
Identifying Microbiome Dynamics in Pediatric IBD: More than a Family Matter
by
 , and
, and
Nikolas Dovrolis
1,2,† ,
,
Anastasia Moschoviti
3,†,
Smaragdi Fessatou
3,
George Karamanolis
4,
George Kolios
1,2 and
Maria Gazouli
5,6,* 1
Laboratory of Pharmacology, Department of Medicine, Democritus University of Thrace, 68100 Alexandroupolis, Greece
2
Individualised Medicine & Pharmacological Research Solutions Center (IMPReS), 68100 Alexandroupolis, Greece
3
Third Department of Pediatrics, “Attikon” General University Hospital, Medical School, National and Kapodistrian University of Athens, 12462 Haidari, Greece
4
Gastroenterology Unit, Second Department of Surgery, Aretaieio Hospital, Medical School, National and Kapodistrian University of Athens, 11527 Athens, Greece
5
Laboratory of Biology, Department of Basic Medical Sciences, Medical School, National and Kapodistrian University of Athens, 11527 Athens, Greece
6
School of Science and Technology, Hellenic Open University, 26335 Patra, Greece
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomedicines 2023, 11(7), 1979; https://doi.org/10.3390/biomedicines11071979
Submission received: 5 June 2023
/
Revised: 9 July 2023
/
Accepted: 11 July 2023
/
Published: 13 July 2023
(This article belongs to the Special Issue Gut Dysbiosis: Molecular Mechanisms and Therapies 2.0)
Abstract
:Background: Pediatric inflammatory bowel disease (IBD) is a chronic inflammatory intestinal disease that affects both children and adolescents. Symptoms can significantly affect a child’s growth, development, and quality of life, making early diagnosis and effective management crucial. This study focuses on treatment-naïve pediatric IBD patients and their immediate families to identify the role of the microbiome in disease onset. Methods: Nine families with pediatric IBD were recruited, comprising seven drug-naïve Crohn’s disease (CD) patients and two drug-naïve ulcerative colitis (UC) patients, as well as twenty-four healthy siblings/parents. Fecal samples were collected for 16S ribosomal RNA gene sequencing and bioinformatics analysis. Results: We identified patterns of dysbiosis and hallmark microbial taxa among patients who shared ethnic, habitual, and dietary traits with themselves and their families. In addition, we examined the impact of the disease on specific microbial taxa and how these could serve as potential biomarkers for early detection. Conclusions: Our results suggest a potential role of maternal factors in the establishment and modulation of the early life microbiome, consistent with the current literature, which may have implications for understanding the etiology and progression of IBD.
1. Introduction
Pediatric IBD is a chronic inflammatory bowel disease that affects children and adolescents. The two main types of IBD are Crohn’s disease (CD) and ulcerative colitis (UC), which are characterized by inflammation and damage to the digestive tract. Children with IBD may experience a variety of symptoms, including abdominal pain, diarrhea, rectal bleeding, weight loss, and fatigue [1]. These symptoms can significantly impact a child’s growth, development, and quality of life, making early diagnosis and effective management crucial. Diagnosis of pediatric IBD can be challenging, as symptoms can be similar to those of other digestive disorders or infections. Pediatric gastroenterologists will often perform a variety of tests, including blood work, stool samples, endoscopy, and imaging studies, to help confirm a diagnosis [2]. Once a diagnosis has been made, treatment will typically involve a combination of medications and dietary changes [3,4]. Some children may require surgery in more severe cases [5]. One of the unique challenges of managing pediatric IBD is ensuring that treatment plans are appropriate for a child’s age and developmental stage. Medications may need to be adjusted based on weight and age, and some medications may not be appropriate for use in young children. In addition, dietary changes may be necessary to help manage symptoms, but it is important to ensure that a child’s nutritional needs are being met [4]. In addition to physical symptoms, pediatric IBD can also impact a child’s emotional well-being. Children with IBD may experience anxiety, depression, and social isolation, especially if they are unable to participate in normal childhood activities [6]. Overall, managing pediatric IBD requires a multidisciplinary approach, involving gastroenterologists, dietitians, mental health professionals, and other healthcare providers. Regular monitoring and follow-up appointments are important to ensure that symptoms are managed effectively and to identify any potential complications. With proper diagnosis, treatment, and support, children with IBD can lead healthy and fulfilling lives.
Although the cause of IBD is not fully understood, various potential factors have been linked to its development and progression. These include gene expression dysregulation and polymorphisms [7,8], lifestyle choices, and the role of microbiota [9,10]. Despite research, it remains unclear why the disease affects people of all ages. Previous studies have suggested that genetics play an important role in developing IBD [11,12,13] with an up to 12% increased risk for disease occurrence [14]. More recently, with the rise of microbiome-focused research, it has been shown that the intrafamilial microbiome can be linked to specific genetic traits and be studied for its behavior during dysbiosis [15,16]. It has also been reported that microbiota composition is shared between cohabitating individuals [17,18,19] but also displays patterns of similarity that differ between families [20]. Perhaps the most important questions shared by scientists today revolve around whether the microbiome can act as a mediator, a causative agent of disease, or is just its innocent victim [21,22,23,24,25].
Microbiome analysis via bioinformatics (metagenomics analysis) is a rapidly growing field that focuses on analyzing the genetic material of microbial communities present in different environments, including the human body [26]. This involves the use of computational tools and statistical methods to study the diversity and abundance of microorganisms and their functional characteristics [27]. With the advent of high-throughput sequencing technologies, it has become possible to study the microbiome at an unprecedented level of detail, enabling researchers to explore the relationship between microbial communities and human health and disease [28,29]. These techniques play a crucial role in identifying specific microbial taxa associated with different diseases and understanding their functional roles. It also aids in the development of diagnostic and therapeutic tools that can modulate the microbiome to treat various diseases. Moreover, metagenomics analysis is essential for interpreting the vast amount of data generated by microbiome studies, allowing researchers to identify patterns and relationships that are otherwise difficult to detect. As the field continues to expand, it is likely to lead to new insights into the complex interplay between microbial communities and their host organisms, paving the way for the development of personalized medicine approaches that consider an individual’s unique microbiome composition.
In this spirit, our study explores the dynamics of the microbiome within families of pediatric IBD patients, attempting to understand its characteristics during dysbiosis and the different microbial profiles formed within a family. Our methodology was structured for the identification of dysbiosis patterns and hallmark microbial taxa among these patients and their healthy first-degree relatives (parents and siblings). This allowed us not only to present the pediatric microbiome in a unique way but also to identify potential markers of diagnosis and treatability.
2. Results
2.1. IBD versus HC Overall
To identify differences between IBD patients and healthy controls (HC), several analyses were performed for evaluating the quantitative and qualitative differences between them. This allows us to study the potential of microbial taxa as biomarkers for IBD in general and also compare our results to previous works.
2.1.1. Microbiota Composition and Univariate Analysis
The overall relative abundance of microbial phyla when comparing HC and IBD samples presents an increase in Bacteroidetes (10% increase) and Proteobacteria (7% increase) in patients while Firmicutes are decreased (16% decrease) (Figure 1A). On the genus taxonomic level in relative abundance, the patient samples are characterized by an increase in Bacteroides (17%) and a decrease in Faecalibacterium (7%), among other changes (Figure 1B). The univariate analysis of phyla provides statistical significance (p.adjust < 0.05) for Bacteroidetes and Proteobacteria only when the genera Veillonella, Haemophilus, Granulicatella, Erysipelatoclostridium, Shigella, and Streptococcus are significantly increased and members of Candidatus Soleaferrea are decreased. In addition, the species Veillonella parvula, Streptococcus parasanguinis, Haemophilus parainfluenzae, Granulicatella paradiacens, Ruminococcus flavefaciens, Dorea massiliensis, Shigella sonnei, Bacteroides fragilis, Bacteroides acidofaciens, and Bacteroides caccae are significantly increased (FC > 8 and p.adjust < 0.05) while Ruminococcus flavefaciens and Alistipes massiliensis are significantly decreased (FC > 6 and p.adjust < 0.05). The top 10 over- and under-abundant species and genera in the patient group versus all healthy controls are depicted in Table 1. All univariate analysis results for the aforementioned taxonomic levels via DESEQ2 can be found in Supplementary File S1.
