Progress in Adenoviral Capsid-Display Vaccines

Abstract
:
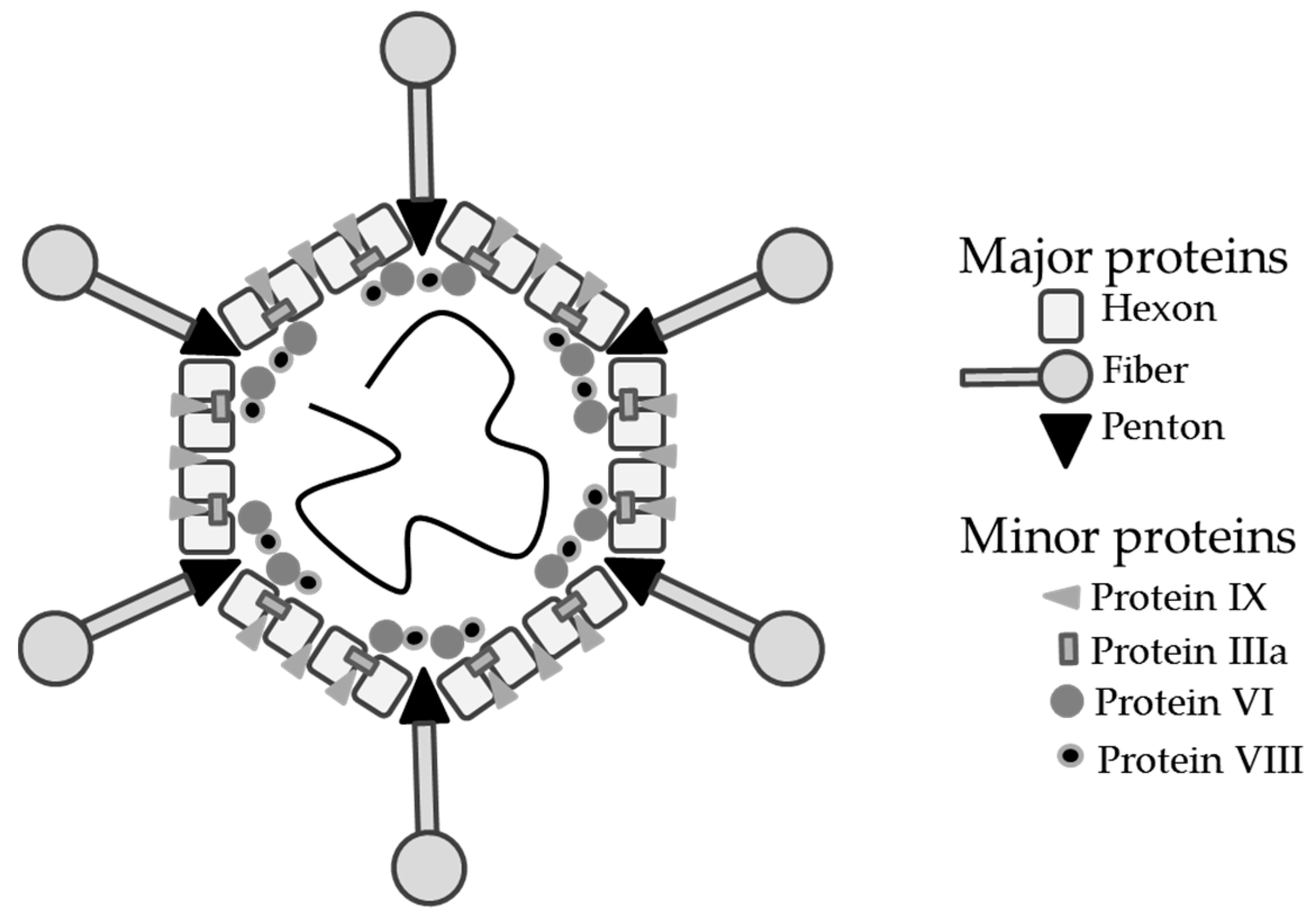
1. Introduction
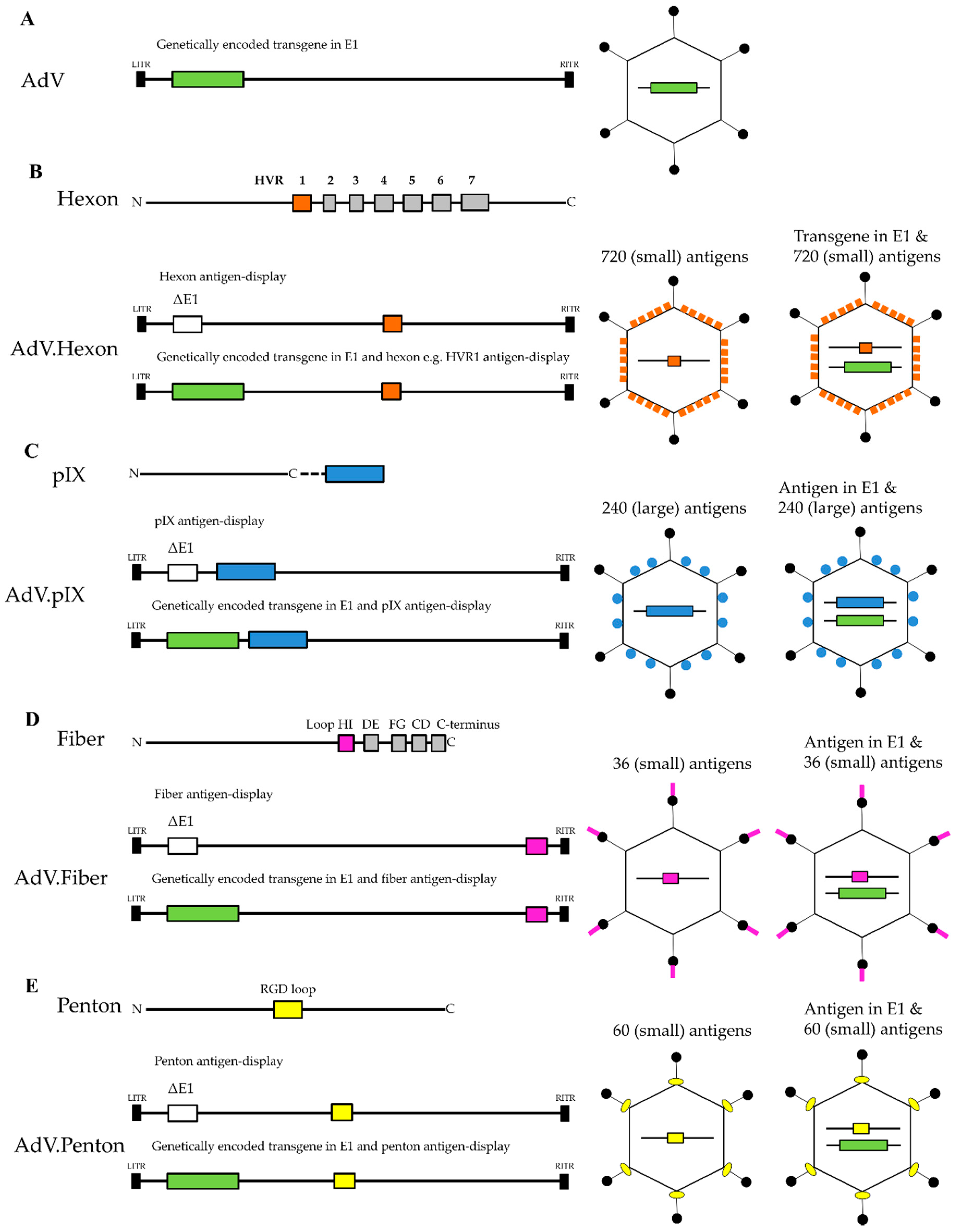
2. Hexon Antigen Display
4. Fiber- and Penton-Display Vectors
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegrist, C. Vaccine immunology. In Vaccines. Saunders Elsevier, 5th ed.; Plotkin, S., Orenstein, W., Offit, P., Eds.; Saunders/Elsevier: Philadelphia, PA, USA, 2008; pp. 17–36. [Google Scholar]
- Rodriguez, A.; Mintardjo, R.; Tax, D.; Gillissen, G.; Custers, J.; Pau, M.G.; Klap, J.; Santra, S.; Balachandran, H.; Letvin, N.L.; et al. Evaluation of a prime-boost vaccine schedule with distinct adenovirus vectors against malaria in rhesus monkeys. Vaccine 2009, 27, 6226–6233. [Google Scholar] [CrossRef] [PubMed]
- Capone, S.; Meola, A.; Ercole, B.B.; Vitelli, A.; Pezzanera, M.; Ruggeri, L.; Davies, M.E.; Tafi, R.; Santini, C.; Luzzago, A.; et al. A novel adenovirus type 6 (Ad6)-based hepatitis C virus vector that overcomes preexisting anti-ad5 immunity and induces potent and broad cellular immune responses in rhesus macaques. J. Virol. 2006, 80, 1688–1699. [Google Scholar] [CrossRef] [PubMed]
- Folgori, A.; Capone, S.; Ruggeri, L.; Meola, A.; Sporeno, E.; Ercole, B.B.; Pezzanera, M.; Tafi, R.; Arcuri, M.; Fattori, E.; et al. A T-cell HCV vaccine eliciting effective immunity against heterologous virus challenge in chimpanzees. Nat. Med. 2006, 12, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Teigler, J.E.; Iampietro, M.J.; Barouch, D.H. Vaccination with adenovirus serotypes 35, 26, and 48 elicits higher levels of innate cytokine responses than adenovirus serotype 5 in rhesus monkeys. J. Virol. 2012, 86, 9590–9598. [Google Scholar] [CrossRef] [PubMed]
- Zahn, R.; Gillisen, G.; Roos, A.; Koning, M.; van der Helm, E.; Spek, D.; Weijtens, M.; Pau, M.G.; Radosevic, K.; Weverling, G.J.; et al. Ad35 and ad26 vaccine vectors induce potent and cross-reactive antibody and T-cell responses to multiple filovirus species. PLoS ONE 2012, 7, e44115. [Google Scholar] [CrossRef] [PubMed]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Fallaux, F.J.; Kranenburg, O.; Cramer, S.J.; Houweling, A.; van Ormondt, H.; Hoeben, R.C.; van der Eb, A.J. Characterization of 911: A new helper cell line for the titration and propagation of early region 1-deleted adenoviral vectors. Hum. Gene Ther. 1996, 7, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Fallaux, F.J.; Bout, A.; van der Velde, I.; van den Wollenberg, D.J.; Hehir, K.M.; Keegan, J.; Auger, C.; Cramer, S.J.; van Ormondt, H.; van der Eb, A.J.; et al. New helper cells and matched early region 1-deleted adenovirus vectors prevent generation of replication-competent adenoviruses. Hum. Gene Ther. 1998, 9, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, J.; Smith, J.P.; Lipiec, A.; Majhen, D.; Lemckert, A.; van Ooij, M.; Ives, P.; Yallop, C.; Custers, J.; Havenga, M. Challenges in manufacturing adenoviral vectors for global vaccine product deployment. Hum. Gene Ther. 2014, 25, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Havenga, M.J.; Lemckert, A.A.; Ophorst, O.J.; van Meijer, M.; Germeraad, W.T.; Grimbergen, J.; van den Doel, M.A.; Vogels, R.; van Deutekom, J.; Janson, A.A.; et al. Exploiting the natural diversity in adenovirus tropism for therapy and prevention of disease. J. Virol. 2002, 76, 4612–4620. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; Walsh, S.R.; Seaman, M.S.; Tucker, R.P.; Krause, K.H.; Patel, A.; Johnson, J.A.; Kleinjan, J.; Yanosick, K.E.; Perry, J.; et al. First-in-Human Evaluation of the Safety and Immunogenicity of a Recombinant Adenovirus Serotype 26 HIV-1 Env Vaccine (IPCAVD 001). J. Infect. Dis. 2013, 207, 240–247. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, G.A.; Duncan, C.J.; Ewer, K.J.; Collins, K.A.; Elias, S.C.; Halstead, F.D.; Goodman, A.L.; Edwards, N.J.; Reyes-Sandoval, A.; Bird, P.; et al. Clinical assessment of a recombinant simian adenovirus ChAd63: A potent new vaccine vector. J. Infect. Dis. 2012, 205, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, A.; Tiono, A.B.; Kargougou, D.; Yaro, J.B.; Ouedraogo, E.; Kabore, Y.; Kangoye, D.; Bougouma, E.C.; Gansane, A.; Henri, N.; et al. A phase 1b randomized, controlled, double-blinded dosage-escalation trial to evaluate the safety, reactogenicity and immunogenicity of an adenovirus type 35 based circumsporozoite malaria vaccine in Burkinabe healthy adults 18 to 45 years of age. PLoS ONE 2013, 8, e78679. [Google Scholar] [CrossRef] [PubMed]
- De Santis, O.; Audran, R.; Pothin, E.; Warpelin-Decrausaz, L.; Vallotton, L.; Wuerzner, G.; Cochet, C.; Estoppey, D.; Steiner-Monard, V.; Lonchampt, S.; et al. Safety and immunogenicity of a chimpanzee adenovirus-vectored Ebola vaccine in healthy adults: A randomised, double-blind, placebo-controlled, dose-finding, phase 1/2a study. Lancet Infect. Dis. 2016, 16, 311–320. [Google Scholar] [CrossRef]
