Osteoblastogenesis Alters Small RNA Profiles in EVs Derived from Bone Marrow Stem Cells (BMSCs) and Adipose Stem Cells (ASCs)


Abstract
:
1. Introduction
2. Experimental Section
2.1. Cell Culture and Osteoblastic Differentiation
2.2. EV Purification
2.3. Nanoparticle Tracking Analysis (NTA) of EVs
2.4. Western Blotting of EVs
2.5. RNA Extraction
2.6. Small RNA Library Preparation and Sequencing
2.7. Raw Data Processing
3. Results
3.1. Characterization of EVs from BMSCs and ASCs
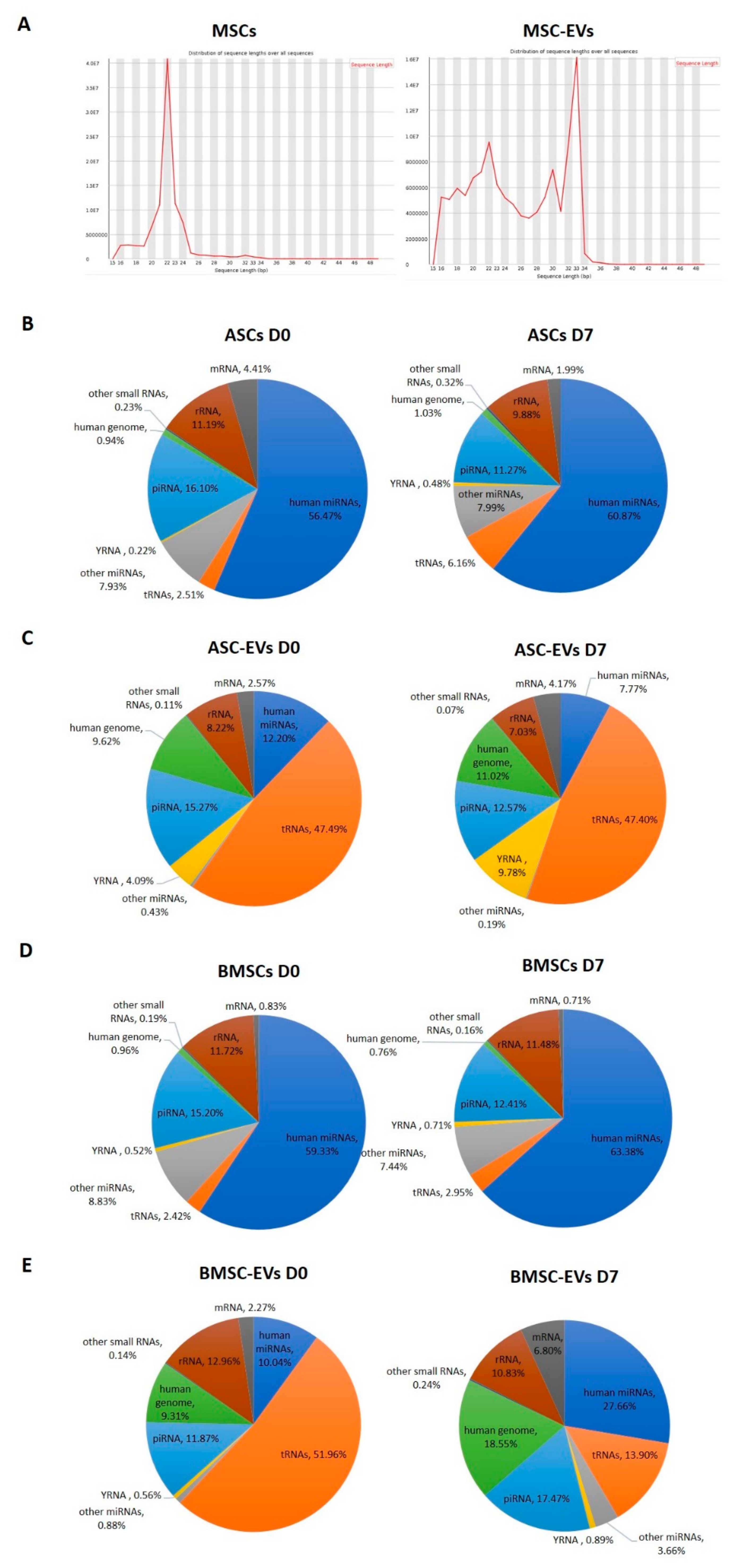
3.2. Small RNA Profiles of MSCs and MSC-EVs
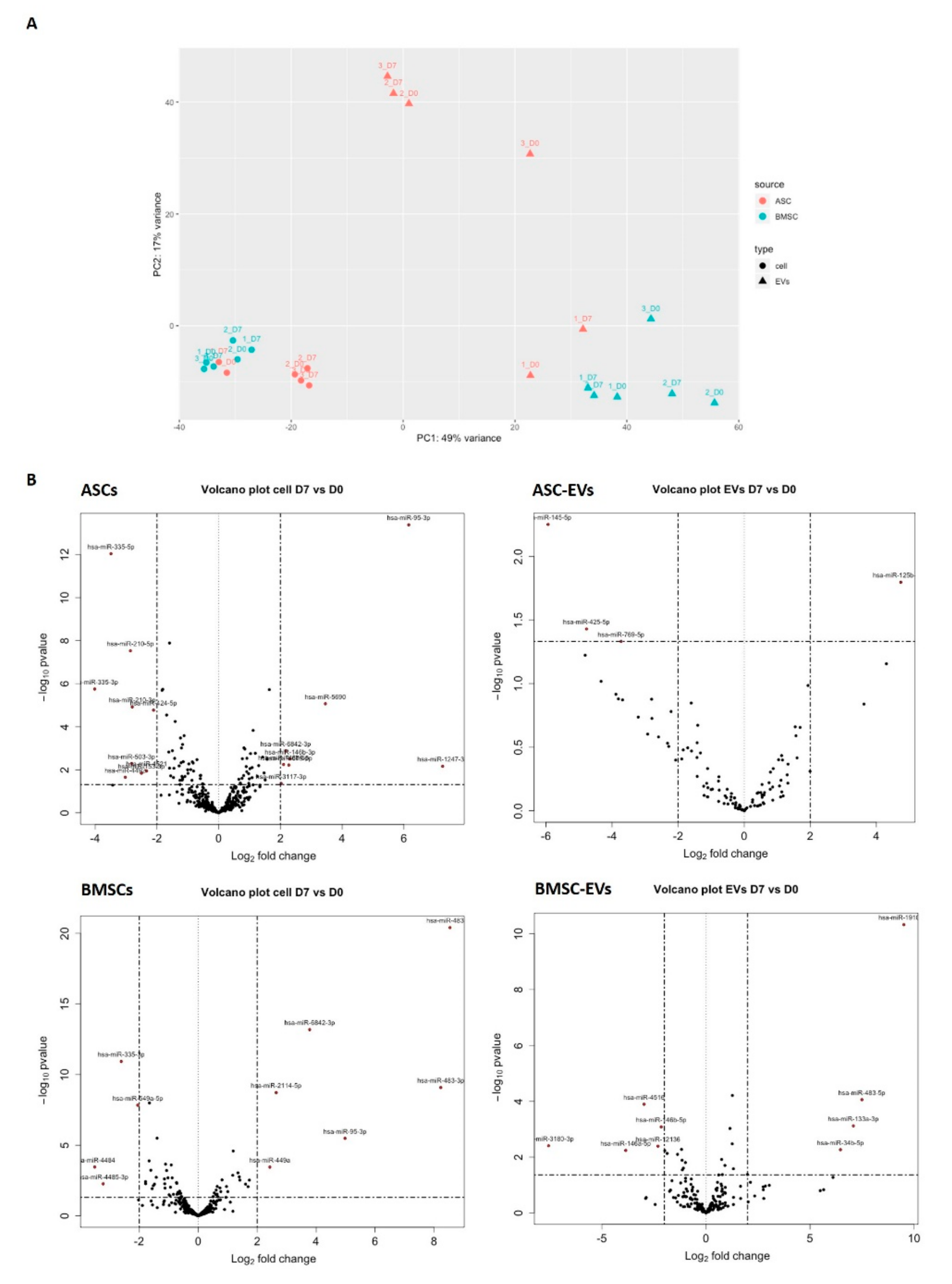
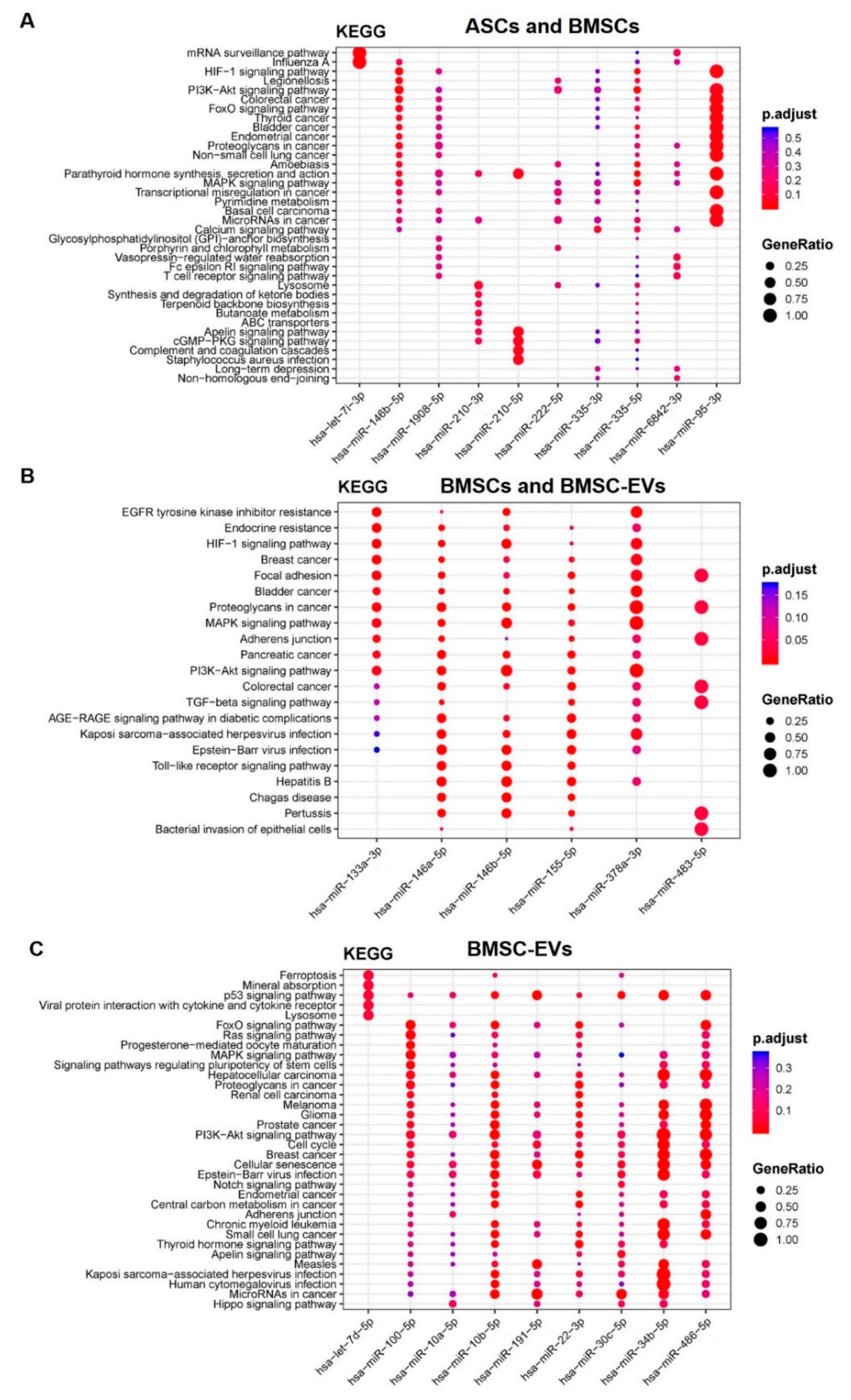
3.3. Osteoblastic Differentiation Alters miRNA Expression in MSCs and MSC-EVs
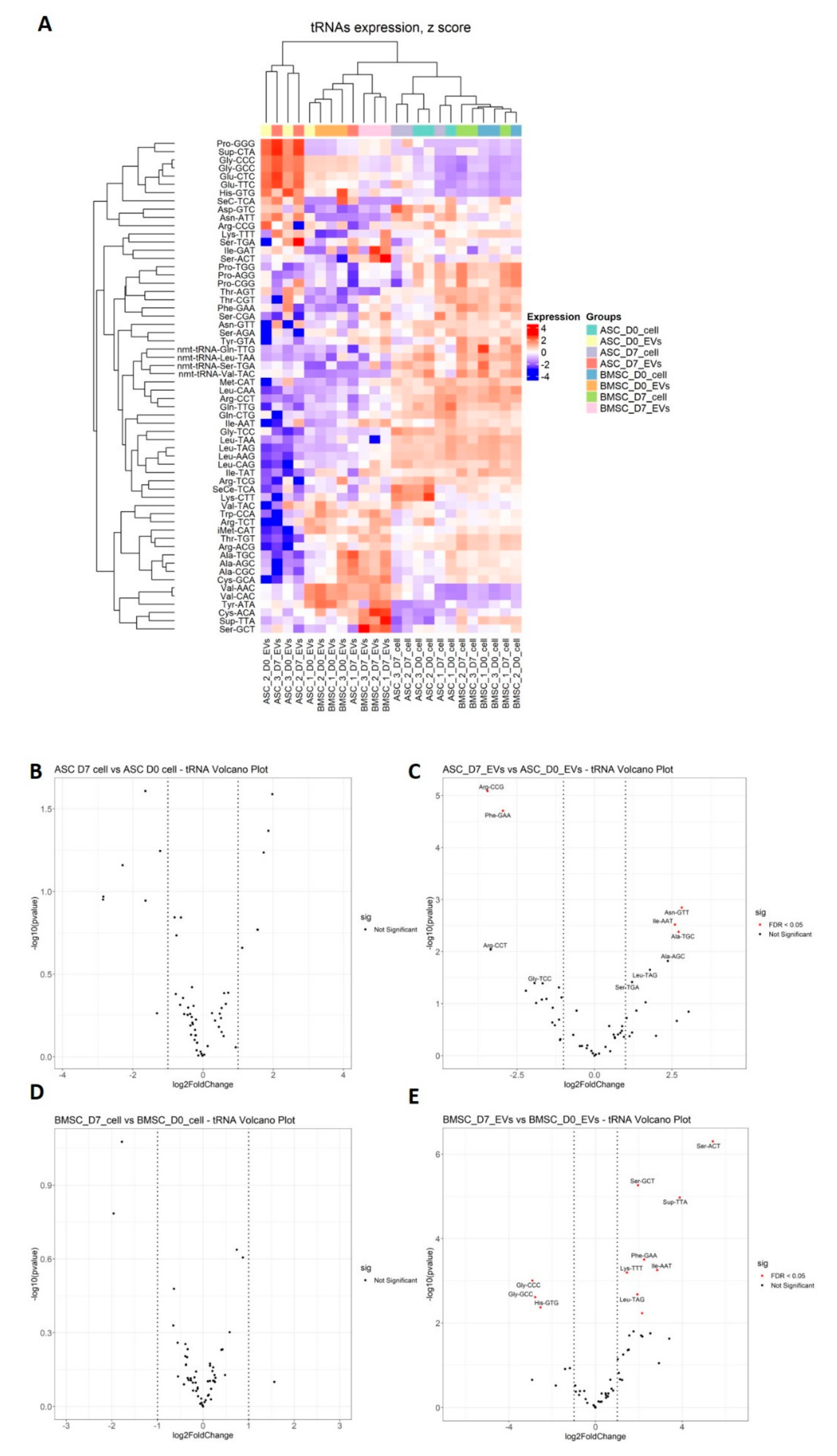
3.4. Differential tsRNA Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baglio, S.R.; Rooijers, K.; Koppers-Lalic, D.; Verweij, F.J.; Lanzón, M.P.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.; Baldini, N.; et al. Human bone marrow-and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 2015, 6, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oryan, A.; Kamali, A.; Moshiri, A.; Eslaminejad, M.B. Role of mesenchymal stem cells in bone regenerative medicine: What is the evidence? Cells Tissues Organs 2017, 204, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhu, W.; Lu, J.; Xie, K.; Fang, S.; Kan, L. Potential Therapeutic Applications of Exosomes in Bone Regenerative Medicine. In Osteogenesis and Bone Regeneration; IntechOpen: London, UK, 2018. [Google Scholar]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, K.C.; Penfornis, P.; Dhule, S.; Guillonneau, F.; Adams, K.V.; Mo, Y.Y.; Xu, R.; Liu, Y.; Watabe, K.; Vemuri, M.C. Extracellular vesicles from bone marrow mesenchymal stem/stromal cells transport tumor regulatory microRNA, proteins, and metabolites. Oncotarget 2015, 6, 4953. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Würdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [Green Version]
