

The Phenomenon of Antiretroviral Drug Resistance in the Context of Human Immunodeficiency Virus Treatment: Dynamic and Ever Evolving Subject Matter


 ,
,  and
and
Abstract
:1. Introduction
2. HIV Structure and Genome
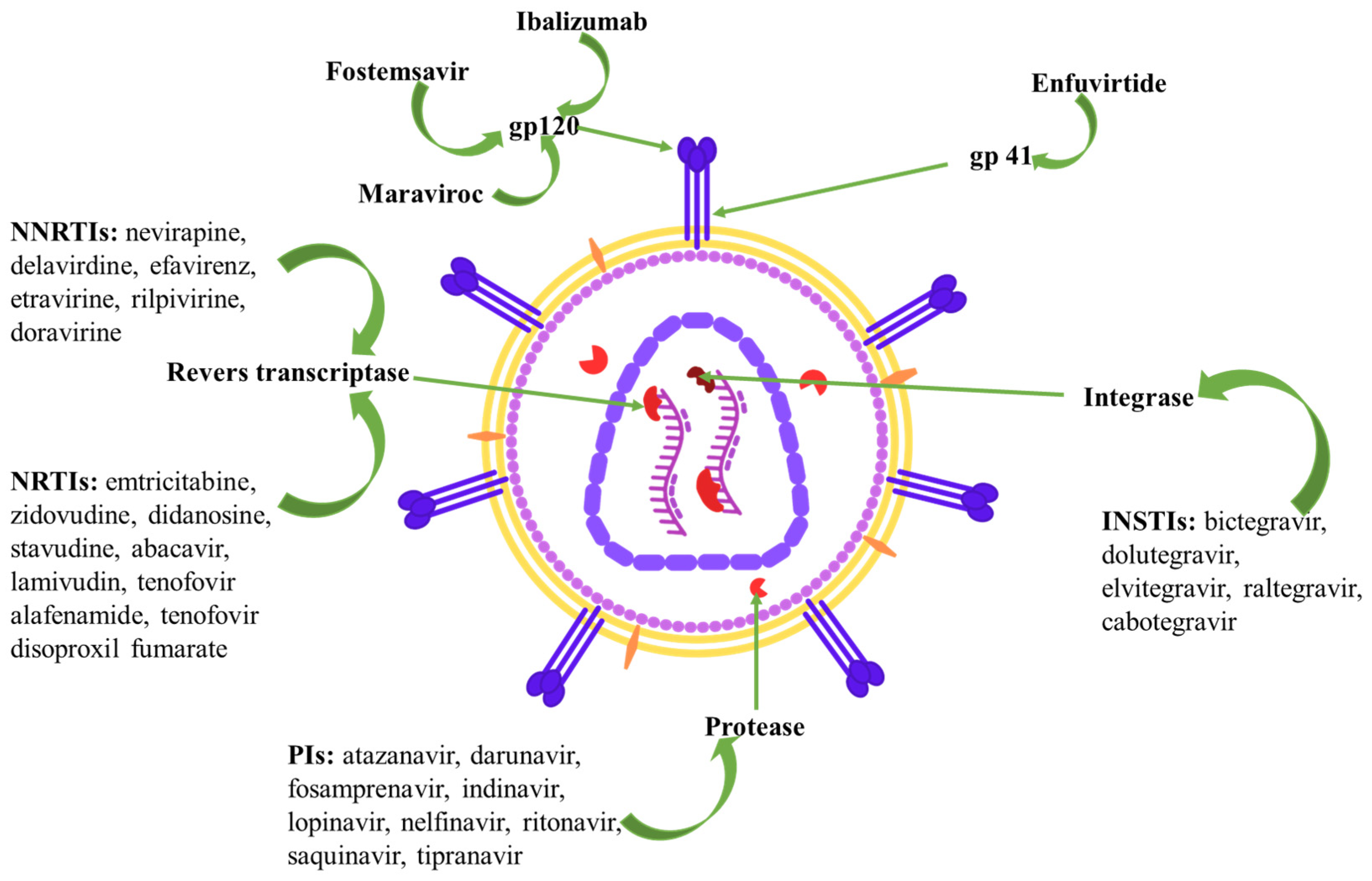
3. HIV Therapeutic Targets and Current Pharmacological Approaches
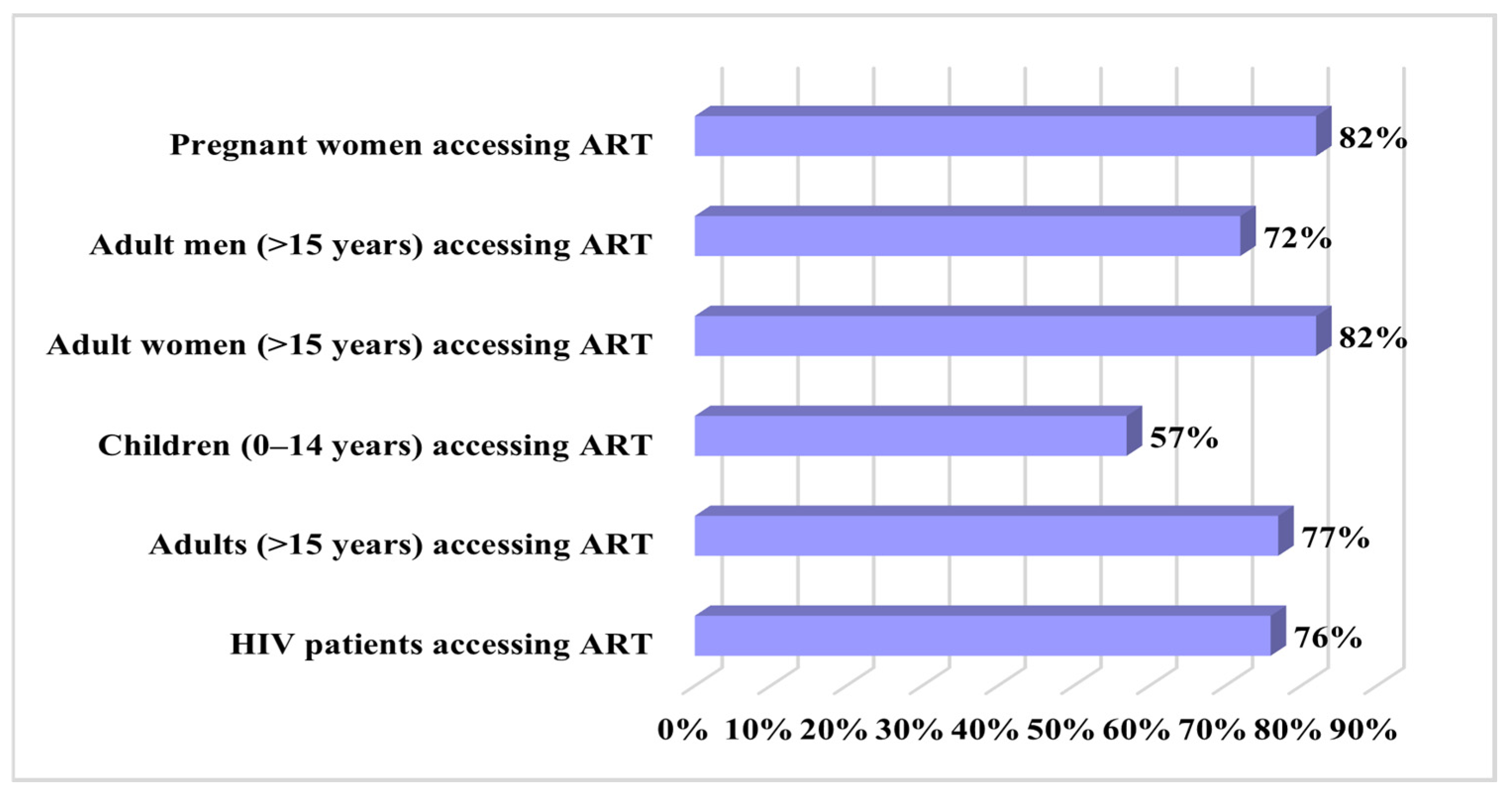
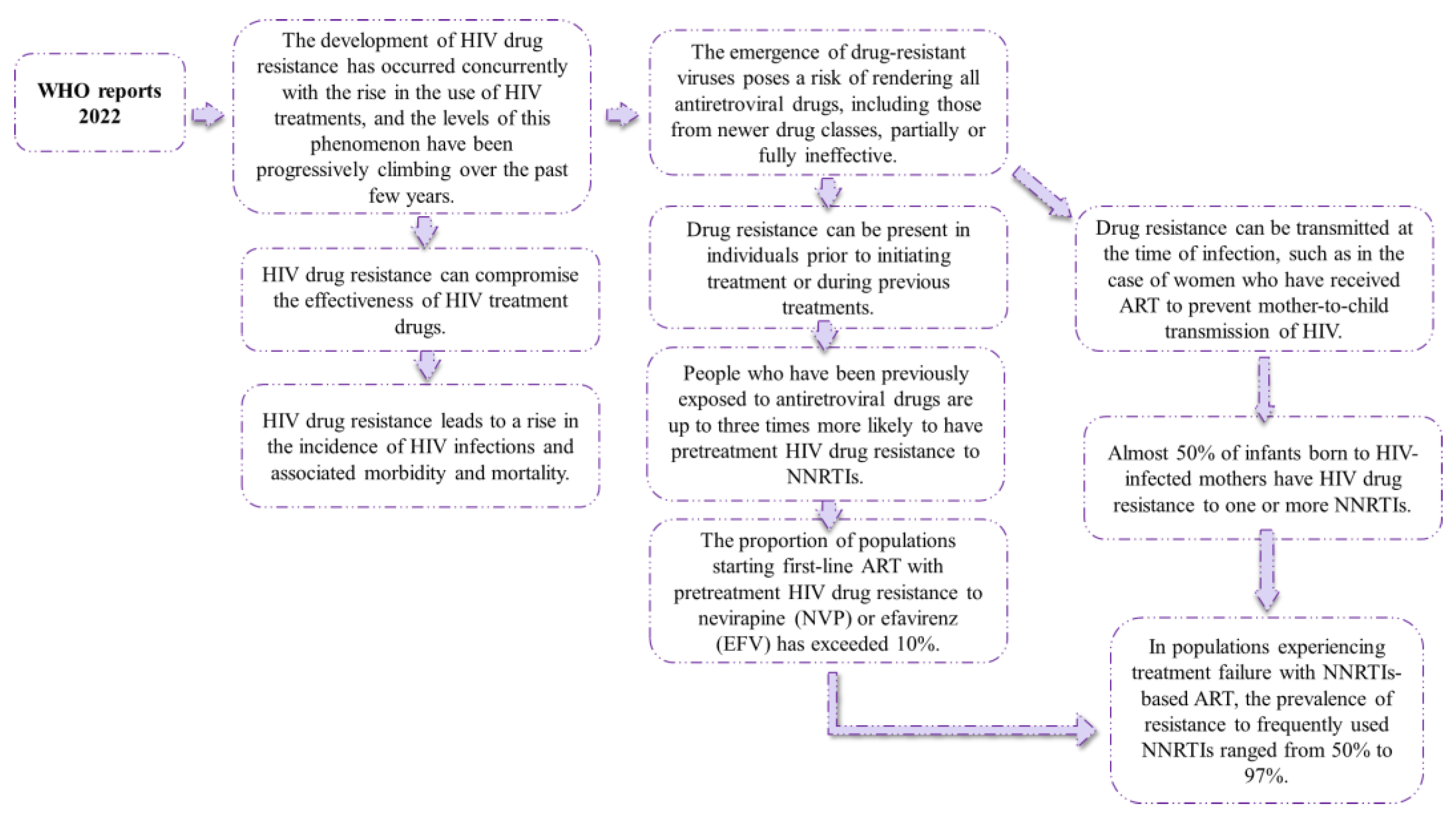
4. HIV Drug Resistance Data
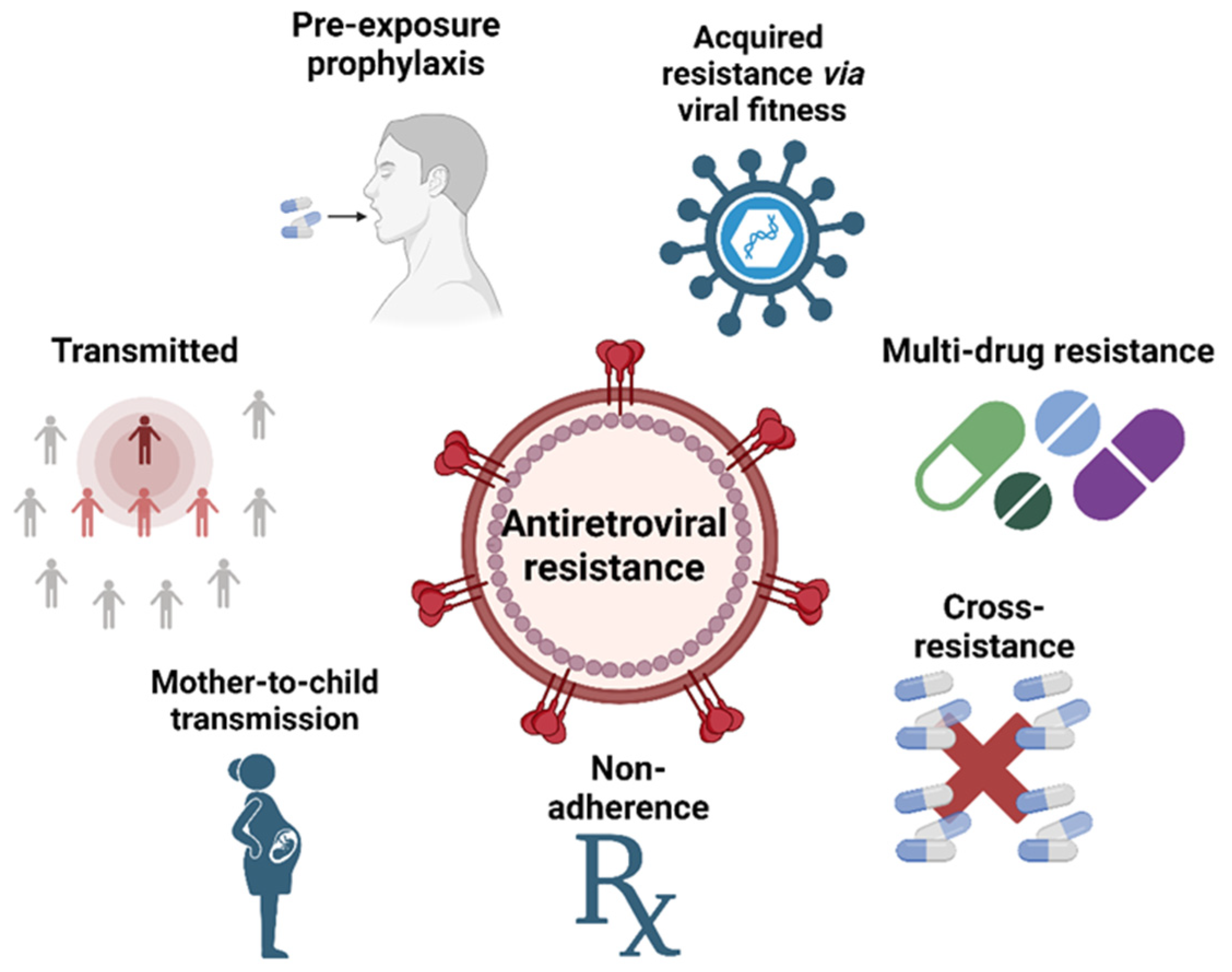
5. HIV Drug Resistance Mechanisms
5.1. Transmitted HIV Drug Resistance
5.2. Acquired HIV Drug Resistance via Viral Mutations
5.3. Cross-Resistance
5.4. Multidrug Resistance (MDR HIV)
5.5. Nonadherence in Drug Resistance
5.6. Mother-to-Child Drug Resistance
5.7. Preexposure Prophylaxis
6. Challenges and Opportunities in Overcoming HIV Drug Resistance
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The Joint United Nations Programme on HIV/AIDS (UNAIDS). Aids in Danger: UNAIDS Global AIDS Update 2022; UNAIDS: Geneva, Switzerland, 2022. [Google Scholar]
- Faria, N.R.; Rambaut, A.; Suchard, M.A.; Baele, G.; Bedford, T.; Ward, M.J.; Tatem, A.J.; Sousa, J.D.; Arinaminpathy, N.; Pépin, J.; et al. The Early Spread and Epidemic Ignition of HIV-1 in Human Populations. Science 2014, 346, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Nastri, B.M.; Pagliano, P.; Zannella, C.; Folliero, V.; Masullo, A.; Rinaldi, L.; Galdiero, M.; Franci, G. HIV and Drug-Resistant Subtypes. Microorganisms 2023, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Nyamweya, S.; Hegedus, A.; Jaye, A.; Rowland-Jones, S.; Flanagan, K.L.; Macallan, D.C. Comparing HIV-1 and HIV-2 Infection: Lessons for Viral Immunopathogenesis. Rev. Med. Virol. 2013, 23, 221–240. [Google Scholar] [CrossRef] [PubMed]
- Esbjörnsson, J.; Jansson, M.; Jespersen, S.; Månsson, F.; Hønge, B.L.; Lindman, J.; Medina, C.; da Silva, Z.J.; Norrgren, H.; Medstrand, P.; et al. HIV-2 as a Model to Identify a Functional HIV Cure. AIDS Res. Ther. 2019, 16, 24. [Google Scholar] [CrossRef] [PubMed]
- Rawson, J.M.O.; Landman, S.R.; Reilly, C.S.; Mansky, L.M. HIV-1 and HIV-2 Exhibit Similar Mutation Frequencies and Spectra in the Absence of G-to-A Hypermutation. Retrovirology 2015, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- McCutchan, F.E. Understanding the Genetic Diversity of HIV-1. AIDS 2000, 14 (Suppl. S3), S31–S44. [Google Scholar] [PubMed]
- Korber, B.; Gaschen, B.; Yusim, K.; Thakallapally, R.; Kesmir, C.; Detours, V. Evolutionary and Immunological Implications of Contemporary HIV-1 Variation. Br. Med. Bull. 2001, 58, 19–42. [Google Scholar] [CrossRef]
- Oguntibeju, O. Quality of Life of People Living with HIV and AIDS and Antiretroviral Therapy. HIV/AIDS—Res. Palliat. Care 2012, 4, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Verma, A.; Kashyap, M.; Gautam, P. ART in Prevention of Mother-to-Child Transmission of HIV. J. Obstet. Gynecol. India 2020, 70, 18–22. [Google Scholar] [CrossRef]
- Mugwaneza, P.; Lyambabaje, A.; Umubyeyi, A.; Humuza, J.; Tsague, L.; Mwanyumba, F.; Mutabazi, V.; Nsanzimana, S.; Ribakare, M.; Irakoze, A.; et al. Impact of Maternal ART on Mother-to-Child Transmission (MTCT) of HIV at Six Weeks Postpartum in Rwanda. BMC Public Health 2018, 18, 1248. [Google Scholar] [CrossRef]
- Ciaranello, A.L.; Seage, G.R.; Freedberg, K.A.; Weinstein, M.C.; Lockman, S.; Walensky, R.P. Antiretroviral Drugs for Preventing Mother-to-Child Transmission of HIV in Sub-Saharan Africa: Balancing Efficacy and Infant Toxicity. AIDS 2008, 22, 2359–2369. [Google Scholar] [CrossRef]
- Campbell, E.M.; Hope, T.J. HIV-1 Capsid: The Multifaceted Key Player in HIV-1 Infection. Nat. Rev. Microbiol. 2015, 13, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, J.I. Morphogenesis of the Infectious HIV-1 Virion. Front. Microbiol. 2011, 2, 242. [Google Scholar] [CrossRef]
- Fanales-Belasio, E.; Raimondo, M.; Suligoi, B.; Buttò, S. HIV Virology and Pathogenetic Mechanisms of Infection: A Brief Overview. Ann. Ist. Super. Sanita 2010, 46, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Kalinichenko, S.; Komkov, D.; Mazurov, D. HIV-1 and HTLV-1 Transmission Modes: Mechanisms and Importance for Virus Spread. Viruses 2022, 14, 152. [Google Scholar] [CrossRef] [PubMed]
- Seitz, R. Human Immunodeficiency Virus (HIV). Transfus. Med. Hemotherapy 2016, 43, 203–222. [Google Scholar] [CrossRef]
- Li, G.; Piampongsant, S.; Faria, N.R.; Voet, A.; Pineda-Peña, A.-C.; Khouri, R.; Lemey, P.; Vandamme, A.-M.; Theys, K. An Integrated Map of HIV Genome-Wide Variation from a Population Perspective. Retrovirology 2015, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- van Heuvel, Y.; Schatz, S.; Rosengarten, J.F.; Stitz, J. Infectious RNA: Human Immunodeficiency Virus (HIV) Biology, Therapeutic Intervention, and the Quest for a Vaccine. Toxins 2022, 14, 138. [Google Scholar] [CrossRef]
- Shah, S.; Alexaki, A.; Pirrone, V.; Dahiya, S.; Nonnemacher, M.R.; Wigdahl, B. Functional Properties of the HIV-1 Long Terminal Repeat Containing Single-Nucleotide Polymorphisms in Sp Site III and CCAAT/Enhancer Binding Protein Site I. Virol. J. 2014, 11, 92. [Google Scholar] [CrossRef]
- Levy, J.A.; Steele, F.R. Hiv and the Pathogenesis of Aids. Nat. Med. 1995, 1, 273. [Google Scholar] [CrossRef]
- Kuiken, C.; Foley, B.; Leitner, T.; Apetrei, C.; Hahn, B.; Mizrachi, I.; Mullins, J.; Rambaut, A.; Wolinsky, S.; Korber, B. HIV Sequence Compendium 2010; Los Alamos National Laboratory: Los Alamos, NM, USA, 2010. [Google Scholar]
- Lu, K.; Heng, X.; Summers, M.F. Structural Determinants and Mechanism of HIV-1 Genome Packaging. J. Mol. Biol. 2011, 410, 609–633. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.; Nava, B.; Caputi, M. Tat Is a Multifunctional Viral Protein That Modulates Cellular Gene Expression and Functions. Oncotarget 2017, 8, 27569–27581. [Google Scholar] [CrossRef] [PubMed]