- White, M.T.; Bejon, P.; Olotu, A.; Griffin, J.T.; Riley, E.M.; Kester, K.E.; Ockenhouse, C.F.; Ghani, A.C. The relationship between RTS,S vaccine-induced antibodies, CD4(+) T cell responses and protection against Plasmodium falciparum infection. PLoS ONE 2013, 8, e61395. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Ockenhouse, C.F.; Regules, J.A.; Dutta, S.; Wallqvist, A.; Jongert, E.; Waters, N.C.; Lemiale, F.; Bergmann-Leitner, E. The biological function of antibodies induced by the RTS,S/AS01 malaria vaccine candidate is determined by their fine specificity. Malar. J. 2016, 15, 301. [Google Scholar] [CrossRef] [PubMed]
- Handisurya, A.; Schellenbacher, C.; Haitel, A.; Senger, T.; Kirnbauer, R. Human papillomavirus vaccination induces neutralising antibodies in oral mucosal fluids. Br. J. Cancer 2016, 114, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, M.G.; Bruden, D.; Hurlburt, D.; Zanis, C.; Thompson, G.; Rea, L.; Toomey, M.; Townshend-Bulson, L.; Rudolph, K.; Bulkow, L.; et al. Antibody Levels and Protection After Hepatitis B Vaccine: Results of a 30-Year Follow-up Study and Response to a Booster Dose. J. Infect. Dis. 2016, 214, 16–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palladini, A.; Thrane, S.; Janitzek, C.M.; Pihl, J.; Clemmensen, S.B.; de Jongh, W.A.; Clausen, T.M.; Nicoletti, G.; Landuzzi, L.; Penichet, M.L.; et al. Virus-like particle display of HER2 induces potent anti-cancer responses. Oncoimmunology 2018, 7, e1408749. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Liu, J.; Peter, L.; Abbink, P.; Iampietro, M.J.; Cheung, A.; Alter, G.; Chung, A.; Dugast, A.S.; Frahm, N.; et al. Characterization of humoral and cellular immune responses elicited by a recombinant adenovirus serotype 26 HIV-1 Env vaccine in healthy adults (IPCAVD 001). J. Infect. Dis. 2013, 207, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Lu, S. Heterologous prime-boost vaccination. Curr. Opin. Immunol. 2009, 21, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Alter, G.; Broge, T.; Linde, C.; Ackerman, M.E.; Brown, E.P.; Borducchi, E.N.; Smith, K.M.; Nkolola, J.P.; Liu, J.; et al. Protective efficacy of adenovirus/protein vaccines against SIV challenges in rhesus monkeys. Science 2015, 349, 320–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, V.A.; McGrath, S.M.; Dubois, P.M.; Pau, M.G.; Mettens, P.; Shott, J.; Cobb, M.; Burge, J.R.; Larson, D.; Ware, L.A.; et al. Priming with an adenovirus 35-circumsporozoite protein (CS) vaccine followed by RTS,S/AS01B boosting significantly improves immunogenicity to Plasmodium falciparum CS compared to that with either malaria vaccine alone. Infect. Immun. 2007, 75, 2283–2290. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, A.M.; Naddeo, M.; Capone, S.; Ammendola, V.; Hu, K.; Meredith, L.; Verhoye, L.; Rychlowska, M.; Rappuoli, R.; Ulmer, J.B.; et al. Combined adenovirus vector and hepatitis C virus envelope protein prime-boost regimen elicits T cell and neutralizing antibody immune responses. J. Virol. 2014, 88, 5502–5510. [Google Scholar] [CrossRef] [PubMed]
- Keele, B.F.; Li, W.; Borducchi, E.N.; Nkolola, J.P.; Abbink, P.; Chen, B.; Seaman, M.S.; Barouch, D.H. Adenovirus prime, Env protein boost vaccine protects against neutralization-resistant SIVsmE660 variants in rhesus monkeys. Nat. Commun. 2017, 8, 15740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, V.S.; Natchiar, S.K.; Stewart, P.L.; Nemerow, G.R. Crystal structure of human adenovirus at 3.5 A resolution. Science 2010, 329, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.S.; Natchiar, S.K.; Gritton, L.; Mullen, T.M.; Stewart, P.L.; Nemerow, G.R. Crystallization and preliminary X-ray diffraction analysis of human adenovirus. Virology 2010, 402, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jin, L.; Koh, S.B.; Atanasov, I.; Schein, S.; Wu, L.; Zhou, Z.H. Atomic structure of human adenovirus by cryo-EM reveals interactions among protein networks. Science 2010, 329, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, J.; van der Heijdt, S.; Hoeben, R.C. The adenovirus capsid: Major progress in minor proteins. J. Gen. Virol. 2005, 86 Pt 6, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wu, L.; Sun, R.; Zhou, Z.H. Atomic Structures of Minor Proteins VI and VII in the Human Adenovirus. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.R.; Maxfield, L.F.; Lynch, D.M.; Iampietro, M.J.; Borducchi, E.N.; Barouch, D.H. Adenovirus serotype 5-specific neutralizing antibodies target multiple hexon hypervariable regions. J. Virol. 2012, 86, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Ahi, Y.S.; Bangari, D.S.; Mittal, S.K. Adenoviral vector immunity: Its implications and circumvention strategies. Curr. Gene Ther. 2011, 11, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, Q.; Ertl, H.C.; Wilson, J.M. Cellular and humoral immune responses to viral antigens create barriers to lung-directed gene therapy with recombinant adenoviruses. J. Virol. 1995, 69, 2004–2015. [Google Scholar] [PubMed]