- Chiou, N.-T.; Kageyama, R.; Ansel, K.M. Selective export into extracellular vesicles and function of tRNA fragments during T cell activation. Cell Rep. 2018, 25, 3356.e4–3370.e4. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.-F.; Yang, G.-H.; Pan, X.-H.; Zhang, S.-J.; Zhao, C.; Qiu, B.-S.; Gu, H.-F.; Hong, J.-F.; Cao, L.; Chen, Y. Altered microRNA expression profile in exosomes during osteogenic differentiation of human bone marrow-derived mesenchymal stem cells. PLoS ONE 2014, 9, e114627. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuscu, C.; Kumar, P.; Kiran, M.; Su, Z.; Malik, A.; Dutta, A. tRNA fragments (tRFs) guide Ago to regulate gene expression post-transcriptionally in a Dicer-independent manner. Rna 2018, 24, 1093–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricker, R.; Brogli, R.; Luidalepp, H.; Wyss, L.; Fasnacht, M.; Joss, O.; Zywicki, M.; Helm, M.; Schneider, A.; Cristodero, M. A tRNA half modulates translation as stress response in Trypanosoma brucei. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.; Feng, J.; Liu, Q.; Sun, F.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Stress induces tRNA cleavage by angiogenin in mammalian cells. Febs Lett. 2009, 583, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhahbi, J.M. 5′ tRNA halves: The next generation of immune signaling molecules. Front. Immunol. 2015, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Rossmanith, W. Of P and Z: Mitochondrial tRNA processing enzymes. Biochim. Biophys. Acta Bba Gene Regul. Mech. 2012, 1819, 1017–1026. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yu, X.; Zhu, L.; Li, T.; Yan, Z.; Guo, J. Transfer RNA-derived fragments and tRNA halves: Biogenesis, biological functions and their roles in diseases. J. Mol. Med. 2018, 96, 1167–1176. [Google Scholar] [CrossRef]