2.1.2. Diversity Metrics
When comparing the pool of the nine pediatric IBD patients against the pool of healthy family members (Figure 2A), alpha diversity appears to be reduced, but without achieving statistical significance between the two groups (Kruskal–Wallis p = 0.11). The same effects are also true when subgrouping the HC group according to their familial relationship with the patients (Figure 2B), where the mother/father/brother/sister subgroups appear to exhibit comparable alpha diversities. The loss of statistical significance can be attributed to the wide distribution of values in the patient group, which will be discussed later on. The NMDS plot (Figure 3), used to depict beta diversity, presents clear clustering of the HC samples, while the patient samples are distributed differently. ANOSIM statistics of R = 0.19878 (p < 0.04) signify a high similarity between the groups.
2.2. Intrafamilial Microbiome Changes
By analyzing each family individually, we were able to identify patterns of microbiota behavior during IBD in our pediatric patients and ascertain that the microbiome is heavily affected by the disease in individuals who share lifestyles with our healthy controls.
2.2.1. Biodiversity
Comparing how diverse the microbiomes of family members are shows that pediatric IBD patients diverge significantly. In Figure 4, all a-diversity indices of members of the nine families are shown individually. In six of the nine families, the patients exhibit less biodiversity compared to their healthy family members, while in the rest, they appear to be richer in microbiota from their parents/siblings. This finding, although surprising, can be explained when comparing the alpha diversity indices of patients only, which shows that they are more or less equal, with the exception of Patients 4 and 5, for whom the alpha diversity indices were further decreased. For example, Patients 2 and 9 appear to have lower and higher, respectively, Shannon indices in Figure 4, but the comparison in Figure 2C shows that there is no discernable difference between them. In addition, we observed no differences between children with CD and UC.
2.2.2. Similarity Clustering
As part of our investigation into the dynamics of the microbiome within families of pediatric IBD patients, we utilized Ward’s hierarchical clustering to identify closer similarities among the microbiomes of specific family members. Figure 5 displays the resulting dendrograms, which revealed that in most cases, the microbiome of the patient was found to be either closer to that of the mother or diverging from all other family member microbiomes.
2.2.3. Microbial Composition
To extend our previous analyses of microbiota differential abundance, we performed the same analysis in each family, comparing the pediatric patients against a pool of their family members. Additionally, we calculated the intersection of microbial genera (Figure 6A) and species (Figure 6B) with increases and decreases in their counts. This analysis enabled us to identify patients who exhibit shared differentially abundant taxa. For instance, patients belonging to Families 1 and 2 have only two enriched genera in common (Butyricimonas and Lachnoclostridium) but share 12 genera (Anaerosporobacter, Anaerobacterium, Prevotella, Dorea, Holdemanella, Paraprevotella, Clostridium, Ruminococcus, Lachnobacterium, Pantoea, Peptococcus, and Shigella) that demonstrate decreased population levels. Similarly, Patients 3 and 6 share six enriched genera (Veillonella, Haemophilus, Shigella, Pantoea, Bifidobacterium, and Flavonifractor) and eighteen genera (Phascolarctobacterium, Akkermansia, Anaerobacterium, Trigonala, Holdemanella, Intestinimonas, Spiroplasma, Alistipes, Caloramator, Clostridium, Ruminiclostridium, Sporobacter, Vallitalea, Victivallis, Holdemania, Odoribacter, Coprobacter, and Porphyromonas) displaying reductions in population size. Because of the paucity of samples in each family, we disregarded statistical significance and considered all taxa with a fold change in abundance greater than two (FC > 2). The genera Butyricimonas and Veillonella exhibited increased abundance in the patients of at least six families (considering all multiple combinations), while Anaerosporobacter, Phascolarctobacterium, Akkermansia, Anaerobacterium, Anaerovorax, Lachnospira, Prevotella, and Trigonala were decreased. In addition, the species Butyricimonas virosa and Bacteroides xylanisolvens were more abundant in the patients of at least six families (considering all multiple combinations), while Lachnoclostridium xylanolyticum, Ruminococcus albus, Eubacterium eligens, Ruminococcus lactaris, Blautia luti, Anaerosporobacter mobilis, Holdemanella biforme, Ruminococcus bromii, Anaerobacterium chartisolvens, Trigonala elaeagnus, Coprococcus eutactus, Ruminococcus flavefaciens, Dorea formicigenerans, Akkermansia muciniphila, Anaerovorax odorimutans, Lachnospira pectinoschiza, Lactobacillus rogosae, Streptococcus salivarius, and Sporobacter termitidis showed decreased numbers when compared with the healthy family members. Supplementary File S2 includes the comparative analysis of differential abundance between patients and their healthy relatives within each family. It also provides information on the shared genera and species that are either overabundant or underabundant across multiple families as those appear in Figure 6.
3. Discussion
This work focuses on treatment-naïve pediatric IBD patients and their immediate families to identify the role of the microbiome in disease onset. Our results elucidate the fact that even though family members share lifestyle and dietary habits, the patients exhibit unique microbiome patterns which could only be attributed to the disease. IBD-related dysbiosis shown here carries common characteristics among patients who share ethnic, habitual, and dietary traits among themselves and their families. In addition, we further examine the disease’s impact on specific microbial taxa and how those can serve as potential biomarkers for early detection.
In terms of comparing IBD patients with healthy controls, our study successfully replicates and validates the findings of previous research, such as the loss of Firmicutes [30,31] and the increase in the phylum Bacteroidetes and the genus Bacteroides [32,33] along with Proteobacteria [34], which are well documented in IBD. In addition, several studies have shown that Faecalibacterium prausnitzii, which exhibits anti-inflammatory potential [35], is decreased significantly in IBD [36], a fact that can only partially be confirmed by this study since the genus Faecalibacterium shows a statistically significant reduction in patients but the specific species fails to achieve the required p-value cutoff of reporting. This can be attributed to the wide distribution of F. prausnitzii among all samples due to its universal presence but also given the fact that there are studies showing an actual increase in some pediatric patients [37], balancing out its statistical power. We also report on several other bacterial taxa differentially abundant between patients and healthy controls able to serve as potential IBD biomarkers as we have in previous works with disease activity [38] and response to treatment [39]. Our findings regarding Prevotella align with previous research [40,41], which observed a significant decrease or even depletion of this genus in IBD patients. In a systematic review of gut microbiota profiles in pediatric IBD patients [42], the genus Lachnospira was highlighted as being consistently underabundant in several studies [16,43,44,45,46], in agreement with our own findings. Lachnospira along with Prevotella are two well-established short-chain fatty acid (SCFA)-producing bacterial genera, and their paucity in IBD, and especially CD, is a known factor in promoting inflammation [47]. The role of short-chain fatty acids (SCFAs) in maintaining homeostasis and supporting overall health has been extensively discussed, particularly in relation to dietary habits, which have a significant impact on the composition of the microbiota [48]. Interestingly, despite the influence of geopolitical designations, religion, and daily life on dietary habits, a study conducted on new-onset pediatric Crohn’s patients from Saudi Arabia [49] reported findings consistent with ours regarding Holdemanella and Lactobacillus. Additionally, Sila et al. [50] reported a similar underabundance of Lactobacillus in newly diagnosed pediatric patients with IBD. Furthermore, a study by Malham et al. [51], investigating the role of the microbiome in predicting disease severity and diagnosis in pediatric IBD patients, revealed similar findings to our research regarding the underabundance of Catenibacterium and Akkermansia in patients. Notably, Akkermansia has also been reported to exhibit lower abundance in patients from at least two other studies [52,53]. Another interesting result was the overabundance of Alcaligenes, which, in a previous study [54], the authors had shown that in innate lymphoid-cell-depleted mice, the presence of Alcaligenes was sufficient to promote systemic inflammation, while systemic immune responses to it have been associated with Crohn’s disease. It is not uncommon for studies conducted in different countries, which inherently involve variations in microbiome profiles, to report on the same bacterial taxa with differences in terms of changes in abundance for a given condition. Streptococcus serves as an example, as it has been found to be overabundant in our pediatric IBD patients, consistent with findings from Kowalska-Duplaga et al. [43], Tang et al. [44], Wang et al. [55], Ijaz et al. [45], and Lewis et al. [56], while El Mouzan et al. [49] and Assa et al. [57] report an underabundance in pediatric IBD.
