

The Roles of AGTRAP, ALKBH3, DIVERSIN, NEDD8 and RRM1 in Glioblastoma Pathophysiology and Prognosis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
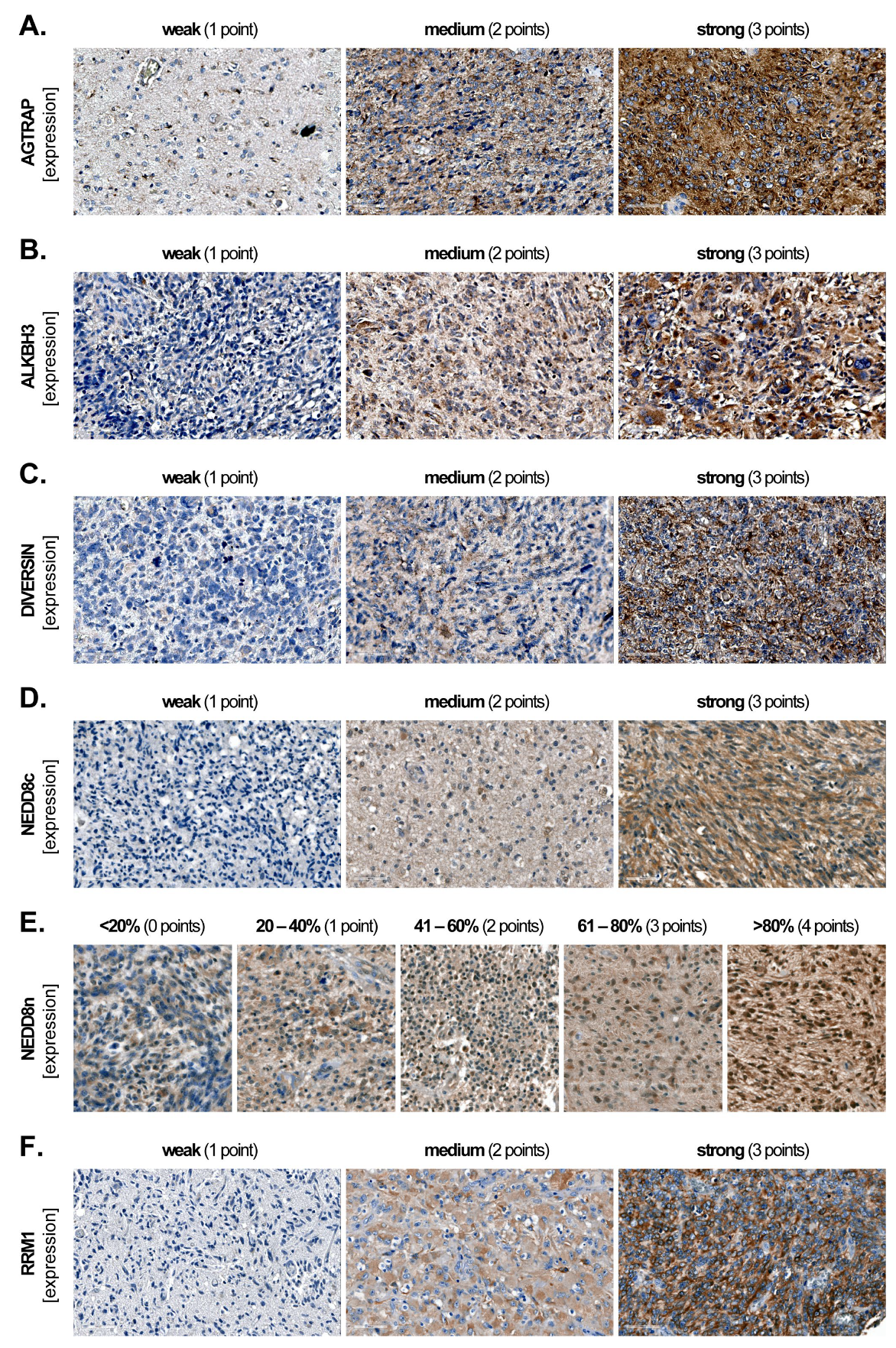
2.2. Marker Expression and Scoring
2.3. Statistical Analysis
3. Results
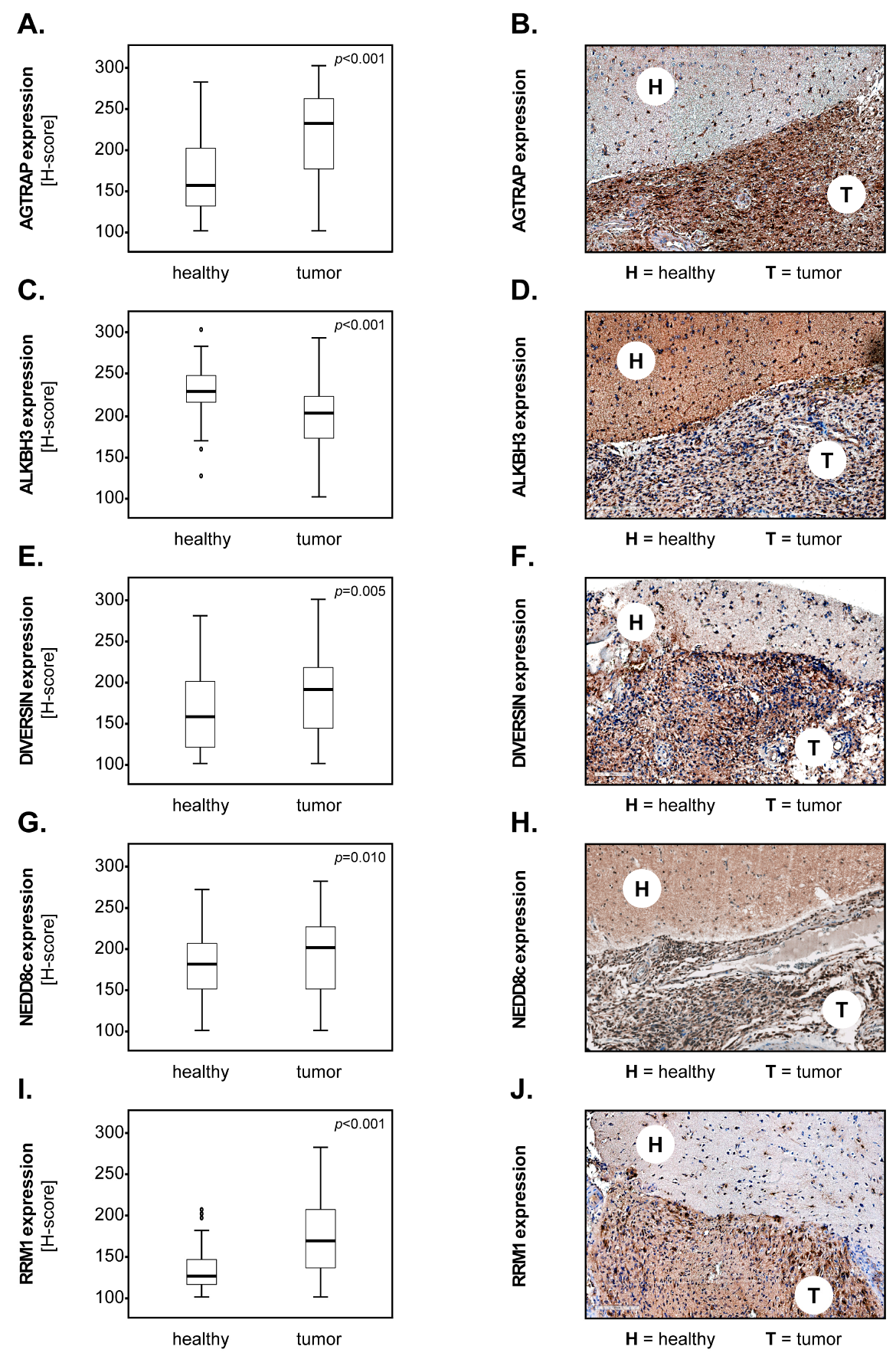
3.1. Marker Expression in Tumour versus Healthy Brain Tissues
3.2. Marker Expression and the Overall Survival of GBM Patients
3.3. Marker Expression and the Progression-Free Survival of GBM Patients
3.4. Marker Combinations and the Survival of GBM Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Cioffi, G.; Gittleman, H.; Patil, N.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2012–2016. Neuro Oncol. 2019, 21, v1–v100. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.C.; Ashley, D.M.; Lopez, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ilasaca, M.; Liu, X.; Tamura, K.; Dzau, V.J. The angiotensin II type I receptor-associated protein, ATRAP, is a transmembrane protein and a modulator of angiotensin II signaling. Mol. Biol. Cell 2003, 14, 5038–5050. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, W.; Li, X.; Zhang, L.; Gao, Y.; Peng, Q.; Du, C.; Jiang, N. AGTRAP Is a Prognostic Biomarker Correlated with Immune Infiltration in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 713017. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jin, X.; Lei, M.; Jiang, Y.; Liu, Y.; Yu, F.; Guo, Y.; Han, B.; Yang, Y.; Sun, W.; et al. USF1-ATRAP-PBX3 Axis Promote Breast Cancer Glycolysis and Malignant Phenotype by Activating AKT/mTOR Signaling. Int. J. Biol. Sci. 2022, 18, 2452–2471. [Google Scholar] [CrossRef]
- Hong, K.; Zhang, Y.; Yao, L.; Zhang, J.; Sheng, X.; Song, L.; Guo, Y.; Guo, Y. Pan-cancer analysis of the angiotensin II receptor-associated protein as a prognostic and immunological gene predicting immunotherapy responses in pan-cancer. Front. Cell Dev. Biol. 2022, 10, 913684. [Google Scholar] [CrossRef]
- Rajecka, V.; Skalicky, T.; Vanacova, S. The role of RNA adenosine demethylases in the control of gene expression. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 343–355. [Google Scholar] [CrossRef]
- Hotta, K.; Sho, M.; Fujimoto, K.; Shimada, K.; Yamato, I.; Anai, S.; Harada, H.; Tsujikawa, K.; Konishi, N.; Shinohara, N.; et al. Clinical significance and therapeutic potential of prostate cancer antigen-1/ALKBH3 in human renal cell carcinoma. Oncol. Rep. 2015, 34, 648–654. [Google Scholar] [CrossRef]