- Ledgerwood, J.E.; Costner, P.; Desai, N.; Holman, L.; Enama, M.E.; Yamshchikov, G.; Mulangu, S.; Hu, Z.; Andrews, C.A.; Sheets, R.A.; et al. A replication defective recombinant Ad5 vaccine expressing Ebola virus GP is safe and immunogenic in healthy adults. Vaccine 2010, 29, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Fausther-Bovendo, H.; Kobinger, G.P. Pre-existing immunity against Ad vectors: Humoral, cellular, and innate response, what’s important? Hum. Vaccines Immunother. 2014, 10, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.G.; Crystal, R.G.; Falck-Pedersen, E. Construction and characterization of hexon-chimeric adenoviruses: Specification of adenovirus serotype. J. Virol. 1998, 72, 10260–10264. [Google Scholar] [PubMed]
- Roy, S.; Shirley, P.S.; McClelland, A.; Kaleko, M. Circumvention of immunity to the adenovirus major coat protein hexon. J. Virol. 1998, 72, 6875–6879. [Google Scholar] [PubMed]
- Wu, H.; Han, T.; Belousova, N.; Krasnykh, V.; Kashentseva, E.; Dmitriev, I.; Kataram, M.; Mahasreshti, P.J.; Curiel, T.D. Identification of sites in adenovirus hexon for foreign peptide incorporation. J. Virol. 2005, 79, 3382–3390. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.M.; Nanda, A.; Havenga, M.J.; Abbink, P.; Lynch, D.M.; Ewald, B.A.; Liu, J.; Thorner, A.R.; Swanson, P.E.; Gorgone, D.A.; et al. Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 2006, 441, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; Walsh, S.R.; Seaman, M.S.; Johnson, J.A.; Tucker, R.P.; Kleinjan, J.A.; Gothing, J.A.; Engelson, B.A.; Carey, B.R.; Oza, A.; et al. First-in-human evaluation of a hexon chimeric adenovirus vector expressing HIV-1 Env (IPCAVD 002). J. Infect. Dis. 2014, 210, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Arnberg, N. Adenovirus receptors: Implications for targeting of viral vectors. Trends Pharmacol. Sci. 2012, 33, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Nicol, C.G.; Graham, D.; Miller, W.H.; White, S.J.; Smith, T.A.; Nicklin, S.A.; Stevenson, S.C.; Baker, A.H. Effect of adenovirus serotype 5 fiber and penton modifications on in vivo tropism in rats. Mol. Ther. 2004, 10, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Beatty, M.S.; Curiel, D.T. Chapter two—Adenovirus strategies for tissue-specific targeting. Adv. Cancer Res. 2012, 115, 39–67. [Google Scholar] [PubMed]
- Hogg, R.T.; Thorpe, P.; Gerard, R.D. Retargeting adenoviral vectors to improve gene transfer into tumors. Cancer Gene Ther. 2011, 18, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Furcinitti, P.S.; van Oostrum, J.; Burnett, R.M. Adenovirus polypeptide IX revealed as capsid cement by difference images from electron microscopy and crystallography. EMBO J. 1989, 8, 3563–3570. [Google Scholar] [PubMed]
- Van Oostrum, J.; Burnett, R.M. Molecular composition of the adenovirus type 2 virion. J. Virol. 1985, 56, 439–448. [Google Scholar] [PubMed]
- Meulenbroek, R.A.; Sargent, K.L.; Lunde, J.; Jasmin, B.J.; Parks, R.J. Use of adenovirus protein IX (pIX) to display large polypeptides on the virion—Generation of fluorescent virus through the incorporation of pIX-GFP. Mol. Ther. 2004, 9, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Matthews, Q.L.; Sibley, D.A.; Wu, H.; Li, J.; Stoff-Khalili, M.A.; Waehler, R.; Mathis, J.M.; Curiel, D.T. Genetic incorporation of a herpes simplex virus type 1 thymidine kinase and firefly luciferase fusion into the adenovirus protein IX for functional display on the virion. Mol. Imaging 2006, 5, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, I.P.; Kashentseva, E.A.; Curiel, D.T. Engineering of adenovirus vectors containing heterologous peptide sequences in the C terminus of capsid protein IX. J. Virol. 2002, 76, 6893–6899. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, J.; de Vrij, J.; Myhre, S.; Uil, T.; Martineau, P.; Lindholm, L.; Hoeben, R.C. Efficient incorporation of a functional hyper-stable single-chain antibody fragment protein-IX fusion in the adenovirus capsid. Gene Ther. 2007, 14, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.S.; Glasgow, J.N.; Borovjagin, A.; Beatty, M.S.; Kashentseva, E.A.; Curiel, D.T.; Marabini, R.; Dmitriev, I.P. Localization of the N-terminus of minor coat protein IIIa in the adenovirus capsid. J. Mol. Biol. 2008, 383, 923–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curiel, D.T. Capsid-Modified Recombinant Adenovirus and Methods of Use. Patent US20030175244A1, 18 September 2003. [Google Scholar]
- Vujadinovic, M.; Vellinga, J.; Janssen Infectious Diseases and Vaccines, Pharmaceutical Companies of Johnson and Johnson, 2301 CA Leiden, The Netherlands. Personal Communication, 2015.
