

The Efficacy of Hispidin and Magnesium Nanoparticles against Zearalenone-Induced Fungal Toxicity Causing Polycystic Ovarian Syndrome in Rats
 , ,
, ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. Preparation of Zea and Hispidin Solutions
2.4. Experimental Design
2.5. Sampling and Tissue Preparation
2.6. Biochemical Assays
2.7. Determination of Biomarkers of Oxidative Stress
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) for Aromatase (Cyp19α1)
2.9. Histopathological Analysis
2.10. Statistical Analysis
3. Results
3.1. Hormones and Biochemical Markers
3.1.1. Hormones
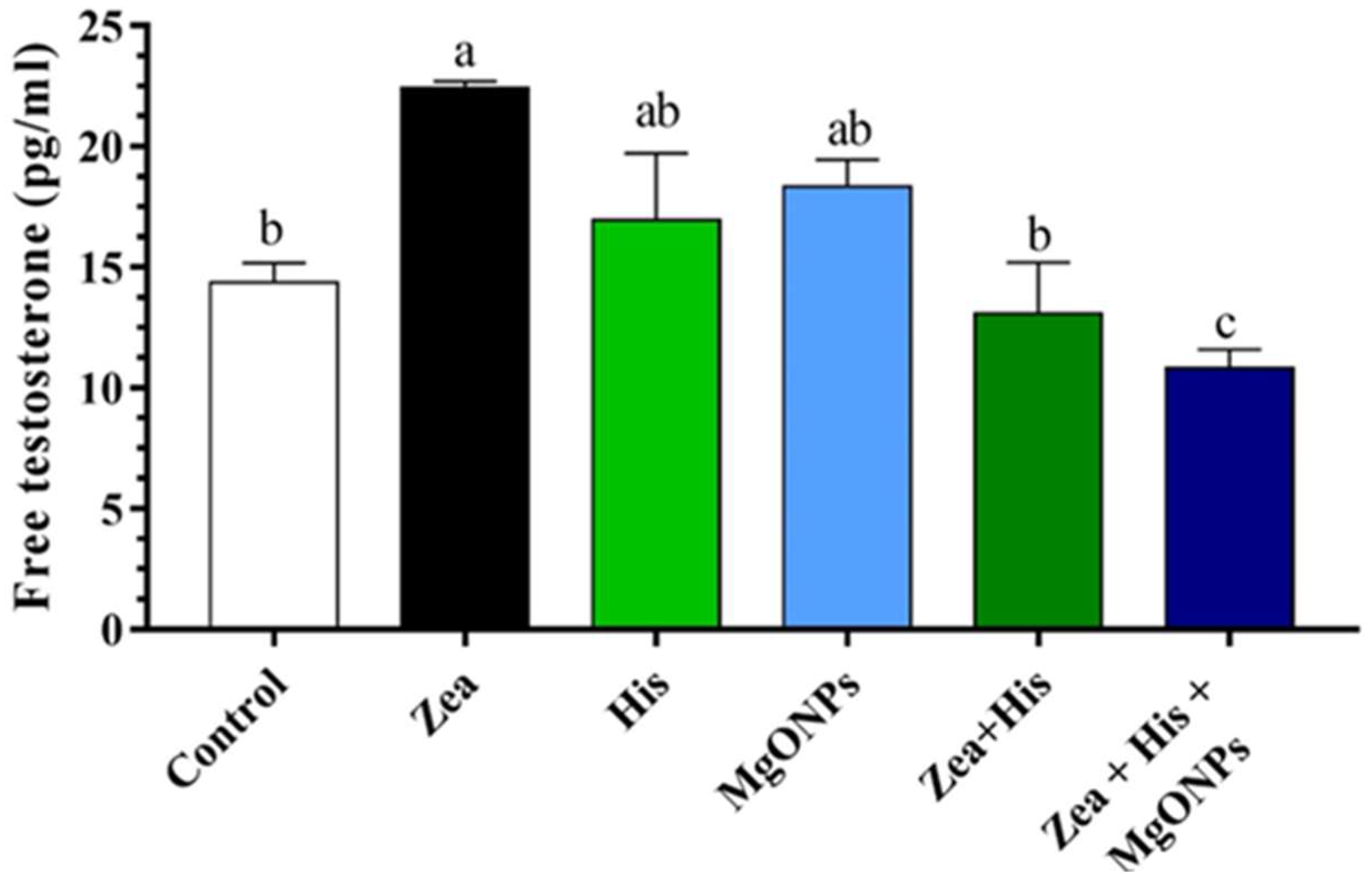
Free Testosterone
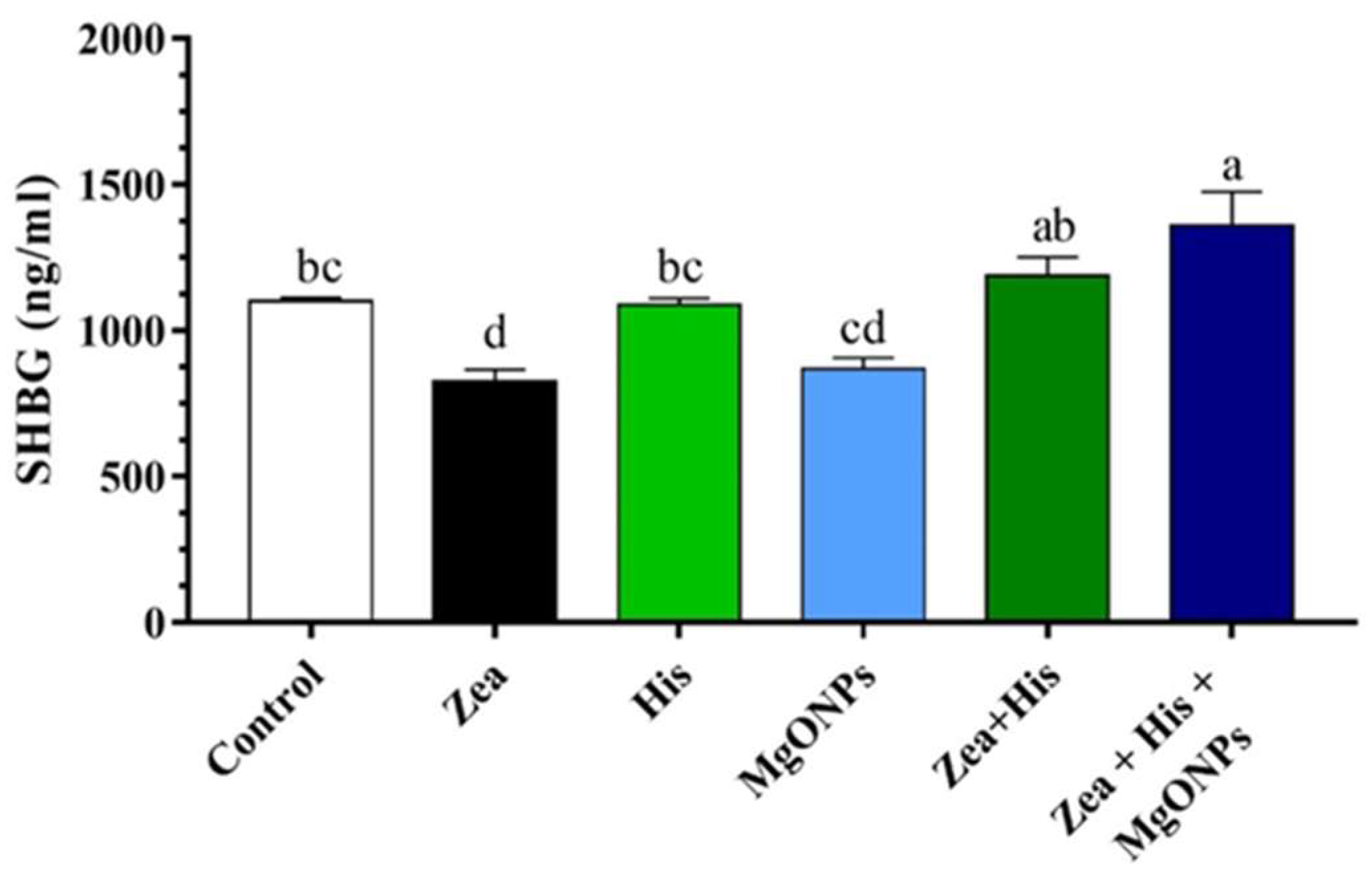
Sex Hormone-Binding Globulin (SHBG)
Luteinizing Hormone (LH)
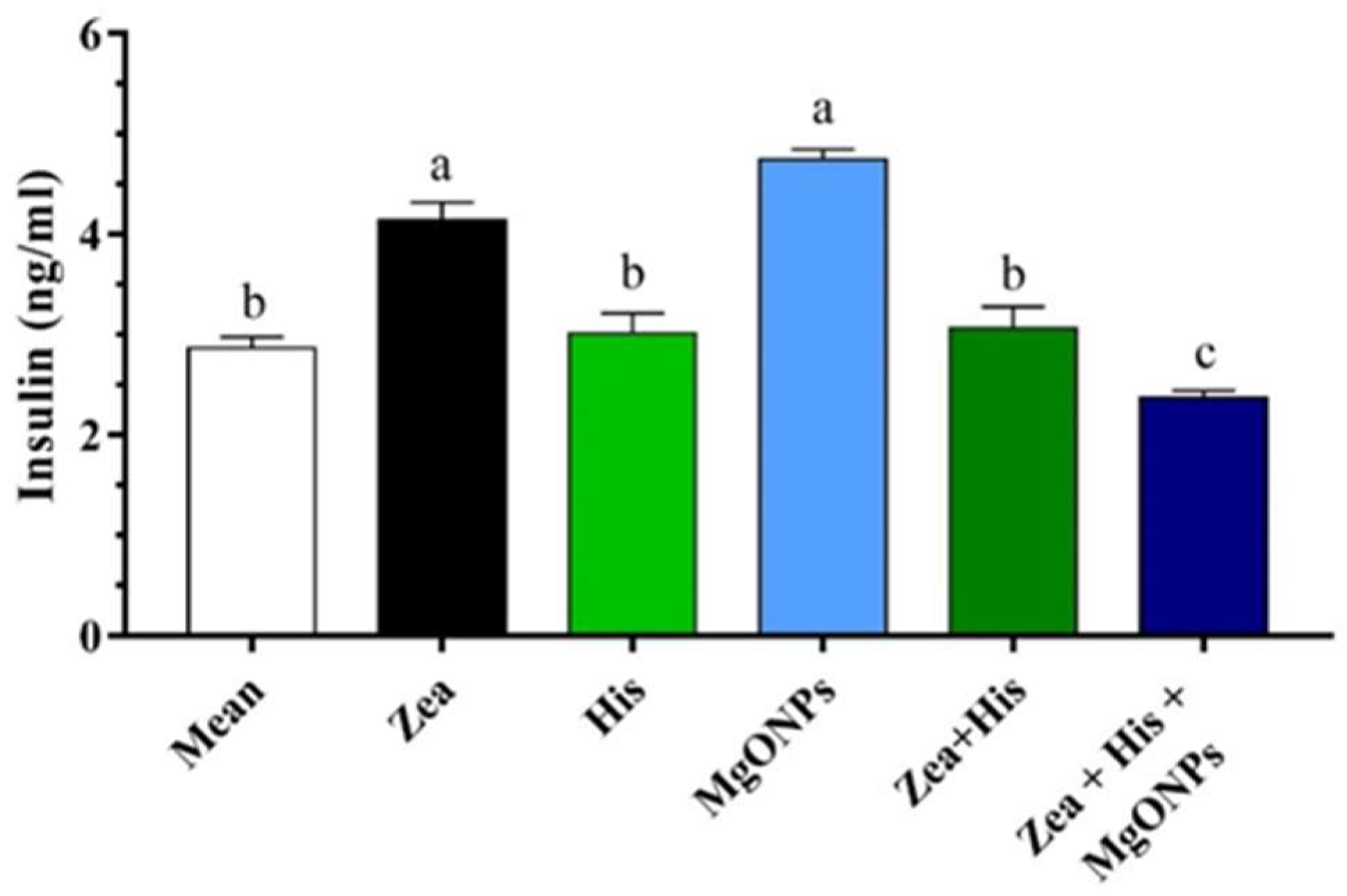
Insulin
3.1.2. Biochemical Markers
Glucose
Lipid Profiles
3.2. Oxidative and Antioxidative Biomarkers
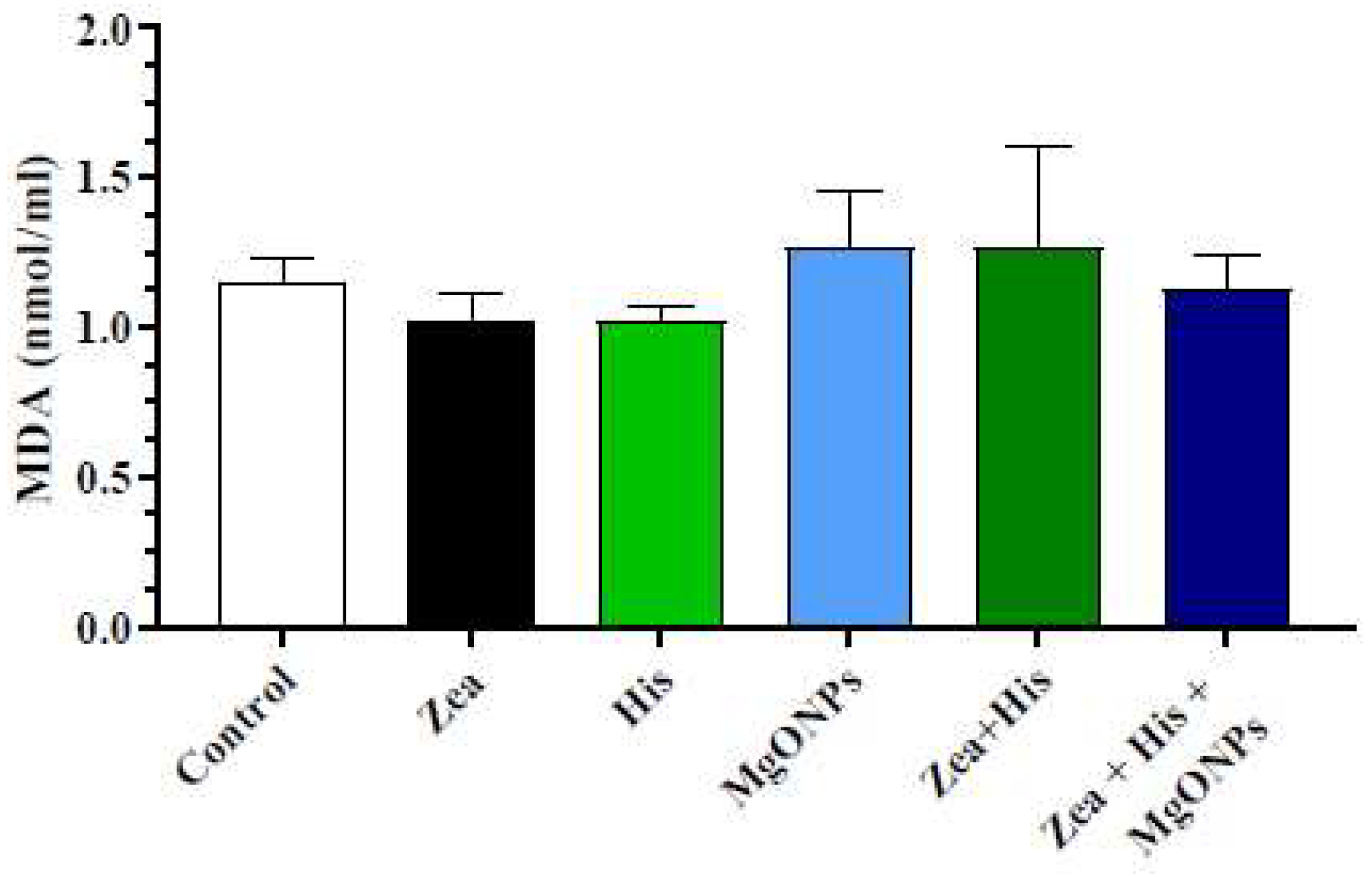
3.2.1. Lipid Peroxides (Malonaldehyde, MDA)
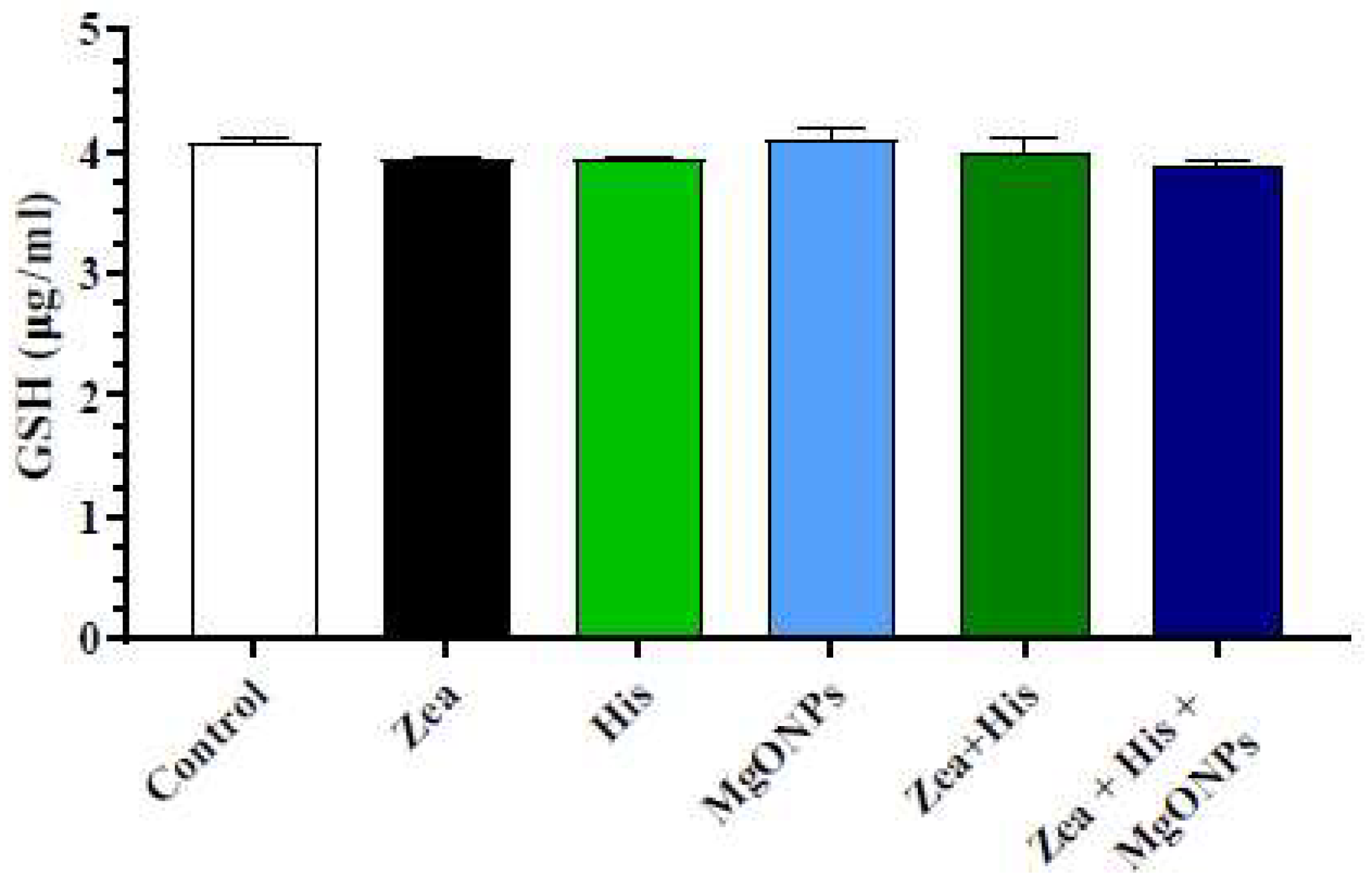
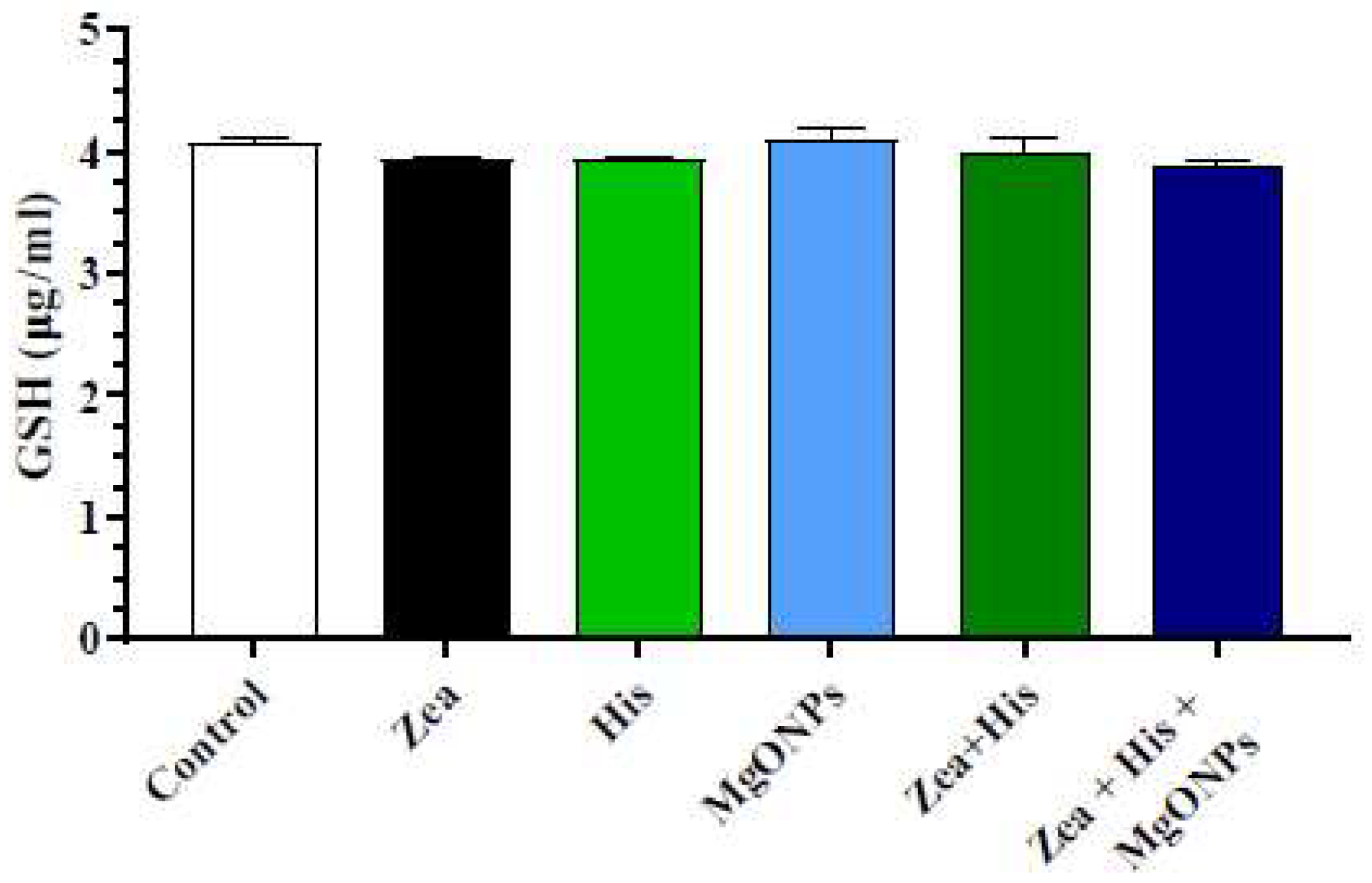
3.2.2. Glutathione (GSH)
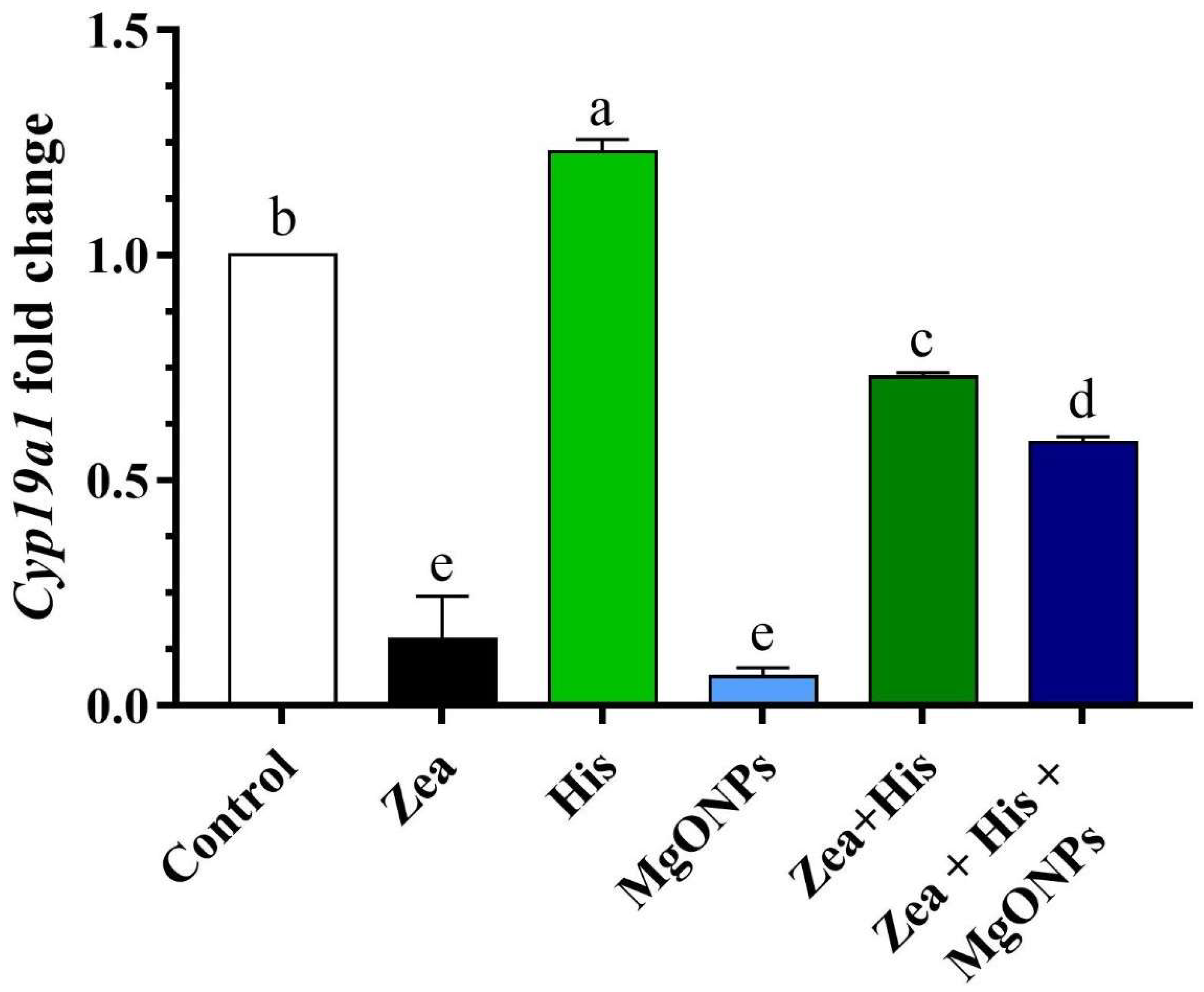
3.3. Aromatase (Cyp19α1) Gene Expression in Ovaries
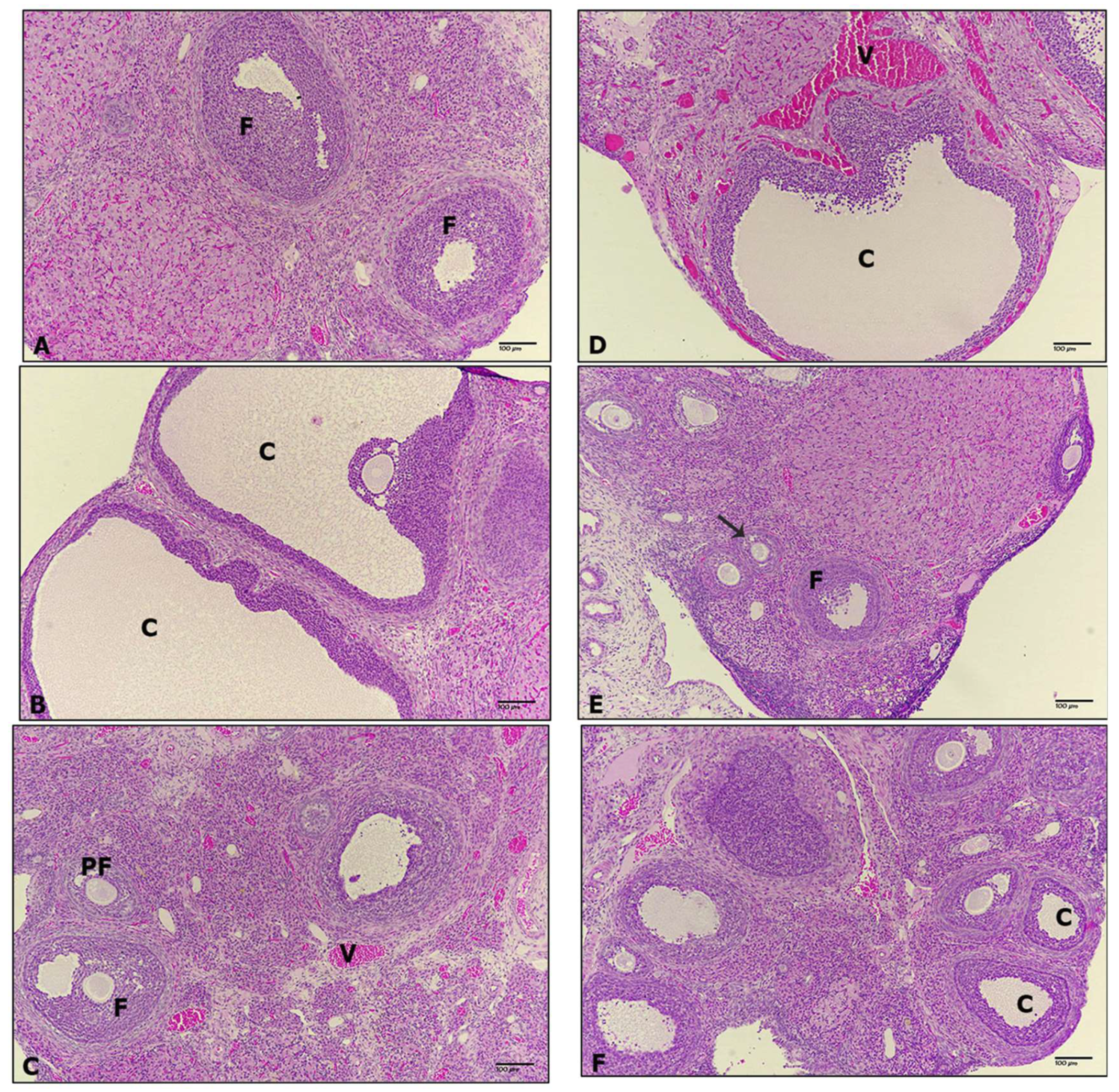
3.4. Ovarian Histopathology
4. Discussion
5. Conclusions
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang’Ethe, E.K.; A Lang’A, K. Aflatoxin B1 and M1 contamination of animal feeds and milk from urban centers in Kenya. Afr. Health Sci. 2009, 9, 218–226. [Google Scholar] [PubMed]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaal, B.; Salama, S.; Al-Qasmi, N.; Jaganjac, M. Mycotoxin contamination of food and feed in the Gulf Cooperation Council countries and its detection. Toxicon 2019, 171, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Tittlemier, S.A.; Cramer, B.; Dall’Asta, C.; Iha, M.H.; Lattanzio, V.M.T.; Maragos, C.; Solfrizzo, M.; Stranska, M.; Stroka, J.; Sumarah, M. Developments in mycotoxin analysis: An update for 2018-19. World Mycotoxin J. 2020, 13, 3–24. [Google Scholar] [CrossRef]
- Tiemann, U.; Brü, K.-P.; Jonas, L.; Pö, R.; Schneider, F.; Dä, S. Effects of diets with cereal grains contaminated by graded levels of two Fusarium toxins on selected immunological and histological measurements in the spleen of gilts 1,2. J. Anim. Sci. 2006, 84, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A.; Leslie, J.F. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135–144. [Google Scholar] [CrossRef]