- Pilzys, T.; Marcinkowski, M.; Kukwa, W.; Garbicz, D.; Dylewska, M.; Ferenc, K.; Mieczkowski, A.; Kukwa, A.; Migacz, E.; Wolosz, D.; et al. ALKBH overexpression in head and neck cancer: Potential target for novel anticancer therapy. Sci. Rep. 2019, 9, 13249. [Google Scholar] [CrossRef] [PubMed]
- Yamato, I.; Sho, M.; Shimada, K.; Hotta, K.; Ueda, Y.; Yasuda, S.; Shigi, N.; Konishi, N.; Tsujikawa, K.; Nakajima, Y. PCA-1/ALKBH3 contributes to pancreatic cancer by supporting apoptotic resistance and angiogenesis. Cancer Res. 2012, 72, 4829–4839. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Puig, R.; Climent, F.; Piñeyro, D.; Domingo-Domènech, E.; Davalos, V.; Encuentra, M.; Rea, A.; Espejo-Herrera, N.; Soler, M.; Lopez, M.; et al. Epigenetic loss of m1A RNA demethylase ALKBH3 in Hodgkin lymphoma targets collagen, conferring poor clinical outcome. Blood 2021, 137, 994–999. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, O.A.; Hermanowicz, S.; van der Horst, J.; Hilmarsdottir, H.; Staszczak, Z.; Jonasson, J.G.; Tryggvadottir, L.; Gudjonsson, T.; Sigurdsson, S. CpG promoter methylation of the ALKBH3 alkylation repair gene in breast cancer. BMC Cancer 2017, 17, 469. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.T.; Yan, Y.L.; Cai, Y.; Peng, B.; Li, J.N.; Huang, J.Z.; Xu, Z.J.; Zhou, J.H. ALKBH1-8 and FTO: Potential Therapeutic Targets and Prognostic Biomarkers in Lung Adenocarcinoma Pathogenesis. Front. Cell Dev. Biol. 2021, 9, 633927. [Google Scholar] [CrossRef]
- Schwarz-Romond, T.; Asbrand, C.; Bakkers, J.; Kuhl, M.; Schaeffer, H.J.; Huelsken, J.; Behrens, J.; Hammerschmidt, M.; Birchmeier, W. The ankyrin repeat protein Diversin recruits Casein kinase Iepsilon to the beta-catenin degradation complex and acts in both canonical Wnt and Wnt/JNK signaling. Genes. Dev. 2002, 16, 2073–2084. [Google Scholar] [CrossRef]
- Luan, L.; Li, N.Y.; Zhang, K.Y.; Wang, X.J.; Pan, H. Diversin upregulates the proliferative ability of colorectal cancer by inducing cell cycle proteins. Exp. Mol. Pathol. 2023, 129, 104850. [Google Scholar] [CrossRef] [PubMed]
- Luan, L.; Zhao, Y.; Xu, Z.; Jiang, G.; Zhang, X.; Fan, C.; Liu, D.; Zhao, H.; Xu, K.; Wang, M.; et al. Diversin increases the proliferation and invasion ability of non-small-cell lung cancer cells via JNK pathway. Cancer Lett. 2014, 344, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.H.; Yu, X.M.; Dong, Q.Z.; Wang, Y.J. Diversin is overexpressed in human gliomas and its depletion inhibits proliferation and invasion. Tumor Biol. 2014, 35, 7905–7909. [Google Scholar] [CrossRef]