- Crompton, J.; Toogood, C.I.; Wallis, N.; Hay, R.T. Expression of a foreign epitope on the surface of the adenovirus hexon. J. Gen. Virol. 1994, 75 Pt 1, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, C.; Overstreet, M.G.; Guedon, J.M.; Hoiczyk, E.; Ward, C.; Karen, K.A.; Zavala, F.; Ketner, G. Adenovirus particles that display the Plasmodium falciparum circumsporozoite protein NANP repeat induce sporozoite-neutralizing antibodies in mice. Vaccine 2011, 29, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Karen, K.A.; Deal, C.; Adams, R.J.; Nielsen, C.; Ward, C.; Espinosa, D.A.; Xie, J.; Zavala, F.; Ketner, G. A replicating adenovirus capsid display recombinant elicits antibodies against Plasmodium falciparum sporozoites in Aotus nancymaae monkeys. Infect. Immun. 2015, 83, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Krendelchtchikova, V.; Krendelchtchikov, A.; Oster, R.A.; Fujihashi, K.; Matthews, Q.L. A recombinant adenovirus-based vector elicits a specific humoral immune response against the V3 loop of HIV-1 gp120 in mice through the “Antigen Capsid-Incorporation” strategy. Virol. J. 2014, 11, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Krendelchtchikova, V.; Krendelchtchikov, A.; Farrow, A.L.; Derdeyn, C.A.; Matthews, Q.L. Adenoviral vectors elicit humoral immunity against variable loop 2 of clade C HIV-1 gp120 via “Antigen Capsid-Incorporation” strategy. Virology 2016, 487, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Li, Z.C.; Krendelchtchikov, A.; Krendelchtchikova, V.; Wu, H.; Matthews, Q.L. Using multivalent adenoviral vectors for HIV vaccination. PLoS ONE 2013, 8, e60347. [Google Scholar] [CrossRef] [PubMed]
- Farrow, A.L.; Rachakonda, G.; Gu, L.; Krendelchtchikova, V.; Nde, P.N.; Pratap, S.; Lima, M.F.; Villalta, F.; Matthews, Q.L. Immunization with Hexon modified adenoviral vectors integrated with gp83 epitope provides protection against Trypanosoma cruzi infection. PLoS Negl. Trop. Dis. 2014, 8, e3089. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.H.; Alkutkar, T.; Karanam, B.; Roden, R.B.; Ketner, G.; Ibeanu, O.A. Capsid display of a conserved human papillomavirus L2 peptide in the adenovirus 5 hexon protein: A candidate prophylactic hpv vaccine approach. Virol. J. 2015, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Su, X.; Li, X.; Li, H.; Li, T.; Zhou, Z.; Zhong, T.; Zhou, R. Protection against enterovirus 71 with neutralizing epitope incorporation within adenovirus type 3 hexon. PLoS ONE 2012, 7, e41381. [Google Scholar] [CrossRef]
- Xue, C.; Tian, X.; Li, X.; Zhou, Z.; Su, X.; Zhou, R. Construction and characterization of a recombinant human adenovirus type 3 vector containing two foreign neutralizing epitopes in hexon. Virus Res. 2014, 183, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Wu, T.L.; Emmer, K.L.; Kurupati, R.; Tuyishime, S.; Li, Y.; Giles-Davis, W.; Zhou, X.; Xiang, Z.; Liu, Q.; et al. Hexon-modified recombinant E1-deleted adenovirus vectors as dual specificity vaccine carriers for influenza virus. Mol. Ther. 2013, 21, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Y.; Chi, Y.; Yin, J.; Yan, L.; Ku, Z.; Liu, Q.; Huang, Z.; Zhou, D. Hexon-modified recombinant E1-deleted adenoviral vectors as bivalent vaccine carriers for Coxsackievirus A16 and Enterovirus 71. Vaccine 2015, 33, 5087–5094. [Google Scholar] [CrossRef] [PubMed]
- Matthews, Q.L.; Fatima, A.; Tang, Y.; Perry, B.A.; Tsuruta, Y.; Komarova, S.; Timares, L.; Zhao, C.; Makarova, N.; Borovjagin, A.V.; et al. HIV antigen incorporation within adenovirus hexon hypervariable 2 for a novel HIV vaccine approach. PLoS ONE 2010, 5, e11815. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Danthinne, X.; Imperiale, M.J. Characterization of a permissive epitope insertion site in adenovirus hexon. J. Virol. 2006, 80, 5361–5370. [Google Scholar] [CrossRef] [PubMed]
- Matthews, Q.L.; Yang, P.; Wu, Q.; Belousova, N.; Rivera, A.A.; Stoff-Khalili, M.A.; Waehler, R.; Hsu, H.C.; Li, Z.; Li, J.; et al. Optimization of capsid-incorporated antigens for a novel adenovirus vaccine approach. Virol. J. 2008, 5, 98. [Google Scholar] [CrossRef] [PubMed]
- Worgall, S.; Krause, A.; Qiu, J.; Joh, J.; Hackett, N.R.; Crystal, R.G. Protective immunity to Pseudomonas aeruginosa induced with a capsid-modified adenovirus expressing P. aeruginosa OprF. J. Virol. 2007, 81, 13801–13808. [Google Scholar] [CrossRef] [PubMed]
- Worgall, S.; Krause, A.; Rivara, M.; Hee, K.K.; Vintayen, E.V.; Hackett, N.R.; Roelvink, P.W.; Bruder, J.T.; Wickham, T.J.; Kovesdi, I.; et al. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J. Clin. Investig. 2005, 115, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.L.; Sofer-Podesta, C.; Ang, J.; Hackett, N.R.; Chiuchiolo, M.J.; Senina, S.; Perlin, D.; Crystal, R.G. Protective immunity against a lethal respiratory Yersinia pestis challenge induced by V antigen or the F1 capsular antigen incorporated into adenovirus capsid. Hum. Gene Ther. 2010, 21, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Johrden, L.; Tenbusch, M.; Lietz, R.; Bonsmann, M.S.; Niezold, T.; Wildner, O.; Bayer, W. Comparison of polystyrene nanoparticles and UV-inactivated antigen-displaying adenovirus for vaccine delivery in mice. Virol. J. 2013, 10, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrow, A.L.; Peng, B.J.; Gu, L.; Krendelchtchikov, A.; Matthews, Q.L. A Novel Vaccine Approach for Chagas Disease Using Rare Adenovirus Serotype 48 Vectors. Viruses 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Bayer, W.; Tenbusch, M.; Lietz, R.; Johrden, L.; Schimmer, S.; Uberla, K.; Dittmer, U.; Wildner, O. Vaccination with an adenoviral vector that encodes and displays a retroviral antigen induces improved neutralizing antibody and CD4+ T-cell responses and confers enhanced protection. J. Virol. 2010, 84, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Salisch, N.C.; Vujadinovic, M.; van der Helm, E.; Spek, D.; Vorthoren, L.; Serroyen, J.; Kuipers, H.; Schuitemaker, H.; Zahn, R.; Custers, J.; et al. Antigen capsid-display on human adenovirus 35 via pIX fusion is a potent vaccine platform. PLoS ONE 2017, 12, e0174728. [Google Scholar] [CrossRef] [PubMed]