- Charnay, N.; Ivanyi-Nagy, R.; Soto-Rifo, R.; Ohlmann, T.; López-Lastra, M.; Darlix, J.-L. Mechanism of HIV-1 Tat RNA Translation and Its Activation by the Tat Protein. Retrovirology 2009, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Alcamí, J. Ciclo Replicativo Del VIH. Dianas Terapéuticas Consolidadas y Dianas Potenciales. Enferm. Infecc. Microbiol. Clin. 2008, 26, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, F. HIV Life Cycle: Overview. In Encyclopedia of AIDS; Springer: New York, NY, USA, 2013; pp. 1–9. [Google Scholar]
- Mann, J.K.; Byakwaga, H.; Kuang, X.T.; Le, A.Q.; Brumme, C.J.; Mwimanzi, P.; Omarjee, S.; Martin, E.; Lee, G.Q.; Baraki, B.; et al. Ability of HIV-1 Nef to Downregulate CD4 and HLA Class I Differs among Viral Subtypes. Retrovirology 2013, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Malim, M.H.; Emerman, M. HIV-1 Accessory Proteins—Ensuring Viral Survival in a Hostile Environment. Cell Host Microbe 2008, 3, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Hokello, J.; Tyagi, K.; Owor, R.O.; Sharma, A.L.; Bhushan, A.; Daniel, R.; Tyagi, M. New Insights into HIV Life Cycle, Th1/Th2 Shift during HIV Infection and Preferential Virus Infection of Th2 Cells: Implications of Early HIV Treatment Initiation and Care. Life 2024, 14, 104. [Google Scholar] [CrossRef]
- Patel, K.; Zhang, A.; Zhang, M.H.; Bunachita, S.; Baccouche, B.M.; Hundal, H.; Lavado, L.K.; Agarwal, A.; Malik, P.; Patel, U.K. Forty Years Since the Epidemic: Modern Paradigms in HIV Diagnosis and Treatment. Cureus 2021, 31, e14805. [Google Scholar] [CrossRef] [PubMed]
- McNeil, R.; Kerr, T.; Coleman, B.; Maher, L.; Milloy, M.J.; Small, W. Antiretroviral Therapy Interruption among HIV Postive People Who Use Drugs in a Setting with a Community-Wide HIV Treatment-as-Prevention Initiative. AIDS Behav. 2017, 21, 402–409. [Google Scholar] [CrossRef]
- Tsuda, H.; Koga, M.; Nojima, M.; Senkoji, T.; Kubota, M.; Kikuchi, T.; Adachi, E.; Ikeuchi, K.; Tsutsumi, T.; Koibuchi, T.; et al. Changes in Survival and Causes of Death among People Living with HIV: Three Decades of Surveys from Tokyo, One of the Asian Metropolitan Cities. J. Infect. Chemother. 2021, 27, 949–956. [Google Scholar] [CrossRef]
- Deeks, S.G.; Lewin, S.R.; Havlir, D. V The End of AIDS: HIV Infection as a Chronic Disease. Lancet 2013, 382, 1525–1533. [Google Scholar] [CrossRef]
- Masters, M.C.; Krueger, K.M.; Williams, J.L.; Morrison, L.; Cohn, S.E. Beyond One Pill, Once Daily: Current Challenges of Antiretroviral Therapy Management in the United States. Expert Rev. Clin. Pharmacol. 2019, 12, 1129–1143. [Google Scholar] [CrossRef] [PubMed]
- WHO. Consolidated Guidelines on HIV Prevention, Testing, Treatment, Service Delivery and Monitoring: Recommendations for a Public Health Approach; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Pomerantz, R.J.; Horn, D.L. Twenty Years of Therapy for HIV-1 Infection. Nat. Med. 2003, 9, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Pettit, S.C.; Moody, M.D.; Wehbie, R.S.; Kaplan, A.H.; Nantermet, P.V.; Klein, C.A.; Swanstrom, R. The P2 Domain of Human Immunodeficiency Virus Type 1 Gag Regulates Sequential Proteolytic Processing and Is Required to Produce Fully Infectious Virions. J. Virol. 1994, 68, 8017–8027. [Google Scholar] [CrossRef] [PubMed]
- Konvalinka, J.; Kräusslich, H.G.; Müller, B. Retroviral Proteases and Their Roles in Virion Maturation. Virology 2015, 479–480, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Oroszlan, S.; Luftig, R.B. Retroviral Proteinases. Curr. Top. Microbiol. Immunol. 1990, 157, 153–189. [Google Scholar]
- Li, P.; Stephenson, A.J.; Brennan, P.A.; Karageorgos, L.; Kok, T.; Kuiper, L.J.; Swift, L.J.; Burrell, C.J. Initiation of Reverse Transcription during Cell-to-Cell Transmission of Human Immunodeficiency Virus Infection Uses Pre-Existing Reverse Transcriptase. J. Gen. Virol. 1994, 75, 1917–1926. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, D.; Evans, D.B.; Deibel, M.R.; Vosters, A.F.; Eckenrode, F.M.; Einspahr, H.M.; Hui, J.O.; Tomasselli, A.G.; Zurcher-Neely, H.A.; Heinrikson, R.L.; et al. Purification and Characterization of Heterodimeric Human Immunodeficiency Virus Type 1 (HIV-1) Reverse Transcriptase Produced by in Vitro Processing of P66 with Recombinant HIV-1 Protease. J. Biol. Chem. 1992, 267, 14227–14232. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.D.; Farnet, C.M.; Bushman, F.D. Human Immunodeficiency Virus Type 1 Preintegration Complexes: Studies of Organization and Composition. J. Virol. 1997, 71, 5382–5390. [Google Scholar] [CrossRef]
- Fujiwara, T.; Mizuuchi, K. Retroviral DNA Integration: Structure of an Integration Intermediate. Cell 1988, 54, 497–504. [Google Scholar] [CrossRef]
- Brown, P.O.; Bowerman, B.; Varmus, H.E.; Bishop, J.M. Retroviral Integration: Structure of the Initial Covalent Product and Its Precursor, and a Role for the Viral IN Protein. Proc. Natl. Acad. Sci. USA 1989, 86, 2525–2529. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.; Mizuuchi, K.; Craigie, R. HIV-1 DNA Integration: Mechanism of Viral DNA Cleavage and DNA Strand Transfer. Cell 1991, 67, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.; Hughes, S.; Varmus, H. The Interactions of Retroviruses and Their Hosts. In Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997; ISBN 0879695714. [Google Scholar]
- Engelman, A.; Cherepanov, P. The Structural Biology of HIV-1: Mechanistic and Therapeutic Insights. Nat. Rev. Microbiol. 2012, 10, 279–290. [Google Scholar] [CrossRef]
- Engelman, A. The Roles of Cellular Factors in Retroviral Integration. Curr. Top. Microbiol. Immunol. 2003, 281, 209–238. [Google Scholar] [PubMed]
- Yoder, K.E.; Bushman, F.D. Repair of Gaps in Retroviral DNA Integration Intermediates. J. Virol. 2000, 74, 11191–11200. [Google Scholar] [CrossRef] [PubMed]
- Usach, I.; Melis, V.; Peris, J.-E. Non-Nucleoside Reverse Transcriptase Inhibitors: A Review on Pharmacokinetics, Pharmacodynamics, Safety and Tolerability. J. Int. AIDS Soc. 2013, 16, 18567. [Google Scholar] [CrossRef] [PubMed]
- Rizza, S.A.; Bhatia, R.; Zeuli, J.; Temesgen, Z. Ibalizumab for the Treatment of Multidrug-Resistant HIV-1 Infection. Drugs Today 2019, 55, 25. [Google Scholar] [CrossRef] [PubMed]