- Yu, X.; Wang, M.; Dong, Q.; Jin, F. Diversin is overexpressed in breast cancer and accelerates cell proliferation and invasion. PLoS ONE 2014, 9, e98591. [Google Scholar] [CrossRef]
- Bai, R.; Wu, D.H.; Shi, Z.; Hu, W.X.; Li, J.; Chen, Y.Y.; Ge, W.T.; Yuan, Y.; Zheng, S. Pan-cancer analyses demonstrate that ANKRD6 is associated with a poor prognosis and correlates with M2 macrophage infiltration in colon cancer. Chin. J. Cancer Res. 2021, 33, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Kamitani, T.; Kito, K.; Nguyen, H.P.; Yeh, E.T. Characterization of NEDD8, a developmentally down-regulated ubiquitin-like protein. J. Biol. Chem. 1997, 272, 28557–28562. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.S.; Jiang, Y.Y.; Luo, Q.; Li, L.H.; Jia, L.J. Neddylation: A novel modulator of the tumor microenvironment. Mol. Cancer 2019, 18, 77. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Jia, L. Targeting Protein Neddylation for Cancer Therapy. Adv. Exp. Med. Biol. 2020, 1217, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, P.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 2006, 75, 681–706. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.W.; Lee, B.; Park, W.S.; Han, N.; Hong, E.K.; Park, E.Y.; Han, S.S.; Park, S.J.; Kim, T.H.; Lee, W.J.; et al. RRM1 Expression as a Prognostic Biomarker for Unresectable or Recurrent Biliary Tract Cancer Treated with Gemcitabine plus Cisplatin. J. Clin. Med. 2021, 10, 4652. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.Y.; Tan, Y.; Pan, L.L.; Tian, Z.F.; Lin, L.J.; Su, M.X.; Ou, G.S.; Chen, Y.T. Prognostic value of RRM1 and its effect on chemoresistance in pancreatic cancer. Cancer Chemother. Pharm. 2023, 93, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.C.; Liu, X.Y.; Zhou, J.C.; Huang, Y.S.; Zhang, S.J.; Shen, J.G.; Loera, S.; Yuan, X.M.; Chen, W.J.; Jin, M.; et al. Ribonucleotide Reductase Large Subunit M1 Predicts Poor Survival Due to Modulation of Proliferative and Invasive Ability of Gastric Cancer. PLoS ONE 2013, 8, e70191. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.M.; Lian, X.Y.; Bi, Y.H.; Hu, C.C.; Liang, Y.W.; Li, Q.S. Prognostic value of ribonucleotide reductase subunit M1 (RRM1) in non-small cell lung cancer: A meta-analysis. Clin. Chim. Acta 2018, 485, 67–73. [Google Scholar] [CrossRef]
