
Gut Microbiota Signatures with Potential Clinical Usefulness in Colorectal and Non-Small Cell Lung Cancers

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. DNA Preparation and Sequencing
2.3. Statistical Analysis
3. Results
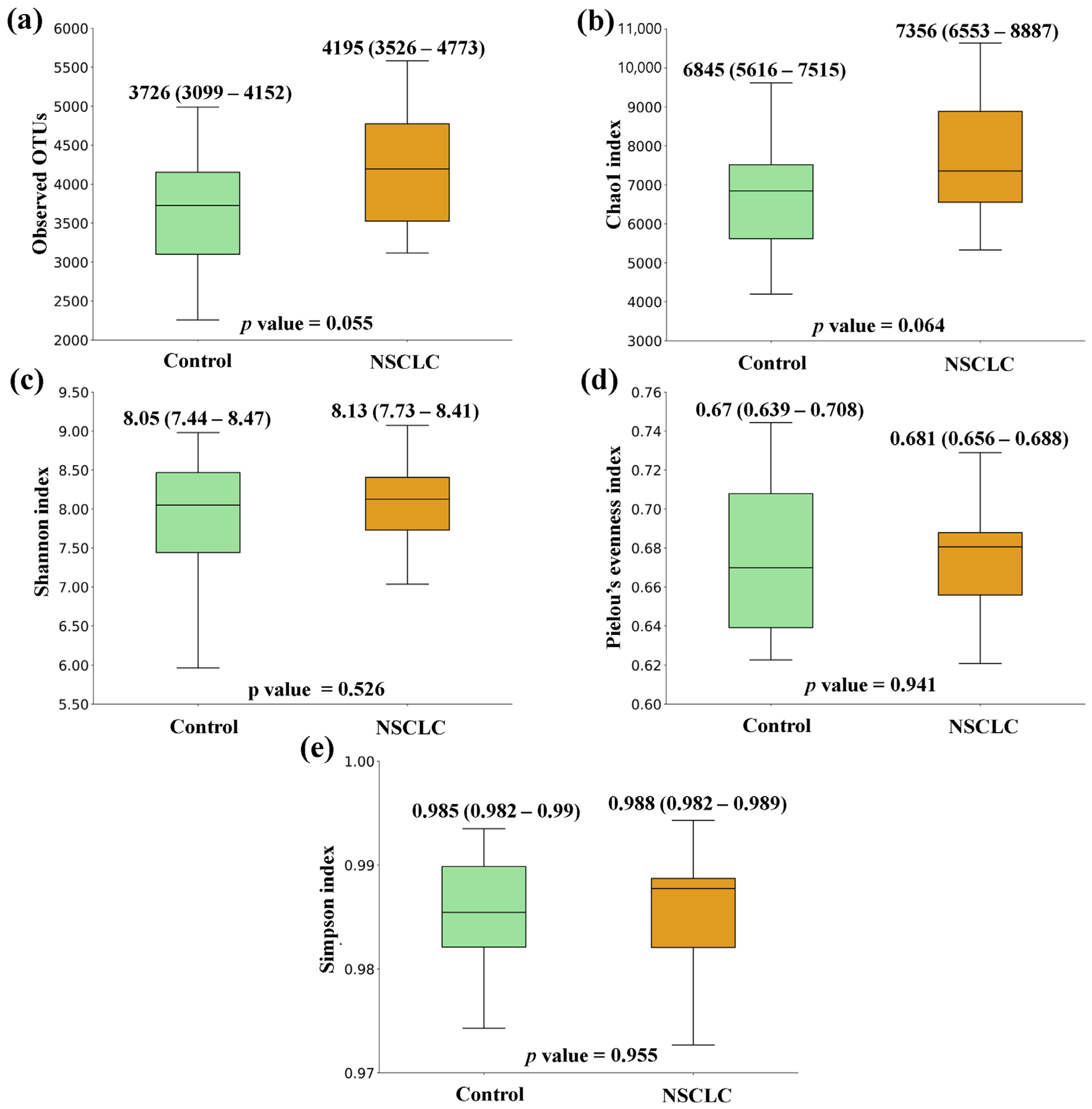
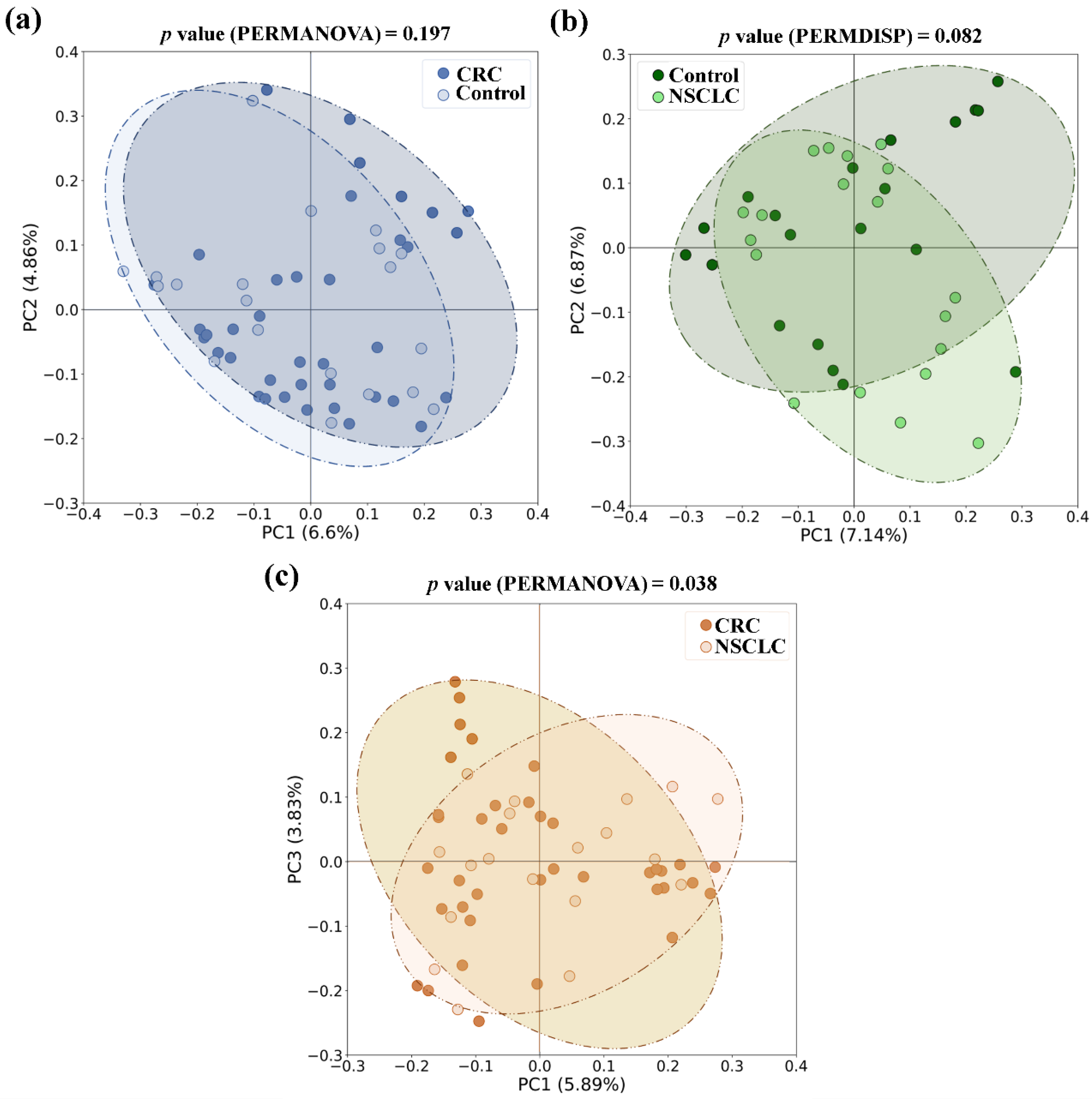
3.1. Comparison of Microbial Diversity between Feces from CRC Patients, NSCLC Patients, and Control Group
3.2. Taxonomic Comparison of Microbiota between Feces from CRC Patients, NSCLC Patients, and Controls
3.3. Logistic Regression Analysis of Differentially Increased Bacterial Genera in Patients with CRC or NSCLC
3.4. Comparison of Fecal Microbiota Panels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharma, R. Mapping of global, regional and national incidence, mortality and mortality-to-incidence ratio of lung cancer in 2020 and 2050. Int. J. Clin. Oncol. 2022, 27, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hannun, Y. The importance of extrinsic factors in the development of cancers. Mol. Cell. Oncol. 2016, 3, e1143079. [Google Scholar] [CrossRef]
- El Tekle, G.; Garrett, W.S. Bacteria in cancer initiation, promotion and progression. Nat. Rev. Cancer 2023, 23, 600–618. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Jia, W.; Xie, G.; Jia, W. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut microbiota-derived short-chain fatty acids and colorectal cancer: Ready for clinical translation? Cancer Lett. 2022, 526, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S.; et al. Commensal Microbiota Promote Lung Cancer Development via γδ T Cells. Cell 2019, 176, 998–1013.e16. [Google Scholar] [CrossRef]
- Wang, N.; Fang, J.-Y. Fusobacterium nucleatum, a key pathogenic factor and microbial biomarker for colorectal cancer. Trends Microbiol. 2023, 31, 159–172. [Google Scholar] [CrossRef]
- Riquelme, E.; Zhang, Y.; Zhang, L.; Montiel, M.; Zoltan, M.; Dong, W.; Quesada, P.; Sahin, I.; Chandra, V.; San Lucas, A.; et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell 2019, 178, 795–806.e12. [Google Scholar] [CrossRef]
- Manzoor, S.S.; Doedens, A.; Burns, M.B. The promise and challenge of cancer microbiome research. Genome Biol. 2020, 21, 131. [Google Scholar] [CrossRef]
- Dumont-Leblond, N.; Veillette, M.; Racine, C.; Joubert, P.; Duchaine, C. Non-small cell lung cancer microbiota characterization: Prevalence of enteric and potentially pathogenic bacteria in cancer tissues. PLoS ONE 2021, 16, e0249832. [Google Scholar] [CrossRef] [PubMed]