- Boeira, S.P.; Filho, C.B.; Del’Fabbro, L.; Roman, S.S.; Royes, L.F.F.; Fighera, M.R.; Jessé, C.R.; Oliveira, M.S.; Furian, A.F. Lycopene treatment prevents hematological, reproductive and histopathological damage induced by acute zearalenone administration in male Swiss mice. Exp. Toxicol. Pathol. 2014, 66, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.E.; Song, J.; Lee, H.J.; Kim, M.; Kim, D.W.; Jung, H.J.; Kim, B.; Lee, Y.; Yu, D.; Kim, D.W.; et al. Effects of high levels of deoxynivalenol and zearalenone on growth performance, and hematological and immunological parameters in pigs. Toxins 2018, 10, 114. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xue, K.S.; Sun, X.; Williams, P.L.; Wang, J.S.; Tang, L. Toxicogenomic responses to zearalenone in Caenorhabditis elegans reveal possible molecular mechanisms of reproductive toxicity. Food Chem. Toxicol. 2018, 122, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.S.; Takagi, M.; Talukder, A.K.; Marey, M.A.; Kowsar, R.; Abdel-Razek, A.R.K.; Shimizu, T.; Fink-Gremmels, J.; Miyamoto, A. Zearalenone (ZEN) disrupts the anti-inflammatory response of bovine oviductal epithelial cells to sperm in vitro. Reprod. Toxicol. 2017, 74, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones: A review. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, C.; Ma, Y.; Xiao, J.; Luo, G.; Li, Y.; Wu, D. Multi-system reproductive metabolic disorder: Significance for the pathogenesis and therapy of polycystic ovary syndrome (PCOS). Life Sci. 2019, 228, 167–175. [Google Scholar] [CrossRef]
- Ghanati, K.; Mahdi, J.; Shakoori, A.; Saeed, A.-B.; Ali, K.-R.; Parisa, S. The association between polycystic ovary syndrome and environmental pollutants based on animal and human study; a systematic review. Rev. Environ. Health 2023. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Koymans, L.; Kamataki, T.; Stegeman, J.J.; Feyereisen, R.; Waxman, D.J.; Waterman, M.R.; Gotoh, O.; Coon, M.J.; Estabrook, R.W.; et al. P450 superfamily: Update on new sequences, gene mapping, accession numbers and nomenclature. Pharmacogenetics 1996, 6, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.M.; Hryb, D.J.; Nakhla, A.M.; Romas, N.A.; Rosner, W. Beyond Carrier Proteins Sex hormone-binding globulin is synthesized in target cells The sex hormone-binding globulin (SHBG) gene. J. Endocrinol. 2002, 175, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.L.; Bhasin, S.; Wu, F.C.W.; Krishna, M.; Matsumoto, A.M.; Jasuja, R. A reappraisal of testosterone’s binding in circulation: Physiological and clinical implications. Endocr. Rev. 2017, 38, 302–324. [Google Scholar] [CrossRef]