Considering the low biodiversity exhibited by patients in most families when compared to their healthy first-degree relatives, a few things can be deduced. Starting from the fact that even though the patients live in a microbiota-rich environment, they appear to be “resistant” to becoming hosts of more bacteria, and bacteria that attempt to colonize them are either instantly killed by an overactive immune system or do not find enough space to thrive. The spread of pathogenic taxa must either create a hostile microenvironment via metabolism or absorb all the nutrients which are crucial for the survival of the commensal ones. These hypotheses are in line with our current knowledge of dysbiosis [58,59]. Regarding the three families in which healthy relatives show reduced biodiversity when compared to patients, there might be a few reasons to explain the phenomenon, like pathogenic bacteria invasion in patients, strong but low diversity colonies in healthy controls which regulate their immunities, and differences in socioeconomic/dietary/environmental factors when compared to the other families. The fact that alpha diversity among patients is comparable in most cases hints at immunity mechanisms targeting taxa of specific functions, which needs to be further investigated since we were not able to find a specific cause for this when comparing patient samples according to their metadata.
Clustering microbiome profiles according to their similarities has provided evidence that in most cases, pediatric patient microflora either resembles that of the mother or differs significantly from those of all healthy family members. This suggests a potential role for maternal factors in the establishment and modulation of the microbiome in early life, which is consistent with the literature and may have implications for understanding the etiology and progression of IBD. Previous works have shown that the human microbiome is shaped early in life (first 2–3 years) [60,61] and is modulated by maternal-related factors like mode of birth [62,63] and breastfeeding [64,65]. In our results, we could not identify these factors as co-factors of differentiation, but the close resemblance between the patient and maternal microbiome profiles is always an interesting finding, especially when we expand studies beyond neonates, infants, and children into adolescence. This is important because even though the maternal microbiome is essential in early life, adolescents (ages 10–19), due to school and social lives, often diverge from the pure familial environment, with studies expanding beyond common pathophysiological changes in known microbiota-associated disorders like IBD [66,67].
Another consideration for future works is how the non-IBD siblings’ microbiome might show signs of divergence from the parents’ “healthy plane”, as defined by Jacobs et al. [16]. For example, in our study, the siblings’ microbiomes are mostly clustered closer to those of the parents but show, in total, a slight drop in alpha diversity, as shown in Figure 2B. Additionally, some of the differentially abundant taxa characterizing patients observed here might also be prominent in those children. For example, since Butyricimonas and Veillonella appear to be prominent in the patients of most families, along with their increased abundance noted in the literature, they have the potential to serve as prognostic markers of disease occurrence. In Wang et al. [55], Veillonella not only demonstrates increased prevalence in patients undergoing anti-TNF treatment, but it also maintains a notable abundance even after therapy, suggesting resistance to regulation by external factors. In addition, Veillonella appears to be more abundant in several pediatric Crohn’s studies, as reported by a recent meta-analysis [42], suggesting a constant factor among diverse populations. Furthermore, Butyricimonas, with its elevated prevalence, can reveal immunological dysfunctions and impacts, as highlighted by Chen et al. [68], whose research suggests that adhesive bacteria, such as Butyricimonas found in the terminal ileum of pediatric patients, contribute to heightened activation of Th17 cells and the secretion of immunoglobulin A in the gut lumen. These mechanisms can contribute to the inflammatory processes, and given Butyricimonas’ proximity to the intestinal wall, it represents a promising candidate for further investigation. Interestingly, butyrate-producing Clostridia, like Erysipelatoclostridium and Butyricimonas, were found in our study to be enriched in pediatric IBD patients, but not in other studies. This observation is intriguing, as butyrate production has been correlated with the mitigation of inflammation [69]. These discrepancies may be even more pronounced within other families, underscoring the importance of vigilant monitoring in children who may be susceptible to developing IBD.
As is true for all works based on 16S rRNA amplicon sequencing, this study has the potential for small sample sizes to affect the accuracy and reliability of the results. This is because the technique relies on amplifying and sequencing a specific region of the 16S rRNA gene, which may not represent the entire microbial community present in a given sample (in our case the V3/V4 hypervariable regions). Additionally, small sample sizes can also increase the likelihood of sampling bias, as there may be significant variation in microbial communities between individual samples [70]. Furthermore, 16S rRNA amplicon sequencing is limited in its ability to accurately identify certain microbial species and may provide only a broad taxonomic classification of the microorganisms present. This can limit the depth of analysis and hinder the ability to detect important microbial interactions [71]. We have tried to alleviate this by utilizing a pipeline with strict taxonomic identification parameters and cross-checking the results with the literature, but uncertainty still exists when it comes to species-level analyses. Finally, the use of fecal samples provides more diluted information about IBD-related dysbiosis when compared to biopsies [72].
By delving into the intrafamilial microbiome, our investigation yields additional evidence that underscores the complex dynamics involved in the development and progression of IBD, particularly concerning treatment approaches. The modulation of some of the taxa highlighted here has been investigated as a potential therapeutic strategy, and our findings may provide more targets. Candidatus Soleaferrea, shown in our study to have reduced abundance in pediatric patients, has been found to be upregulated in Svolos et al. [73] after Exclusive Enteral Nutrition (ENT) and a food-based diet that simulates it. ENT is considered a first-line treatment for CD and its sub-phenotypes [74]. Haemophilus and the specific species Haemophilus parainfluenzae highlighted in this study are also promising candidates for modulation-based therapies since they were also highlighted as some of the taxa more abundant in first-diagnosis pediatric CD patients by Kansal et al. [75]. Haemophilus parainfluenzae is an opportunistic pathogen that can also be detected in the oral cavity and respiratory tract which a recent study [76] has linked to CD progression and severity. In a recent study by our group [77], we reported on a reverse correlation between Granulicatella and IFN type II expression levels, which holds promise as a prognostic marker for anti-TNF treatment. Similarly, modulating this taxon may have the potential to not only influence the occurrence and outcomes of IBD but also enhance the effectiveness of treatment. There is no definitive proof that modulating dysregulated bacterial populations can treat IBD effectively, but symptom alleviation can significantly improve a patient’s life.
However, it is still unclear why only some family members who share genetic and environmental traits develop IBD. Is it a stroke of luck or a “perfect storm” of aligning factors that cause the disease? Should we blame the specific taxa found to be differentially abundant in this work for mediating the inflammatory process, or do genetic traits spark the onset and the microbiome follows? Further research on this subject remains essential for collecting additional data and delivering answers.