- Montessori, V.; Press, N.; Harris, M.; Akagi, L.; Montaner, J.S.G. Adverse Effects of Antiretroviral Therapy for HIV Infection. CMAJ 2004, 170, 229–238. [Google Scholar] [PubMed]
- Margolis, A.M.; Heverling, H.; Pham, P.A.; Stolbach, A. A Review of the Toxicity of HIV Medications. J. Med. Toxicol. 2014, 10, 26–39. [Google Scholar] [CrossRef]
- Eckhardt, B.J.; Gulick, R.M. Drugs for HIV Infection. In Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1293–1308.e2. [Google Scholar]
- European AIDS Clinical Society. EACS Guidelines 2022; Version 11.1; European AIDS Clinical Society: Brussels, Belgium, 2022. [Google Scholar]
- FDA Cabenuva (Cabotegravir Extended-Release Injectable Suspension; Rilpivirine Extended-Release Injectable Suspension), Co-Packaged for Intramuscular Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2021/212888s000lbl.pdf (accessed on 4 September 2023).
- Durham, S.H.; Chahine, E.B. Cabotegravir-Rilpivirine: The First Complete Long-Acting Injectable Regimen for the Treatment of HIV-1 Infection. Ann. Pharmacother. 2021, 55, 1397–1409. [Google Scholar] [CrossRef]
- Patel, P.H.; Zulfiqar, H. Reverse Transcriptase Inhibitors. In Frontiers in HIV Research; StatPearls Publishing: St. Petersburg, FL, USA, 2015; pp. 44–61. [Google Scholar]
- Bazzoli, C.; Jullien, V.; Le Tiec, C.; Rey, E.; Mentré, F.; Taburet, A.M. Intracellular Pharmacokinetics of Antiretroviral Drugs in HIV-Infected Patients, and Their Correlation with Drug Action. Clin. Pharmacokinet. 2010, 49, 17–45. [Google Scholar] [CrossRef]
- Gao, W.Y.; Shirasaka, T.; Johns, D.G.; Broder, S.; Mitsuya, H. Differential Phosphorylation of Azidothymidine, Dideoxycytidine, and Dideoxyinosine in Resting and Activated Peripheral Blood Mononuclear Cells. J. Clin. Investig. 1993, 91, 2326–2333. [Google Scholar] [CrossRef] [PubMed]
- Robbins, B.L.; Wilcox, C.K.; Fridland, A.; Rodman, J.H. Metabolism of Tenofovir and Didanosine in Quiescent or Stimulated Human Peripheral Blood Mononuclear Cells. Pharmacotherapy 2003, 23, 695–701. [Google Scholar] [CrossRef]
- Arts, E.J.; Hazuda, D.J. HIV-1 Antiretroviral Drug Therapy. Cold Spring Harb. Perspect. Med. 2012, 2, a007161. [Google Scholar] [CrossRef]
- De Clercq, E. The Design of Drugs for HIV and HCV. Nat. Rev. Drug Discov. 2007, 6, 1001–1018. [Google Scholar] [CrossRef]
- Martins, S.; Ramos, M.; Fernandes, P. The Current Status of the NNRTI Family of Antiretrovirals Used Against HIV Infection. Curr. Med. Chem. 2008, 15, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Chen, X.; Li, D.; Fang, Z.; De Clercq, E.; Liu, X. HIV-1 NNRTIs: Structural Diversity, Pharmacophore Similarity, and Impliations for Drug Design. Med. Res. Rev. 2013, 33, E1–E72. [Google Scholar] [CrossRef]
- Wang, Y.; De Clercq, E.; Li, G. Current and Emerging Non-Nucleoside Reverse Transcriptase Inhibitors (NNRTIs) for HIV-1 Treatment. Expert Opin. Drug Metab. Toxicol. 2019, 15, 813–829. [Google Scholar] [CrossRef]
- Zhuang, C.; Pannecouque, C.; De Clercq, E.; Chen, F. Development of Non-Nucleoside Reverse Transcriptase Inhibitors (NNRTIs): Our Past Twenty Years. Acta Pharm. Sin. B 2020, 10, 961–978. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Martinez, S.E.; DeStefano, J.J.; Arnold, E. Structure of HIV-1 RT/DsRNA Initiation Complex Prior to Nucleotide Incorporation. Proc. Natl. Acad. Sci. USA 2019, 116, 7308–7313. [Google Scholar] [CrossRef]
- Namasivayam, V.; Vanangamudi, M.; Kramer, V.G.; Kurup, S.; Zhan, P.; Liu, X.; Kongsted, J.; Byrareddy, S.N. The Journey of HIV-1 Non-Nucleoside Reverse Transcriptase Inhibitors (NNRTIs) from Lab to Clinic. J. Med. Chem. 2019, 62, 4851–4883. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Diabetes and Digestive and Kidney Diseases. LiverTox: Clinical and Research Information on Drug-Induced Liver Injury [Internet]; Protease Inhibitors; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2012. [Google Scholar]
- Wang, Y.; Lv, Z.; Chu, Y. HIV Protease Inhibitors: A Review of Molecular Selectivity and Toxicity. HIV/AIDS—Res. Palliat. Care 2015, 7, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Oliveira Passos, D.; Shan, Z.; Smith, S.J.; Sun, Q.; Biswas, A.; Choudhuri, I.; Strutzenberg, T.S.; Haldane, A.; Deng, N.; et al. Mechanisms of HIV-1 Integrase Resistance to Dolutegravir and Potent Inhibition of Drug-Resistant Variants. Sci. Adv. 2023, 9, eadg5953. [Google Scholar] [CrossRef]
- Delelis, O.; Carayon, K.; Saïb, A.; Deprez, E.; Mouscadet, J.-F. Integrase and Integration: Biochemical Activities of HIV-1 Integrase. Retrovirology 2008, 5, 114. [Google Scholar] [CrossRef]
- Hazuda, D.J.; Felock, P.; Witmer, M.; Wolfe, A.; Stillmock, K.; Grobler, J.A.; Espeseth, A.; Gabryelski, L.; Schleif, W.; Blau, C.; et al. Inhibitors of Strand Transfer That Prevent Integration and Inhibit HIV-1 Replication in Cells. Science 2000, 287, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Irlbeck, D.M.; Amrine-Madsen, H.; Kitrinos, K.M.; LaBranche, C.C.; Demarest, J.F. Chemokine (C-C Motif) Receptor 5-Using Envelopes Predominate in Dual/Mixed-Tropic HIV from the Plasma of Drug-Naive Individuals. AIDS 2008, 22, 1425–1431. [Google Scholar] [CrossRef]
- Mosier, D.E. How HIV Changes Its Tropism: Evolution and Adaptation? Curr. Opin. HIV AIDS 2008, 4, 125–130. [Google Scholar] [CrossRef]
- Vandekerckhove, L.; Verhofstede, C.; Vogelaers, D. Maraviroc: Perspectives for Use in Antiretroviral-Naive HIV-1-Infected Patients. J. Antimicrob. Chemother. 2009, 63, 1087–1096. [Google Scholar] [CrossRef]
- Gulick, R.M.; Fatkenheuer, G.; Burnside, R.; Hardy, W.D.; Nelson, M.R.; Goodrich, J.; Mukwaya, G.; Portsmouth, S.; Heera, J.R. Five-Year Safety Evaluation of Maraviroc in HIV-1–Infected Treatment-Experienced Patients. JAIDS J. Acquir. Immune Defic. Syndr. 2014, 65, 78–81. [Google Scholar] [CrossRef]
- Saag, M.; Goodrich, J.; Fätkenheuer, G.; Clotet, B.; Clumeck, N.; Sullivan, J.; Westby, M.; van der Ryst, E.; Mayer, H. A Double-Blind, Placebo-Controlled Trial of Maraviroc in Treatment-Experienced Patients Infected with Non-R5 HIV-1. J. Infect. Dis. 2009, 199, 1638–1647. [Google Scholar] [CrossRef]