- Bello, S.; Vengoechea, J.J.; Ponce-Alonso, M.; Figueredo, A.L.; Mincholé, E.; Rezusta, A.; Gambó, P.; Pastor, J.M.; Galeano, J.; del Campo, R. Core Microbiota in Central Lung Cancer with Streptococcal Enrichment as a Possible Diagnostic Marker. Arch. de Bronconeumol. 2021, 57, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cheng, Y.; Zang, D.; Zhang, M.; Li, X.; Liu, D.; Gao, B.; Zhou, H.; Sun, J.; Han, X.; et al. The Role of Gut Microbiota in Lung Cancer: From Carcinogenesis to Immunotherapy. Front. Oncol. 2021, 11, 720842. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.M.; Fidelle, M.; Routy, B.; Kroemer, G.; Wargo, J.A.; Segata, N.; Zitvogel, L. Gut OncoMicrobiome Signatures (GOMS) as next-generation biomarkers for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2023, 20, 583–603. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; An, H.; He, Y.; Fu, G.; Jiang, Z. Comprehensive analysis of microbiota signature across 32 cancer types. Front. Oncol. 2023, 13, 1127225. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Yu, J. Gut microbiota in colorectal cancer development and therapy. Nat. Rev. Clin. Oncol. 2023, 20, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Birla, P.; Shaikh, F.Y. De-“bug”-ing the microbiome in lung cancer. Cancer Metastasis Rev. 2022, 41, 335–346. [Google Scholar] [CrossRef] [PubMed]
- AJCC Cancer Staging Manual [Internet]. Available online: https://link.springer.com/book/9783319406176 (accessed on 19 December 2023).
