaus, W.R.; Gopalan, A.; Dowling, C.; Wanjala, J.N.; Undvall, E.A.; Arora, V.K.; et al. Organoid Cultures Derived from Patients with Advanced Prostate Cancer. Cell 2014, 159, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Drost, J.; Karthaus, W.R.; Gao, D.; Driehuis, E.; Sawyers, C.L.; Chen, Y.; Clevers, H. Organoid Culture Systems for Prostate Epithelial and Cancer Tissue. Nat. Protoc. 2016, 11, 347–358. [Google Scholar] [CrossRef]
- Lang, S.H.; Stark, M.; Collins, A.; Paul, A.B.; Stower, M.J.; Maitland, N.J. Experimental Prostate Epithelial Morphogenesis in Response to Stroma and Three-Dimensional Matrigel Culture. Cell Growth Differ. 2001, 12, 631–640. [Google Scholar]
- Servant, R.; Garioni, M.; Vlajnic, T.; Blind, M.; Pueschel, H.; Müller, D.C.; Zellweger, T.; Templeton, A.J.; Garofoli, A.; Maletti, S.; et al. Prostate Cancer Patient-Derived Organoids: Detailed Outcome from a Prospective Cohort of 81 Clinical Specimens. J. Pathol. 2021, 254, 543–555. [Google Scholar] [CrossRef]
- Puca, L.; Bareja, R.; Prandi, D.; Shaw, R.; Benelli, M.; Karthaus, W.R.; Hess, J.; Sigouros, M.; Donoghue, A.; Kossai, M.; et al. Patient Derived Organoids to Model Rare Prostate Cancer Phenotypes. Nat. Commun. 2018, 9, 2404. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Liu, R.; Bi, H.; Xu, C.; Rosenholm, J.M.; Åkerfelt, M. 3D Modeling of Epithelial Tumors-The Synergy between Materials Engineering, 3D Bioprinting, High-Content Imaging, and Nanotechnology. Int. J. Mol. Sci. 2021, 22, 6225. [Google Scholar] [CrossRef] [PubMed]
- Eder, T.; Eder, I.E. 3D Hanging Drop Culture to Establish Prostate Cancer Organoids. Methods Mol. Biol. 2017, 1612, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Weinstein, H.N.W.; Allegakoen, P.; Wadsworth, M.H.; Xie, J.; Yang, H.; Castro, E.A.; Lu, K.L.; Stohr, B.A.; Feng, F.Y.; et al. Single-Cell Analysis of Human Primary Prostate Cancer Reveals the Heterogeneity of Tumor-Associated Epithelial Cell States. Nat. Commun. 2022, 13, 141. [Google Scholar] [CrossRef] [PubMed]
- Stoyanova, T.; Cooper, A.R.; Drake, J.M.; Liu, X.; Armstrong, A.J.; Pienta, K.J.; Zhang, H.; Kohn, D.B.; Huang, J.; Witte, O.N.; et al. Prostate Cancer Originating in Basal Cells Progresses to Adenocarcinoma Propagated by Luminal-like Cells. Proc. Natl. Acad. Sci. USA 2013, 110, 20111–20116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.A.; Mitrofanova, A.; Bergren, S.K.; Abate-Shen, C.; Cardiff, R.D.; Califano, A.; Shen, M.M. Lineage analysis of basal epithelial cells reveals their unexpected plasticity and supports a cell-of-origin model for prostate cancer heterogeneity. Nat. Cell Biol. 2013, 15, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Karthaus, W.R.; Iaquinta, P.J.; Drost, J.; Gracanin, A.; van Boxtel, R.; Wongvipat, J.; Dowling, C.M.; Gao, D.; Begthel, H.; Sachs, N.; et al. Identification of Multipotent Luminal Progenitor Cells in Human Prostate Organoid Cultures. Cell 2014, 159, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.; Yeleswarapu, S.; Heinrich, M.A.; Prakash, J.; Pati, F. Mimicking Tumor Microenvironment by 3D Bioprinting: 3D Cancer Modeling. Biofabrication 2022, 14, 032002. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, C.; Zhang, Y.; Shi, C. Application of Organoid Models in Prostate Cancer Research. Front. Oncol. 2021, 11, 736431. [Google Scholar] [CrossRef]