4. Materials and Methods
4.1. Samples
Nine CD (n = 7) and UC (n = 2) probands younger than the age of 16 were recruited from the Pediatric Department of “Attikon” General University Hospital (Table 2). All patients were newly diagnosed and drug-naïve at the time of fecal sample collection. The diagnosis was based on clinical symptoms, laboratory tests, and histological, radiological, and endoscopic findings based on the Porto Criteria recommended by The European Society for Pediatric Gastroenterology Hepatology and Nutrition (ESPGHAN). Family members of these probands were recruited and none of them had a history of IBD or other immune- and/or inflammation-related disorders. Fecal samples from all participants were obtained by Fecal Swab Collection and Preservation System (Norgen BioTek Corp, Thorold, ON, Canada) according to the manufacturer’s instructions. Fecal calprotectin was measured by an enzyme-linked immunosorbent assay (MyBioSource, Inc., San Diego, CA, USA) according to the manufacturer’s instructions. Informed consent was obtained from the parents, and methods were carried out in accordance with relevant guidelines and regulations. The research and all associated experimental protocols were performed in accordance with institutional approval from the hospital ethics committee.
4.2. DNA Extraction and 16S rRNA Amplicon Sequencing
Total DNA was purified from the fecal samples using the Stool DNA Isolation Kit (Norgen BioTek Corp, Thorold, ON, Canada) following the manufacturer’s instructions. Sequencing services were performed by external independent facilities (MR DNA -Molecular Research LP, Shallowater, TX, USA). Sequencing was performed at MR DNA (www.mrdnalab.com (accessed on 1 July 2023), Shallowater, TX, USA) on a MiSeq following the manufacturer’s guidelines. Sequenced reads were quality-controlled, with sequences < 150 bp and containing ambiguous base calls removed. After dereplication, the unique sequences were denoised and had chimeras removed based on the standard QIIME2 [78] pipeline using DADA2 [79], providing a denoised sequence or zOTU. Final zOTUs were taxonomically classified using BLASTn [80] (99% sequence similarity) against a curated database derived from NCBI. The final library contained samples with an average of 30,000 aligned reads.
4.3. 16S rRNA Bioinformatics Analysis
Aligned raw read counts, sample metadata, and taxonomy information files were formatted and used as input to the MicrobiomeAnalyst [81] platform for analysis. The original total of 1099 zOTUs was filtered to 384 after accounting for low read counts (<20% prevalence in all samples). Samples were normalized using total sum scaling (TSS) to avoid isolation/sequencing biases. All samples were included for the IBD versus healthy controls (HC) comparisons and sub-grouped for the per-family analyses. Alpha diversity analysis (unfiltered raw counts) was performed using the Shannon index on the zOTU level, beta diversity was calculated via NMDS and ANOSIM, and univariate differential abundance calculations were performed on the genera and species levels using DESEQ2 considering their fold change (FC) and false discovery rate adjusted p-values (p.adjust). Intrafamily microbiome similarity clustering was performed using Ward’s hierarchical clustering method. To ensure the reproducibility of results, we include in the supplementary material the necessary input files for MicrobiomeAnalyst from our data: “Supplementary File S3.txt”, “Supplementary File S4.txt”, and “Supplementary File S4.txt”; these files contain the raw counts, metadata, and taxonomic classification of our samples, respectively. Intersections of microbial taxa between families were created using the multiple-list intersection of the Molbiotools online platform [82].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biomedicines11071979/s1.
Author Contributions
Conceptualization, M.G. and G.K. (George Kolios); methodology, A.M., N.D., S.F. and G.K. (George Kolios); investigation, A.M., M.G., N.D., S.F., G.K. (George Karamanolisand) and G.K. (George Kolios); resources, S.F. and M.G.; writing—original draft preparation, N.D. and M.G.; writing—review and editing, A.M., N.D., S.F., G.K. (George Karamanolisand) and G.K. (George Kolios); funding acquisition, S.F. and M.G. All authors have read and agreed to the published version of the manuscript.
Funding
S.F., G.K. (George Karamanolisand) and M.G. are funded by the project 2020EOMIFNEp3 financed by the Hellenic Idiopathic Inflammatory Bowel Disease Study Group. N.D. and G.K. (George Kolios) are funded by the project IMPReS (MIS 5047189) financed by the Program “Competitiveness, Entrepreneurship and Innovation” (NSRF 2014–2020) and co-financed by Greece and the European Union (European Regional Development Fund).
Data Availability Statement
Data available as Supplementary Files S3–S5.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peloquin, J.M.; Goel, G.; Villablanca, E.J.; Xavier, R.J. Mechanisms of pediatric inflammatory bowel disease. Annu. Rev. Immunol. 2016, 34, 31–64. [Google Scholar] [CrossRef] [PubMed]
- Conrad, M.A.; Rosh, J.R. Pediatric inflammatory bowel disease. Pediatr. Clin. 2017, 64, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Corica, D.; Romano, C. Biological therapy in pediatric inflammatory bowel disease. J. Clin. Gastroenterol. 2017, 51, 100–110. [Google Scholar] [CrossRef]
- Miele, E.; Shamir, R.; Aloi, M.; Assa, A.; Braegger, C.; Bronsky, J.; De Ridder, L.; Escher, J.C.; Hojsak, I.; Kolaček, S. Nutrition in pediatric inflammatory bowel disease: A position paper on behalf of the Porto Inflammatory Bowel Disease Group of the European Society of Pediatric Gastroenterology, Hepatology and Nutrition. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 687–708. [Google Scholar] [CrossRef] [Green Version]
- Ricciuto, A.; Aardoom, M.; Orlanski-Meyer, E.; Navon, D.; Carman, N.; Aloi, M.; Bronsky, J.; Däbritz, J.; Dubinsky, M.; Hussey, S. Predicting outcomes in pediatric crohn’s disease for management optimization: Systematic review and consensus statements from the pediatric inflammatory bowel disease–ahead program. Gastroenterology 2021, 160, 403–436.e426. [Google Scholar] [CrossRef]
- Goldstein-Leever, A.; Bass, J.A.; Goyal, A.; Maddux, M.H. Health-related quality of life predicts psychology referral in youth with inflammatory bowel disease. J. Pediatr. Nurs. 2019, 47, 73–77. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Gazouli, M.; Dovrolis, N.; Franke, A.; Spyrou, G.M.; Sechi, L.A.; Kolios, G. Differential genetic and functional background in inflammatory bowel disease phenotypes of a Greek population: A systems bioinformatics approach. Gut Pathog. 2019, 11, 31. [Google Scholar] [CrossRef]
- Caruso, R.; Lo, B.C.; Núñez, G. Host–microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- Kumar, A.; Al-Hassi, H.O.; Steed, H.; Phipps, O.; Brookes, M.J. Bile acids and the microbiome: Making sense of this dynamic relationship in their role and management in Crohn’s disease. Can. J. Gastroenterol. Hepatol. 2022, 2022, 8416578. [Google Scholar] [CrossRef]
- Russell, R.K.; Satsangi, J. Does IBD run in families? Inflamm. Bowel Dis. 2008, 14 (Suppl. 2), S20–S21. [Google Scholar] [CrossRef] [PubMed]
- Spencer, E.A.; Helmus, D.; Telesco, S.; Colombel, J.-F.; Dubinsky, M.C. Inflammatory bowel disease clusters within affected sibships in Ashkenazi Jewish multiplex families. Gastroenterology 2020, 159, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Colombel, J.F.; Grandbastien, B.; Gower-Rousseau, C.; Plegat, S.; Evrard, J.P.; Dupas, J.L.; Gendre, J.P.; Modigliani, R.; Belaiche, J.; Hostein, J. Clinical characteristics of Crohn’s disease in 72 families. Gastroenterology 1996, 111, 604–607. [Google Scholar] [CrossRef]
