
Effects of Deuterium Depletion on Age-Declining Thymopoiesis In Vivo
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Thymic Lymphocytes Culture Preparation
2.3. Flow Cytometry
2.4. Ethical Approval
2.5. Statistical Analysis
3. Results
3.1. Body Weight Changes during the Experiment
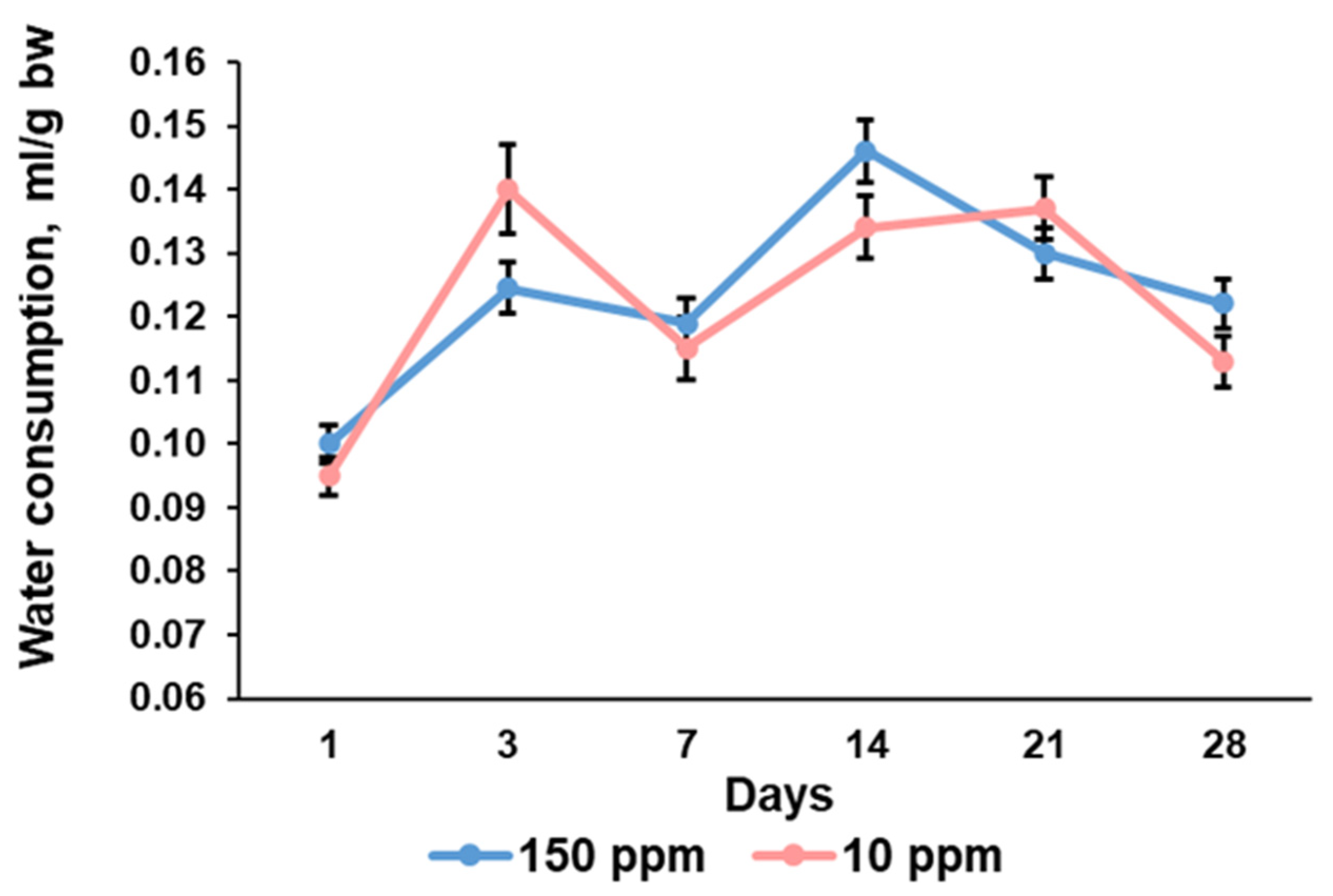
3.2. Parameters of Water Consumption
3.3. Thymus Weight
3.4. Evaluation of Thymic Lymphocyte Subpopulations
3.4.1. CD4−CD8− Progenitors
3.4.2. CD4+CD8+ Lymphocytes
3.4.3. CD3+ Lymphocytes
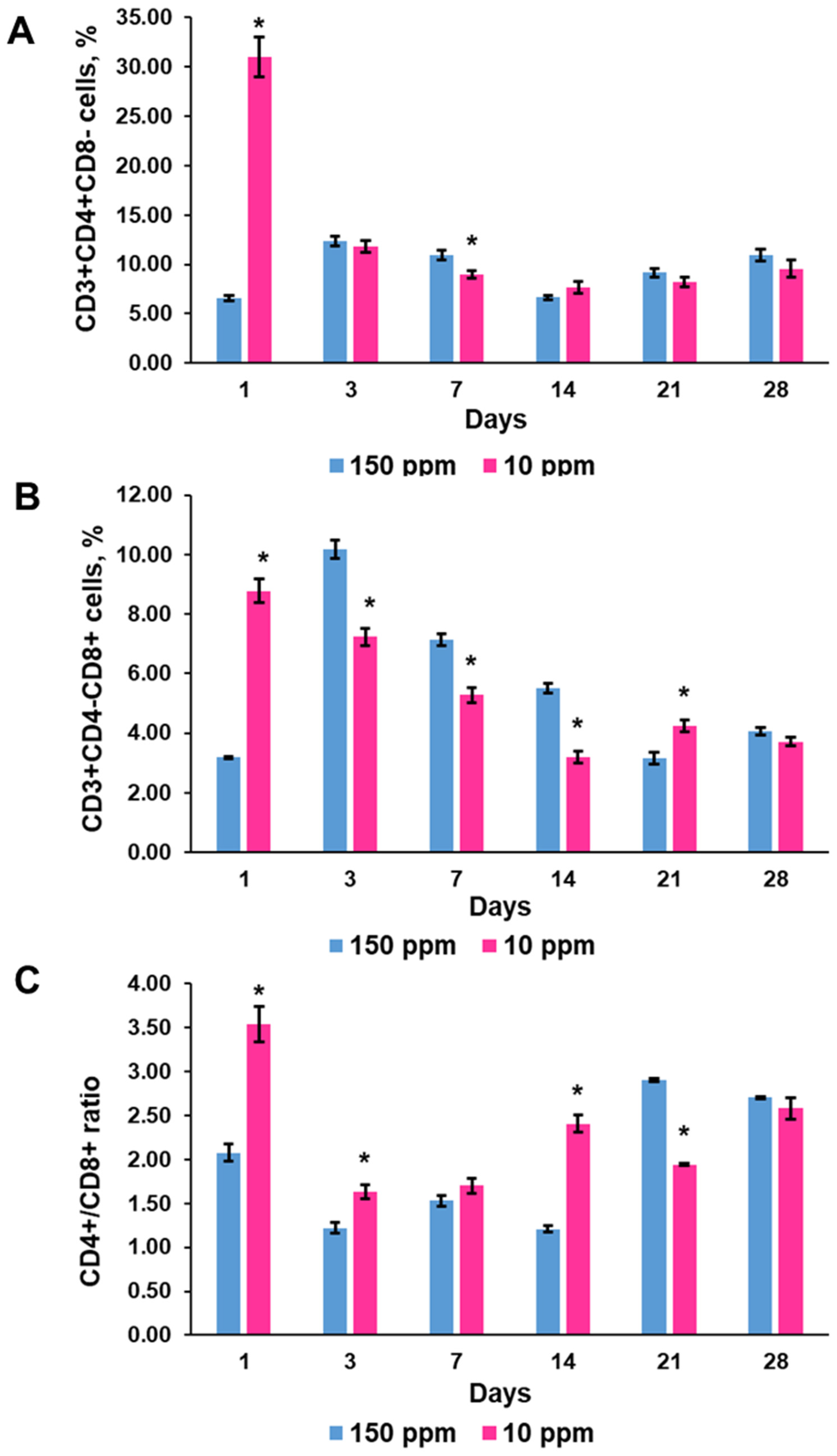
3.4.4. T-Helper CD3+CD4+ and T-Cytotoxic CD3+CD8+ Lymphocytes
3.5. T Cell Efflux to the Blood
4. Discussion
5. Conclusions
Limitations of the Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boursalian, T.E.; Golob, J.; Soper, D.M.; Cooper, C.J.; Fink, P.J. Continued maturation of thymic emigrants in the periphery. Nat. Immunol. 2004, 5, 418–425. [Google Scholar] [CrossRef]
- Martins, V.C.; Ruggiero, E.; Schlenner, S.M.; Madan, V.; Schmidt, M.; Fink, P.J.; von Kalle, C.; Rodewald, H.R. Thymus-autonomous T cell development in the absence of progenitor import. J. Exp. Med. 2012, 209, 1409–1417. [Google Scholar] [CrossRef]
- Min, H.; Montecino-Rodriguez, E.; Dorshkind, K. Reduction in the Developmental Potential of Intrathymic T Cell Progenitors with Age. J. Immunol. 2004, 173, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.S.; Boursalian, T.E.; Turk, G.L.; Fink, P.J. Thymic output in aged mice. Proc. Natl. Acad. Sci. USA 2006, 103, 8447–8852. [Google Scholar] [CrossRef]
- Palathumpat, V.C.; Vandeputte, M.M.; Waer, M. Effects of thymus irradiation on the immune competence of T cells after total-lymphoid irradiation. Transplantation 1990, 50, 95–100. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Timokhina, E.P.; Yaglov, V.V. Effects of low-dose dichlorodiphenyltrichloroethane on the morphology and function of rat thymus. Bull. Exp. Biol. Med. 2013, 155, 701–704. [Google Scholar] [CrossRef]
- Obernikhin, S.S.; Yaglova, N.V. Morphological and functional changes in the thymus and spleen of mouse offspring in the development of systemic inflammatory response after a single immunity stimulation in early pregnancy. Bull. Exp. Biol. Med. 2014, 157, 812–815. [Google Scholar] [CrossRef]
- Schaue, D.A. Century of Radiation Therapy and Adaptive Immunity. Front. Immunol. 2017, 8, 431. [Google Scholar] [CrossRef]