- Tan, Q.; Zhu, Y.; Li, J.; Chen, Z.; Han, G.W.; Kufareva, I.; Li, T.; Ma, L.; Fenalti, G.; Li, J.; et al. Structure of the CCR5 Chemokine Receptor–HIV Entry Inhibitor Maraviroc Complex. Science 2013, 341, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Seval, N.; Frank, C.; Kozal, M. Fostemsavir for the Treatment of HIV. Expert Rev. Anti. Infect. Ther. 2021, 19, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Kozal, M.; Aberg, J.; Pialoux, G.; Cahn, P.; Thompson, M.; Molina, J.-M.; Grinsztejn, B.; Diaz, R.; Castagna, A.; Kumar, P.; et al. Fostemsavir in Adults with Multidrug-Resistant HIV-1 Infection. N. Engl. J. Med. 2020, 382, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Meanwell, N.A.; Krystal, M.R.; Nowicka-Sans, B.; Langley, D.R.; Conlon, D.A.; Eastgate, M.D.; Grasela, D.M.; Timmins, P.; Wang, T.; Kadow, J.F. Inhibitors of HIV-1 Attachment: The Discovery and Development of Temsavir and Its Prodrug Fostemsavir. J. Med. Chem. 2018, 61, 62–80. [Google Scholar] [CrossRef] [PubMed]
- Pancera, M.; Lai, Y.-T.; Bylund, T.; Druz, A.; Narpala, S.; O’Dell, S.; Schön, A.; Bailer, R.T.; Chuang, G.-Y.; Geng, H.; et al. Crystal Structures of Trimeric HIV Envelope with Entry Inhibitors BMS-378806 and BMS-626529. Nat. Chem. Biol. 2017, 13, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Lalezari, J.P.; Henry, K.; O’Hearn, M.; Montaner, J.S.G.; Piliero, P.J.; Trottier, B.; Walmsley, S.; Cohen, C.; Kuritzkes, D.R.; Eron, J.J.; et al. Enfuvirtide, an HIV-1 Fusion Inhibitor, for Drug-Resistant HIV Infection in North and South America. N. Engl. J. Med. 2003, 348, 2175–2185. [Google Scholar] [CrossRef] [PubMed]
- Matthews, T.; Salgo, M.; Greenberg, M.; Chung, J.; DeMasi, R.; Bolognesi, D. Enfuvirtide: The First Therapy to Inhibit the Entry of HIV-1 into Host CD4 Lymphocytes. Nat. Rev. Drug Discov. 2004, 3, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Lazzarin, A. Enfuvirtide: The First HIV Fusion Inhibitor. Expert Opin. Pharmacother. 2005, 6, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Jamjian, M.C.; McNicholl, I.R. Enfuvirtide: First Fusion Inhibitor for Treatment of HIV Infection. Am. J. Health-Syst. Pharm. 2004, 61, 1242–1247. [Google Scholar] [CrossRef]
- Follis, K.E.; Larson, S.J.; Lu, M.; Nunberg, J.H. Genetic Evidence That Interhelical Packing Interactions in the Gp41 Core Are Critical for Transition of the Human Immunodeficiency Virus Type 1 Envelope Glycoprotein to the Fusion-Active State. J. Virol. 2002, 76, 7356–7362. [Google Scholar] [CrossRef]
- Beccari, M.V.; Mogle, B.T.; Sidman, E.F.; Mastro, K.A.; Asiago-Reddy, E.; Kufel, W.D. Ibalizumab, a Novel Monoclonal Antibody for the Management of Multidrug-Resistant HIV-1 Infection. Antimicrob. Agents Chemother. 2019, 63, 10–1128. [Google Scholar] [CrossRef]
- Bettiker, R.L.; Koren, D.E.; Jacobson, J.M. Ibalizumab. Curr. Opin. HIV AIDS 2018, 13, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.S.; Fordyce, M.W.; Franco, D.; Kao, C.-Y.; Seaman, M.S.; Ho, D.D. Anti-CD4 Monoclonal Antibody Ibalizumab Exhibits Breadth and Potency Against HIV-1, with Natural Resistance Mediated by the Loss of a V5 Glycan in Envelope. JAIDS J. Acquir. Immune Defic. Syndr. 2013, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Franco, D.; Kao, C.-Y.; Yu, F.; Huang, Y.; Ho, D.D. Epitope Mapping of Ibalizumab, a Humanized Anti-CD4 Monoclonal Antibody with Anti-HIV-1 Activity in Infected Patients. J. Virol. 2010, 84, 6935–6942. [Google Scholar] [CrossRef]
- Freeman, M.M.; Seaman, M.S.; Rits-Volloch, S.; Hong, X.; Kao, C.-Y.; Ho, D.D.; Chen, B. Crystal Structure of HIV-1 Primary Receptor CD4 in Complex with a Potent Antiviral Antibody. Structure 2010, 18, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Iacob, S.A.; Iacob, D.G. Ibalizumab Targeting CD4 Receptors, An Emerging Molecule in HIV Therapy. Front. Microbiol. 2017, 8, 2323. [Google Scholar] [CrossRef] [PubMed]
- Gulick, R.M.; Flexner, C. Long-Acting HIV Drugs for Treatment and Prevention. Annu. Rev. Med. 2019, 70, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Gendelman, H.E.; McMillan, J.; Bade, A.N.; Edagwa, B.; Kevadiya, B.D. The Promise of Long-Acting Antiretroviral Therapies: From Need to Manufacture. Trends Microbiol. 2019, 27, 593–606. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.M.; Hall, E.; Tomlin, R. Injectable Long-Acting Cabotegravir–Rilpivirine Therapy for People Living With HIV/AIDS: Addressing Implementation Barriers From the Start. J. Assoc. Nurses AIDS Care 2023, 34, 216–220. [Google Scholar] [CrossRef]
- Slama, L.; Porcher, R.; Linard, F.; Chakvetadze, C.; Cros, A.; Carillon, S.; Gallardo, L.; Viard, J.-P.; Molina, J.-M. Injectable Long Acting Antiretroviral for HIV Treatment and Prevention: Perspectives of Potential Users. BMC Infect. Dis. 2023, 23, 98. [Google Scholar] [CrossRef]
- Moreno, S.; Rivero, A.; Ventayol, P.; Falcó, V.; Torralba, M.; Schroeder, M.; Neches, V.; Vallejo-Aparicio, L.A.; Mackenzie, I.; Turner, M.; et al. Cabotegravir and Rilpivirine Long-Acting Antiretroviral Therapy Administered Every 2 Months Is Cost-Effective for the Treatment of HIV-1 in Spain. Infect. Dis. Ther. 2023, 12, 2039–2055. [Google Scholar] [CrossRef] [PubMed]
- Bares, S.H.; Scarsi, K.K. A New Paradigm for Antiretroviral Delivery: Long-Acting Cabotegravir and Rilpivirine for the Treatment and Prevention of HIV. Curr. Opin. HIV AIDS 2022, 17, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Thoueille, P.; Choong, E.; Cavassini, M.; Buclin, T.; Decosterd, L.A. Long-Acting Antiretrovirals: A New Era for the Management and Prevention of HIV Infection. J. Antimicrob. Chemother. 2022, 77, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Cobb, D.A.; Smith, N.A.; Edagwa, B.J.; McMillan, J.M. Long-Acting Approaches for Delivery of Antiretroviral Drugs for Prevention and Treatment of HIV: A Review of Recent Research. Expert Opin. Drug Deliv. 2020, 17, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS Global HIV & AIDS Statistics—Fact Sheet. 2023. Available online: https://www.unaids.org/sites/default/files/media_asset/UNAIDS_FactSheet_en.pdf (accessed on 4 April 2024).