- Deswal, R.; Yadav, A.; Dang, A.S. Sex hormone binding globulin—An important biomarker for predicting PCOS risk: A systematic review and meta-analysis. Syst. Biol. Reprod. Med. 2018, 64, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.S.; Bae, K.; Kuk Yoo, J.; Ryoo, I.J.; Kim, B.Y.; Seog Ahn, J.; Yoo, I.D. Inhibition of human neutrophil elastase by ergosterol derivatives from the mycelium of Phellinus linteus. J. Antibiot. 2012, 65, 437–440. [Google Scholar] [CrossRef]
- Chen, W.; Feng, L.; Shen, Y.; Su, H.; Li, Y.; Zhuang, J.; Zhang, L.; Zheng, X. Myricitrin inhibits acrylamide-mediated cytotoxicity in human caco-2 cells by preventing oxidative stress. BioMed Res. Int. 2013, 2013, 724183. [Google Scholar] [CrossRef] [PubMed]
- Abbasian, N.; Momtaz, S.; Baeeri, M.; Navaei-Nigjeh, M.; Hosseini, R.; Abdollahi, M. Molecular and biochemical evidence on the role of zearalenone in rat polycystic ovary. Toxicon 2018, 154, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Mazaheri, N.; Naghsh, N.; Karimi, A.; Salavati, H. In vivo toxicity investigation of magnesium oxide nanoparticles in rat for environmental and biomedical applications. Iran. J. Biotechnol. 2019, 17, e1543. [Google Scholar] [CrossRef] [PubMed]
- Rezvanfar, M.A.; Shojaei Saadi, H.A.; Gooshe, M.; Abdolghaffari, A.H.; Baeeri, M.; Abdollahi, M. Ovarian aging-like phenotype in the hyperandrogenism-induced murine model of polycystic ovary. Oxidative Med. Cell. Longev. 2014, 2014, 948951. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, Y.; Ma, L.; Chen, X.; Lü, Y.; Ge, F.; Chen, Y.; Chen, X.; Lian, Q.; Jin, X.D.; et al. Zearalenone delays rat leydig cell regeneration. Toxicol. Sci. 2018, 164, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Del Fabbro, L.; Jesse, C.R.; de Gomes, M.G.; Borges Filho, C.; Donato, F.; Souza, L.C.; Goes, A.R.; Furian, A.F.; Boeira, S.P. The flavonoid chrysin protects against zearalenone induced reproductive toxicity in male mice. Toxicon 2019, 165, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yang, S.; Zhang, W.; Zhang, Y.; Li, P.; Guo, Y.; Wang, Y.; Chen, X.; He, J. The Influence of Selenium Yeast on Hematological, Biochemical and Reproductive Hormone Level Changes in Kunming Mice Following Acute Exposure to Zearalenone. Biol. Trace Elem. Res. 2016, 174, 362–368. [Google Scholar] [CrossRef]