- Duah, M.; Li, L.; Sen, J.; Lan, Q.; Pan, B.; Xu, K. Thymus Degeneration and Regeneration. Front. Immunol. 2021, 12, 706244. [Google Scholar] [CrossRef]
- Thomas, R.; Wang, W.; Su, D.M. Contributions of Age-Related Thymic Involution to Immunosenescence and Inflammaging. Immun. Ageing 2020, 17, 2. [Google Scholar] [CrossRef]
- Kinsella, S.; Dudakov, J.A. When the Damage Is Done: Injury and Repair in Thymus Function. Front. Immunol. 2020, 11, 1745. [Google Scholar] [CrossRef] [PubMed]
- Sunaoshi, M.; Blyth, B.J.; Shang, Y.; Tsuruoka, C.; Morioka, T.; Shinagawa, M.; Ogawa, M.; Shimada, Y.; Tachibana, A.; Iizuka, D.; et al. Post-Irradiation Thymic Regeneration in B6C3F1 Mice Is Age Dependent and Modulated by Activation of the PI3K-AKT-mTOR Pathway. Biology 2022, 11, 449. [Google Scholar] [CrossRef] [PubMed]
- Kreins, A.Y.; Graham Davies, E. Replacing Defective Thymus Function. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Gaudeaux, P.; Moirangthem, R.D.; Bauquet, A.; Simons, L.; Joshi, A.; Cavazzana, M.; Negre, O.; Soheili, S.; Andre, I. T-Cell Progenitors as a new immunotherapy to bypass hurdles of allogeneic hematopoietic stem cell transplantation. Front. Immunol. 2022, 13, 956919. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.S.; Spyroglou, L.; Muirhead, J.L.; Heng, T.S.; Prieto-Hinojosa, A.; Prince, H.M.; Chidgey, A.P.; Schwarer, A.P.; Boyd, R.L. Enhanced immune system regeneration in humans following allogeneic or autologous hemopoietic stem cell transplantation by temporary sex steroid blockade. Clin. Cancer Res. 2008, 14, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Bodart, G.; Farhat, K.; Renard-Charlet, C.; Becker, G.; Plenevaux, A.; Salvatori, R.; Geenen, V.; Martens, H. The severe deficiency of the somatotrope GH-releasing hormone/Growth Hormone/Insulin-Like Growth Factor 1 axis of Ghrh-/- mice is associated with an important splenic atrophy and relative B lymphopenia. Front. Endocrinol. 2018, 9, 296. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Duah, M.; Xu, Y.; Liang, Y.; Wang, D.; Xia, F.; Li, L.; Sun, Z.; Yan, Z.; Xu, K.; et al. Dynamic of plasma IL-22 level is an indicator of thymic output after allogeneic hematopoietic cell transplantation. Life Sci. 2021, 265, 118849. [Google Scholar] [CrossRef] [PubMed]