- WHO. HIV Drug Resistance Report 2021; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Pennings, P.S. HIV Drug Resistance: Problems and Perspectives. Infect. Dis. Rep. 2013, 5, e5. [Google Scholar] [CrossRef]
- Zdanowicz, M.M. The Pharmacology of HIV Drug Resistance. Am. J. Pharm. Educ. 2006, 70, 100. [Google Scholar] [CrossRef] [PubMed]
- Richman, D.D.; Morton, S.C.; Wrin, T.; Hellmann, N.; Berry, S.; Shapiro, M.F.; Bozzette, S.A. The Prevalence of Antiretroviral Drug Resistance in the United States. AIDS 2004, 18, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Frentz, D.; van de Vijver, D.; Abecasis, A.; Albert, J.; Hamouda, O.; Jørgensen, L.; Kücherer, C.; Struck, D.; Schmit, J.-C.; Vercauteren, J.; et al. Patterns of Transmitted HIV Drug Resistance in Europe Vary by Risk Group. PLoS ONE 2014, 9, e94495. [Google Scholar] [CrossRef]
- Yerly, S.; Vora, S.; Rizzardi, P.; Chave, J.-P.; Vernazza, P.L.; Flepp, M.; Telenti, A.; Battegay, M.; Veuthey, A.-L.; Bru, J.-P.; et al. Acute HIV Infection: Impact on the Spread of HIV and Transmission of Drug Resistance. AIDS 2001, 15, 2287–2292. [Google Scholar] [CrossRef]
- Pao, D.; Fisher, M.; Hué, S.; Dean, G.; Murphy, G.; Cane, P.A.; Sabin, C.A.; Pillay, D. Transmission of HIV-1 during Primary Infection: Relationship to Sexual Risk and Sexually Transmitted Infections. AIDS 2005, 19, 85–90. [Google Scholar] [CrossRef]
- Brenner, B.G.; Roger, M.; Routy, J.; Moisi, D.; Ntemgwa, M.; Matte, C.; Baril, J.; Thomas, R.; Rouleau, D.; Bruneau, J.; et al. High Rates of Forward Transmission Events after Acute/Early HIV-1 Infection. J. Infect. Dis. 2007, 195, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Temereanca, A.; Ruta, S. Strategies to Overcome HIV Drug Resistance-Current and Future Perspectives. Front. Microbiol. 2023, 14, 1133407. [Google Scholar] [CrossRef] [PubMed]
- Kühnert, D.; Kouyos, R.; Shirreff, G.; Pečerska, J.; Scherrer, A.U.; Böni, J.; Yerly, S.; Klimkait, T.; Aubert, V.; Günthard, H.F.; et al. Quantifying the Fitness Cost of HIV-1 Drug Resistance Mutations through Phylodynamics. PLoS Pathog. 2018, 14, e1006895. [Google Scholar] [CrossRef] [PubMed]
- Wittkop, L.; Günthard, H.F.; de Wolf, F.; Dunn, D.; Cozzi-Lepri, A.; de Luca, A.; Kücherer, C.; Obel, N.; von Wyl, V.; Masquelier, B.; et al. Effect of Transmitted Drug Resistance on Virological and Immunological Response to Initial Combination Antiretroviral Therapy for HIV (EuroCoord-CHAIN Joint Project): A European Multicohort Study. Lancet Infect. Dis. 2011, 11, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Jordan, M.R.; Sultan, B.J.; Hill, A.; Davis, D.H.; Gregson, J.; Sawyer, A.W.; Hamers, R.L.; Ndembi, N.; Pillay, D.; et al. Global Trends in Antiretroviral Resistance in Treatment-Naive Individuals with HIV after Rollout of Antiretroviral Treatment in Resource-Limited Settings: A Global Collaborative Study and Meta-Regression Analysis. Lancet 2012, 380, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gregson, J.; Parkin, N.; Haile-Selassie, H.; Tanuri, A.; Andrade Forero, L.; Kaleebu, P.; Watera, C.; Aghokeng, A.; Mutenda, N.; et al. HIV-1 Drug Resistance before Initiation or Re-Initiation of First-Line Antiretroviral Therapy in Low-Income and Middle-Income Countries: A Systematic Review and Meta-Regression Analysis. Lancet Infect. Dis. 2018, 18, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.N.S.; Pingarilho, M.; Pimentel, V.; Martins, M.d.R.O.; Kaiser, R.; Seguin-Devaux, C.; Paredes, R.; Zazzi, M.; Incardona, F.; Abecasis, A.B. Trends of Transmitted and Acquired Drug Resistance in Europe from 1981 to 2019: A Comparison between the Populations of Late Presenters and Non-Late Presenters. Front. Microbiol. 2022, 13, 846943. [Google Scholar] [CrossRef]
- Stadeli, K.M.; Richman, D.D. Rates of Emergence of HIV Drug Resistance in Resource-Limited Settings: A Systematic Review. Antivir. Ther. 2013, 18, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.; Goldstein, F.; Reichmuth, M.L.; Kouyos, R.D.; Wandeler, G.; Egger, M.; Riou, J. Acquired HIV Drug Resistance Mutations on First-Line Antiretroviral Therapy in Southern Africa: Systematic Review and Bayesian Evidence Synthesis. J. Clin. Epidemiol. 2022, 148, 135–145. [Google Scholar] [CrossRef]
- Shirasaka, T.; Kavlick, M.F.; Ueno, T.; Gao, W.Y.; Kojima, E.; Alcaide, M.L.; Chokekijchai, S.; Roy, B.M.; Arnold, E.; Yarchoan, R. Emergence of Human Immunodeficiency Virus Type 1 Variants with Resistance to Multiple Dideoxynucleosides in Patients Receiving Therapy with Dideoxynucleosides. Proc. Natl. Acad. Sci. USA 1995, 92, 2398–2402. [Google Scholar] [CrossRef]
- Marcelin, A.-G. Resistance to Nucleoside Reverse Transcriptase Inhibitors. In Antiretroviral Resistance in Clinical Practice; Mediscript: London, UK, 2006; ISBN 9780955166907. [Google Scholar]
- Winters, M.A.; Coolley, K.L.; Girard, Y.A.; Levee, D.J.; Hamdan, H.; Shafer, R.W.; Katzenstein, D.A.; Merigan, T.C. A 6-Basepair Insert in the Reverse Transcriptase Gene of Human Immunodeficiency Virus Type 1 Confers Resistance to Multiple Nucleoside Inhibitors. J. Clin. Investig. 1998, 102, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- de Jong, J.J.; Goudsmit, J.; Lukashov, V.V.; Hillebrand, M.E.; Baan, E.; Huismans, R.; Danner, S.A.; ten Veen, J.H.; de Wolf, F.; Jurriaans, S. Insertion of Two Animo Acids Combined with Changes in Reverse Transcriptase Containing Tyrosine-215 of HIV-1 Resistant to Multiple Nucleoside Analogs. AIDS 1999, 13, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Richman, D.D. Antiretroviral Drug Resistance: Mechanisms, Pathogenesis, Clinical Significance. In Antiviral Chemotherapy 4. Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1996; pp. 383–395. [Google Scholar]
- Shafer, R.W.; Schapiro, J.M. HIV-1 Drug Resistance Mutations: An Updated Framework for the Second Decade of HAART. AIDS Rev. 2008, 10, 67–84. [Google Scholar] [PubMed]
- Brenner, B.; Turner, D.; Oliveira, M.; Moisi, D.; Detorio, M.; Carobene, M.; Marlink, R.G.; Schapiro, J.; Roger, M.; Wainberg, M.A. A V106M Mutation in HIV-1 Clade C Viruses Exposed to Efavirenz Confers Cross-Resistance to Non-Nucleoside Reverse Transcriptase Inhibitors. AIDS 2003, 17, F1–F5. [Google Scholar] [CrossRef] [PubMed]
- Sarafianos, S.G.; Das, K.; Hughes, S.H.; Arnold, E. Taking Aim at a Moving Target: Designing Drugs to Inhibit Drug-Resistant HIV-1 Reverse Transcriptases. Curr. Opin. Struct. Biol. 2004, 14, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Stammers, D.K. Structural Basis for Drug Resistance Mechanisms for Non-Nucleoside Inhibitors of HIV Reverse Transcriptase. Virus Res. 2008, 134, 157–170. [Google Scholar] [CrossRef]
- Lecossier, D.; Shulman, N.S.; Morand-Joubert, L.; Shafer, R.W.; Joly, V.; Zolopa, A.R.; Clavel, F.; Hance, A.J. Detection of Minority Populations of HIV-1 Expressing the K103N Resistance Mutation in Patients Failing Nevirapine. JAIDS J. Acquir. Immune Defic. Syndr. 2005, 38, 37–42. [Google Scholar] [CrossRef]
- Rhee, S.-Y.; Taylor, J.; Wadhera, G.; Ben-Hur, A.; Brutlag, D.L.; Shafer, R.W. Genotypic Predictors of Human Immunodeficiency Virus Type 1 Drug Resistance. Proc. Natl. Acad. Sci. USA 2006, 103, 17355–17360. [Google Scholar] [CrossRef]
- Bacheler, L.; Jeffrey, S.; Hanna, G.; D’Aquila, R.; Wallace, L.; Logue, K.; Cordova, B.; Hertogs, K.; Larder, B.; Buckery, R.; et al. Genotypic Correlates of Phenotypic Resistance to Efavirenz in Virus Isolates from Patients Failing Nonnucleoside Reverse Transcriptase Inhibitor Therapy. J. Virol. 2001, 75, 4999–5008. [Google Scholar] [CrossRef]
- Vingerhoets, J.; Azijn, H.; Fransen, E.; De Baere, I.; Smeulders, L.; Jochmans, D.; Andries, K.; Pauwels, R.; de Béthune, M.-P. TMC125 Displays a High Genetic Barrier to the Development of Resistance: Evidence from In Vitro Selection Experiments. J. Virol. 2005, 79, 12773–12782. [Google Scholar] [CrossRef]
- Parkin, N.T.; Gupta, S.; Chappey, C.; Petropoulos, C.J. The K101P and K103R/V179D Mutations in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Confer Resistance to Nonnucleoside Reverse Transcriptase Inhibitors. Antimicrob. Agents Chemother. 2006, 50, 351–354. [Google Scholar] [CrossRef]
- Rhee, S.-Y. Human Immunodeficiency Virus Reverse Transcriptase and Protease Sequence Database. Nucleic Acids Res. 2003, 31, 298–303. [Google Scholar] [CrossRef]
- Brown, A.J.L.; Precious, H.M.; Whitcomb, J.M.; Wong, J.K.; Quigg, M.; Huang, W.; Daar, E.S.; D’Aquila, R.T.; Keiser, P.H.; Connick, E.; et al. Reduced Susceptibility of Human Immunodeficiency Virus Type 1 (HIV-1) from Patients with Primary HIV Infection to Nonnucleoside Reverse Transcriptase Inhibitors Is Associated with Variation at Novel Amino Acid Sites. J. Virol. 2000, 74, 10269–10273. [Google Scholar] [CrossRef]
- Ceccherini-Silberstein, F.; Svicher, V.; Sing, T.; Artese, A.; Santoro, M.M.; Forbici, F.; Bertoli, A.; Alcaro, S.; Palamara, G.; d’Arminio Monforte, A.; et al. Characterization and Structural Analysis of Novel Mutations in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Involved in the Regulation of Resistance to Nonnucleoside Inhibitors. J. Virol. 2007, 81, 11507–11519. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Chapsal, B.D.; Weber, I.T.; Mitsuya, H. Design of HIV Protease Inhibitors Targeting Protein Backbone: An Effective Strategy for Combating Drug Resistance. Acc. Chem. Res. 2008, 41, 78–86. [Google Scholar] [CrossRef]
- Wensing, A.M.J.; van Maarseveen, N.M.; Nijhuis, M. Fifteen Years of HIV Protease Inhibitors: Raising the Barrier to Resistance. Antivir. Res. 2010, 85, 59–74. [Google Scholar] [CrossRef]
- Johnson, V.A.; Brun-Vezinet, F.; Clotet, B.; Gunthard, H.F.; Kuritzkes, D.R.; Pillay, D.; Schapiro, J.M.; Richman, D.D. Update of the Drug Resistance Mutations in HIV-1. Top. HIV Med. 2008, 16, 138–145. [Google Scholar]
- Cooper, D.A.; Steigbigel, R.T.; Gatell, J.M.; Rockstroh, J.K.; Katlama, C.; Yeni, P.; Lazzarin, A.; Clotet, B.; Kumar, P.N.; Eron, J.E.; et al. Subgroup and Resistance Analyses of Raltegravir for Resistant HIV-1 Infection. N. Engl. J. Med. 2008, 359, 355–365. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nakahara, K.; Seki, T.; Miki, S.; Kawauchi, S.; Suyama, A.; Wakasamorimoto, C.; Kodama, M.; Endoh, T.; Oosugi, E. Selection of Diverse and Clinically Relevant Integrase Inhibitor-Resistant Human Immunodeficiency Virus Type 1 Mutants. Antivir. Res. 2008, 80, 213–222. [Google Scholar] [CrossRef]