- Jordheim, L.P.; Seve, P.; Tredan, O.; Dumontet, C. The ribonucleotide reductase large subunit (RRM1) as a predictive factor in patients with cancer. Lancet Oncol. 2011, 12, 693–702. [Google Scholar] [CrossRef]
- Jiang, K.; Zhi, T.L.; Xu, W.H.; Xu, X.P.; Wu, W.N.; Yu, T.F.; Nie, E.; Zhou, X.; Bao, Z.Y.; Jin, X.; et al. MicroRNA-1468-5p inhibits glioma cell proliferation and induces cell cycle arrest by targeting RRM1. Am. J. Cancer Res. 2017, 7, 784–800. [Google Scholar] [PubMed]
- Ariey-Bonnet, J.; Berges, R.; Montero, M.P.; Mouysset, B.; Piris, P.; Muller, K.; Pinna, G.; Failes, T.W.; Arndt, G.M.; Morando, P.; et al. Combination drug screen targeting glioblastoma core vulnerabilities reveals pharmacological synergisms. Ebiomedicine 2023, 95, 104752. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, C.A.; Schroder, H.; Schafer, F.T.A.; Aust, J.F.; Kresse, N.; Siebert, C.L.R.; Stein, K.P.; Haghikia, A.; Wilkens, L.; Mawrin, C.; et al. Progesterone Receptor Membrane Component 1 (PGRMC1) Modulates Tumour Progression, the Immune Microenvironment and the Response to Therapy in Glioblastoma. Cells 2023, 12, 2498. [Google Scholar] [CrossRef] [PubMed]
- Kresse, N.; Schroder, H.; Stein, K.P.; Wilkens, L.; Mawrin, C.; Sandalcioglu, I.E.; Dumitru, C.A. PLOD2 Is a Prognostic Marker in Glioblastoma That Modulates the Immune Microenvironment and Tumor Progression. Int. J. Mol. Sci. 2022, 23, 6037. [Google Scholar] [CrossRef]
- Chambless, L.B.; Kistka, H.M.; Parker, S.L.; Hassam-Malani, L.; McGirt, M.J.; Thompson, R.C. The relative value of postoperative versus preoperative Karnofsky Performance Scale scores as a predictor of survival after surgical resection of glioblastoma multiforme. J. Neuro-Oncol. 2015, 121, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Liang, H.; Cheng, P.; Yang, H.; Zhao, P. Gross Total vs. Subtotal Resection on Survival Outcomes in Elderly Patients with High-Grade Glioma: A Systematic Review and Meta-Analysis. Front. Oncol. 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Schiffgens, S.; Wilkens, L.; Brandes, A.A.; Meier, T.; Franceschi, E.; Ermani, M.; Hartmann, C.; Sandalcioglu, I.E.; Dumitru, C.A. Sex-specific clinicopathological significance of novel (Frizzled-7) and established (MGMT, IDH1) biomarkers in glioblastoma. Oncotarget 2016, 7, 55169–55180. [Google Scholar] [CrossRef] [PubMed]