- Tesolato, S.; Ortega-Hernández, A.; Gómez-Garre, D.; Claver, P.; De Juan, C.; De la Serna, S.; Paz, M.; Domínguez-Serrano, I.; Dziakova, J.; Rivera, D.; et al. Gut microbiota profiles in feces and paired tumor and non-tumor tissues from Colorectal Cancer patients. Relationship to the Body Mass Index. PLoS ONE 2023, 18, e0292551. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Budczies, J.; Klauschen, F.; Sinn, B.V.; Győrffy, B.; Schmitt, W.D.; Darb-Esfahani, S.; Denkert, C. Cutoff Finder: A comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PLoS ONE 2012, 7, e51862. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.R.; DeLong, D.M.; Clarke-Pearson, D.L. Comparing the areas under two or more correlated receiver operating characteristic curves: A nonparametric approach. Biometrics 1988, 44, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Ai, D.; Pan, H.; Li, X.; Gao, Y.; Liu, G.; Xia, L.C. Identifying Gut Microbiota Associated with Colorectal Cancer Using a Zero-Inflated Lognormal Model. Front. Microbiol. 2019, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, R.; Shu, R.; Yu, J.; Li, H.; Long, H.; Jin, S.; Li, S.; Hu, Q.; Yao, F.; et al. Study of the Relationship between Microbiome and Colorectal Cancer Susceptibility Using 16SrRNA Sequencing. Biomed. Res. Int. 2020, 2020, 7828392. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Cheng, L.; Wang, Y.; Zhang, Y.-K.; Zhao, M.-F.; Liang, G.-D.; Zhang, M.-C.; Li, Y.-G.; Zhao, J.-B.; Gao, Y.-N.; et al. Dysbiosis of the Gut Microbiome in Lung Cancer. Front. Cell Infect. Microbiol. 2019, 9, 112. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.P.; Redinbo, M.R.; Bultman, S.J. The Role of the Microbiome in Cancer Development and Therapy. CA Cancer J. Clin. 2017, 67, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Grimes, S.M.; Greer, S.U.; Kubit, M.; Lee, H.; Nadauld, L.D.; Ji, H.P. Characterization of the consensus mucosal microbiome of colorectal cancer. NAR Cancer 2021, 3, zcab049. [Google Scholar] [CrossRef]
- Cullin, N.; Azevedo Antunes, C.; Straussman, R.; Stein-Thoeringer, C.K.; Elinav, E. Microbiome and cancer. Cancer Cell 2021, 39, 1317–1341. [Google Scholar] [CrossRef]
- van Elsland, D.M.; Duijster, J.W.; Zhang, J.; Stévenin, V.; Zhang, Y.; Zha, L.; Xia, Y.; Franz, E.; Sun, J.; Mughini-Gras, L.; et al. Repetitive non-typhoidal Salmonella exposure is an environmental risk factor for colon cancer and tumor growth. Cell Rep. Med. 2022, 3, 100852. [Google Scholar] [CrossRef]
- Qi, Z.; Zhibo, Z.; Jing, Z.; Zhanbo, Q.; Shugao, H.; Weili, J.; Jiang, L.; Shuwen, H. Prediction model of poorly differentiated colorectal cancer (CRC) based on gut bacteria. BMC Microbiol. 2022, 22, 312. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, E.; Avan, A.; Ryzhikov, M.; Ferns, G.; Khazaei, M.; Soleimanpour, S. Role of lactobacillus strains in the management of colorectal cancer: An overview of recent advances. Nutrition 2022, 103–104, 111828. [Google Scholar] [CrossRef]