- Moller, F.T.; Andersen, V.; Wohlfahrt, J.; Jess, T. Familial risk of inflammatory bowel disease: A population-based cohort study 1977–2011. Off. J. Am. Coll. Gastroenterol. 2015, 110, 564–571. [Google Scholar] [CrossRef]
- Sharma, A.; Junge, O.; Szymczak, S.; Rühlemann, M.C.; Enderle, J.; Schreiber, S.; Laudes, M.; Franke, A.; Lieb, W.; Krawczak, M. Network-based quantitative trait linkage analysis of microbiome composition in inflammatory bowel disease families. Front. Genet. 2023, 14, 1048312. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.P.; Goudarzi, M.; Singh, N.; Tong, M.; McHardy, I.H.; Ruegger, P.; Asadourian, M.; Moon, B.H.; Ayson, A.; Borneman, J.; et al. A Disease-Associated Microbial and Metabolomics State in Relatives of Pediatric Inflammatory Bowel Disease Patients. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 750–766. [Google Scholar] [CrossRef] [Green Version]
- Song, S.J.; Lauber, C.; Costello, E.K.; Lozupone, C.A.; Humphrey, G.; Berg-Lyons, D.; Caporaso, J.G.; Knights, D.; Clemente, J.C.; Nakielny, S. Cohabiting family members share microbiota with one another and with their dogs. Elife 2013, 2, e00458. [Google Scholar] [CrossRef]
- Mosites, E.; Sammons, M.; Otiang, E.; Eng, A.; Noecker, C.; Manor, O.; Hilton, S.; Thumbi, S.M.; Onyango, C.; Garland-Lewis, G. Microbiome sharing between children, livestock and household surfaces in western Kenya. PLoS ONE 2017, 12, e0171017. [Google Scholar] [CrossRef] [Green Version]
- Valles-Colomer, M.; Blanco-Míguez, A.; Manghi, P.; Asnicar, F.; Dubois, L.; Golzato, D.; Armanini, F.; Cumbo, F.; Huang, K.D.; Manara, S. The person-to-person transmission landscape of the gut and oral microbiomes. Nature 2023, 614, 125–135. [Google Scholar] [CrossRef]
- Schloss, P.D.; Iverson, K.D.; Petrosino, J.F.; Schloss, S.J. The dynamics of a family’s gut microbiota reveal variations on a theme. Microbiome 2014, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Vijay, A.; Valdes, A.M. Role of the gut microbiome in chronic diseases: A narrative review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and disease markers correlate with gut microbiome composition across thousands of people. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chang, E.B. Inflammatory bowel diseases (IBD) and the microbiome—Searching the crime scene for clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Olbjørn, C.; Småstuen, M.C.; Moen, A.E.F. Targeted analysis of the gut microbiome for diagnosis, prognosis and treatment individualization in pediatric inflammatory bowel disease. Microorganisms 2022, 10, 1273. [Google Scholar] [CrossRef] [PubMed]
- Riesenfeld, C.S.; Schloss, P.D.; Handelsman, J. Metagenomics: Genomic analysis of microbial communities. Annu. Rev. Genet. 2004, 38, 525–552. [Google Scholar] [CrossRef] [Green Version]
- Calle, M.L. Statistical analysis of metagenomics data. Genom. Inform. 2019, 17, e6. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: Mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Loh, G.; Blaut, M. Role of commensal gut bacteria in inflammatory bowel diseases. Gut Microbes 2012, 3, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.T.; Amos, G.C.; Murphy, A.R.; Murch, S.; Wellington, E.M.; Arasaradnam, R.P. Microbial imbalance in inflammatory bowel disease patients at different taxonomic levels. Gut Pathog. 2020, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhi, F. Lower Level of Bacteroides in the Gut Microbiota Is Associated with Inflammatory Bowel Disease: A Meta-Analysis. Biomed Res. Int. 2016, 2016, 5828959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, K.; Ishikawa, D. Bacteroidetes Species Are Correlated with Disease Activity in Ulcerative Colitis. 2021, 10, 1749. J. Clin. Med. 2021, 10, 1749. [Google Scholar] [CrossRef]
- Vester-Andersen, M.; Mirsepasi-Lauridsen, H.; Prosberg, M.; Mortensen, C.; Träger, C.; Skovsen, K.; Thorkilgaard, T.; Nøjgaard, C.; Vind, I.; Krogfelt, K.A. Increased abundance of proteobacteria in aggressive Crohn’s disease seven years after diagnosis. Sci. Rep. 2019, 9, 13473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez-Humaran, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Eppinga, H.; Sperna Weiland, C.J.; Thio, H.B.; van der Woude, C.J.; Nijsten, T.E.; Peppelenbosch, M.P.; Konstantinov, S.R. Similar depletion of protective Faecalibacterium prausnitzii in psoriasis and inflammatory bowel disease, but not in hidradenitis suppurativa. J. Crohn’s Colitis 2016, 10, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Hansen, R.; Russell, R.K.; Reiff, C.; Louis, P.; McIntosh, F.; Berry, S.H.; Mukhopadhya, I.; Bisset, W.M.; Barclay, A.R.; Bishop, J.; et al. Microbiota of de-novo pediatric IBD: Increased Faecalibacterium prausnitzii and reduced bacterial diversity in Crohn’s but not in ulcerative colitis. Am. J. Gastroenterol. 2012, 107, 1913–1922. [Google Scholar] [CrossRef] [Green Version]
- Dovrolis, N.; Drygiannakis, I.; Filidou, E.; Kandilogiannakis, L.; Arvanitidis, K.; Tentes, I.; Kolios, G.; Valatas, V. Gut Microbial Signatures Underline Complicated Crohn’s Disease but Vary Between Cohorts; An In Silico Approach. Inflamm. Bowel Dis. 2019, 25, 217–225. [Google Scholar] [CrossRef]
- Dovrolis, N.; Michalopoulos, G.; Theodoropoulos, G.E.; Arvanitidis, K.; Kolios, G.; Sechi, L.A.; Eliopoulos, A.G.; Gazouli, M. The interplay between mucosal microbiota composition and host gene-expression is linked with infliximab response in inflammatory bowel diseases. Microorganisms 2020, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Rausch, P.; Rehman, A.; Künzel, S.; Häsler, R.; Ott, S.J.; Schreiber, S.; Rosenstiel, P.; Franke, A.; Baines, J.F. Colonic mucosa-associated microbiota is influenced by an interaction of Crohn disease and FUT2 (Secretor) genotype. Proc. Natl. Acad. Sci. USA 2011, 108, 19030–19035. [Google Scholar] [CrossRef]
- Zhuang, X.; Liu, C.; Zhan, S.; Tian, Z.; Li, N.; Mao, R.; Zeng, Z.; Chen, M. Gut microbiota profile in pediatric patients with inflammatory bowel disease: A systematic review. Front. Pediatr. 2021, 9, 626232. [Google Scholar] [CrossRef]
- Kowalska-Duplaga, K.; Gosiewski, T.; Kapusta, P.; Sroka-Oleksiak, A.; Wędrychowicz, A.; Pieczarkowski, S.; Ludwig-Słomczyńska, A.H.; Wołkow, P.P.; Fyderek, K. Differences in the intestinal microbiome of healthy children and patients with newly diagnosed Crohn’s disease. Sci. Rep. 2019, 9, 18880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Huang, Y.; Shi, P.; Wang, Y.; Zhang, Y.; Xue, A.; Tang, Z.; Hu, W.; Sun, H.; Zhang, P. Effect of exclusive enteral nutrition on the disease process, nutrition status, and gastrointestinal microbiota for Chinese children with Crohn’s disease. J. Parenter. Enter. Nutr. 2021, 45, 826–838. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, U.Z.; Quince, C.; Hanske, L.; Loman, N.; Calus, S.T.; Bertz, M.; Edwards, C.A.; Gaya, D.R.; Hansen, R.; McGrogan, P. The distinct features of microbial ‘dysbiosis’ of Crohn’s disease do not occur to the same extent in their unaffected, genetically-linked kindred. PLoS ONE 2017, 12, e0172605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Kaplan, J.L.; Gold, B.D.; Bhasin, M.K.; Ward, N.L.; Kellermayer, R.; Kirschner, B.S.; Heyman, M.B.; Dowd, S.E.; Cox, S.B. Detecting microbial dysbiosis associated with pediatric Crohn disease despite the high variability of the gut microbiota. Cell Rep. 2016, 14, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Cheng, S.; Yao, J.; Lin, X.; Li, Y.; Wang, W.; Weng, J.; Zou, Y.; Zhu, L.; Zhi, M. Correlation between altered gut microbiota and elevated inflammation markers in patients with Crohn’s disease. Front. Immunol. 2022, 13, 947313. [Google Scholar] [CrossRef]