- Siker, M.L.; Wang, M.; Porter, K.; Nelson, D.F.; Curran, W.J.; Michalski, J.M.; Souhami, L.; Chakravarti, A.; Yung, W.K.; Delrowe, J.; et al. Age as an independent prognostic factor in patients with glioblastoma: A Radiation Therapy Oncology Group and American College of Surgeons National Cancer Data Base comparison. J. Neuro-Oncol. 2011, 104, 351–356. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, W.; Yang, T.; He, L.; Liao, Y.; Lu, J. Construction and Comprehensive Analysis of a Stratification System Based on AGTRAP in Patients with Hepatocellular Carcinoma. Dis. Markers 2021, 2021, 6144476. [Google Scholar] [CrossRef]
- Mendaza, S.; Fernandez-Irigoyen, J.; Santamaria, E.; Zudaire, T.; Guarch, R.; Guerrero-Setas, D.; Vidal, A.; Santos-Salas, J.; Matias-Guiu, X.; Ausin, K.; et al. Absence of Nuclear p16 Is a Diagnostic and Independent Prognostic Biomarker in Squamous Cell Carcinoma of the Cervix. Int. J. Mol. Sci. 2020, 21, 2125. [Google Scholar] [CrossRef]
- Zeng, H.; Li, H.; Zhao, Y.; Chen, L.; Ma, X. Transcripto-based network analysis reveals a model of gene activation in tongue squamous cell carcinomas. Head. Neck 2019, 41, 4098–4110. [Google Scholar] [CrossRef] [PubMed]
- Vidyarthi, A.; Agnihotri, T.; Khan, N.; Singh, S.; Tewari, M.K.; Radotra, B.D.; Chatterjee, D.; Agrewala, J.N. Predominance of M2 macrophages in gliomas leads to the suppression of local and systemic immunity. Cancer Immunol. Immunother. 2019, 68, 1995–2004. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.S.; Xu, Z.J.; Peng, J.W.; Zhang, M.Y. The AlkB Family: Potential Prognostic Biomarkers and Therapeutic Targets in Glioblastoma. Front. Oncol. 2022, 12, 847821. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Majid, M.C.; Soll, J.M.; Brickner, J.R.; Dango, S.; Mosammaparast, N. Noncanonical regulation of alkylation damage resistance by the OTUD4 deubiquitinase. EMBO J. 2015, 34, 1687–1703. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.W.; Wu, Z.L.; Jiang, L.M.; Gao, J.; Wu, C.L.; Hu, H.L. Neural precursor cell expressed, developmentally downregulated 8 promotes tumor progression and predicts poor prognosis of patients with bladder cancer. Cancer Sci. 2019, 110, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Xian, J.R.; Wang, S.W.; Jiang, Y.Y.; Li, L.H.; Cai, L.L.; Chen, P.; Liu, Y.; Zeng, X.F.; Chen, G.A.; Ding, C.; et al. Overexpressed NEDD8 as a potential therapeutic target in esophageal squamous cell carcinoma. Cancer Biol. Med. 2022, 19, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.; Li, C.J.; Yang, Z.X.; Li, L.H.; Jiang, Y.A.; Yu, G.Y.; Zhu, W.; Liu, Z.Y.; Duan, S.Z.; Chu, Y.W.; et al. Suppression of glioblastoma by targeting the overactivated protein neddylation pathway. Neuro-Oncol. 