- Steigbigel, R.T.; Cooper, D.A.; Teppler, H.; Eron, J.J.; Gatell, J.M.; Kumar, P.N.; Rockstroh, J.K.; Schechter, M.; Katlama, C.; Markowitz, M.; et al. Long-Term Efficacy and Safety of Raltegravir Combined with Optimized Background Therapy in Treatment-Experienced Patients with Drug-Resistant HIV Infection: Week 96 Results of the BENCHMRK 1 and 2 Phase III Trials. Clin. Infect. Dis. 2010, 50, 605–612. [Google Scholar] [CrossRef]
- Van Wesenbeeck, L.; Rondelez, E.; Feyaerts, M.; Verheyen, A.; Van der Borght, K.; Smits, V.; Cleybergh, C.; De Wolf, H.; Van Baelen, K.; Stuyver, L.J. Cross-Resistance Profile Determination of Two Second-Generation HIV-1 Integrase Inhibitors Using a Panel of Recombinant Viruses Derived from Raltegravir-Treated Clinical Isolates. Antimicrob. Agents Chemother. 2011, 55, 321–325. [Google Scholar] [CrossRef]
- Bar-Magen, T.; Sloan, R.D.; Faltenbacher, V.H.; Donahue, D.A.; Kuhl, B.D.; Oliveira, M.; Xu, H.; Wainberg, M.A. Comparative Biochemical Analysis of HIV-1 Subtype B and C Integrase Enzymes. Retrovirology 2009, 6, 103. [Google Scholar] [CrossRef]
- Bar-Magen, T.; Sloan, R.D.; Donahue, D.A.; Kuhl, B.D.; Zabeida, A.; Xu, H.; Oliveira, M.; Hazuda, D.J.; Wainberg, M.A. Identification of Novel Mutations Responsible for Resistance to MK-2048, a Second-Generation HIV-1 Integrase Inhibitor. J. Virol. 2010, 84, 9210–9216. [Google Scholar] [CrossRef]
- Li, X.; Krishnan, L.; Cherepanov, P.; Engelman, A. Structural Biology of Retroviral DNA Integration. Virology 2011, 411, 194–205. [Google Scholar] [CrossRef]
- Bacchi, A.; Carcelli, M.; Compari, C.; Fisicaro, E.; Pala, N.; Rispoli, G.; Rogolino, D.; Sanchez, T.W.; Sechi, M.; Sinisi, V.; et al. Investigating the Role of Metal Chelation in HIV-1 Integrase Strand Transfer Inhibitors. J. Med. Chem. 2011, 54, 8407–8420. [Google Scholar] [CrossRef]
- Hare, S.; Smith, S.J.; Métifiot, M.; Jaxa-Chamiec, A.; Pommier, Y.; Hughes, S.H.; Cherepanov, P. Structural and Functional Analyses of the Second-Generation Integrase Strand Transfer Inhibitor Dolutegravir (S/GSK1349572). Mol. Pharmacol. 2011, 80, 565–572. [Google Scholar] [CrossRef]
- Hare, S.; Vos, A.M.; Clayton, R.F.; Thuring, J.W.; Cummings, M.D.; Cherepanov, P. Molecular Mechanisms of Retroviral Integrase Inhibition and the Evolution of Viral Resistance. Proc. Natl. Acad. Sci. USA 2010, 107, 20057–20062. [Google Scholar] [CrossRef]
- Hare, S.; Gupta, S.S.; Valkov, E.; Engelman, A.; Cherepanov, P. Retroviral Intasome Assembly and Inhibition of DNA Strand Transfer. Nature 2010, 464, 232–236. [Google Scholar] [CrossRef]
- Espeseth, A.S.; Felock, P.; Wolfe, A.; Witmer, M.; Grobler, J.; Anthony, N.; Egbertson, M.; Melamed, J.Y.; Young, S.; Hamill, T.; et al. HIV-1 Integrase Inhibitors That Compete with the Target DNA Substrate Define a Unique Strand Transfer Conformation for Integrase. Proc. Natl. Acad. Sci. USA 2000, 97, 11244–11249. [Google Scholar] [CrossRef]
- Mesplède, T.; Quashie, P.K.; Wainberg, M.A. Resistance to HIV Integrase Inhibitors. Curr. Opin. HIV AIDS 2012, 7, 401–408. [Google Scholar] [CrossRef]
- Mbhele, N.; Chimukangara, B.; Gordon, M. HIV-1 Integrase Strand Transfer Inhibitors: A Review of Current Drugs, Recent Advances and Drug Resistance. Int. J. Antimicrob. Agents 2021, 57, 106343. [Google Scholar] [CrossRef]
- Temesgen, Z. Raltegravir: First in Class HIV Integrase Inhibitor. Ther. Clin. Risk Manag. 2008, 4, 493–500. [Google Scholar] [CrossRef]
- Wohl, D.A.; Cohen, C.; Gallant, J.E.; Mills, A.; Sax, P.E.; DeJesus, E.; Zolopa, A.; Liu, H.C.; Plummer, A.; White, K.L.; et al. A Randomized, Double-Blind Comparison of Single-Tablet Regimen Elvitegravir/Cobicistat/Emtricitabine/Tenofovir DF Versus Single-Tablet Regimen Efavirenz/Emtricitabine/Tenofovir DF for Initial Treatment of HIV-1 Infection. JAIDS J. Acquir. Immune Defic. Syndr. 2014, 65, e118–e120. [Google Scholar] [CrossRef]
- Clumeck, N.; Molina, J.-M.; Henry, K.; Gathe, J.; Rockstroh, J.K.; DeJesus, E.; Wei, X.; White, K.; Fordyce, M.W.; Rhee, M.S.; et al. A Randomized, Double-Blind Comparison of Single-Tablet Regimen Elvitegravir/Cobicistat/Emtricitabine/Tenofovir DF vs Ritonavir-Boosted Atazanavir Plus Emtricitabine/Tenofovir DF for Initial Treatment of HIV-1 Infection. JAIDS J. Acquir. Immune Defic. Syndr. 2014, 65, e121–e124. [Google Scholar] [CrossRef]
- Lübke, N.; Jensen, B.; Hüttig, F.; Feldt, T.; Walker, A.; Thielen, A.; Däumer, M.; Obermeier, M.; Kaiser, R.; Knops, E.; et al. Failure of Dolutegravir First-Line ART with Selection of Virus Carrying R263K and G118R. N. Engl. J. Med. 2019, 381, 887–889. [Google Scholar] [CrossRef]
- Wensing, A.M.; Calvez, V.; Ceccherini-Silberstein, F.; Charpentier, C.; Günthard, H.F.; Paredes, R.; Shafer, R.W.; Richman, D.D. 2019 Update of the Drug Resistance Mutations in HIV-1. Top. Antivir. Med. 2019, 27, 111–121. [Google Scholar]
- Labrosse, B.; Labernardière, J.-L.; Dam, E.; Trouplin, V.; Skrabal, K.; Clavel, F.; Mammano, F. Baseline Susceptibility of Primary Human Immunodeficiency Virus Type 1 to Entry Inhibitors. J. Virol. 2003, 77, 1610–1613. [Google Scholar] [CrossRef]
- Reeves, J.D.; Gallo, S.A.; Ahmad, N.; Miamidian, J.L.; Harvey, P.E.; Sharron, M.; Pöhlmann, S.; Sfakianos, J.N.; Derdeyn, C.A.; Blumenthal, R.; et al. Sensitivity of HIV-1 to Entry Inhibitors Correlates with Envelope/Coreceptor Affinity, Receptor Density, and Fusion Kinetics. Proc. Natl. Acad. Sci. USA 2002, 99, 16249–16254. [Google Scholar] [CrossRef]
- Reeves, J.D.; Miamidian, J.L.; Biscone, M.J.; Lee, F.-H.; Ahmad, N.; Pierson, T.C.; Doms, R.W. Impact of Mutations in the Coreceptor Binding Site on Human Immunodeficiency Virus Type 1 Fusion, Infection, and Entry Inhibitor Sensitivity. J. Virol. 2004, 78, 5476–5485. [Google Scholar] [CrossRef]
- Ray, N. Maraviroc in the Treatment of HIV Infection. Drug Des. Dev. Ther. 2008, 2, 151–161. [Google Scholar] [CrossRef]
- Poveda, E.; Rodés, B.; Toro, C.; Martín-Carbonero, L.; Gonzalez-Lahoz, J.; Soriano, V. Evolution of the Gp41 Env Region in HIV-Infected Patients Receiving T-20, a Fusion Inhibitor. AIDS 2002, 16, 1959–1961. [Google Scholar] [CrossRef] [PubMed]
- Rimsky, L.T.; Shugars, D.C.; Matthews, T.J. Determinants of Human Immunodeficiency Virus Type 1 Resistance to Gp41-Derived Inhibitory Peptides. J. Virol. 1998, 72, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Decker, J.M.; Liu, H.; Zhang, Z.; Arani, R.B.; Kilby, J.M.; Saag, M.S.; Wu, X.; Shaw, G.M.; Kappes, J.C. Emergence of Resistant Human Immunodeficiency Virus Type 1 in Patients Receiving Fusion Inhibitor (T-20) Monotherapy. Antimicrob. Agents Chemother. 2002, 46, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Mink, M.; Mosier, S.M.; Janumpalli, S.; Davison, D.; Jin, L.; Melby, T.; Sista, P.; Erickson, J.; Lambert, D.; Stanfield-Oakley, S.A.; et al. Impact of Human Immunodeficiency Virus Type 1 Gp41 Amino Acid Substitutions Selected during Enfuvirtide Treatment on Gp41 Binding and Antiviral Potency of Enfuvirtide In Vitro. J. Virol. 2005, 79, 12447–12454. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Deeks, S.G.; Hoh, R.; Beatty, G.; Kuritzkes, B.A.; Martin, J.N.; Kuritzkes, D.R. Rapid Emergence of Enfuvirtide Resistance in HIV-1-Infected Patients. JAIDS J. Acquir. Immune Defic. Syndr. 2006, 43, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Pozniak, A.; Wildfire, A.; Stanfield-Oakley, S.A.; Mosier, S.M.; Ratcliffe, D.; Workman, J.; Joall, A.; Myers, R.; Smit, E.; et al. Emergence and Evolution of Enfuvirtide Resistance Following Long-Term Therapy Involves Heptad Repeat 2 Mutations within Gp41. Antimicrob. Agents Chemother. 2005, 49, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.L. Resistance to Enfuvirtide, the First HIV Fusion Inhibitor. J. Antimicrob. Chemother. 2004, 54, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.H.M.; Chen, J.H.K.; Wong, K.H.; Chan, K.C.; Lam, H.Y.; Cheng, V.C.C.; Yuen, K.Y.; Yam, W.C. High Prevalence of Primary Enfuvirtide (ENF) Resistance-Associated Mutations in HIV-1-Infected Patients in Hong Kong. J. Clin. Virol. 2010, 47, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.A.; Robinson, L.; Tseng, A.; MacArthur, R.D. New Antiretroviral Drugs: A Review of the Efficacy, Safety, Pharmacokinetics, and Resistance Profile of Tipranavir, Darunavir, Etravirine, Rilpivirine, Maraviroc, and Raltegravir. Expert Opin. Pharmacother. 2009, 10, 2445–2466. [Google Scholar] [CrossRef]
- Pugach, P.; Marozsan, A.J.; Ketas, T.J.; Landes, E.L.; Moore, J.P.; Kuhmann, S.E. HIV-1 Clones Resistant to a Small Molecule CCR5 Inhibitor Use the Inhibitor-Bound Form of CCR5 for Entry. Virology 2007, 361, 212–228. [Google Scholar] [CrossRef]
- Roche, M.; Salimi, H.; Duncan, R.; Wilkinson, B.L.; Chikere, K.; Moore, M.S.; Webb, N.E.; Zappi, H.; Sterjovski, J.; Flynn, J.K.; et al. A Common Mechanism of Clinical HIV-1 Resistance to the CCR5 Antagonist Maraviroc despite Divergent Resistance Levels and Lack of Common Gp120 Resistance Mutations. Retrovirology 2013, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Trkola, A.; Kuhmann, S.E.; Strizki, J.M.; Maxwell, E.; Ketas, T.; Morgan, T.; Pugach, P.; Xu, S.; Wojcik, L.; Tagat, J.; et al. HIV-1 Escape from a Small Molecule, CCR5-Specific Entry Inhibitor Does Not Involve CXCR4 Use. Proc. Natl. Acad. Sci. USA 2002, 99, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Hiryak, K.; Koren, D.E. Fostemsavir: A Novel Attachment Inhibitor for Patients With Multidrug-Resistant HIV-1 Infection. Ann. Pharmacother. 2021, 55, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Berruti, M.; Pincino, R.; Taramasso, L.; Di Biagio, A. Evaluating Fostemsavir as a Therapeutic Option for Patients with HIV. Expert Opin. Pharmacother. 2021, 22, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Nowicka-Sans, B.; McAuliffe, B.; Ray, N.; Eggers, B.; Fang, H.; Fan, L.; Healy, M.; Langley, D.R.; Hwang, C.; et al. Genotypic Correlates of Susceptibility to HIV-1 Attachment Inhibitor BMS-626529, the Active Agent of the Prodrug BMS-663068. J. Antimicrob. Chemother. 2014, 69, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Lataillade, M.; Zhou, N.; Joshi, S.R.; Lee, S.; Stock, D.A.; Hanna, G.J.; Krystal, M. Viral Drug Resistance Through 48 Weeks, in a Phase 2b, Randomized, Controlled Trial of the HIV-1 Attachment Inhibitor Prodrug, Fostemsavir. JAIDS J. Acquir. Immune Defic. Syndr. 2018, 77, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Lepore, L.; Fabrizio, C.; Bavaro, D.F.; Milano, E.; Volpe, A.; Lagioia, A.; Angarano, G.; Saracino, A.; Monno, L. Gp120 Substitutions at Positions Associated with Resistance to Fostemsavir in Treatment-Naive HIV-1-Positive Individuals. J. Antimicrob. Chemother. 2020, 75, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Gradt, E.; Charpentier, C.; Leoz, M.; Mourez, T.; Descamps, D.; Plantier, J.-C. Impact of Natural Polymorphisms of HIV-1 Non-Group M on Genotypic Susceptibility to the Attachment Inhibitor Fostemsavir. J. Antimicrob. Chemother. 2018, 73, 2716–2720. [Google Scholar] [CrossRef] [PubMed]