- Vujadinovic, M.; Khan, S.; Oosterhuis, K.; Uil, T.G.; Wunderlich, K.; Damman, S.; Boedhoe, S.; Verwilligen, A.; Knibbe, J.; Serroyen, J.; et al. Adenovirus based HPV L2 vaccine induces broad cross-reactive humoral immune responses. Vaccine 2018, 36, 4462–4470. [Google Scholar] [CrossRef] [PubMed]
- Krause, A.; Joh, J.H.; Hackett, N.R.; Roelvink, P.W.; Bruder, J.T.; Wickham, T.J.; Kovesdi, I.; Crystal, R.G.; Worgall, S. Epitopes expressed in different adenovirus capsid proteins induce different levels of epitope-specific immunity. J. Virol. 2006, 80, 5523–5530. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, A.; Youssef, G.B.; Perricaudet, M.; Benihoud, K. Anti-adenovirus humoral responses influence on the efficacy of vaccines based on epitope display on adenovirus capsid. Vaccine 2011, 29, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Krause, A.; Xu, Y.; Sung, B.; Wu, W.; Worgall, S. Adenovirus-based vaccine with epitopes incorporated in novel fiber sites to induce protective immunity against Pseudomonas aeruginosa. PLoS ONE 2013, 8, e56996. [Google Scholar] [CrossRef] [PubMed]
- Rux, J.J.; Kuser, P.R.; Burnett, R.M. Structural and phylogenetic analysis of adenovirus hexons by use of high-resolution X-ray crystallographic, molecular modeling, and sequence-based methods. J. Virol. 2003, 77, 9553–9566. [Google Scholar] [CrossRef] [PubMed]
- Crawford-Miksza, L.; Schnurr, D.P. Analysis of 15 adenovirus hexon proteins reveals the location and structure of seven hypervariable regions containing serotype-specific residues. J. Virol. 1996, 70, 1836–1844. [Google Scholar] [PubMed]
- Rux, J.J.; Burnett, R.M. Type-specific epitope locations revealed by X-ray crystallographic study of adenovirus type 5 hexon. Mol. Ther. 2000, 1, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Youil, R.; Toner, T.J.; Su, Q.; Chen, M.; Tang, A.; Bett, A.J.; Casimiro, D. Hexon gene switch strategy for the generation of chimeric recombinant adenovirus. Hum. Gene Ther. 2002, 13, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Okuda, K.; Ura, T.; Kondo, A.; Yoshida, A.; Yoshizaki, S.; Mizuguchi, H.; Klinman, D.; Shimada, M. Adenovirus type 5 with modified hexons induces robust transgene-specific immune responses in mice with pre-existing immunity against adenovirus type 5. J. Gene Med. 2009, 11, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Shiratsuchi, T.; Rai, U.; Krause, A.; Worgall, S.; Tsuji, M. Replacing adenoviral vector HVR1 with a malaria B cell epitope improves immunogenicity and circumvents preexisting immunity to adenovirus in mice. J. Clin. Investig. 2010, 120, 3688–3701. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.E.; Moodie, Z.; Metch, B.; Gilbert, P.B.; Bekker, L.G.; Churchyard, G.; Nchabeleng, M.; Mlisana, K.; Laher, F.; Roux, S.; et al. Recombinant adenovirus type 5 HIV gag/pol/nef vaccine in South Africa: Unblinded, long-term follow-up of the phase 2b HVTN 503/Phambili study. Lancet Infect. Dis. 2014, 14, 388–396. [Google Scholar] [CrossRef]
- Fitzgerald, D.W.; Janes, H.; Robertson, M.; Coombs, R.; Frank, I.; Gilbert, P.; Loufty, M.; Mehrotra, D.; Duerr, A.; Step Study Protocol Team. An Ad5-vectored HIV-1 vaccine elicits cell-mediated immunity but does not affect disease progression in HIV-1-infected male subjects: Results from a randomized placebo-controlled trial (the Step study). J. Infect. Dis. 2011, 203, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Kik, S.V.; Weverling, G.J.; Dilan, R.; King, S.L.; Maxfield, L.F.; Clark, S.; Ng’ang’a, D.; Brandariz, K.L.; Abbink, P.; et al. International seroepidemiology of adenovirus serotypes 5, 26, 35, and 48 in pediatric and adult populations. Vaccine 2011, 29, 5203–5209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledgerwood, J.E.; DeZure, A.D.; Stanley, D.A.; Coates, E.E.; Novik, L.; Enama, M.E.; Berkowitz, N.M.; Hu, Z.; Joshi, G.; Ploquin, A.; et al. Chimpanzee Adenovirus Vector Ebola Vaccine. N. Engl. J. Med. 2017, 376, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, J.; van den Wollenberg, D.J.; van der Heijdt, S.; Rabelink, M.J.; Hoeben, R.C. The coiled-coil domain of the adenovirus type 5 protein IX is dispensable for capsid incorporation and thermostability. J. Virol. 2005, 79, 3206–3210. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.; Rosa-Calatrava, M.; Kedinger, C. The product of the adenovirus intermediate gene IX is a transcriptional activator. J. Virol. 1997, 71, 5102–5109. [Google Scholar] [PubMed]