2015, 17, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, M.; Parcon, P.A.; Bose, C.; Liu, L.; Jones, R.A.; Farlow, M.R.; Mrak, R.E.; Barger, S.W.; Griffin, W.S.T. Interleukin-1β drives NEDD8 nuclear-to-cytoplasmic translocation, fostering parkin activation via NEDD8 binding to the P-ubiquitin activating site. J. Neuroinflamm. 2019, 16, 275. [Google Scholar] [CrossRef] [PubMed]
- Yeung, Y.T.; McDonald, K.L.; Grewal, T.; Munoz, L. Interleukins in glioblastoma pathophysiology: Implications for therapy. Br. J. Pharmacol. 2013, 168, 591–606. [Google Scholar] [CrossRef]
- Molenaar, R.J.; Verbaan, D.; Lamba, S.; Zanon, C.; Jeuken, J.W.; Boots-Sprenger, S.H.; Wesseling, P.; Hulsebos, T.J.; Troost, D.; van Tilborg, A.A.; et al. The combination of IDH1 mutations and MGMT methylation status predicts survival in glioblastoma better than either IDH1 or MGMT alone. Neuro-Oncol. 2014, 16, 1263–1273. [Google Scholar] [CrossRef]
- Nystrom, H.; Tavelin, B.; Bjorklund, M.; Naredi, P.; Sund, M. Improved tumour marker sensitivity in detecting colorectal liver metastases by combined type IV collagen and CEA measurement. Tumour Biol. 2015, 36, 9839–9847. [Google Scholar] [CrossRef] [PubMed]
- Weller, P.; Bankfalvi, A.; Gu, X.; Dominas, N.; Lehnerdt, G.F.; Zeidler, R.; Lang, S.; Brandau, S.; Dumitru, C.A. The role of tumour FoxP3 as prognostic marker in different subtypes of head and neck cancer. Eur. J. Cancer 2014, 50, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Zemans, R.L.; Jacobson, S.; Keene, J.; Kechris, K.; Miller, B.E.; Tal-Singer, R.; Bowler, R.P. Multiple biomarkers predict disease severity, progression and mortality in COPD. Respir. Res. 2017, 18, 117. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Parameter | Number | Percentage |
|---|---|---|
| Sex | ||
| female | 85 | 45.7 |
| male | 101 | 54.3 |
| Karnofsky performance scale | ||
| 10 | 3 | 1.6 |
| 20 | 2 | 1.1 |
| 30 | 11 | 5.9 |
| 40 | 8 | 4.3 |
| 50 | 29 | 15.6 |
| 60 | 33 | 17.7 |
| 70 | 60 | 32.3 |
| 80 | 19 | 10.2 |
| 90 | 19 | 10.2 |
| 100 | 0 | 0 |
| n.d. | 2 | 1.1 |
| Therapy | ||
| surgery | 35 | 18.8 |
| surgery+RTX | 21 | 11.3 |
| surgery+CTX | 1 | 0.5 |
| surgery+RCTX | 123 | 66.1 |