- Xiong, R.-G.; Zhou, D.-D.; Wu, S.-X.; Huang, S.-Y.; Saimaiti, A.; Yang, Z.-J.; Shang, A.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Health benefits and side effects of short-chain fatty acids. Foods 2022, 11, 2863. [Google Scholar] [CrossRef]
- El Mouzan, M.I.; Winter, H.S.; Assiri, A.A.; Korolev, K.S.; Al Sarkhy, A.A.; Dowd, S.E.; Al Mofarreh, M.A.; Menon, R. Microbiota profile in new-onset pediatric Crohn’s disease: Data from a non-Western population. Gut Pathog. 2018, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Sila, S.; Jelic, M.; Trivic, I.; Andraševic, A.T.; Hojsak, I.; Kolacek, S. Altered gut microbiota is present in newly diagnosed pediatric patients with inflammatory bowel disease. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 497–502. [Google Scholar] [CrossRef]
- Malham, M.; Lilje, B.; Houen, G.; Winther, K.; Andersen, P.S.; Jakobsen, C. The microbiome reflects diagnosis and predicts disease severity in paediatric onset inflammatory bowel disease. Scand. J. Gastroenterol. 2019, 54, 969–975. [Google Scholar] [CrossRef]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H. Mucolytic bacteria with increased prevalence in IBD mucosa augmentin vitroutilization of mucin by other bacteria. Off. J. Am. Coll. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- Höyhtyä, M.; Korpela, K.; Saqib, S.; Junkkari, S.; Nissilä, E.; Nikkonen, A.; Dikareva, E.; Salonen, A.; de Vos, W.M.; Kolho, K.-L. Quantitative fecal microbiota profiles relate to therapy response during induction with tumor necrosis factor α antagonist infliximab in pediatric inflammatory bowel disease. Inflamm. Bowel Dis. 2023, 29, 116–124. [Google Scholar] [CrossRef]
- Sonnenberg, G.F.; Monticelli, L.A.; Alenghat, T.; Fung, T.C.; Hutnick, N.A.; Kunisawa, J.; Shibata, N.; Grunberg, S.; Sinha, R.; Zahm, A.M. Innate lymphoid cells promote anatomical containment of lymphoid-resident commensal bacteria. Science 2012, 336, 1321–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gao, X.; Ghozlane, A.; Hu, H.; Li, X.; Xiao, Y.; Li, D.; Yu, G.; Zhang, T. Characteristics of faecal microbiota in paediatric Crohn’s disease and their dynamic changes during infliximab therapy. J. Crohn’s Colitis 2018, 12, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.D.; Chen, E.Z.; Baldassano, R.N.; Otley, A.R.; Griffiths, A.M.; Lee, D.; Bittinger, K.; Bailey, A.; Friedman, E.S.; Hoffmann, C. Inflammation, antibiotics, and diet as environmental stressors of the gut microbiome in pediatric Crohn’s disease. Cell Host Microbe 2015, 18, 489–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assa, A.; Butcher, J.; Li, J.; Elkadri, A.; Sherman, P.M.; Muise, A.M.; Stintzi, A.; Mack, D. Mucosa-associated ileal microbiota in new-onset pediatric Crohn’s disease. Inflamm. Bowel Dis. 2016, 22, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Santana, P.T.; Rosas, S.L.B.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S. Dysbiosis in inflammatory bowel disease: Pathogenic role and potential therapeutic targets. Int. J. Mol. Sci. 2022, 23, 3464. [Google Scholar] [CrossRef]
- Sultan, S.; El-Mowafy, M.; Elgaml, A.; Ahmed, T.A.; Hassan, H.; Mottawea, W. Metabolic influences of gut microbiota dysbiosis on inflammatory bowel disease. Front. Physiol. 2021, 12, 715506. [Google Scholar] [CrossRef]
- Stinson, L.F. Establishment of the early-life microbiome: A DOHaD perspective. J. Dev. Orig. Health Dis. 2020, 11, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Ríos-Covian, D.; Langella, P.; Martín, R. From short-to long-term effects of c-section delivery on microbiome establishment and host health. Microorganisms 2021, 9, 2122. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, N.T.; Bakacs, E.; Combellick, J.; Grigoryan, Z.; Dominguez-Bello, M.G. The infant microbiome development: Mom matters. Trends Mol. Med. 2015, 21, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casas, L.; Karvonen, A.M.; Kirjavainen, P.V.; Täubel, M.; Hyytiäinen, H.; Jayaprakash, B.; Lehmann, I.; Standl, M.; Pekkanen, J.; Heinrich, J. Early life home microbiome and hyperactivity/inattention in school-age children. Sci. Rep. 2019, 9, 17355. [Google Scholar] [CrossRef] [Green Version]
- Flannery, J.; Callaghan, B.; Sharpton, T.; Fisher, P.; Pfeifer, J. Is adolescence the missing developmental link in Microbiome–Gut–Brain axis communication? Dev. Psychobiol. 2019, 61, 783–795. [Google Scholar] [CrossRef]
- Chen, B.; Ye, D.; Luo, L.; Liu, W.; Peng, K.; Shu, X.; Gu, W.; Wang, X.; Xiang, C.; Jiang, M. Adhesive bacteria in the terminal ileum of children correlates with increasing Th17 cell activation. Front. Pharmacol. 2020, 11, 588560. [Google Scholar] [CrossRef]
- Chen, J.; Vitetta, L. The role of butyrate in attenuating pathobiont-induced hyperinflammation. Immune Netw. 2020, 20, e15. [Google Scholar] [CrossRef]
- Poretsky, R.; Rodriguez-R, L.M.; Luo, C.; Tsementzi, D.; Konstantinidis, K.T. Strengths and limitations of 16S rRNA gene amplicon sequencing in revealing temporal microbial community dynamics. PLoS ONE 2014, 9, e93827. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Jin, G.; Wang, G.; Liu, T.; Liu, X.; Wang, B.; Cao, H. Current sampling methods for gut microbiota: A call for more precise devices. Front. Cell. Infect. Microbiol. 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Svolos, V.; Hansen, R.; Nichols, B.; Quince, C.; Ijaz, U.Z.; Papadopoulou, R.T.; Edwards, C.A.; Watson, D.; Alghamdi, A.; Brejnrod, A. Treatment of active Crohn’s disease with an ordinary food-based diet that replicates exclusive enteral nutrition. Gastroenterology 2019, 156, 1354–1367.e1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamji, M.; Day, A.S. An overview of the role of exclusive enteral nutrition for complicated Crohn’s disease. Intest. Res. 2019, 17, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Kansal, S.; Catto-Smith, A.G.; Boniface, K.; Thomas, S.; Cameron, D.J.; Oliver, M.; Alex, G.; Kirkwood, C.D.; Wagner, J. The microbiome in paediatric crohn’s disease—A longitudinal, prospective, single-centre study. J. Crohn’s Colitis 2019, 13, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Sohn, J.; Li, L.; Zhang, L.; Genco, R.J.; Falkner, K.L.; Tettelin, H.; Rowsam, A.M.; Smiraglia, D.J.; Novak, J.M.; Diaz, P.I. Periodontal disease is associated with increased gut colonization of pathogenic Haemophilus parainfluenzae in patients with Crohn’s disease. Cell Rep. 2023, 42, 112120. [Google Scholar] [CrossRef]
- Mavragani, C.P.; Nezos, A.; Dovrolis, N.; Andreou, N.-P.; Legaki, E.; Sechi, L.A.; Bamias, G.; Gazouli, M. Type I and II interferon signatures can predict the response to anti-TNF agents in inflammatory bowel disease patients: Involvement of the microbiota. Inflamm. Bowel Dis. 2020, 26, 1543–1553. [Google Scholar] [CrossRef]
- Estaki, M.; Jiang, L.; Bokulich, N.A.; McDonald, D.; González, A.; Kosciolek, T.; Martino, C.; Zhu, Q.; Birmingham, A.; Vázquez-Baeza, Y. QIIME 2 enables comprehensive end-to-end analysis of diverse microbiome data and comparative studies with publicly available data. Curr. Protoc. Bioinform. 2020, 70, e100. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.molbiotools.com/listcompare.php (accessed on 1 July 2023).