- Rosa-Calatrava, M.; Puvion-Dutilleul, F.; Lutz, P.; Dreyer, D.; de The, H.; Chatton, B.; Kedinger, C. Adenovirus protein IX sequesters host-cell promyelocytic leukaemia protein and contributes to efficient viral proliferation. EMBO Rep. 2003, 4, 969–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sargent, K.L.; Ng, P.; Evelegh, C.; Graham, F.L.; Parks, R.J. Development of a size-restricted pIX-deleted helper virus for amplification of helper-dependent adenovirus vectors. Gene Ther. 2004, 11, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Colby, W.W.; Shenk, T. Adenovirus type 5 virions can be assembled in vivo in the absence of detectable polypeptide IX. J. Virol. 1981, 39, 977–980. [Google Scholar] [PubMed]
- Ghosh-Choudhury, G.; Haj-Ahmad, Y.; Graham, F.L. Protein IX, a minor component of the human adenovirus capsid, is essential for the packaging of full length genomes. EMBO J. 1987, 6, 1733–1739. [Google Scholar] [PubMed]
- Reddy, V.S.; Nemerow, G.R. Structures and organization of adenovirus cement proteins provide insights into the role of capsid maturation in virus entry and infection. Proc. Natl. Acad. Sci. USA 2014, 111, 11715–11720. [Google Scholar] [CrossRef] [PubMed]
- Akalu, A.; Liebermann, H.; Bauer, U.; Granzow, H.; Seidel, W. The subgenus-specific C-terminal region of protein IX is located on the surface of the adenovirus capsid. J. Virol. 1999, 73, 6182–6187. [Google Scholar] [PubMed]
- Vellinga, J.; Rabelink, M.J.; Cramer, S.J.; van den Wollenberg, D.J.; van der Meulen, H.; Leppard, K.N.; Fallaux, F.J.; Hoeben, R.C. Spacers increase the accessibility of peptide ligands linked to the carboxyl terminus of adenovirus minor capsid protein IX. J. Virol. 2004, 78, 3470–3479. [Google Scholar] [CrossRef] [PubMed]
- Campos, S.K.; Barry, M.A. Comparison of adenovirus fiber, protein IX, and hexon capsomeres as scaffolds for vector purification and cell targeting. Virology 2006, 349, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Campos, S.K.; Parrott, M.B.; Barry, M.A. Avidin-based targeting and purification of a protein IX-modified, metabolically biotinylated adenoviral vector. Mol. Ther. 2004, 9, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Le, L.P.; Everts, M.; Dmitriev, I.P.; Davydova, J.G.; Yamamoto, M.; Curiel, D.T. Fluorescently labeled adenovirus with pIX-EGFP for vector detection. Mol. Imaging 2004, 3, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Bauer, U.; Flunker, G.; Bruss, K.; Kallwellis, K.; Liebermann, H.; Luettich, T.; Motz, M.; Seidel, W. Detection of antibodies against adenovirus protein IX, fiber, and hexon in human sera by immunoblot assay. J. Clin. Microbiol. 2005, 43, 4426–4433. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Komiya, K.; Murata, M.; Kimura, T.; Kanaoka, M.; Kanegae, Y.; Saito, I. Expression of pIX gene induced by transgene promoter: Possible cause of host immune response in first-generation adenoviral vectors. Hum. Gene Ther. 2007, 18, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Dong, X.; Wu, X.; Wen, B.; Ji, G.; Cheng, L.; Liu, H. Conserved fiber-penton base interaction revealed by nearly atomic resolution cryo-electron microscopy of the structure of adenovirus provides insight into receptor interaction. J. Virol. 2012, 86, 12322–12329. [Google Scholar] [CrossRef] [PubMed]
- Arnberg, N. Adenovirus receptors: Implications for tropism, treatment and targeting. Rev. Med. Virol. 2009, 19, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Nicklin, S.A.; Wu, E.; Nemerow, G.R.; Baker, A.H. The influence of adenovirus fiber structure and function on vector development for gene therapy. Mol. Ther. 2005, 12, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Law, L.K.; Davidson, B.L. What does it take to bind CAR? Mol. Ther. 2005, 12, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Zubieta, C.; Schoehn, G.; Chroboczek, J.; Cusack, S. The structure of the human adenovirus 2 penton. Mol. Cell 2005, 17, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Kamata, T.; Takada, Y.; Ruggeri, Z.M.; Nemerow, G.R. Adenovirus interaction with distinct integrins mediates separate events in cell entry and gene delivery to hematopoietic cells. J. Virol. 1996, 70, 4502–4508. [Google Scholar] [PubMed]
- Roelvink, P.W.; Kovesdi, I.; Wickham, T.J. Comparative analysis of adenovirus fiber-cell interaction: Adenovirus type 2 (Ad2) and Ad9 utilize the same cellular fiber receptor but use different binding strategies for attachment. J. Virol. 1996, 70, 7614–7621. [Google Scholar] [PubMed]
- Michael, S.I.; Hong, J.S.; Curiel, D.T.; Engler, J.A. Addition of a short peptide ligand to the adenovirus fiber protein. Gene Ther. 1995, 2, 660–668. [Google Scholar] [PubMed]
- Krasnykh, V.N.; Mikheeva, G.V.; Douglas, J.T.; Curiel, D.T. Generation of recombinant adenovirus vectors with modified fibers for altering viral tropism. J. Virol. 1996, 70, 6839–6846. [Google Scholar] [PubMed]