- Madani, N.; Perdigoto, A.L.; Srinivasan, K.; Cox, J.M.; Chruma, J.J.; LaLonde, J.; Head, M.; Smith, A.B.; Sodroski, J.G. Localized Changes in the Gp120 Envelope Glycoprotein Confer Resistance to Human Immunodeficiency Virus Entry Inhibitors BMS-806 and #155. J. Virol. 2004, 78, 3742–3752. [Google Scholar] [CrossRef]
- Nowicka-Sans, B.; Gong, Y.-F.; McAuliffe, B.; Dicker, I.; Ho, H.-T.; Zhou, N.; Eggers, B.; Lin, P.-F.; Ray, N.; Wind-Rotolo, M.; et al. In Vitro Antiviral Characteristics of HIV-1 Attachment Inhibitor BMS-626529, the Active Component of the Prodrug BMS-663068. Antimicrob. Agents Chemother. 2012, 56, 3498–3507. [Google Scholar] [CrossRef]
- Ray, N.; Hwang, C.; Healy, M.D.; Whitcomb, J.; Lataillade, M.; Wind-Rotolo, M.; Krystal, M.; Hanna, G.J. Prediction of Virological Response and Assessment of Resistance Emergence to the HIV-1 Attachment Inhibitor BMS-626529 During 8-Day Monotherapy With Its Prodrug BMS-663068. JAIDS J. Acquir. Immune Defic. Syndr. 2013, 64, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Soulie, C.; Lambert-Niclot, S.; Fofana, D.B.; Fourati, S.; Ait-Arkoub, Z.; Sayon, S.; Simon, A.; Katlama, C.; Calvez, V.; Marcelin, A.-G. Frequency of Amino Acid Changes Associated with Resistance to Attachment Inhibitor BMS-626529 in R5- and X4-Tropic HIV-1 Subtype B. J. Antimicrob. Chemother. 2013, 68, 1243–1245. [Google Scholar] [CrossRef] [PubMed]
- Fofana, D.B.; Charpentier, C.; Maiga, A.I.; Lambert-Niclot, S.; Sayon, S.; Desire, N.; Simon, A.; Yazdanpanah, Y.; Katlama, C.; Descamps, D.; et al. Genetic Barrier for Attachment Inhibitor BMS-626529 Resistance in HIV-1 B and Non-B Subtypes. J. Antimicrob. Chemother. 2015, 70, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Ballana, E.; Esté, J.A. BMS-663068, a Safe and Effective HIV-1 Attachment Inhibitor. Lancet HIV 2015, 2, e404–e405. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, C.; Larrouy, L.; Visseaux, B.; Landman, R.; Levittas, M.; Storto, A.; Damond, F.; Yazdanpanah, Y.; Yeni, P.; Brun-Vezinet, F.; et al. Prevalence of Subtype-Related Polymorphisms Associated with in Vitro Resistance to Attachment Inhibitor BMS-626529 in HIV-1 ‘Non-B’-Infected Patients. J. Antimicrob. Chemother. 2012, 67, 1459–1461. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, N.; Sun, Y.; Ray, N.; Lataillade, M.; Hanna, G.J.; Krystal, M. Activity of the HIV-1 Attachment Inhibitor BMS-626529, the Active Component of the Prodrug BMS-663068, against CD4-Independent Viruses and HIV-1 Envelopes Resistant to Other Entry Inhibitors. Antimicrob. Agents Chemother. 2013, 57, 4172–4180. [Google Scholar] [CrossRef]
- Rose, R.; Gartland, M.; Li, Z.; Zhou, N.; Cockett, M.; Beloor, J.; Lataillade, M.; Ackerman, P.; Krystal, M. Clinical Evidence for a Lack of Cross-Resistance between Temsavir and Ibalizumab or Maraviroc. AIDS 2022, 36, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Bouba, Y.; Berno, G.; Fabeni, L.; Carioti, L.; Salpini, R.; Aquaro, S.; Svicher, V.; Perno, C.F.; Ceccherini-Silberstein, F.; Santoro, M.M. Identification of Gp120 Polymorphisms in HIV-1 B Subtype Potentially Associated with Resistance to Fostemsavir. J. Antimicrob. Chemother. 2020, 75, 1778–1786. [Google Scholar] [CrossRef]
- Toma, J.; Weinheimer, S.P.; Stawiski, E.; Whitcomb, J.M.; Lewis, S.T.; Petropoulos, C.J.; Huang, W. Loss of Asparagine-Linked Glycosylation Sites in Variable Region 5 of Human Immunodeficiency Virus Type 1 Envelope Is Associated with Resistance to CD4 Antibody Ibalizumab. J. Virol. 2011, 85, 3872–3880. [Google Scholar] [CrossRef]
- Fessel, W.J.; Anderson, B.; Follansbee, S.E.; Winters, M.A.; Lewis, S.T.; Weinheimer, S.P.; Petropoulos, C.J.; Shafer, R.W. The Efficacy of an Anti-CD4 Monoclonal Antibody for HIV-1 Treatment. Antivir. Res. 2011, 92, 484–487. [Google Scholar] [CrossRef]
- De Feo, C.; Weiss, C. Escape from Human Immunodeficiency Virus Type 1 (HIV-1) Entry Inhibitors. Viruses 2012, 4, 3859–3911. [Google Scholar] [CrossRef] [PubMed]
- Steegen, K.; Chandiwana, N.; Sokhela, S.; Venter, W.D.F.; Hans, L. Impact of Rilpivirine Cross-Resistance on Long-Acting Cabotegravir-Rilpivirine in Low and Middle-Income Countries. AIDS 2023, 37, 1009–1011. [Google Scholar] [CrossRef] [PubMed]
- van Welzen, B.J.; Van Lelyveld, S.F.L.; Ter Beest, G.; Gisolf, J.H.; Geerlings, S.E.; Prins, J.M.; Van Twillert, G.; Van Nieuwkoop, C.; Van der Valk, M.; Burger, D.; et al. Virological Failure after Switch to Long-Acting Cabotegravir and Rilpivirine Injectable Therapy: An In-Depth Analysis. Clin. Infect. Dis. 2024, ciae016. [Google Scholar] [CrossRef] [PubMed]
- Cervo, A.; Russo, A.; Di Carlo, D.; De Vito, A.; Fabeni, L.; D’Anna, S.; Duca, L.; Colpani, A.; Fois, M.; Zauli, B.; et al. Long-Acting Combination of Cabotegravir plus Rilpivirine: A Picture of Potential Eligible and Ineligible HIV-Positive Individuals from the Italian ARCA Cohort. J. Glob. Antimicrob. Resist. 2023, 34, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Iyidogan, P.; Anderson, K. Current Perspectives on HIV-1 Antiretroviral Drug Resistance. Viruses 2014, 6, 4095–4139. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Patil, S.; Guo, X.; Wen, F.; Zhang, X.; Zhong, Z.; Wang, X. Clinical, Epidemiological, and Drug Resistance Insights into HIV-Positive Patients in Meizhou, China. Front. Cell. Infect. Microbiol. 2024, 13, 1330826. [Google Scholar] [CrossRef] [PubMed]
- Su, C.T.-T.; Ling, W.-L.; Lua, W.-H.; Haw, Y.-X.; Gan, S.K.-E. Structural Analyses of 2015-Updated Drug-Resistant Mutations in HIV-1 Protease: An Implication of Protease Inhibitor Cross-Resistance. BMC Bioinform. 2016, 17, 500. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.-Y.; Taylor, J.; Fessel, W.J.; Kaufman, D.; Towner, W.; Troia, P.; Ruane, P.; Hellinger, J.; Shirvani, V.; Zolopa, A.; et al. HIV-1 Protease Mutations and Protease Inhibitor Cross-Resistance. Antimicrob. Agents Chemother. 2010, 54, 4253–4261. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.M.; Kuritzkes, D.R.; Godofsky, E.; DeJesus, E.; Larson, J.A.; Weinheimer, S.P.; Lewis, S.T. Safety, Pharmacokinetics, and Antiretroviral Activity of Multiple Doses of Ibalizumab (Formerly TNX-355), an Anti-CD4 Monoclonal Antibody, in Human Immunodeficiency Virus Type 1-Infected Adults. Antimicrob. Agents Chemother. 2009, 53, 450–457. [Google Scholar] [CrossRef]
- Blair, H.A. Ibalizumab: A Review in Multidrug-Resistant HIV-1 Infection. Drugs 2020, 80, 189–196. [Google Scholar] [CrossRef]
- Grover, D.; Copas, A.; Green, H.; Edwards, S.G.; Dunn, D.T.; Sabin, C.; Phillips, A.; Allen, E.; Pillay, D. What Is the Risk of Mortality Following Diagnosis of Multidrug-Resistant HIV-1? J. Antimicrob. Chemother. 2008, 61, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, L.; Blaschke, T. Adherence to Medication. N. Engl. J. Med. 2005, 353, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhu, P.; Zhang, Y.; Li, J.; Ma, X.; Li, N.; Wang, Q.; Xue, X.; Luo, L.; Li, Z.; et al. Analysis of Social and Genetic Factors Influencing Heterosexual Transmission of HIV within Serodiscordant Couples in the Henan Cohort. PLoS ONE 2015, 10, e0129979. [Google Scholar] [CrossRef] [PubMed]
- Brawner, B.M.; Kerr, J.; Castle, B.F.; Bannon, J.A.; Bonett, S.; Stevens, R.; James, R.; Bowleg, L. A Systematic Review of Neighborhood-Level Influences on HIV Vulnerability. AIDS Behav. 2022, 26, 874–934. [Google Scholar] [CrossRef] [PubMed]
- Nachega, J.B.; Marconi, V.C.; van Zyl, G.U.; Gardner, E.M.; Preiser, W.; Hong, S.Y.; Mills, E.J.; Gross, R. HIV Treatment Adherence, Drug Resistance, Virologic Failure: Evolving Concepts. Infect. Disord.—Drug Targets 2011, 11, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Maggiolo, F.; Ravasio, L.; Ripamonti, D.; Gregis, G.; Quinzan, G.; Arici, C.; Airoldi, M.; Suter, F. Similar Adherence Rates Favor Different Virologic Outcomes for Patients Treated with Nonnucleoside Analogues or Protease Inhibitors. Clin. Infect. Dis. 2005, 40, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Del Cacho, E.; Codina, C.; Tuset, M.; De Lazzari, E.; Mallolas, J.; Miró, J.-M.; Gatell, J.M.; Ribas, J. Relationship between Adherence Level, Type of the Antiretroviral Regimen, and Plasma HIV Type 1 RNA Viral Load: A Prospective Cohort Study. AIDS Res. Hum. Retroviruses 2008, 24, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.K.; Celentano, D.D.; Gange, S.J.; Moore, R.D.; Gallant, J.E. Association between Adherence to Antiretroviral Therapy and Human Immunodeficiency Virus Drug Resistance. Clin. Infect. Dis. 2003, 37, 1112–1118. [Google Scholar] [CrossRef]
- Weidle, P.J.; Nesheim, S. HIV Drug Resistance and Mother-to-Child Transmission of HIV. Clin. Perinatol. 2010, 37, 825–842. [Google Scholar] [CrossRef]
- Mirochnick, M.; Thomas, T.; Capparelli, E.; Zeh, C.; Holland, D.; Masaba, R.; Odhiambo, P.; Fowler, M.G.; Weidle, P.J.; Thigpen, M.C. Antiretroviral Concentrations in Breast-Feeding Infants of Mothers Receiving Highly Active Antiretroviral Therapy. Antimicrob. Agents Chemother. 2009, 53, 1170–1176. [Google Scholar] [CrossRef]