- Sobhani, I.; Tap, J.; Roudot-Thoraval, F.; Roperch, J.P.; Letulle, S.; Langella, P.; Corthier, G.; Nhieu, J.T.V.; Furet, J.P. Microbial Dysbiosis in Colorectal Cancer (CRC) Patients. PLoS ONE 2011, 6, e16393. [Google Scholar] [CrossRef]
- Zheng, Y.; Fang, Z.; Xue, Y.; Zhang, J.; Zhu, J.; Gao, R.; Yao, S.; Ye, Y.; Wang, S.; Lin, C.; et al. Specific gut microbiome signature predicts the early-stage lung cancer. Gut Microbes 2020, 11, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, K.; Sampath, V.; Umar, S. Bacterial Infections and Cancer: Exploring This Association and Its Implications for Cancer Patients. Int. J. Mol. Sci. 2023, 24, 3110. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, X.-X.; Han, X.; Chen, B.-X.; Zhang, X.-H.; Gao, S.; Xu, D.-Q.; Wang, Y.; Gao, Z.-K.; Yu, L.; et al. Fecal microbiota transplantation inhibits colorectal cancer progression: Reversing intestinal microbial dysbiosis to enhance anti-cancer immune responses. Front. Microbiol. 2023, 14, 1126808. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Hou, S.; Wu, X.; Liu, J.; Wan, X. Analyses of Potential Driver and Passenger Bacteria in Human Colorectal Cancer. Cancer Manag. Res. 2020, 12, 11553–11561. [Google Scholar] [CrossRef]
- Ni, B.; Kong, X.; Yan, Y.; Fu, B.; Zhou, F.; Xu, S. Combined analysis of gut microbiome and serum metabolomics reveals novel biomarkers in patients with early-stage non-small cell lung cancer. Front. Cell Infect. Microbiol. 2023, 13, 1091825. [Google Scholar] [CrossRef]
- Hakozaki, T.; Nolin-Lapalme, A.; Kogawa, M.; Okuma, Y.; Nakamura, S.; Moreau-Amaru, D.; Tamura, T.; Hosomi, Y.; Takeyama, H.; Richard, C.; et al. Cancer Cachexia among Patients with Advanced Non-Small-Cell Lung Cancer on Immunotherapy: An Observational Study with Exploratory Gut Microbiota Analysis. Cancers 2022, 14, 5405. [Google Scholar] [CrossRef]
- Maddi, A.; Sabharwal, A.; Violante, T.; Manuballa, S.; Genco, R.; Patnaik, S.; Yendamuri, S. The microbiome and lung cancer. J. Thorac. Dis. 2019, 11, 280–291. [Google Scholar] [CrossRef]
- Bagheri, Z.; Moeinzadeh, L.; Razmkhah, M. Roles of Microbiota in Cancer: From Tumor Development to Treatment. J. Oncol. 2022, 2022, 3845104. [Google Scholar] [CrossRef] [PubMed]
- Hosmer, D.W.; Lemeshow, S. Area under the ROC curve. In Applied Logistic Regression, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2000; pp. 160–164. [Google Scholar]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- NIH Human Microbiome Project—HMQCP. Available online: https://hmpdacc.org/HMQCP/ (accessed on 16 January 2024).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | CRC Group (N = 38) | NSCLC Group (N = 19) | Control Group (N = 20) | p Value (CRC vs. Control) | p Value (NSCLC vs. Control) |
|---|---|---|---|---|---|
| Age (Mean ± SD, years) | 71.24 ± 12 | 72.79 ± 7.91 | 54.80 ± 14.97 | <0.001 1,* | <0.001 2,* |
| Gender, N (%) | 0.016 3,* | 0.265 3 | |||
| Male | 24 (63.2) | 9 (47.4) | 6 (30) | ||