- Rezvanfar, M.A.; Rezvanfar, M.A.; Ahmadi, A.; Shojaei-Saadi, H.A.; Baeeri, M.; Abdollahi, M. Molecular mechanisms of a novel selenium-based complementary medicine which confers protection against hyperandrogenism-induced polycystic ovary. Theriogenology 2012, 78, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Erickson, G.F.; Magoffin, D.A.; Cragun, J.R.; Chang, R.J. The Effects of Insulin and Insulin-Like Growth Factors-I and-II on Estradiol Production by Granulosa Cells of Polycystic Ovaries. J. Clin. Endocrinol. Metab. 1990, 70, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of Insulin Synthesis and Secretion and Pancreatic Beta-Cell Dysfunction in Diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- González, F. Inflammation in Polycystic Ovary Syndrome: Underpinning of insulin resistance and ovarian dysfunction. Steroids 2012, 77, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Ortega, I.; Villanueva, J.A.; Wong, D.H.; Cress, A.B.; Sokalska, A.; Stanley, S.D.; Duleba, A.J. Resveratrol potentiates effects of simvastatin on inhibition of rat ovarian theca-interstitial cells steroidogenesis. J. Ovarian Res. 2014, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Et Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Spaczynski, R.Z.; Arici, A.; Duleba, A.J. Tumor Necrosis Factor-Stimulates Proliferation of Rat Ovarian Theca-Interstitial Cells 1. Biol. Reprod. 1999, 61, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Hammond, G.L.; Wu, T.S.; Simard, M. Evolving utility of sex hormone-binding globulin measurements in clinical medicine. Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Saddick, S.Y. Identifying genes associated with the development of human polycystic ovary syndrome. Saudi J. Biol. Sci. 2020, 27, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.J. The reproductive phenotype in polycystic ovary syndrome. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Zhang, J.; Zhao, H.; He, B. Effect of Sex Hormone-Binding Globulin on Polycystic Ovary Syndrome: Mechanisms, Manifestations, Genetics, and Treatment. Int. J. Women’s Health 2022, 14, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Babapour, M.; Mohammadi, H.; Kazemi, M.; Hadi, A.; Rezazadegan, M.; Askari, G. Associations Between Serum Magnesium Concentrations and Polycystic Ovary Syndrome Status: A Systematic Review and Meta-analysis. Biol. Trace Elem. Res. 2021, 199, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Z.; Huang, Y.; Hu, K.; Ma, B.; Yang, Y. The effect of magnesium alone or its combination with other supplements on the markers of inflammation, OS and metabolism in women with polycystic ovarian syndrome (PCOS): A systematic review. Front. Endocrinol. 2022, 13, 974042. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Karandish, M.; Asghari Jafarabadi, M.; Heidari, L.; Nikbakht, R.; Babaahmadi Rezaei, H.; Mousavi, R. Metabolic and hormonal effects of melatonin and/or magnesium supplementation in women with polycystic ovary syndrome: A randomized, double-blind, placebo-controlled trial. Nutr. Metab. 2021, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Virk, P.; Al-mukhaizeem, N.A.R.; Bin Morebah, S.H.; Fouad, D.; Elobeid, M. Protective effect of resveratrol against toxicity induced by the mycotoxin, zearalenone in a rat model. Food Chem. Toxicol. 2020, 146, 111840. [Google Scholar] [CrossRef] [PubMed]
- El-Nekeety, A.A.; El-Kady, A.A.; Abdel-Wahhab, K.G.; Hassan, N.S.; Abdel-Wahhab, M.A. Reduction of individual or combined toxicity of fumonisin B1 and zearalenone via dietary inclusion of organo-modified nano-montmorillonite in rats. Environ. Sci. Pollut. Res. 2017, 24, 20770–20783. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Gao, J.; Liu, F.X.; Broomhead, J.; Chi, F. Effects of purified zearalenone on growth performance, organ size, serum metabolites, and oxidative stress in postweaning gilts. J. Anim. Sci. 2011, 89, 3008–3015. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Liu, M.; Qu, Z.; Zhang, Y.; Yin, S.; Shan, A. Toxic effects of zearalenone on oxidative stress, inflammatory cytokines, biochemical and pathological changes induced by this toxin in the kidney of pregnant rats. Environ. Toxicol. Pharmacol. 2014, 37, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Ren, Z.; Gao, S.; Chen, Y.; Yang, Y.; Yang, D.; Deng, J.; Zuo, Z.; Wang, Y.; Shen, L. Individual and combined effects of deoxynivalenol and zearalenone on mouse kidney. Environ. Toxicol. Pharmacol. 