- Gebetsberger, J.; Wyss, L.; Mleczko, A.M.; Reuther, J.; Polacek, N. A tRNA-derived fragment competes with mRNA for ribosome binding and regulates translation during stress. Rna Biol. 2017, 14, 1364–1373. [Google Scholar] [CrossRef] [Green Version]
- Gebetsberger, J.; Zywicki, M.; Künzi, A.; Polacek, N. tRNA-derived fragments target the ribosome and function as regulatory non-coding RNA in Haloferax volcanii. Archaea 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, P.; Emara, M.M.; Villen, J.; Gygi, S.P.; Anderson, P. Angiogenin-induced tRNA fragments inhibit translation initiation. Mol. Cell 2011, 43, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigematsu, M.; Kirino, Y. tRNA-derived short non-coding RNA as interacting partners of argonaute proteins. Gene Regul. Syst. Biol. 2015, 9, GRSB-S29411. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Karaiskos, S.; Grigoriev, A. Inferring targeting modes of Argonaute-loaded tRNA fragments. Rna Biol. 2019, 17, 1070–1080. [Google Scholar] [CrossRef]
- Raynaud, C.M.; Rafii, A. The necessity of a systematic approach for the use of MSCs in the clinical setting. Stem Cells Int. 2013, 2013, 892340. [Google Scholar] [CrossRef] [PubMed]
- Dumic-Cule, I.; Pecina, M.; Jelic, M.; Jankolija, M.; Popek, I.; Grgurevic, L.; Vukicevic, S. Biological aspects of segmental bone defects management. Int. Orthop. 2015, 39, 1005–1011. [Google Scholar] [CrossRef] [PubMed]
- Decambron, A.; Fournet, A.; Bensidhoum, M.; Manassero, M.; Sailhan, F.; Petite, H.; Logeart-Avramoglou, D.; Viateau, V. Low-dose BMP-2 and MSC dual delivery onto coral scaffold for critical-size bone defect regeneration in sheep. J. Orthop. Res. 2017, 35, 2637–2645. [Google Scholar] [CrossRef] [PubMed]
- Lindroos, B.; Suuronen, R.; Miettinen, S. The potential of adipose stem cells in regenerative medicine. Stem Cell Rev. Rep. 2011, 7, 269–291. [Google Scholar] [CrossRef]
- Qin, Y.; Sun, R.; Wu, C.; Wang, L.; Zhang, C. Exosome: A novel approach to stimulate bone regeneration through regulation of osteogenesis and angiogenesis. Int. J. Mol. Sci. 2016, 17, 712. [Google Scholar] [CrossRef]