| Female | 14 (36.8) | 10 (52.6) | 14 (70) | ||
| BMI group, N (%) | 0.004 3,* | 0.008 3,* | |||
| Normal weight (BMI < 25 kg/m2) | 8 (21.1) | 6 (31.6) | 3 (15.0) | ||
| Overweight (BMI ≥ 25 kg/m2 and <30 kg/m2) | 20 (52.6) | 9 (47.4) | 3 (15.0) | ||
| Obesity (BMI ≥ 30 kg/m2) | 10 (26.3) | 4 (21.1) | 14 (70.0) | ||
| Tumor location, N (%) | |||||
| Right colon | 20 (52.6) | ||||
| Left colon | 13 (34.2) | ||||
| Rectum | 5 (13.2) | ||||
| TNM stage, N (%) | |||||
| I–II | 17 (44.7) | 13 (68.4) | |||
| III–IV | 21 (55.3) | 6 (31.6) |
| (a) | (b) | ||
| Bacterial Genus | p Values (LEfSe) | Bacterial Genus | p Values (LEfSe) |
| Control group | |||
| g__Sutterella | 0.019 | ||
| g__Blautia | 0.005 | Control group | |
| g__Lachnoclostridium | 0.044 | g__Lachnoclostridium | 0.031 |
| g__Dielma | 0.018 | g__Megasphaera | 0.040 |
| g__Olsenella | 0.024 | g__Olsenella | 0.020 |
| CRC group | NSCLC group | ||
| g__Campylobacter | 0.004 | g_Hungatella | 0.009 |
| g__Intestinimonas | 0.049 | g_Turicibacter | 0.013 |
| g__Parvimonas | 0.004 | g_Coprobacillus | 0.007 |
| g__Turicibacter | 0.006 | g_Clostridium sensu stricto 3 | 0.015 |
| g__Hydrogenoanaerobacterium | 0.008 | g_Intestinimonas | 0.038 |
| g__Gemella | 0.016 | g_Eisenbergiella | 0.028 |
| g__Eisenbergiella | 0.008 | g_Salmonella | 0.015 |
| g__Clostridium sensu stricto 1 | 0.021 | g_DTU089 | 0.034 |
| g__Peptostreptococcus | 0.006 | g_Frisingicoccus | 0.040 |
| g__S5-A14a | 0.023 | g_Granulicatella | 0.035 |
| g__Ruminococcaceae | 0.022 | g_Incertae Sedis | 0.010 |
| g__Lactobacillus | 0.024 | g__Lactobacillus | 0.017 |
| g__Salmonella | 0.011 | g__Agathobacter | 0.038 |
| g__Eikenella | 0.034 | ||
| g__CAG-352 | 0.035 | ||
| g__Fusobacterium | 0.003 | ||
| g__Escherichia-Shigella | 0.018 | ||
| (c) | |||
| Bacterial Genus | p Values (LEfSe) | ||
| NSCLC group | |||
| g__Bifidobacterium | 0.005 | ||
| g__Agathobacter | 0.004 | ||
| g__Roseburia | 0.050 | ||
| g__Blautia | 0.009 | ||
| g__CAG-873 | 0.029 | ||
| g__Incertae Sedis | 0.039 | ||
| g__Oscillibacter | 0.005 | ||
| g__Ammoniphilus | 0.023 | ||
| g__Pseudobutyrivibrio | 0.005 | ||
| CRC group | |||
| g__Campylobacter | 0.030 | ||
| g__Ruminococcaceae | 0.042 | ||
| g__Hydrogenoanaerobacterium | 0.016 | ||
| g__Sellimonas | 0.021 | ||
| g__Solobacterium | 0.002 | ||
| g__Sarcina | 0.034 | ||
| g__Parvimonas | 0.005 | ||
| g__Peptostreptococcus | 0.003 | ||
| g__Eikenella | 0.033 | ||
| g__CAG-352 | 0.037 | ||
| g__Fusobacterium | 0.004 |
| CRC Bacterial Genera (Abundance Threshold) | Group | Unadjusted OR (95% CI) | Unadjusted p Value | Adjusted OR (95% CI) | Adjusted p Value |
|---|---|---|---|---|---|
| Campylobacter (0.003313) | Control | reference | - | reference | - |
| Cancer | 6.13 (1.72–21.9) | 0.005 * | 4.44 (0.88–22.4) | 0.071 | |
| Intestinimonas (0.01827) | Control | reference | - | reference | - |
| Cancer | 2.35 (0.78–7.11) | 0.130 | 1.34 (0.28–6.32) | 0.705 | |
| Parvimonas (0.001859) | Control | reference | - | reference | - |