- Go, Y.-H.; Choi, W.H.; Bae, W.J.; Jung, S.-I.; Cho, C.-H.; Lee, S.A.; Park, J.S.; Ahn, J.M.; Kim, S.W.; Lee, K.J.; et al. Modeling Pancreatic Cancer with Patient-Derived Organoids Integrating Cancer-Associated Fibroblasts. Cancers 2022, 14, 2077. [Google Scholar] [CrossRef]
- Silva-Pedrosa, R.; Salgado, A.J.; Ferreira, P.E. Revolutionizing Disease Modeling: The Emergence of Organoids in Cellular Systems. Cells 2023, 12, 930. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-M.; Cieślik, M.; Lonigro, R.J.; Vats, P.; Reimers, M.A.; Cao, X.; Ning, Y.; Wang, L.; Kunju, L.P.; de Sarkar, N.; et al. Inactivation of CDK12 Delineates a Distinct Immunogenic Class of Advanced Prostate Cancer. Cell 2018, 173, 1770–1782.e14. [Google Scholar] [CrossRef] [PubMed]
- Perera, M.P.J.; Thomas, P.B.; Risbridger, G.P.; Taylor, R.; Azad, A.; Hofman, M.S.; Williams, E.D.; Vela, I. Chimeric Antigen Receptor T-Cell Therapy in Metastatic Castrate-Resistant Prostate Cancer. Cancers 2022, 14, 503. [Google Scholar] [CrossRef] [PubMed]
- Karbach, J.; Kiselicki, D.; Brand, K.; Wahle, C.; Sinelnikov, E.; Gustavus, D.; Hoffmeister, H.; Prisack, H.-B.; Atmaca, A.; Jäger, E. Tumor-Infiltrating Lymphocytes Mediate Complete and Durable Remission in a Patient with NY-ESO-1 Expressing Prostate Cancer. J. Immunother. Cancer 2023, 11, e005847. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Zaidi, S.; Love, J.R.; Zhao, J.L.; Setty, M.; Wadosky, K.M.; Gopalan, A.; Choo, Z.-N.; Persad, S.; Choi, J.; et al. Lineage Plasticity in Prostate Cancer Depends on JAK/STAT Inflammatory Signaling. Science 2022, 377, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.I.; Decker, S.; Zaharevitz, D.; Rubinstein, L.V.; Venditti, J.M.; Schepartz, S.; Kalyandrug, S.; Christian, M.; Arbuck, S.; Hollingshead, M.; et al. Relationships between Drug Activity in NCI Preclinical in Vitro and in Vivo Models and Early Clinical Trials. Br. J. Cancer 2001, 84, 1424–1431. [Google Scholar] [CrossRef] [PubMed]
- Tosca, E.M.; Ronchi, D.; Facciolo, D.; Magni, P. Replacement, Reduction, and Refinement of Animal Experiments in Anticancer Drug Development: The Contribution of 3D In Vitro Cancer Models in the Drug Efficacy Assessment. Biomedicines 2023, 11, 1058. [Google Scholar] [CrossRef]
- Stewart, E.L.; Tsao, M.S. Modeling Drug Resistance in PDX Models. In Patient-Derived Xenograft Models of Human Cancer; Wang, Y., Lin, D., Gout, P.W., Eds.; Molecular and Translational Medicine; Springer International Publishing: Cham, Switzerland, 2017; pp. 115–126. ISBN 978-3-319-55825-7. [Google Scholar]
- Lin, D.; Xue, H.; Wang, Y.; Wu, R.; Watahiki, A.; Dong, X.; Cheng, H.; Wyatt, A.W.; Collins, C.C.; Gout, P.W.; et al. Next Generation Patient-Derived Prostate Cancer Xenograft Models. Asian J. Androl. 2014, 16, 407–412. [Google Scholar] [CrossRef]
- Risbridger, G.P.; Clark, A.K.; Porter, L.H.; Toivanen, R.; Bakshi, A.; Lister, N.L.; Pook, D.; Pezaro, C.J.; Sandhu, S.; Keerthikumar, S.; et al. The MURAL Collection of Prostate Cancer Patient-Derived Xenografts Enables Discovery through Preclinical Models of Uro-Oncology. Nat. Commun. 2021, 12, 5049. [Google Scholar] [CrossRef]
- Driehuis, E.; van Hoeck, A.; Moore, K.; Kolders, S.; Francies, H.E.; Gulersonmez, M.C.; Stigter, E.C.A.; Burgering, B.; Geurts, V.; Gracanin, A.; et al. Pancreatic Cancer Organoids Recapitulate Disease and Allow Personalized Drug Screening. Proc. Natl. Acad. Sci. USA 2019, 116, 26580–26590. [Google Scholar] [CrossRef]