- Gall, J.; Kass-Eisler, A.; Leinwand, L.; Falck-Pedersen, E. Adenovirus type 5 and 7 capsid chimera: Fiber replacement alters receptor tropism without affecting primary immune neutralization epitopes. J. Virol. 1996, 70, 2116–2123. [Google Scholar] [PubMed]
- Stevenson, S.C.; Rollence, M.; Marshall-Neff, J.; McClelland, A. Selective targeting of human cells by a chimeric adenovirus vector containing a modified fiber protein. J. Virol. 1997, 71, 4782–4790. [Google Scholar] [PubMed]
- Coughlan, L.; Alba, R.; Parker, A.L.; Bradshaw, A.C.; McNeish, I.A.; Nicklin, S.A.; Baker, A.H. Tropism-modification strategies for targeted gene delivery using adenoviral vectors. Viruses 2010, 2, 2290–2355. [Google Scholar] [CrossRef] [PubMed]
- Rea, D.; Havenga, M.J.; van den Assem, M.; Sutmuller, R.P.; Lemckert, A.; Hoeben, R.C.; Bout, A.; Melief, C.J.; Offringa, R. Highly efficient transduction of human monocyte-derived dendritic cells with subgroup B fiber-modified adenovirus vectors enhances transgene-encoded antigen presentation to cytotoxic T cells. J. Immunol. 2001, 166, 5236–5244. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Gall, J.G.; Kong, W.P.; Sheets, R.L.; Gomez, P.L.; King, C.R.; Nabel, G.J. Mechanism of ad5 vaccine immunity and toxicity: Fiber shaft targeting of dendritic cells. PLoS Pathog. 2007, 3, e25. [Google Scholar] [CrossRef] [PubMed]
- Someya, K.; Xin, K.Q.; Ami, Y.; Izumi, Y.; Mizuguchi, H.; Ohta, S.; Yamamoto, N.; Honda, M.; Okuda, K. Chimeric adenovirus type 5/35 vector encoding SIV gag and HIV env genes affords protective immunity against the simian/human immunodeficiency virus in monkeys. Virology 2007, 367, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.; Lynch, D.M.; Goudsmit, J.; Lemckert, A.A.; Ewald, B.A.; Sumida, S.M.; Truitt, D.M.; Abbink, P.; Kishko, M.G.; Gorgone, D.A.; et al. Immunogenicity of recombinant fiber-chimeric adenovirus serotype 35 vector-based vaccines in mice and rhesus monkeys. J. Virol. 2005, 79, 14161–14168. [Google Scholar] [CrossRef] [PubMed]
- Xin, K.Q.; Sekimoto, Y.; Takahashi, T.; Mizuguchi, H.; Ichino, M.; Yoshida, A.; Okuda, K. Chimeric adenovirus 5/35 vector containing the clade C HIV gag gene induces a cross-reactive immune response against HIV. Vaccine 2007, 25, 3809–3815. [Google Scholar] [CrossRef] [PubMed]
- Kurachi, S.; Koizumi, N.; Sakurai, F.; Kawabata, K.; Sakurai, H.; Nakagawa, S.; Hayakawa, T.; Mizuguchi, H. Characterization of capsid-modified adenovirus vectors containing heterologous peptides in the fiber knob, protein IX, or hexon. Gene Ther. 2007, 14, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Kurachi, S.; Tashiro, K.; Sakurai, F.; Sakurai, H.; Kawabata, K.; Yayama, K.; Okamoto, H.; Nakagawa, S.; Mizuguchi, H. Fiber-modified adenovirus vectors containing the TAT peptide derived from HIV-1 in the fiber knob have efficient gene transfer activity. Gene Ther. 2007, 14, 1160–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasnykh, V.; Dmitriev, I.; Mikheeva, G.; Miller, C.R.; Belousova, N.; Curiel, D.T. Characterization of an adenovirus vector containing a heterologous peptide epitope in the HI loop of the fiber knob. J. Virol. 1998, 72, 1844–1852. [Google Scholar] [PubMed]
- Magnusson, M.K.; Henning, P.; Myhre, S.; Wikman, M.; Uil, T.G.; Friedman, M.; Andersson, K.M.; Hong, S.S.; Hoeben, R.C.; Habib, N.A.; et al. Adenovirus 5 vector genetically re-targeted by an Affibody molecule with specificity for tumor antigen HER2/neu. Cancer Gene Ther. 2007, 14, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uil, T.G.; Seki, T.; Dmitriev, I.; Kashentseva, E.; Douglas, J.T.; Rots, M.G.; Middeldorp, J.M.; Curiel, D.T. Generation of an adenoviral vector containing an addition of a heterologous ligand to the serotype 3 fiber knob. Cancer Gene Ther. 2003, 10, 121–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, T.J.; Carrion, M.E.; Kovesdi, I. Targeting of adenovirus penton base to new receptors through replacement of its RGD motif with other receptor-specific peptide motifs. Gene Ther. 1995, 2, 750–756. [Google Scholar] [PubMed]
- Einfeld, D.A.; Brough, D.E.; Roelvink, P.W.; Kovesdi, I.; Wickham, T.J. Construction of a pseudoreceptor that mediates transduction by adenoviruses expressing a ligand in fiber or penton base. J. Virol. 1999, 73, 9130–9136. [Google Scholar] [PubMed]
- Custers, J.H.H.V.; Vellinga, J.; Vujadinovic, M.; van der Helm, E. An Improved Adenovirus Based Malaria Vaccine Encoding and Displaying a Malaria Antigen. Patent WO/2017/125463, 27 July 2017. [Google Scholar]
- Vujadinovic, M.; Uil, T.G.; Oosterhuis, K.N.; Vellinga, J.N.; Custers, J. HPV VACCINES. Patent WO/2018/011196, 18 January 2018. [Google Scholar]
- Nieto, K.; Weghofer, M.; Sehr, P.; Ritter, M.; Sedlmeier, S.; Karanam, B.; Seitz, H.; Muller, M.; Kellner, M.; Horer, M.; et al. Development of AAVLP(HPV16/31L2) particles as broadly protective HPV vaccine candidate. PLoS ONE 2012, 7, e39741. [Google Scholar] [CrossRef] [PubMed]