Figure 1.
Stacked bar plots representing the bacterial relative abundance of (A) healthy family members and patients at the phylum level and (B) healthy family members and patients at the genus level.
Figure 1.
Stacked bar plots representing the bacterial relative abundance of (A) healthy family members and patients at the phylum level and (B) healthy family members and patients at the genus level.
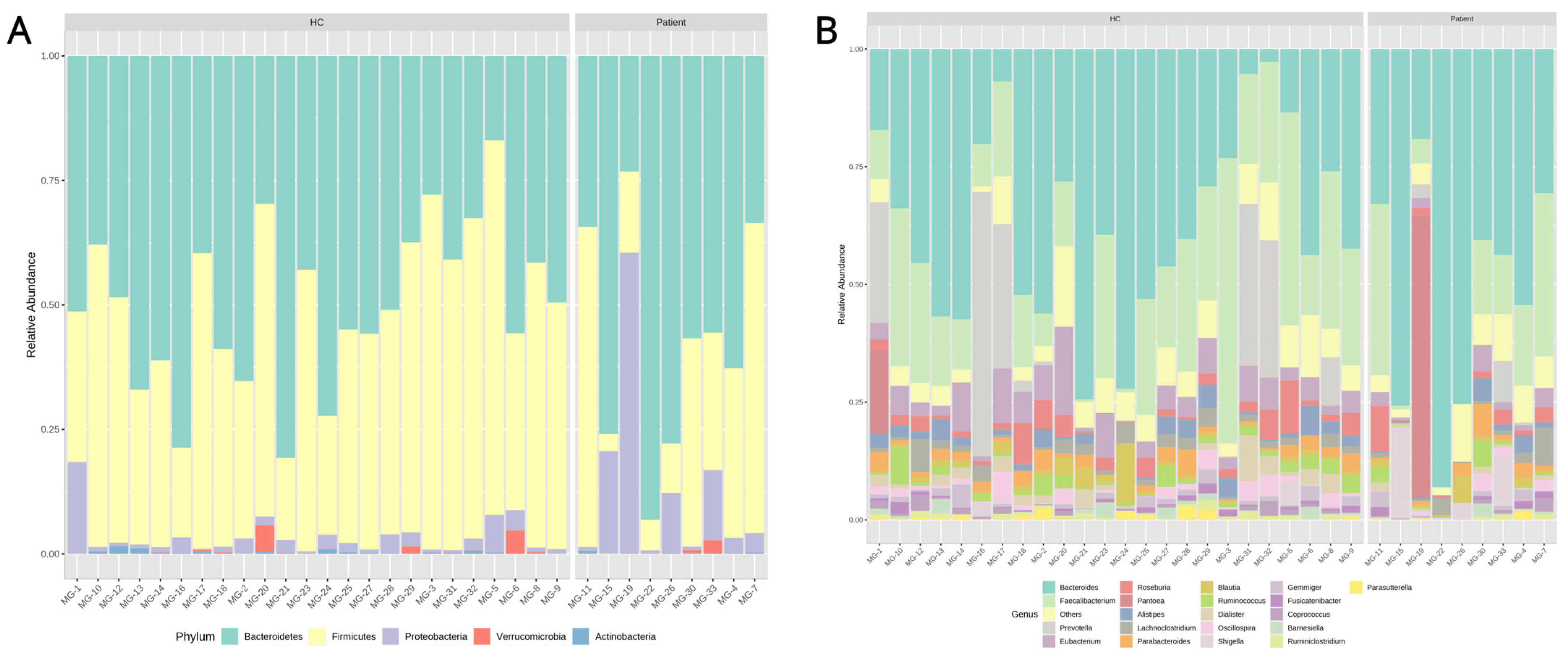
Figure 2.
Alpha diversity indices (Shannon index) representing how biodiverse the sample groupings are. (A) Comparing healthy controls, in total, versus pediatric IBD patients. (B) Comparing healthy controls grouped by their familial relationship with the patients versus those patients. (C) Comparing individual patients from all families among themselves.
Figure 2.
Alpha diversity indices (Shannon index) representing how biodiverse the sample groupings are. (A) Comparing healthy controls, in total, versus pediatric IBD patients. (B) Comparing healthy controls grouped by their familial relationship with the patients versus those patients. (C) Comparing individual patients from all families among themselves.