- Pamarthy, S.; Sabaawy, H.E. Patient Derived Organoids in Prostate Cancer: Improving Therapeutic Efficacy in Precision Medicine. Mol. Cancer 2021, 20, 125. [Google Scholar] [CrossRef]
- Karkampouna, S.; La Manna, F.; Benjak, A.; Kiener, M.; De Menna, M.; Zoni, E.; Grosjean, J.; Klima, I.; Garofoli, A.; Bolis, M.; et al. Patient-Derived Xenografts and Organoids Model Therapy Response in Prostate Cancer. Nat. Commun. 2021, 12, 1117. [Google Scholar] [CrossRef] [PubMed]
- Beardsley, E.K.; Hotte, S.J.; North, S.; Ellard, S.L.; Winquist, E.; Kollmannsberger, C.; Mukherjee, S.D.; Chi, K.N. A Phase II Study of Sorafenib in Combination with Bicalutamide in Patients with Chemotherapy-Naive Castration Resistant Prostate Cancer. Investig. New Drugs 2012, 30, 1652–1659. [Google Scholar] [CrossRef] [PubMed]
- Zurita, A.J.; George, D.J.; Shore, N.D.; Liu, G.; Wilding, G.; Hutson, T.E.; Kozloff, M.; Mathew, P.; Harmon, C.S.; Wang, S.L.; et al. Sunitinib in Combination with Docetaxel and Prednisone in Chemotherapy-Naive Patients with Metastatic, Castration-Resistant Prostate Cancer: A Phase 1/2 Clinical Trial. Ann. Oncol. 2012, 23, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Van Hemelryk, A.; Erkens-Schulze, S.; Lim, L.; de Ridder, C.M.A.; Stuurman, D.C.; Jenster, G.W.; van Royen, M.E.; van Weerden, W.M. Viability Analysis and High-Content Live-Cell Imaging for Drug Testing in Prostate Cancer Xenograft-Derived Organoids. Cells 2023, 12, 1377. [Google Scholar] [CrossRef] [PubMed]
- Tiriac, H.; Belleau, P.; Engle, D.D.; Plenker, D.; Deschênes, A.; Somerville, T.D.D.; Froeling, F.E.M.; Burkhart, R.A.; Denroche, R.E.; Jang, G.-H.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov. 2018, 8, 1112–1129. [Google Scholar] [CrossRef] [PubMed]
- Pappas, K.J.; Choi, D.; Sawyers, C.L.; Karthaus, W.R. Prostate Organoid Cultures as Tools to Translate Genotypes and Mutational Profiles to Pharmacological Responses. J. Vis. Exp. 2019, e60346. [Google Scholar] [CrossRef]
- Fong, E.L.S.; Martinez, M.; Yang, J.; Mikos, A.G.; Navone, N.M.; Harrington, D.A.; Farach-Carson, M.C. Hydrogel-Based 3D Model of Patient-Derived Prostate Xenograft Tumors Suitable for Drug Screening. Mol. Pharm. 2014, 11, 2040–2050. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Vessella, R.L.; Morrissey, C.; Brown, L.G.; Coleman, I.M.; Higano, C.S.; Mostaghel, E.A.; Zhang, X.; True, L.D.; Lam, H.-M.; et al. LuCaP Prostate Cancer Patient-Derived Xenografts Reflect the Molecular Heterogeneity of Advanced Disease and Serve as Models for Evaluating Cancer Therapeutics. Prostate 2017, 77, 654–671. [Google Scholar] [CrossRef]
- Zhou, Q.; Facciponte, J.; Jin, M.; Shen, Q.; Lin, Q. Humanized NOD-SCID IL2rg–/– Mice as a Preclinical Model for Cancer Research and Its Potential Use for Individualized Cancer Therapies. Cancer Lett. 2014, 344, 13–19. [Google Scholar] [CrossRef]
- Craft, N.; Shostak, Y.; Carey, M.; Sawyers, C.L. A Mechanism for Hormone-Independent Prostate Cancer through Modulation of Androgen Receptor Signaling by the HER-2/Neu Tyrosine Kinase. Nat. Med. 1999, 5, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Molife, L.R.; Omlin, A.; Jones, R.J.; Karavasilis, V.; Bloomfield, D.; Lumsden, G.; Fong, P.C.; Olmos, D.; O’Sullivan, J.M.; Pedley, I.; et al. Randomized Phase II Trial of Nintedanib, Afatinib and Sequential Combination in Castration-Resistant Prostate Cancer. Future Oncol. 2014, 10, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Ingersoll, M.A.; Lin, M.-F. ErbB-2 Signaling in Advanced Prostate Cancer Progression and Potential Therapy. Endocr. Relat. Cancer 2019, 26, R195–R209. [Google Scholar] [CrossRef] [PubMed]