- Zhang, L.; Jiao, G.; Ren, S.; Zhang, X.; Li, C.; Wu, W.; Wang, H.; Liu, H.; Zhou, H.; Chen, Y.; et al. Exosomes from bone marrow mesenchymal stem cells enhance fracture healing through the promotion of osteogenesis and angiogenesis in a rat model of nonunion. Stem Cell Res. Ther. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, L.; Gao, Z.; Chen, G.; Zhang, C. Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity and differentiation in vitro and promote bone regeneration in vivo. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Furuta, T.; Miyaki, S.; Ishitobi, H.; Ogura, T.; Kato, Y.; Kamei, N.; Miyado, K.; Higashi, Y.; Ochi, M. Mesenchymal stem cell-derived exosomes promote fracture healing in a mouse model. Stem Cells Transl. Med. 2016, 5, 1620–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Liu, Y.; Zhang, P.; Tang, Y.; Zhou, M.; Jiang, W.; Zhang, X.; Wu, G.; Zhou, Y. Tissue-engineered bone immobilized with human adipose stem cells-derived exosomes promotes bone regeneration. Acs Appl. Mater. Interfaces 2018, 10, 5240–5254. [Google Scholar] [CrossRef]
- Lu, Z.; Chen, Y.; Dunstan, C.; Roohani-Esfahani, S.; Zreiqat, H. Priming adipose stem cells with tumor necrosis factor-alpha preconditioning potentiates their exosome efficacy for bone regeneration. Tissue Eng. Part A 2017, 23, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Licursi, V.; Conte, F.; Fiscon, G.; Paci, P. MIENTURNET: An interactive web tool for microRNA-target enrichment and network-based analysis. BMC Bioinform. 2019, 20, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Wang, Y.; Hao, Y.; Juan, L.; Teng, M.; Zhang, X.; Li, M.; Wang, G.; Liu, Y. miR2Disease: A manually curated database for microRNA deregulation in human disease. Nucleic Acids Res. 2009, 37, D98–D104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linares, R.; Tan, S.; Gounou, C.; Arraud, N.; Brisson, A.R. High-speed centrifugation induces aggregation of extracellular vesicles. J. Extracell. Vesicles 2015, 4, 29509. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Lee, H.; Jung, C.H.; Jeon, T.I.; Ha, T.Y. MicroRNA-146b promotes adipogenesis by suppressing the SIRT1-FOXO1 cascade. EMBO Mol. Med. 2013, 5, 1602–1612. [Google Scholar] [CrossRef]
- Budd, E.; De Andrés, M.C.; Sanchez-Elsner, T.; Oreffo, R.O. MiR-146b is down-regulated during the chondrogenic differentiation of human bone marrow derived skeletal stem cells and up-regulated in osteoarthritis. Sci. Rep. 2017, 7, 46704. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-C.; Venø, M.T.; Chen, L.; Ditzel, N.; Le, D.Q.; Dillschneider, P.; Kassem, M.; Kjems, J. Global microRNA profiling in human bone marrow skeletal—stromal or mesenchymal–stem cells identified candidates for bone regeneration. Mol. Ther. 2018, 26, 593–605. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Hong, X.; Le Bras, A.; Nowak, W.N.; Bhaloo, S.I.; Deng, J.; Xie, Y.; Hu, Y.; Ruan, X.Z.; Xu, Q. Smooth muscle cells differentiated from mesenchymal stem cells are regulated by microRNAs and suitable for vascular tissue grafts. J. Biol. Chem. 2018, 293, 8089–8102. [Google Scholar] [CrossRef] [Green Version]
- Avendaño-Félix, M.; Fuentes-Mera, L.; Ramos-Payan, R.; Aguilar-Medina, M.; Pérez-Silos, V.; Moncada-Saucedo, N.; Marchat, L.A.; González-Barrios, J.A.; Ruiz-García, E.; Astudillo-de la Vega, H. A Novel OsteomiRs Expression Signature for Osteoblast Differentiation of Human Amniotic Membrane-Derived Mesenchymal Stem Cells. Biomed. Res. Int. 2019, 2019, 8987268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Peng, B.; Zhu, X.; Wang, P.; Sun, K.; Lei, X.; He, H.; Tian, Y.; Mo, S.; Zhang, R. MiR-210-3p inhibits osteogenic differentiation and promotes adipogenic differentiation correlated with Wnt signaling in ERα-deficient rBMSCs. J. Cell. Physiol. 2019, 234, 23475–23484. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Chen, S.; Cai, P.; Chen, K.; Li, L.; Yang, X.; Yi, J.; Luo, X.; Du, Y.; Zheng, H.J.M.; et al. MiRNA-483–5p is involved in the pathogenesis of osteoporosis by promoting osteoclast differentiation. Mol. Cell. Probes 2020, 49, 101479. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, S.; Li, J.; Zhao, S.; Yi, Z. Roles for miRNAs in osteogenic differentiation of bone marrow mesenchymal stem cells. Stem Cell Res. Ther. 2019, 10, 197. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.; Zhang, Y.; Ma, L.; Lin, L.; Meng, J.; Jiang, L.; Wang, L.; Zhou, P.; Zhang, Y.J.B.; et al. LncRNA MEG3 inhibited osteogenic differentiation of bone marrow mesenchymal stem cells from postmenopausal osteoporosis by targeting miR-133a-3p. Biomed. Pharmacother. 2017, 89, 1178–1186. [Google Scholar] [CrossRef]
- Liu, H.; Zhong, L.; Yuan, T.; Chen, S.; Zhou, Y.; An, L.; Guo, Y.; Fan, M.; Li, Y.; Sun, Y. MicroRNA-155 inhibits the osteogenic differentiation of mesenchymal stem cells induced by BMP9 via downregulation of BMP signaling pathway. Int. J. Mol. Med. 2018, 41, 3379–3393. [Google Scholar] [CrossRef]
- Xie, Q.; Wei, W.; Ruan, J.; Ding, Y.; Zhuang, A.; Bi, X.; Sun, H.; Gu, P.; Wang, Z.; Fan, X. Effects of miR-146a on the osteogenesis of adipose-derived mesenchymal stem cells and bone regeneration. Sci. Rep. 2017, 7, 42840. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | miRNA Name | BaseMean | Log2Fold Change | Sample | miRNA Name | BaseMean | Log2Fold Change |
|---|---|---|---|---|---|---|---|
| ASC cells | hsa-let-7i-3p | 479 | −1.6 | BMSC cells | hsa-let-7i-3p | 427 | −1.0 |
| hsa-miR-146b-5p | 20821 | 2.1 | hsa-miR-146b-5p | 3630 | −1.1 | ||
| hsa-miR-1908-5p | 39 | 1.5 | hsa-miR-1908-5p | 31 | 1.7 | ||
| hsa-miR-210-3p | 1191 | −2.8 | hsa-miR-210-3p | 1236 | −1.7 | ||
| hsa-miR-210-5p | 89 | −2.9 | hsa-miR-210-5p | 104 | −1.5 | ||
| hsa-miR-222-5p | 151 | −1.5 | hsa-miR-222-5p | 443 | 1.0 | ||