2015, 40, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Llorens, P.; Herrera, M.; Juan-García, A.; Payá, J.J.; Moltó, J.C.; Ariño, A.; Juan, C. Biomarkers of Exposure to Zearalenone in In Vivo and In Vitro Studies. Toxins 2022, 14, 291. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Shen, T.; Ding, Q.; Lv, Y.; Li, L.; Huang, K.; Yan, L.; Song, S. Zearalenone induces ROS-mediated mitochondrial damage in porcine IPEC-J2 cells. J. Biochem. Mol. Toxicol. 2017, 31, e21944. [Google Scholar] [CrossRef]
- Mangalampalli, B.; Dumala, N.; Perumalla Venkata, R.; Grover, P. Genotoxicity, biochemical, and biodistribution studies of magnesium oxide nano and microparticles in albino wistar rats after 28-day repeated oral exposure. Environ. Toxicol. 2018, 33, 396–410. [Google Scholar] [CrossRef] [PubMed]
- Warsy, A.S.; Almukaynizi, F.B.; AlDaihan, S.; Alam, S.; Daghastani, M. Genetic Polymorphisms in Aromatase (CYP19) Gene and Cancer. Genet. Polymorph. 2017. [Google Scholar]
- Aghaie, F.; Khazali, H.; Hedayati, M.; Akbarnejad, A. The effects of exercise on expression of CYP19 and StAR mRNA in steroid-induced polycystic ovaries of female rats. Int. J. Fertil. Steril. 2018, 11, 298–303. [Google Scholar] [PubMed]
- Lee, Y.H.; Yang, H.; Lee, S.R.; Kwon, S.W.; Hong, E.J.; Lee, H.W. Welsh onion root (Allium fistulosum) restores ovarian functions from letrozole induced-polycystic ovary syndrome. Nutrients 2018, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Panghiyangani, R.; Soeharso, P.; Pujianto, A.; Suryandari, D.; Wiweko, B.; Kurniati, M.; Pujianto, D. CYP19A1 gene expression in patients with polycystic ovarian syndrome. J. Hum. Reprod. Sci. 2020, 13, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Suriyakalaa, U.; Ramachandran, R.; Doulathunnisa, J.A.; Aseervatham, S.B.; Sankarganesh, D.; Kamalakkannan, S.; Kadalmani, B.; Angayarkanni, J.; Akbarsha, M.A.; Achiraman, S. Upregulation of Cyp19a1 and PPAR-γ in ovarian steroidogenic pathway by Ficus religiosa: A potential cure for polycystic ovary syndrome. J. Ethnopharmacol. 2021, 267, 113540. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ruan, Y.-C.; Yang, Y.; Wang, K.; Liang, S.; Han, Y.; Teng, X.-M.; Yang, J.-Z. Follicular hyperandrogenism downregulates aromatase in luteinized granulosa cells in PCOS women. Reproduction. 