- Gil, V.; Miranda, S.; Riisnaes, R.; Gurel, B.; D’Ambrosio, M.; Vasciaveo, A.; Crespo, M.; Ferreira, A.; Brina, D.; Troiani, M.; et al. HER3 Is an Actionable Target in Advanced Prostate Cancer. Cancer Res. 2021, 81, 6207–6218. [Google Scholar] [CrossRef] [PubMed]
- Namekawa, T.; Ikeda, K.; Horie-Inoue, K.; Inoue, S. Application of Prostate Cancer Models for Preclinical Study: Advantages and Limitations of Cell Lines, Patient-Derived Xenografts, and Three-Dimensional Culture of Patient-Derived Cells. Cells 2019, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Yang, K.; Wang, Y.Z.; Lin, D. TMEM45B Is a Novel Predictive Biomarker for Prostate Cancer Progression and Metastasis. Neoplasma 2018, 65, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Sowalsky, A.G.; Figueiredo, I.; Lis, R.T.; Coleman, I.; Gurel, B.; Bogdan, D.; Yuan, W.; Russo, J.W.; Bright, J.R.; Whitlock, N.C.; et al. Assessment of Androgen Receptor Splice Variant-7 as a Biomarker of Clinical Response in Castration-Sensitive Prostate Cancer. Clin. Cancer Res. 2022, 28, 3509–3525. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Terada, N.; Fujii, M.; Nagata, Y.; Nakahara, K.; Mukai, S.; Okasho, K.; Kamiyama, Y.; Akamatsu, S.; Kobayashi, T.; et al. Identification of the A2 Chain of Interleukin-13 Receptor as a Potential Biomarker for Predicting Castration Resistance of Prostate Cancer Using Patient-Derived Xenograft Models. Cancer Rep. 2023, 6, e1701. [Google Scholar] [CrossRef]
- Ryniawec, J.M.; Coope, M.R.; Loertscher, E.; Bageerathan, V.; de Oliveira Pessoa, D.; Warfel, N.A.; Cress, A.E.; Padi, M.; Rogers, G.C. GLUT3/SLC2A3 Is an Endogenous Marker of Hypoxia in Prostate Cancer Cell Lines and Patient-Derived Xenograft Tumors. Diagnostics 2022, 12, 676. [Google Scholar] [CrossRef]
- Scher, H.I.; Heller, G. Clinical States in Prostate Cancer: Toward a Dynamic Model of Disease Progression. Urology 2000, 55, 323–327. [Google Scholar] [CrossRef]
- Nakazawa, M.; Kyprianou, N. Epithelial-Mesenchymal-Transition Regulators in Prostate Cancer: Androgens and Beyond. J. Steroid Biochem. Mol. Biol. 2017, 166, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Pantel, K.; Hille, C.; Scher, H.I. Circulating Tumor Cells in Prostate Cancer: From Discovery to Clinical Utility. Clin. Chem. 2019, 65, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Welch, H.G.; Albertsen, P.C. Reconsidering Prostate Cancer Mortality—The Future of PSA Screening. N. Engl. J. Med. 2020, 382, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, D.T.; Sequist, L.V.; Lee, R.J. Circulating Tumour Cells-Monitoring Treatment Response in Prostate Cancer. Nat. Rev. Clin. Oncol. 2014, 11, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Pastushenko, I.; Brisebarre, A.; Sifrim, A.; Fioramonti, M.; Revenco, T.; Boumahdi, S.; Van Keymeulen, A.; Brown, D.; Moers, V.; Lemaire, S.; et al. Identification of the tumour transition states occurring during EMT. Nature 2018, 556, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Garg, M. Emerging Roles of Epithelial-Mesenchymal Plasticity in Invasion-Metastasis Cascade and Therapy Resistance. Cancer Metastasis Rev. 2022, 41, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Blick, T.; Thompson, E.W.; Williams, E.D. Diversity of Epithelial-Mesenchymal Phenotypes in Circulating Tumour Cells from Prostate Cancer Patient-Derived Xenograft Models. Cancers 2021, 13, 2750. [Google Scholar] [CrossRef]