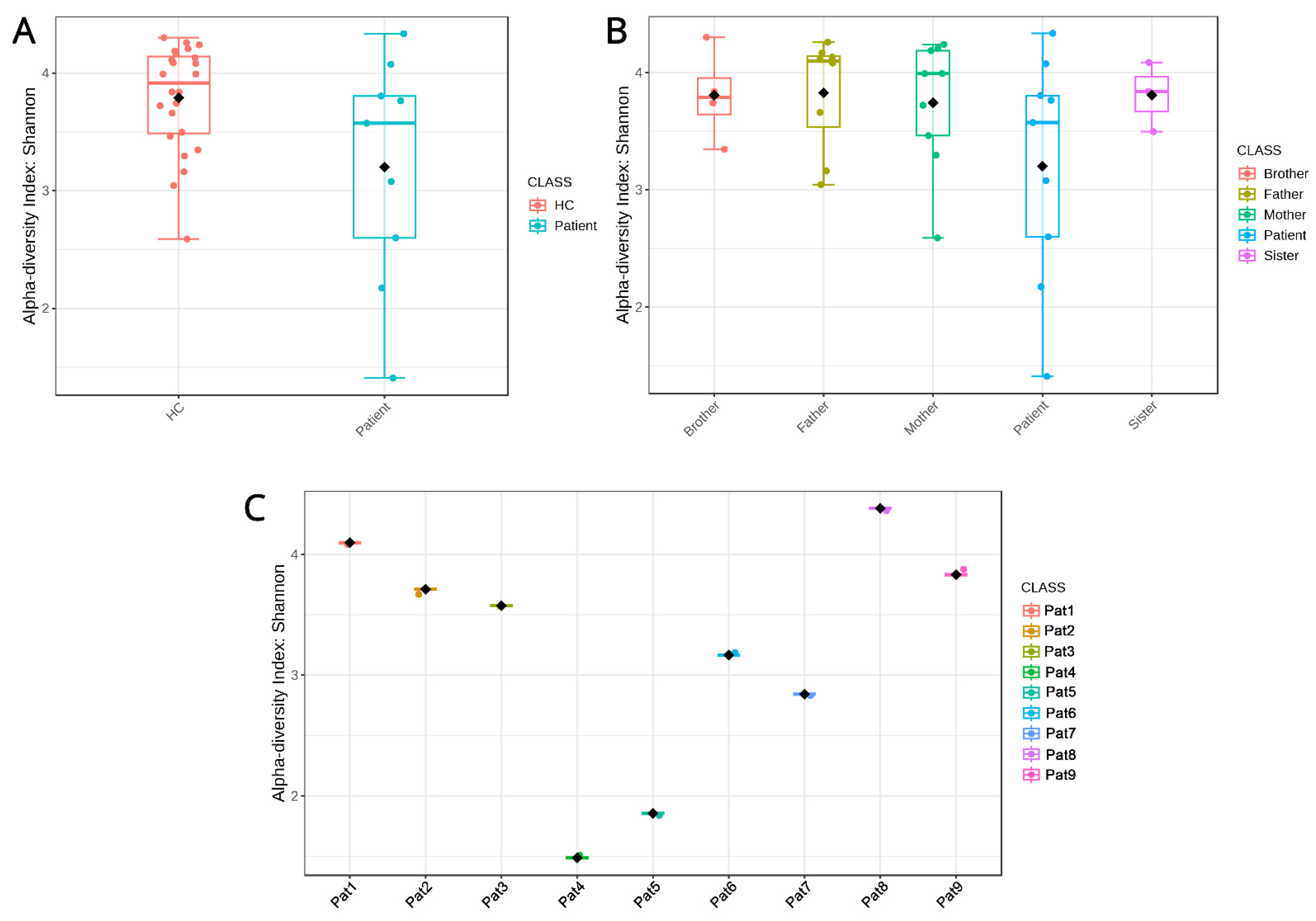
Figure 3.
Non-metric multi-dimensional scaling (NMDS) plot depicting the qualitative and quantitative (dis)similarities between samples of healthy controls and pediatric IBD patients.
Figure 3.
Non-metric multi-dimensional scaling (NMDS) plot depicting the qualitative and quantitative (dis)similarities between samples of healthy controls and pediatric IBD patients.

Figure 4.
Alpha diversity indices (Shannon index) representing how biodiverse the members of each family are when compared to each other. Red boxes highlight the patients in each family.
Figure 4.
Alpha diversity indices (Shannon index) representing how biodiverse the members of each family are when compared to each other. Red boxes highlight the patients in each family.

Figure 5.
Hierarchical clustering trees for each family. Clustering family members within each family based on their microbiota composition allows us to detect similarities among them. Red boxes highlight the patients in each family.
Figure 5.
Hierarchical clustering trees for each family. Clustering family members within each family based on their microbiota composition allows us to detect similarities among them. Red boxes highlight the patients in each family.

Figure 6.
Intersections of differentially abundant taxa (FC > 2 upregulation or downregulation of abundance) among families based on intrafamilial differential abundance comparisons between patients and non-IBD members. (A) Intersections of microbial genera. (B) Intersections of microbial species. Color intensity represents higher intersection values. Numbers in each box are the number of differentially abundant taxa in each intersection.
Figure 6.
Intersections of differentially abundant taxa (FC > 2 upregulation or downregulation of abundance) among families based on intrafamilial differential abundance comparisons between patients and non-IBD members. (A) Intersections of microbial genera. (B) Intersections of microbial species. Color intensity represents higher intersection values. Numbers in each box are the number of differentially abundant taxa in each intersection.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Top 10 over- and under-abundant species and genera in the pediatric patient group versus all healthy controls.
Table 1.
Top 10 over- and under-abundant species and genera in the pediatric patient group versus all healthy controls.
| Genera More Abundant in IBD | Genera Less Abundant in IBD | Species More Abundant in IBD | Species Less Abundant in IBD |
|---|---|---|---|
| Veillonella | Candidatus Soleaferrea | Veillonella_parvula | Ruminococcus__flavefaciens |
| Haemophilus | Prevotella | Haemophilus_parainfluenzae | Bacteroides__massiliensis |
| Granulicatella | Holdemanella | Erysipelatoclostridium_ramosum | Prevotella__copri |
| Sutterella | Anaerobacterium | Granulicatella_paradiacens | Bacteroides__stercoris |
| Shigella | Lactobacillus | Streptococcus_parasanguinis | Dialister__succinatiphilus |
| Erysipelatoclostridium | Holdemania | Bacteroides_caccae | Lachnoclostridium__xylanolyticum |
| Pantoea | Catenibacterium | Shigella_sonnei | Eubacterium__hallii |
| Streptococcus | Lachnospira | Sutterella_wadsworthensis | Alistipes__indistinctus |
| Butyricimonas | Dorea | Bacteroides_fragilis | Holdemanella__biforme |
| Alcaligenes | Anaerovorax | Bacteroides_acidofaciens | Anaerobacterium__chartisolvens |
Table 2.
Patient demographic data.
| Family Identifier | Age at Diagnosis | Gender | IBD Phenotype | Fecal Calprotectin (μg/fecal gr) | CRP 1 (mg/dL) | ESR 2 (mm/h) | PUCAI/PUCDAI 3 | Birth Mode | Prematurity |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 13 | F | CD | 250 | <3 | 1 | 25 (Mild) | Cesarean | Yes |
| 2 | 12 | M | CD | 530 | <3 | 40 | 30 (Moderate) | Cesarean | No |
| 3 | 14 | F | UC | 600 | 8 | 50 | 20 (Mild) | Vaginal | No |
| 4 | 4.5 | F | CD | 95 | 96 | 120 | >40 (Severe) | Vaginal | No |
| 5 | 16 | M | CD | 1660 | 110 | 65 | >40 (Severe) | Vaginal | No |
| 6 | 9 | M | CD | 1200 | 100 | 40 | >40 (Severe) | Cesarean | No |
| 7 | 13 | M | CD | 1500 | 40 | 30 | 37.5 (Moderate) | Cesarean | Yes |
| 8 | 16 | M | CD | 1200 | 20 | 55 | >40 (Severe) | Cesarean | No |
| 9 | 13.5 | M | UC | 1270 | <3 | 15 | 50 (Moderate) | Vaginal | No |
1 CRP: C-reactive protein; 2 ESR: erythrocyte sedimentation rate; 3 PUCAI/PUCDAI: Pediatric Ulcerative Colitis Activity Index/Pediatric Crohn’s Disease Activity Index.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dovrolis, N.; Moschoviti, A.; Fessatou, S.; Karamanolis, G.; Kolios, G.; Gazouli, M. Identifying Microbiome Dynamics in Pediatric IBD: More than a Family Matter. Biomedicines 2023, 11, 1979. https://doi.org/10.3390/biomedicines11071979
AMA Style
Dovrolis N, Moschoviti A, Fessatou S, Karamanolis G, Kolios G, Gazouli M. Identifying Microbiome Dynamics in Pediatric IBD: More than a Family Matter. Biomedicines. 2023; 11(7):1979. https://doi.org/10.3390/biomedicines11071979
Chicago/Turabian StyleDovrolis, Nikolas, Anastasia Moschoviti, Smaragdi Fessatou, George Karamanolis, George Kolios, and Maria Gazouli. 2023. "Identifying Microbiome Dynamics in Pediatric IBD: More than a Family Matter" Biomedicines 11, no. 7: 1979. https://doi.org/10.3390/biomedicines11071979
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.