| hsa-miR-335-3p | 1681 | −4.0 | hsa-miR-335-3p | 3536 | −2.6 | ||
| hsa-miR-335-5p | 1248 | −3.5 | hsa-miR-335-5p | 2835 | −1.7 | ||
| hsa-miR-6842-3p | 281 | 2.2 | hsa-miR-6842-3p | 65 | 3.8 | ||
| hsa-miR-95-3p | 172 | 6.2 | hsa-miR-95-3p | 21 | 5.0 | ||
| ASC cells | hsa-miR-125b-2-3p | 808 | 1.1 | ASC-EVs | hsa-miR-425-5p | 15 | −4.8 |
| hsa-miR-425-5p | 1440 | 1.1 | hsa-miR-125b-2-3p | 29 | 4.7 | ||
| BMSC cells | hsa-miR-133a-3p | 120 | 1.3 | BMSC-EVs | hsa-miR-133a-3p | 15 | 7.1 |
| hsa-miR-146a-5p | 86 | −1.1 | hsa-miR-146a-5p | 26 | −3.9 | ||
| hsa-miR-146b-5p | 3630 | −1.1 | hsa-miR-146b-5p | 1450 | −2.2 | ||
| hsa-miR-155-5p | 871 | −1.4 | hsa-miR-155-5p | 183 | −1.1 | ||
| hsa-miR-378a-3p | 2747 | −1.4 | hsa-miR-378a-3p | 1026 | −1.9 | ||
| hsa-miR-483-5p | 168 | 8.5 | hsa-miR-483-5p | 20 | 7.5 |
| miRNA Name | BaseMean | Log2FoldChange |
|---|---|---|
| hsa-miR-10b-5p | 172,238 | −1.1 |
| hsa-miR-10a-5p | 71,894 | −2.0 |
| hsa-miR-22-3p | 44,127 | 1.3 |
| hsa-miR-191-5p | 30,963 | −1.4 |
| hsa-miR-486-5p | 22,431 | −1.2 |
| hsa-miR-100-5p | 20,117 | −1.1 |
| hsa-miR-146b-5p | 1450 | −2.2 |
| hsa-miR-378a-3p | 1026 | −1.9 |
| hsa-let-7d-5p | 908 | 1.2 |
| hsa-miR-30c-5p | 370 | 1.3 |
| hsa-miR-155-5p | 183 | −1.1 |
| hsa-let-7d-3p | 168 | 1.3 |
| hsa-miR-12136 | 94 | −2.3 |
| hsa-miR-1910-5p | 82 | 9.5 |
| hsa-miR-4516 | 81 | −3.0 |
| hsa-miR-877-5p | 42 | 2.0 |
| hsa-miR-146a-5p | 26 | −3.9 |
| hsa-miR-483-5p | 20 | 7.5 |
| hsa-miR-133a-3p | 15 | 7.1 |
| hsa-miR-3180-3p | 14 | −7.6 |
| hsa-miR-34b-5p | 10 | 6.5 |
| miRNA Name | BaseMean | Log2FoldChange |
|---|---|---|
| hsa-miR-769-5p | 20 | −3.7 |
| hsa-miR-425-5p | 15 | −4.8 |
| hsa-miR-125b-2-3p | 29 | 4.7 |
| hsa-miR-145-5p | 97 | −5.9 |
| Sample | tRNAs | Percentage of Total Reads Mapped to tRNAs | Sample | tRNAs | Percentage of Total Reads Mapped to tRNAs |
|---|---|---|---|---|---|
| ASCs | Gly-GCC | 34.59% | BMSCs | Gly-GCC | 26.99% |
| Val-AAC | 15.97% | Val-CAC | 8.46% | ||