- Hassan, S.; Blick, T.; Wood, J.; Thompson, E.W.; Williams, E.D. Circulating Tumour Cells Indicate the Presence of Residual Disease Post-Castration in Prostate Cancer Patient-Derived Xenograft Models. Front. Cell Dev. Biol. 2022, 10, 858013. [Google Scholar] [CrossRef]
- Wensink, G.E.; Elias, S.G.; Mullenders, J.; Koopman, M.; Boj, S.F.; Kranenburg, O.W.; Roodhart, J.M.L. Patient-derived organoids as a predictive biomarker for treatment response in cancer patients. NPJ Precis. Oncol. 2021, 5, 30. [Google Scholar] [CrossRef]
- Lee, J.; Bogyo, M. Target deconvolution techniques in modern phenotypic profiling. Curr. Opin. Chem. Biol. 2013, 17, 118–126. [Google Scholar] [CrossRef]
- Ravikumar, B.; Aittokallio, T. Improving the Efficacy-Safety Balance of Polypharmacology in Multi-Target Drug Discovery. Expert Opin. Drug Discov. 2018, 13, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Podaza, E.; Kuo, H.-H.; Nguyen, J.; Elemento, O.; Martin, M.L. Next Generation Patient Derived Tumor Organoids. Transl. Res. 2022, 250, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Rausch, M.; Iqbal, N.; Pathak, S.; Owston, H.E.; Ganguly, P. Organoid Models and Next-Generation Sequencing for Bone Marrow and Related Disorders. Organoids 2023, 2, 123–139. [Google Scholar] [CrossRef]
- Zoetemelk, M.; Ramzy, G.M.; Rausch, M.; Koessler, T.; van Beijnum, J.R.; Weiss, A.; Mieville, V.; Piersma, S.R.; de Haas, R.R.; Delucinge-Vivier, C.; et al. Optimized low-dose combinatorial drug treatment boosts selectivity and efficacy of colorectal carcinoma treatment. Mol. Oncol. 2020, 14, 2894–2919. [Google Scholar] [CrossRef] [PubMed]
- Ramzy, G.M.; Norkin, M.; Koessler, T.; Voirol, L.; Tihy, M.; Hany, D.; McKee, T.; Ris, F.; Buchs, N.; Docquier, M.; et al. Platform Combining Statistical Modeling and Patient-Derived Organoids to Facilitate Personalized Treatment of Colorectal Carcinoma. J. Exp. Clin. Cancer Res. 2023, 42, 79. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.J.; Decker, B.; Roberts, E.A.; Horowitz, N.S.; Muto, M.G.; Worley, M.J.; Feltmate, C.M.; Nucci, M.R.; Swisher, E.M.; Nguyen, H.; et al. Prediction of DNA Repair Inhibitor Response in Short-Term Patient-Derived Ovarian Cancer Organoids. Cancer Discov. 2018, 8, 1404–1421. [Google Scholar] [CrossRef]
- Ooft, S.N.; Weeber, F.; Schipper, L.; Dijkstra, K.K.; McLean, C.M.; Kaing, S.; van de Haar, J.; Prevoo, W.; van Werkhoven, E.; Snaebjornsson, P.; et al. Prospective Experimental Treatment of Colorectal Cancer Patients Based on Organoid Drug Responses. ESMO Open 2021, 6, 100103. [Google Scholar] [CrossRef] [PubMed]
- Weng, G.; Tao, J.; Liu, Y.; Qiu, J.; Su, D.; Wang, R.; Luo, W.; Zhang, T. Organoid: Bridging the Gap between Basic Research and Clinical Practice. Cancer Lett. 2023, 572, 216353. [Google Scholar] [CrossRef]

| Models | Tissues/Cells | Notes | References |
|---|---|---|---|
| PDO | Human biopsy samples and circulating tumor cells | The engraftment percentage was 15–20%. Characterization of PCa subtypes. | [37] |
| PDO | Human biopsy samples | The engraftment percentage was 16%. Characterization of neuroendocrine prostate cancer. | [41] |