- Skamel, C.; Ploss, M.; Bottcher, B.; Stehle, T.; Wallich, R.; Simon, M.M.; Nassal, M. Hepatitis B virus capsid-like particles can display the complete, dimeric outer surface protein C and stimulate production of protective antibody responses against Borrelia burgdorferi infection. J. Biol. Chem. 2006, 281, 17474–17481. [Google Scholar] [CrossRef] [PubMed]
- Schellenbacher, C.; Roden, R.; Kirnbauer, R. Chimeric L1-L2 virus-like particles as potential broad-spectrum human papillomavirus vaccines. J. Virol. 2009, 83, 10085–10095. [Google Scholar] [CrossRef] [PubMed]
- Moreno, N.; Mena, I.; Angulo, I.; Gomez, Y.; Crisci, E.; Montoya, M.; Caston, J.R.; Blanco, E.; Barcena, J. Rabbit hemorrhagic disease virus capsid, a versatile platform for foreign B-cell epitope display inducing protective humoral immune responses. Sci. Rep. 2016, 6, 31844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Ku, Z.; Liu, Q.; Wang, X.; Shi, J.; Zhang, Y.; Kong, L.; Cong, Y.; Huang, Z. Chimeric virus-like particle vaccines displaying conserved enterovirus 71 epitopes elicit protective neutralizing antibodies in mice through divergent mechanisms. J. Virol. 2014, 88, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Gahery-Segard, H.; Farace, F.; Godfrin, D.; Gaston, J.; Lengagne, R.; Tursz, T.; Boulanger, P.; Guillet, J.G. Immune response to recombinant capsid proteins of adenovirus in humans: Antifiber and anti-penton base antibodies have a synergistic effect on neutralizing activity. J. Virol. 1998, 72, 2388–2397. [Google Scholar] [PubMed]
- Stallwood, Y.; Fisher, K.D.; Gallimore, P.H.; Mautner, V. Neutralisation of adenovirus infectivity by ascitic fluid from ovarian cancer patients. Gene Ther. 2000, 7, 637–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabel, F.; Kundig, T.M.; Bachmann, M.F. Virus-induced humoral immunity: On how B cell responses are initiated. Curr. Opin. Virol. 2013, 3, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Zinkernagel, R.M. Neutralizing antiviral B cell responses. Annu. Rev. Immunol. 1997, 15, 235–270. [Google Scholar] [CrossRef] [PubMed]
- Scherer, J.; Vallee, R.B. Conformational changes in the adenovirus hexon subunit responsible for regulating cytoplasmic dynein recruitment. J. Virol. 2015, 89, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Danthinne, X.; Imperiale, M.J. Production of first generation adenovirus vectors: A review. Gene Ther. 2000, 7, 1707–1714. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| AdV Location | Vector | Disease and Antigen/Epitope (Size aa) | Other Modifications | Reference | |
|---|---|---|---|---|---|
| Transgene in E1 | Capsid | ||||
| Hexon HVR1 | HAdV5 | Polio type 3 (8 aa) | No | No | [56] |
| Malaria Circumsporozoite protein (CSP) (20, 24 aa) and (12 aa) | No | No | [57,58,59] | ||
| HIVgp120 (10, 21, 24, 26 aa) and HIV gp41 and His6 (7, 6 aa) | Yes | Yes | [60,61] | ||
| Chagas disease Trypanosoma cruzi gp83 (24 aa) | No | No | [62] | ||
| Human papillomavirus (HPV) HPV16 L2 protein (29 aa) | No | Yes | [63] | ||
| HAdV3 | Enterovirus VP1 SP70 (15 aa) and Enterovirus VP1 SP70 and SP55 (15 aa) | Yes | Yes | [64,65] | |
| SAdV25 (AdC68) | Influenza A M2e and NP antigen (13 aa) | Yes | Yes | [66] | |
| Coxsackievirus and Enterovirus VP1 (6, 15 aa) | Yes | Yes | [67] | ||
| Hexon HVR2 | HAdV5 | HIV gp41 and Gag protein (39 aa) | Yes | No | [68] |
| Hexon HVR5 | HAdV5 | Anthrax Bacillus anthracis Protective antigen (PA) (16, 36, 66,143, 245 aa (eGFP)) | No | No | [69] |
| Model antigen RGD motif + His-linker (45, 55, 65, 75, 85, 95 aa) | Yes | Yes | [70] | ||
| Pseudomonas aeruginosa outer membrane protein F (OprF) (14 aa) | Yes | Yes | [71,72] | ||
| pIX C-terminus | HAdV5 | Plague Yersinia pestis (15, 37 aa) | Yes | No | [73] |
| Ovalbumin (Ova) (43 aa) | No | Yes | [74] | ||
| HAdV5 and HAdV48 | Chagas-disease vaccine ASP2 and gp83 (26, 24 aa) | Yes | No | [75] | |
| HAdV5 | HIV gp120 (67 aa) | No | Yes | [60] | |
| Friend Murine Leukemia Virus gp70, Gag and Ova (22, 43, 60, 70, 92) HAdV5/HAdV5.F35 | Yes | Yes | [76] | ||
| HAdV35 | Malaria P. falciparum Circumsprozoite protein (CSP) (17, 20, 40 aa) | Yes | No | [77] | |
| Human papillomavirus (HPV) HPV6, 11, 16, 18, 31, 45, 52/58 L2 protein (93 aa) | No | No | [78] | ||
| Fiber Knob-loop | HAdV5 | Influenza A HA (9 aa) | Yes | Yes | [79] |
| Ovalbumin (20, 22 aa) | Yes | Yes | [80] | ||
| Pseudomonas aeruginosa (14 aa) | Yes | Yes | [81] | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujadinovic, M.; Vellinga, J. Progress in Adenoviral Capsid-Display Vaccines. Biomedicines 2018, 6, 81. https://doi.org/10.3390/biomedicines6030081
Vujadinovic M, Vellinga J. Progress in Adenoviral Capsid-Display Vaccines. Biomedicines. 2018; 6(3):81. https://doi.org/10.3390/biomedicines6030081
Chicago/Turabian StyleVujadinovic, Marija, and Jort Vellinga. 2018. "Progress in Adenoviral Capsid-Display Vaccines" Biomedicines 6, no. 3: 81. https://doi.org/10.3390/biomedicines6030081
APA StyleVujadinovic, M., & Vellinga, J. (2018). Progress in Adenoviral Capsid-Display Vaccines. Biomedicines, 6(3), 81. https://doi.org/10.3390/biomedicines6030081