2015, 150, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Li, W.; Zhang, G. The protective effect of resveratrol against cytotoxicity induced by mycotoxin, zearalenone. Food Funct. 2016, 7, 3703–3715. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Heuck, C.J.; Fazzari, M.J.; Mehta, J.; Singhal, S.; Greally, J.M.; Verma, A. DNA methylation alterations in multiple myeloma as a model for epigenetic changes in cancer. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.Y.; Sun, C.X.; Liu, Y.K.; Li, Y.; Wang, L.; Zhang, W. Promoter methylation of CYP19A1 gene in chinese polycystic ovary syndrome patients. Gynecol. Obstet. Investig. 2013, 76, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Song, B.; Wu, J.; Zhang, Y.; Chen, A.; Shao, L. Potential adverse effects of nanoparticles on the reproductive system. Int. J. Nanomed. 2018, 13, 8487–8506. [Google Scholar] [CrossRef] [PubMed]
- Anway, M.D.; Cupp, A.S.; Uzumcu, N.; Skinner, M.K. Toxicology: Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005, 308, 1466–1469. [Google Scholar] [CrossRef]
- Li, N.; Liu, X.L.; Zhang, F.L.; Tian, Y.; Zhu, M.; Meng, L.Y.; Dyce, P.W.; Shen, W.; Li, L. Whole-transcriptome analysis of the toxic effects of zearalenone exposure on ceRNA networks in porcine granulosa cells. Environ. Pollut. 2020, 261, 114007. [Google Scholar] [CrossRef] [PubMed]
- Azouz, A.A.; Ali, S.E.; Abd-Elsalam, R.M.; Emam, S.R.; Galal, M.K.; Elmosalamy, S.H.; Alsherbiny, M.A.; Hassan, B.B.; Li, C.G.; El Badawy, S.A. Modulation of steroidogenesis by Actaea racemosa and vitamin C combination, in letrozole induced polycystic ovarian syndrome rat model: Promising activity without the risk of hepatic adverse effect. Chin. Med. 2021, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Güney, M.; Demirin, H.; Oral, B.; Özgüner, M.; Bayhan, G.; Altuntas, I. Ovarian toxicity in rats caused by methidathion and ameliorating effect of vitamins E and C. Hum. Exp. Toxicol. 2007, 26, 491–498. [Google Scholar] [CrossRef]
- Ahmad, B.; Shrivastava, V.K.; Saleh, R.; Henkel, R.; Agarwal, A. Protective effects of saffron against zearalenoneinduced alterations in reproductive hormones in female mice (Mus musculus). Clin. Exp. Reprod. Med. 2018, 45, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Mangalampalli, B.; Dumala, N.; Grover, P. Acute oral toxicity study of magnesium oxide nanoparticles and microparticles in female albino Wistar rats. Regul. Toxicol. Pharmacol. 