| Cancer | 10.9 (2.69–44.1) | 0.001 * | 53.3 (3.26–870.5) | 0.005 * | |
| Turicibacter (0.01017) | Control | reference | - | reference | - |
| Cancer | 6.13 (1.72–21.9) | 0.005 * | 3.98 (0.83–19.0) | 0.084 | |
| Hydrogenoanaerobacterium (0.00367) | Control | reference | - | reference | - |
| Cancer | 4.56 (1.44–14.5) | 0.010 * | 1.42 (0.295–6.82) | 0.662 | |
| Gemella (0.006006) | Control | reference | - | reference | - |
| Cancer | 3.68 (1.18–11.5) | 0.025 * | 6.01 (1.20–30.0) | 0.029 * | |
| Eisenbergiella (0.008012) | Control | reference | - | reference | - |
| Cancer | 4.56 (1.44–14.5) | 0.010 * | 5.35 (1.08–26.5) | 0.040 * | |
| Clostridium sensu stricto 1 (0.08304) | Control | reference | - | reference | - |
| Cancer | 7.79 (1.95–31.2) | 0.004 * | 4.98 (0.93–26.6) | 0.061 | |
| Peptostreptococcus (0.0003669) | Control | reference | - | reference | - |
| Cancer | 4.13 (1.24–13.7) | 0.021 * | 9.42 (1.38–64.2) | 0.022 * | |
| S5-A14a (0.0003232) | Control | reference | - | reference | - |
| Cancer | 6.55 (1.32–32.3) | 0.021 * | 2.54 (0.41–15.8) | 0.318 | |
| Ruminococcaceae (0.004992) | Control | reference | - | reference | - |
| Cancer | 8.5 (2.19–33.0) | 0.002 * | 3.67 (0.62–21.7) | 0.151 | |
| Lactobacillus (0.04309) | Control | reference | - | reference | - |
| Cancer | 5.67 (1.42–22.6) | 0.014 * | 6.72 (1.05–43.0) | 0.044 * | |
| Salmonella (0.00081) | Control | reference | - | reference | - |
| Cancer | 8.22 (2.40–28.2) | 0.001 * | 5.44 (1.02–28.8) | 0.047 * | |
| Eikenella (0.0001615) | Control | reference | - | reference | - |
| Cancer | 7.74 (0.92–65.1) | 0.060 | 8.20 (0.73–92.1) | 0.088 | |
| CAG-352 (0.008875) | Control | reference | - | reference | - |
| Cancer | 3.42 (1.10–10.7) | 0.034 * | 2.14 (0.47–9.61) | 0.323 | |
| Fusobacterium (0.01714) | Control | reference | - | reference | - |
| Cancer | 11.2 (3.02–41.6) | 0.000 * | 78.9 (4.48–1389.0) | 0.003 * | |
| Escherichia_Shigella (0.1569) | Control | reference | - | reference | - |
| Cancer | 5.2 (1.61–16.7) | 0.006 * | 3.81 (0.80–18.2) | 0.094 |
| NSCLC Bacterial Genera (Abundance Threshold) | Group | Unadjusted OR (95% CI) | Unadjusted p Value | Adjusted OR (95% CI) | Adjusted p Value |
|---|---|---|---|---|---|
| Hungatella (0.002103) | Control | reference | - | reference | - |
| Cancer | 6.53 (1.61–26.5) | 0.009 * | 7.17 (0.92–56.0) | 0.060 | |
| Turicibacter (0.006865) | Control | reference | - | reference | - |
| Cancer | 5.20 (1.32–20.5) | 0.019 * | 4.61 (0.73–29.1) | 0.104 | |
| Coprobacillus (0.0006056) | Control | reference | - | reference | - |
| Cancer | 6.86 (1.63–28.9) | 0.009 * | 7.00 (0.59–82.9) | 0.123 | |
| Clostridium sensu stricto 3 (0.0004242) | Control | reference | - | reference | - |
| Cancer | 5.50 (1.32–22.9) | 0.019 * | 6.11 (0.84–44.5) | 0.074 | |
| Intestinimonas (0.0208) | Control | reference | - | reference | - |
| Cancer | 5.63 (1.36–23.3) | 0.017 * | 3.50 (0.45–27.3) | 0.232 | |