- Kselíková, V.; Vítová, M.; Bišová, K. Deuterium and its impact on living organisms. Folia Microbiol. 2019, 64, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Yaglova, N.V.; Timokhina, E.P.; Obernikhin, S.S.; Yaglov, V.V. Emerging Role of Deuterium/Protium Disbalance in Cell Cycle and Apoptosis. Int. J. Mol. Sci. 2023, 24, 3107. [Google Scholar] [CrossRef] [PubMed]
- Jancso, G. Isotope effects. In Handbook of Nuclear Chemistry; Vértes, A., Nagy, S., Klencsar, Z., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2013; Volume 2, pp. 85–116. [Google Scholar]
- Somlyai, G.; Javaheri, B.; Davari, H.; Gyöngyi, Z.; Somlyai, I.; Tamaddon, K.A.; Boros, L.G. Pre-Clinical and Clinical Data Confirm the Anticancer Effect of Deuterium Depletion. Biomacromol. J. 2016, 2, 1–7. [Google Scholar] [CrossRef]
- Yaglova, N.V.; Obernikhin, S.S.; Yaglov, V.V.; Nazimova, S.V. Time-dependent effect of deuterium depletion on tumor growth and metastasis. Russ. Open Med. J. 2020, 9, e0210. [Google Scholar] [CrossRef]
- Boros, L.G.; Somlyai, I.; Kovács, B.Z.; Puskás, L.G.; Nagy, L.I.; Dux, L.; Farkas, G.; Somlyai, G. Deuterium Depletion Inhibits Cell Proliferation, RNA and Nuclear Membrane Turnover to Enhance Survival in Pancreatic Cancer. Cancer Control 2021, 28, 1073274821999655. [Google Scholar] [CrossRef] [PubMed]
- Gyöngyi, Z.; Budán, F.; Szabó, I.; Ember, I.; Kiss, I.; Krempels, K.; Somlyai, I.; Somlyai, G. Deuterium depleted water effects on survival of lung cancer patients and expression of Kras, Bcl2, and Myc genes in mouse lung. Nutr. Cancer. 2013, 65, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gaetani, M.; Chernobrovkin, A.; Zubarev, R.A. Anticancer Effect of Deuterium Depleted Water—Redox Disbalance Leads to Oxidative Stress. Mol. Cell. Proteom. 2019, 18, 2373–2387. [Google Scholar] [CrossRef] [PubMed]
- Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Yaglov, V.V. Response of Pituitary-Thyroid Axis to a Short-Term Shift in Deuterium Content in the Body. Bull. Exp. Biol. Med. 2021, 171, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Nazimova, S.V.; Yaglov, V.V. Reactive Alterations in Thymic Lymphocytopoiesis to Short-Term Decrease in Deuterium Content in the Body. Bull. Exp. Biol. Med. 2022, 173, 494–496. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and negative selection of the T cell repertoire: What thymocytes see (and don’t see). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Ohigashi, I.; Takahama, Y. Thymus machinery for T-cell selection. Int. Immunol. 2019, 31, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Diatroptov, M.E.; Kondrashevskaia, M.V.; Makarova, O.V.; Obernikhin, S.S. Infradian rhythms of the thymic morphofunctional state in male Wistar rats. Ross. Fiziol. Zhurnal Im. IM Sechenova 2013, 99, 729–736. (In Russian) [Google Scholar]
- Dzhimak, S.S.; Basov, A.A.; Baryshev, M.G. Content of deuterium in biological fluids and organs: Influence of deuterium depleted water on D/H gradient and the process of adaptation. Dokl. Biochem. Biophys. 2015, 465, 370–373. [Google Scholar] [CrossRef]
- Kosenkov, A.V.; Lobyshev, V.I.; Gulyaev, M.V.; Yusubalieva, G.M.; Baklaushev, V.P. The reversible effect of deuteration on tissue fluid and biopolymers in normal and tumor tissues of mice. Biophysics 2018, 63, 820–824. [Google Scholar] [CrossRef]
- Malcolm, T.I.; Hodson, D.J.; Macintyre, E.A.; Turner, S.D. Challenging perspectives on the cellular origins of lymphoma. Open Biol. 2016, 6, 160232. [Google Scholar] [CrossRef]
- Kravtsov, A.; Kozin, S.; Basov, A.; Butina, E.; Baryshev, M.; Malyshko, V.; Moiseev, A.; Elkina, A.; Dzhimak, S. Reduction of deuterium level supports resistance of neurons to glucose deprivation and hypoxia: Study in cultures of neurons and on animals. Molecules 2022, 27, 243. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, J.; Hennig, D. Breather solutions of a nonlinear DNA model including a longitudinal degree of freedom. Phys. A Stat. Mech. Appl. 2003, 323, 519–533. [Google Scholar] [CrossRef]
- Sulaiman, A.; Zen, F.P.; Alatas, H.; Handoko, L.T. Dynamics of DNA breathing in the Peyrard–Bishop model with damping and external force. Phys. D Nonlinear Phenom. 2012, 241, 1640–1647. [Google Scholar] [CrossRef]
- Nikitiuk, A.S.; Bayandin, Y.V.; Naimark, O.B. Statistical thermodynamics of DNA with open states. Phys. A Stat. Mech. Appl. 2022, 607, 128156. [Google Scholar] [CrossRef]
- Dzhimak, S.S.; Drobotenko, M.I.; Basov, A.A.; Svidlov, A.A.; Fedulova, L.V.; Lyasota, O.M.; Baryshev, M.G. Mathematical Modeling of Open State in DNA Molecule Depending on the Deuterium Concentration in the Surrounding Liquid Media at Different Values of Hydrogen Bond Disruption Energy. Dokl. Biochem. Biophys. 2018, 483, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Basov, A.; Drobotenko, M.; Svidlov, A.; Gerasimenko, E.; Malyshko, V.; Elkina, A.; Baryshev, M.; Dzhimak, S. Inequality in the frequency of the open states occurrence depends on single 2H/1H replacement in DNA. Molecules 2020, 25, 3753. [Google Scholar] [CrossRef] [PubMed]
- Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Yaglov, V.V.; Tsomartova, D.A.; Nazimova, S.V.; Tsomartova, E.S.; Ivanova, M.Y.; Chereshneva, E.V.; Lomanovskaya, T.A. Bilateral Shifts in Deuterium Supply Similarly Change Physiology of the Pituitary–Thyroid Axis, but Differentially Influence Na+/I- Symporter Production. Int. J. Mol. Sci. 2023, 24, 6803. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Verdon, D.J.; Mulazzani, M.; Jenkins, M.R. Cellular and Molecular Mechanisms of CD8+ T Cell Differentiation, Dysfunction and Exhaustion. Int. J. Mol. Sci. 2020, 21, 7357. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Ha, S.-J.; Kaech, S.M.; Haining, W.N.; Sarkar, S.; Kalia, V.; Subramaniam, S.; Blattman, J.N.; Barber, D.L.; Ahmed, R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity 2007, 27, 670–676. [Google Scholar] [CrossRef] [PubMed]
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. CD8 T Cell Exhaustion During Chronic Viral Infection and Cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaglova, N.V.; Obernikhin, S.S.; Timokhina, E.P.; Tsomartova, D.A.; Yaglov, V.V.; Nazimova, S.V.; Tsomartova, E.S.; Ivanova, M.Y.; Chereshneva, E.V.; Lomanovskaya, T.A. Effects of Deuterium Depletion on Age-Declining Thymopoiesis In Vivo. Biomedicines 2024, 12, 956. https://doi.org/10.3390/biomedicines12050956
Yaglova NV, Obernikhin SS, Timokhina EP, Tsomartova DA, Yaglov VV, Nazimova SV, Tsomartova ES, Ivanova MY, Chereshneva EV, Lomanovskaya TA. Effects of Deuterium Depletion on Age-Declining Thymopoiesis In Vivo. Biomedicines. 2024; 12(5):956. https://doi.org/10.3390/biomedicines12050956
Chicago/Turabian StyleYaglova, Nataliya V., Sergey S. Obernikhin, Ekaterina P. Timokhina, Dibakhan A. Tsomartova, Valentin V. Yaglov, Svetlana V. Nazimova, Elina S. Tsomartova, Marina Y. Ivanova, Elizaveta V. Chereshneva, and Tatiana A. Lomanovskaya. 2024. "Effects of Deuterium Depletion on Age-Declining Thymopoiesis In Vivo" Biomedicines 12, no. 5: 956. https://doi.org/10.3390/biomedicines12050956