| n.d. | 6 | 3.2 |
| Extent of surgical resection | ||
| total | 65 | 34.9 |
| subtotal | 111 | 59.7 |
| n.d. | 10 | 5.4 |
| MGMT methylation status | ||
| unmethylated | 70 | 37.6 |
| methylated | 116 | 62.4 |
| n.d. | 0 | 0 |
| IDH mutation status | ||
| wild-type | 186 | 100 |
| mutated | 0 | 0 |
| n.d. | 0 | 0 |
| Cox Regression: OS | HR | CI [95%] | p-Value |
|---|---|---|---|
| AGTRAPlow | 1 | ||
| AGTRAPhigh | 1.814 | 1.260–2.612 | 0.001 |
| ALKBH3high | 1 | ||
| ALKBH3low | 1.345 | 0.938–1.929 | 0.107 |
| NEDD8clow | 1 | ||
| NEDD8chigh | 1.370 | 0.980–1.914 | 0.065 |
| RRM1low (new cut-off) | 1 | ||
| RRM1high (new cut-off) | 1.746 | 1.099–2.775 | 0.018 |
| Cox Regression: PFS | HR | CI [95%] | p-Value |
|---|---|---|---|
| AGTRAPlow | 1 | ||
| AGTRAPhigh | 1.921 | 1.200–3.075 | 0.007 |
| NEDD8clow | 1 | ||
| NEDD8chigh | 1.498 | 0.979–2.293 | 0.063 |
| Cox Regression: OS | HR | CI [95%] | p-Value |
|---|---|---|---|
| Rest | 1 | ||
| AGTRAPhighALKBH3low | 2.059 | 1.343–3.157 | <0.001 |
| AGTRAPhighNEDD8chigh | 1.516 | 1.025–2.243 | 0.037 |
| AGTRAPhighRRM1high | 1.742 | 1.223–2.480 | 0.002 |
| ALKBH3lowNEDD8chigh | 1.893 | 1.263–2.836 | 0.002 |
| ALKBH3lowRRM1high | 1.576 | 1.096–2.266 | 0.014 |
| NEDD8chighRRM1high | 1.416 | 1.017–1.971 | 0.039 |
| Cox regression: PFS | HR | CI [95%] | p-value |
| Rest | 1 | ||
| AGTRAPhighNEDD8chigh | 1.834 | 1.123–2.995 | 0.015 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumitru, C.A.; Walter, N.; Siebert, C.L.R.; Schäfer, F.T.A.; Rashidi, A.; Neyazi, B.; Stein, K.-P.; Mawrin, C.; Sandalcioglu, I.E. The Roles of AGTRAP, ALKBH3, DIVERSIN, NEDD8 and RRM1 in Glioblastoma Pathophysiology and Prognosis. Biomedicines 2024, 12, 926. https://doi.org/10.3390/biomedicines12040926
Dumitru CA, Walter N, Siebert CLR, Schäfer FTA, Rashidi A, Neyazi B, Stein K-P, Mawrin C, Sandalcioglu IE. The Roles of AGTRAP, ALKBH3, DIVERSIN, NEDD8 and RRM1 in Glioblastoma Pathophysiology and Prognosis. Biomedicines. 2024; 12(4):926. https://doi.org/10.3390/biomedicines12040926
Chicago/Turabian StyleDumitru, Claudia Alexandra, Nikolas Walter, Carl Ludwig Raven Siebert, Frederik Till Alexander Schäfer, Ali Rashidi, Belal Neyazi, Klaus-Peter Stein, Christian Mawrin, and Ibrahim Erol Sandalcioglu. 2024. "The Roles of AGTRAP, ALKBH3, DIVERSIN, NEDD8 and RRM1 in Glioblastoma Pathophysiology and Prognosis" Biomedicines 12, no. 4: 926. https://doi.org/10.3390/biomedicines12040926