| PDO | Healthy mouse and human prostate, human metastatic prostate cancer lesions, and circulating tumor cells | Development of the protocol for the engraftment of normal and tumoral tissues from the prostate. Characterization of the human and mouse organoids. | [38] |
| PDO | PCa specimens from a cohort of 81 patients with different pathological and clinical features | Morphological, immunohistochemical and genomic profiles of whole organoids to define the subtypes correlated to the PCa patients. | [40] |
| PDO | Localized PCa biopsies and radical prostatectomy specimens | Single-cell molecular analyses of established organoids to characterize the heterogeneity of tumor cells, subpopulations of epithelial cells, stromal cells, and tumor microenvironments. | [44] |
| PDX | PCa human cell lines: LNCaP, PC-3, Ca-2, DU145, VCaP | Set-up of the procedures for the generation of prostate cancer PDX models. | [20] |
| PDX | Setup of 80 PDXs derived from 47 human prostate cancer donors. | Some PDXs have generated cell lines to use as working models (MDA-PCa-2a and 2b). The histopathologic, genomic, and molecular characteristics are performed. Treatment with erdafitinib (FGFR inhibitor). | [34] |
| PDX | Setup of 5 PDX models from PCa | This collection included hormone-naïve, androgen-sensitive, and castration-resistant (CRPC) primary tumors, as well as prostate carcinoma with neuroendocrine differentiation (CRPC-NE). Morphological and immunohistochemical description and genomic profiles. Treatment with docetaxel, leuprolenin, enzalutamide, abiraterone, and olaparib. | [35] |
| PDX/PDXO | 59 PDXs established from 41 specimens obtained from 30 PCa patients | Morphological and immunohistochemical description, genomic profiles, and gene expression profiles of MURAL cohort PDXs; 22 PDX tissues were grown as organoids to perform drug screening with apalutamide, enzalutamide, azacytidine, AZD1775 (Wee1 inhibitor), VX-970 (ATR inhibitor), docetaxel, carboplatin, and talazoparib. Drug as a single agent or combination. | [60] |
| PDX/PDXO | PDX model derived from a treatment-naïve soft tissue metastasis (PNPCa), with androgen-sensitive characteristics | Molecular characterization by DNA and RNA sequencing of PDX and establishment of PDXO to assess whether therapy resistance preexists in this treatment-naïve PCa case. | [63] |
| PDO/PDXO | Tissue from human PDX or prostate tissue from genetically engineered mouse model (GEMM) as source material to obtain individual cells | Assess the therapeutic potential of new drugs in the treatment of neuroendocrine prostate cancer (NEPC). Drug as a single agent or combination. | [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rago, V.; Perri, A.; Di Agostino, S. New Therapeutic Perspectives in Prostate Cancer: Patient-Derived Organoids and Patient-Derived Xenograft Models in Precision Medicine. Biomedicines 2023, 11, 2743. https://doi.org/10.3390/biomedicines11102743
Rago V, Perri A, Di Agostino S. New Therapeutic Perspectives in Prostate Cancer: Patient-Derived Organoids and Patient-Derived Xenograft Models in Precision Medicine. Biomedicines. 2023; 11(10):2743. https://doi.org/10.3390/biomedicines11102743
Chicago/Turabian StyleRago, Vittoria, Anna Perri, and Silvia Di Agostino. 2023. "New Therapeutic Perspectives in Prostate Cancer: Patient-Derived Organoids and Patient-Derived Xenograft Models in Precision Medicine" Biomedicines 11, no. 10: 2743. https://doi.org/10.3390/biomedicines11102743