| Val-CAC | 15.93% | Val-AAC | 7.41% | ||
| ASC-Evs | Gly-GCC | 54.59% | BMSC-Evs | Val-CAC | 43.17% |
| Glu-CTC | 16.14% | Val-AAC | 33.07% | ||
| Gly-CCC | 16.09% | Gly-GCC | 13.07% |
| Sample | tRNAs | BaseMean | Log2FoldChange | Note |
|---|---|---|---|---|
| BMSC-EVs | Ser-ACT | 15 | 5.4 | fragment |
| Ser-GCT | 753 | 2.0 | fragment | |
| Sup-TTA | 35 | 3.9 | fragment | |
| Phe-GAA | 826 | 2.2 | fragment | |
| Ile-AAT | 545 | 2.8 | fragment | |
| Lys-TTT | 4439 | 1.4 | fragment | |
| Gly-CCC | 236570 | −2.9 | halve | |
| Leu-TAG | 393 | 1.9 | fragment | |
| Gly-GCC | 804325 | −2.8 | halve | |
| His-GTG | 22356 | −2.5 | halve | |
| Thr-CGT | 234 | 2.1 | fragment | |
| ASC-EVs | Arg-CCG | 204 | −3.5 | halve |
| Phe-GAA | 826 | −3.0 | fragment | |
| Asn-GTT | 147 | 2.8 | fragment | |
| Ile-AAT | 545 | 2.6 | fragment | |
| Ala-TGC | 1188 | 2.7 | fragment |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Y.; Chang, C.; Su, J.; Venø, M.T.; Kjems, J. Osteoblastogenesis Alters Small RNA Profiles in EVs Derived from Bone Marrow Stem Cells (BMSCs) and Adipose Stem Cells (ASCs). Biomedicines 2020, 8, 387. https://doi.org/10.3390/biomedicines8100387
Yan Y, Chang C, Su J, Venø MT, Kjems J. Osteoblastogenesis Alters Small RNA Profiles in EVs Derived from Bone Marrow Stem Cells (BMSCs) and Adipose Stem Cells (ASCs). Biomedicines. 2020; 8(10):387. https://doi.org/10.3390/biomedicines8100387
Chicago/Turabian StyleYan, Yan, Clare Chang, Junyi Su, Morten T. Venø, and Jørgen Kjems. 2020. "Osteoblastogenesis Alters Small RNA Profiles in EVs Derived from Bone Marrow Stem Cells (BMSCs) and Adipose Stem Cells (ASCs)" Biomedicines 8, no. 10: 387. https://doi.org/10.3390/biomedicines8100387
APA StyleYan, Y., Chang, C., Su, J., Venø, M. T., & Kjems, J. (2020). Osteoblastogenesis Alters Small RNA Profiles in EVs Derived from Bone Marrow Stem Cells (BMSCs) and Adipose Stem Cells (ASCs). Biomedicines, 8(10), 387. https://doi.org/10.3390/biomedicines8100387