| Eisenbergiella (0.007163) | Control | reference | - | reference | - |
| Cancer | 4.02 (1.06–15.3) | 0.041 * | 5.49 (0.69–43.9) | 0.108 | |
| Salmonella (0.0008436) | Control | reference | - | reference | - |
| Cancer | 15.8 (2.80–89.0) | 0.002 * | 6.06 (0.73–50.4) | 0.096 | |
| DTU089 (0.007431) | Control | reference | - | reference | - |
| Cancer | 5.50 (1.32–22.9) | 0.019 * | 20.1 (1.35–300.1) | 0.029 * | |
| Frisingicoccus (0.0004159) | Control | reference | - | reference | - |
| Cancer | 5.06 (1.30–19.7) | 0.020 * | 2.88 (0.49–17.1) | 0.244 | |
| Granullicatella (0.008393) | Control | reference | - | reference | - |
| Cancer | 4.02 (1.06–15.3) | 0.041 * | 4.81 (0.63–36.6) | 0.129 | |
| Incertae Sedis (0.1568) | Control | reference | - | reference | - |
| Cancer | 8.00 (1.74–36.7) | 0.007 * | 160.1 (2.44–10,506.36) | 0.017 * | |
| Lactobacillus (0.03697) | Control | reference | - | reference | - |
| Cancer | 4.13 (1.06–16.1) | 0.041 * | 2.07 (0.36–11.8) | 0.414 | |
| Agathobacter (0.5153) | Control | reference | - | reference | - |
| Cancer | 5.625 (1.36–23.3) | 0.017 * | 6.94 (0.97–49.6) | 0.053 |
| Panel | Bacterial Taxa Included | AUC (95% CI) | Se (95% CI) | Sp (95% CI) | p Value |
|---|---|---|---|---|---|
| CRC (proposed) | 7 a | 0.840 (0.720–0.923) | 78.9 (63.7–88.9) | 80.0 (58.4–91.9) | - |
| Thomas et al. [15] | 34 c | 0.557 (0.420–0.687) | 60.5 (44.7–74.4) | 60.0 (38.7–78.1) | 0.004 * |
| Yang et al. [16] | 43 d | 0.511 (0.376–0.644) | 47.4 (32.5–62.7) | 65.0 (43.3–81.9) | 0.015 * |
| NSCLC (proposed) | 2 b | 0.747 (0.583–0.872) | 73.7 (51.2–88.2) | 75.0 (53.1–88.8) | - |
| Thomas et al. [15] | 34 c | 0.447 (0.288–0.615) | 36.8 (19.1–59.0) | 65.0 (43.3–81.3) | 0.038 * |
| Yang et al. [16] | 43 d | 0.413 (0.258–0.582) | 47.4 (27.3–68.3) | 45.0 (25.8–65.8) | 0.002 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesolato, S.; Vicente-Valor, J.; Paz-Cabezas, M.; Gómez-Garre, D.; Sánchez-González, S.; Ortega-Hernández, A.; de la Serna, S.; Domínguez-Serrano, I.; Dziakova, J.; Rivera, D.; et al. Gut Microbiota Signatures with Potential Clinical Usefulness in Colorectal and Non-Small Cell Lung Cancers. Biomedicines 2024, 12, 703. https://doi.org/10.3390/biomedicines12030703
Tesolato S, Vicente-Valor J, Paz-Cabezas M, Gómez-Garre D, Sánchez-González S, Ortega-Hernández A, de la Serna S, Domínguez-Serrano I, Dziakova J, Rivera D, et al. Gut Microbiota Signatures with Potential Clinical Usefulness in Colorectal and Non-Small Cell Lung Cancers. Biomedicines. 2024; 12(3):703. https://doi.org/10.3390/biomedicines12030703
Chicago/Turabian StyleTesolato, Sofía, Juan Vicente-Valor, Mateo Paz-Cabezas, Dulcenombre Gómez-Garre, Silvia Sánchez-González, Adriana Ortega-Hernández, Sofía de la Serna, Inmaculada Domínguez-Serrano, Jana Dziakova, Daniel Rivera, and et al. 2024. "Gut Microbiota Signatures with Potential Clinical Usefulness in Colorectal and Non-Small Cell Lung Cancers" Biomedicines 12, no. 3: 703. https://doi.org/10.3390/biomedicines12030703