2017, 90, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Sudhabose, S.; Sooryakanth, B.; Rajan, M.R. Acute Toxicity, Hematological Profile, and Histopathological Effects of MgO Nanoparticles on Gills, Muscle, Liver of Mrigal, Cirrhinus mrigala. Biol. Trace Elem. Res. 2023, 202, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Dumala, N.; Mangalampalli, B.; Chinde, S.; Kumari, S.I.; Mahoob, M.; Rahman, M.F.; Grover, P. Genotoxicity study of nickel oxide nanoparticles in female Wistar rats after acute oral exposure. Mutagenesis 2017, 32, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Reddy, U.A.; Prabhakar, P.V.; Mahboob, M. Biomarkers of oxidative stress for in vivo assessment of toxicological effects of iron oxide nanoparticles. Saudi J. Biol. Sci. 2017, 24, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Kim, B.; Shin, H.S.; Kwon, H.J.; Park, E.S. The protective effect of hispidin against hydrogen peroxide-induced apoptosis in H9c2 cardiomyoblast cells through Akt/GSK-3β and ERK1/2 signaling pathway. Exp. Cell Res. 2014, 327, 264–275. [Google Scholar] [CrossRef]
- Lai, M.C.; Liu, W.Y.; Liou, S.S.; Liu, I.M. Hispidin in the Medicinal Fungus Protects Dopaminergic Neurons from JNK Activation-Regulated Mitochondrial-Dependent Apoptosis in an MPP+-Induced In Vitro Model of Parkinson’s Disease. Nutrients 2023, 15, 549. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alenazi, A.; Virk, P.; Almoqhem, R.; Alsharidah, A.; Al-Ghadi, M.Q.; Aljabr, W.; Alasmari, F.; Albasher, G. The Efficacy of Hispidin and Magnesium Nanoparticles against Zearalenone-Induced Fungal Toxicity Causing Polycystic Ovarian Syndrome in Rats. Biomedicines 2024, 12, 943. https://doi.org/10.3390/biomedicines12050943
Alenazi A, Virk P, Almoqhem R, Alsharidah A, Al-Ghadi MQ, Aljabr W, Alasmari F, Albasher G. The Efficacy of Hispidin and Magnesium Nanoparticles against Zearalenone-Induced Fungal Toxicity Causing Polycystic Ovarian Syndrome in Rats. Biomedicines. 2024; 12(5):943. https://doi.org/10.3390/biomedicines12050943
Chicago/Turabian StyleAlenazi, Amenah, Promy Virk, Reem Almoqhem, Amani Alsharidah, Muath Q. Al-Ghadi, Waleed Aljabr, Fawaz Alasmari, and Gadah Albasher. 2024. "The Efficacy of Hispidin and Magnesium Nanoparticles against Zearalenone-Induced Fungal Toxicity Causing Polycystic Ovarian Syndrome in Rats" Biomedicines 12, no. 5: 943. https://doi.org/10.3390/biomedicines12050943