- Arrivé, E.; Newell, M.-L.; Ekouevi, D.K.; Chaix, M.-L.; Thiebaut, R.; Masquelier, B.; Leroy, V.; Van de Perre, P.; Rouzioux, C.; Dabis, F. Prevalence of Resistance to Nevirapine in Mothers and Children after Single-Dose Exposure to Prevent Vertical Transmission of HIV-1: A Meta-Analysis. Int. J. Epidemiol. 2007, 36, 1009–1021. [Google Scholar] [CrossRef]
- Pennings, P.S. Standing Genetic Variation and the Evolution of Drug Resistance in HIV. PLoS Comput. Biol. 2012, 8, e1002527. [Google Scholar] [CrossRef] [PubMed]
- Hunt, G.M.; Ledwaba, J.; Salimo, A.; Kalimashe, M.; Dinh, T.-H.; Jackson, D.; Sherman, G.; Puren, A.; Ngandu, N.K.; Lombard, C.; et al. Prevalence of HIV-1 Drug Resistance amongst Newly Diagnosed HIV-Infected Infants Age 4–8 Weeks, Enrolled in Three Nationally Representative PMTCT Effectiveness Surveys, South Africa: 2010, 2011–12 and 2012–13. BMC Infect. Dis. 2019, 19, 787. [Google Scholar] [CrossRef] [PubMed]
- Boyce, C.L.; Sils, T.; Ko, D.; Wong-on-Wing, A.; Beck, I.A.; Styrchak, S.M.; DeMarrais, P.; Tierney, C.; Stranix-Chibanda, L.; Flynn, P.M.; et al. Maternal Human Immunodeficiency Virus (HIV) Drug Resistance Is Associated with Vertical Transmission and Is Prevalent in Infected Infants. Clin. Infect. Dis. 2022, 74, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Castilla, J.; del Romero, J.; Hernando, V.; Marincovich, B.; García, S.; Rodríguez, C. Effectiveness of Highly Active Antiretroviral Therapy in Reducing Heterosexual Transmission of HIV. JAIDS J. Acquir. Immune Defic. Syndr. 2005, 40, 96–101. [Google Scholar] [CrossRef]
- Tetteh, R.A.; Yankey, B.A.; Nartey, E.T.; Lartey, M.; Leufkens, H.G.M.; Dodoo, A.N.O. Pre-Exposure Prophylaxis for HIV Prevention: Safety Concerns. Drug Saf. 2017, 40, 273–283. [Google Scholar] [CrossRef]
- Kirby, T.; Thornber-Dunwell, M. Uptake of PrEP for HIV Slow among MSM. Lancet 2014, 383, 399–400. [Google Scholar] [CrossRef]
- Molina, J.-M.; Capitant, C.; Spire, B.; Pialoux, G.; Cotte, L.; Charreau, I.; Tremblay, C.; Le Gall, J.-M.; Cua, E.; Pasquet, A.; et al. On-Demand Preexposure Prophylaxis in Men at High Risk for HIV-1 Infection. N. Engl. J. Med. 2015, 373, 2237–2246. [Google Scholar] [CrossRef]
- Mortlock, R.; Smith, V.; Nesci, I.; Bertoldi, A.; Ho, A.; El Mekkawi, Z.; Kakuzada, L.; Williams, K.; Pont, L.; De Rubis, G.; et al. A Comparative Evaluation of Propranolol Pharmacokinetics in Obese versus Ideal Weight Individuals: A Blueprint towards a Personalised Medicine. Chem. Biol. Interact. 2023, 371, 110351. [Google Scholar] [CrossRef]
- Gibas, K.M.; van den Berg, P.; Powell, V.E.; Krakower, D.S. Drug Resistance During HIV Pre-Exposure Prophylaxis. Drugs 2019, 79, 609–619. [Google Scholar] [CrossRef]
- Günthard, H.F.; Saag, M.S.; Benson, C.A.; del Rio, C.; Eron, J.J.; Gallant, J.E.; Hoy, J.F.; Mugavero, M.J.; Sax, P.E.; Thompson, M.A.; et al. Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults. JAMA 2016, 316, 191. [Google Scholar] [CrossRef] [PubMed]
- Christopoulos, K.A.; Grochowski, J.; Mayorga-Munoz, F.; Hickey, M.D.; Imbert, E.; Szumowski, J.D.; Dilworth, S.; Oskarsson, J.; Shiels, M.; Havlir, D.; et al. First Demonstration Project of Long-Acting Injectable Antiretroviral Therapy for Persons with and without Detectable Human Immunodeficiency Virus (HIV) Viremia in an Urban HIV Clinic. Clin. Infect. Dis. 2023, 76, e645–e651. [Google Scholar] [CrossRef] [PubMed]
- Edwards, G.G.; Miyashita-Ochoa, A.; Castillo, E.G.; Goodman-Meza, D.; Kalofonos, I.; Landovitz, R.J.; Leibowitz, A.A.; Pulsipher, C.; El Sayed, E.; Shoptaw, S.; et al. Long-Acting Injectable Therapy for People with HIV: Looking Ahead with Lessons from Psychiatry and Addiction Medicine. AIDS Behav. 2023, 27, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Ullah Nayan, M.; Sillman, B.; Hasan, M.; Deodhar, S.; Das, S.; Sultana, A.; Thai Hoang Le, N.; Soriano, V.; Edagwa, B.; Gendelman, H.E. Advances in Long-Acting Slow Effective Release Antiretroviral Therapies for Treatment and Prevention of HIV Infection. Adv. Drug Deliv. Rev. 2023, 200, 115009. [Google Scholar] [CrossRef]
- Cunha, R.F.; Simões, S.; Carvalheiro, M.; Pereira, J.M.A.; Costa, Q.; Ascenso, A. Novel Antiretroviral Therapeutic Strategies for HIV. Molecules 2021, 26, 5305. [Google Scholar] [CrossRef] [PubMed]
- Kirtane, A.R.; Abouzid, O.; Minahan, D.; Bensel, T.; Hill, A.L.; Selinger, C.; Bershteyn, A.; Craig, M.; Mo, S.S.; Mazdiyasni, H.; et al. Development of an Oral Once-Weekly Drug Delivery System for HIV Antiretroviral Therapy. Nat. Commun. 2018, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Oti, V.B. Nanoparticles and Its Implications in HIV/AIDS Therapy. Curr. Drug Discov. Technol. 2020, 17, 448–456. [Google Scholar] [CrossRef]
- Rajoli, R.K.R.; Flexner, C.; Chiong, J.; Owen, A.; Donnelly, R.F.; Larrañeta, E.; Siccardi, M. Modelling the Intradermal Delivery of Microneedle Array Patches for Long-Acting Antiretrovirals Using PBPK. Eur. J. Pharm. Biopharm. 2019, 144, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Baeten, J.M.; Hendrix, C.W.; Hillier, S.L. Topical Microbicides in HIV Prevention: State of the Promise. Annu. Rev. Med. 2020, 71, 361–377. [Google Scholar] [CrossRef]
- Walsh, S.R.; Seaman, M.S. Broadly Neutralizing Antibodies for HIV-1 Prevention. Front. Immunol. 2021, 12, 712122. [Google Scholar] [CrossRef]
- Griffith, S.A.; McCoy, L.E. To BnAb or Not to BnAb: Defining Broadly Neutralising Antibodies Against HIV-1. Front. Immunol. 2021, 12, 708227. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Mechanism | Nervous System | Cardiovascular | Digestive System | Liver | Renal | Skin | Metabolism | Other |
|---|---|---|---|---|---|---|---|---|---|
| NRTIs: emtricitabine, zidovudine, didanosine, stavudine, abacavir, lamivudin, tenofovir alafenamide, tenofovir disoproxil fumarate | reverse transcriptase inhibition by competing with natural nucleosides | headache, peripheral neuropathy | cardiomyopathy | nausea, diarrhea, pancreatitis | hepatic steatosis | Fanconi’s syndrome, renal insufficiency | rash | lactic acidosis, lipodystrophy, hyperlactatemia | hypersensitivity reaction, lipoatrophy, thrombocytopenia, anemia, myelosuppression |
| NNRTIs: nevirapine, delavirdine, efavirenz, etravirine, rilpivirine, doravirine | reverse transcriptase polymerization inhibition by binding to the NNRTI binding pocket | depression, sleep disturbance, headache | nausea, diarrhea | hepatitis | rash | dyslipidemia | neutropenia | ||
| INSTIs: bictegravir, dolutegravir, elvitegravir, raltegravir, cabotegravir | integrase inhibition | intracranial hemorrhage, sleep disturbance, headache | myocardial infarction | nausea, diarrhea | unconjugated hyperbilirubinemia | nephrolithiasis, renal insufficiency | rash | weight gain | myopathy, rhabdomyolysis |
| PIs: atazanavir, darunavir, fosamprenavir, indinavir, lopinavir, nelfinavir, ritonavir, saquinavir, tipranavir | protease inhibition | dizziness, headache | QT prolongation | nausea, diarrhea | hepatotoxicity | nephrolithiasis, renal insufficiency | rash | dyslipidemia, hyperglycemia, fat maldistribution | |
| Maraviroc | binds to CCR5, prevents the interaction of HIV-1 gp120 with CCR5-tropic HIV-1, and inhibits the virus from entering the cell | dizziness, headache | postural hypotension | hepatitis | renal insufficiency | ||||
| Fostemsavir | (prodrug) binds the viral envelope protein gp120 on HIV-1, prevents the conformational change required for attachment of HIV-1 to the host cell | headache, neuropathy | QT prolongation | nausea, vomiting, abdominal pain, diarrhea | rash | myalgia | |||
| Enfuvirtide | binds to HR1 in the gp41 subunit of the viral envelope glycoprotein, prevents the conformational changes required for the fusion of viral and cellular membranes | dizziness, headache, sleep disturbance | nausea, diarrhea | rash | injection site nodules, hypersensitivity | ||||
| Ibalizumab | blocks gp120-CD4 complex conformational alterations that allow for co-receptor attachment and fusion | dizziness, headache | nausea, diarrhea | immune system disorders | |||||
| Long-acting injectable antiretroviral drugs (cabotegravir-rilpivirine) | INSTI + NNRTI | pyrexia, headache, sleep disorders, dizziness, depression | nausea | abnormalities in aspartate aminotransferase, alanine aminotransferase, total bilirubin | rash | fatigue, injection site reactions, musculoskeletal pain |
| Drug | Mutation | ||
|---|---|---|---|
| Major | Accessory | ||
| First-generation INSTI | Raltegravir [156] | R263K, N155H, Q148HKR, Y143RHC, F121Y | L74M, E138AK, G140AS, E92Q, T97A |
| Elvitegravir [157,158] | R263K, N155H, Q148HKR, S147G, F121Y, E92Q, T66I | T97A, E92G, T66AK | |
| Second-generation INSTI | Dolutegravir [159] | R263K, Q148HKR, G118R | N155H, G140AS, E138AKT, T66K, E92Q, F121Y |
| Bictegravir [160] | R263K, Q148H | G140S, E138K, T66K, E92Q, G118R | |
| Cabotegravir [160] | R263K, Q148HKR, G140R, G118R | N155H, S153FY, G140ACS, E138AKT, T66K | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apetroaei, M.-M.; Velescu, B.Ș.; Nedea, M.I.; Dinu-Pîrvu, C.E.; Drăgănescu, D.; Fâcă, A.I.; Udeanu, D.I.; Arsene, A.L. The Phenomenon of Antiretroviral Drug Resistance in the Context of Human Immunodeficiency Virus Treatment: Dynamic and Ever Evolving Subject Matter. Biomedicines 2024, 12, 915. https://doi.org/10.3390/biomedicines12040915
Apetroaei M-M, Velescu BȘ, Nedea MI, Dinu-Pîrvu CE, Drăgănescu D, Fâcă AI, Udeanu DI, Arsene AL. The Phenomenon of Antiretroviral Drug Resistance in the Context of Human Immunodeficiency Virus Treatment: Dynamic and Ever Evolving Subject Matter. Biomedicines. 2024; 12(4):915. https://doi.org/10.3390/biomedicines12040915
Chicago/Turabian StyleApetroaei, Miruna-Maria, Bruno Ștefan Velescu, Marina Ionela (Ilie) Nedea, Cristina Elena Dinu-Pîrvu, Doina Drăgănescu, Anca Ionela Fâcă, Denisa Ioana Udeanu, and Andreea Letiția Arsene. 2024. "The Phenomenon of Antiretroviral Drug Resistance in the Context of Human Immunodeficiency Virus Treatment: Dynamic and Ever Evolving Subject Matter" Biomedicines 12, no. 4: 915. https://doi.org/10.3390/biomedicines12040915