
Impact of Doxycycline Addition on Activated Sludge Microflora and Microbial Communities
by
 , ,
, ,
Hayet Djelal
1,*,
Djouza Haddouche
1,
Megane Lebreton
2,
Vanessa Barros
3,
Cristian Villegas
4 and
Patrick Dabert
2 1
Unilasalle-Ecole des Métiers de l’Environnement, CYCLANN, Campus de Ker Lann, 35170 Bruz, France
2
INRAE, UR1466 OPAALE, 17 Avenue de Cucillé, 35044 Rennes, France
3
Institute of Mathematics and Statistics, Federal University of Bahia, Av. Adhemar de Barros s/n, Ondina, Salvador 40170-110, BA, Brazil
4
Department of Exact Sciences, University of São Paulo, Piracicaba 13418-900, SP, Brazil
*
Author to whom correspondence should be addressed.
Processes 2024, 12(2), 350; https://doi.org/10.3390/pr12020350
Submission received: 11 January 2024
/
Revised: 30 January 2024
/
Accepted: 2 February 2024
/
Published: 7 February 2024
(This article belongs to the Section Environmental and Green Processes)
Abstract
:Municipal wastewater treatment plants (WWTPs) are exposed to high concentrations of micropollutants that can impact conventional activated sludge treatment. The consequences of this include failure to meet discharge standards and the disintegration of flocs, leading to poor sludge settleability. This lab-scale study focuses on the influence of doxycycline, an antibiotic widely used against human and animal diseases, on protozoa, metazoa, and bacterial communities under sludge growing conditions. Doxycycline was added to the mixed liquor of a communal WWTP up to 0, 100, 200, and 400 mg of doxycycline L−1 and incubated in batch conditions for 23 days. The regular addition of nutrient and carbon sources was preformed every 2 days to prevent sludge starvation. Sludge growth, conductivity, and settleability were measured and compared to sludge microbial community structure, determined by microscopic observations and high-throughput 16S rDNA sequencing. The high doxycycline concentration negatively impacted settleability and correlated with a decrease in bacterial diversity and floc disintegration. The addition of doxycycline promoted the enrichment of Proteobacteria Brevundimonas sp., Luteibacter anthropi, and the Bacteroidetes Chryseobacterium massoliae. These species are known to be resistant to a wide spectrum of antibiotics, including tetracyclines. A study of a larger scale may be conducted based on this study’ results.
1. Introduction
Antibiotics are largely consumed by humans and animals and are discharged into the environment because of their low degradability and metabolization by the user. A high proportion of the antibiotics ends up in wastewater from feces and urine [1]. Doxycycline (DC), a semisynthetic antibiotic of the tetracycline class, is efficacious against Gram-negative and Gram-positive bacteria. DC was developed in the early 1960s and was the first broad-spectrum antibiotic taken daily [2]. Doxycycline is effective against intracellular bacteria, and its mode of action involves blocking the synthesis of bacterial proteins by reversibly binding to the 30S ribosomal subunit and preventing the association of aminoacyl-tRNA with the bacterial ribosome [2]. Doxycycline is widely used to treat reproductive, urinary, intestinal, respiratory infections, and teenage acne. More recently, doxycycline was used against SARS-CoV-2 (COVID-19) in therapies as an alternative to azithromycin [3,4]. Doxycycline is also used as a treatment for melanoma, a type of skin cancer [5]. This antibiotic is also utilized in veterinary health [6]. Thus, DC is widely used and discharged in wastewater, and DC in wastewater can be transferred to surface and underground waters due to its solubility (630 mg L−1 at 25 °C) [7].
The activated sludge (AS) process is the most used biological treatment in the world for both domestic and industrial wastewater treatment plants (WWTPs) [8]. In this process, heterotrophic aerobic microorganisms degrade wastewater organic matter into carbon dioxide, water, and minerals. Mixed liquor (ML) refers to a mixture made of raw wastewater and microorganisms. After treatment, this mixed liquor is separated by gravity settling to release treated water and collect the sludge. On sites, the control of the wastewater treatment process is performed mainly by monitoring the discharged water quality, measuring the mixed liquor suspended solids (MLSS) content and sludge settleability (measured using the sludge volume index (SVI)), and sometimes by microscopically observing the sludge [9]. In the existing literature, however, analyses of microbial communities in activated sludge mainly focus on bacteria, and the roles of other microbial groups have often been neglected [10].
Actually, AS is a complex ecosystem composed of a high biodiversity of bacteria, archaea, fungi, protozoans (10–200 µm), and metazoans (50 µm–3 mm), and of several non-biological compounds that contain organic and inorganic particles [11]. Its bacterial community structures can be highly variable over one year. The main factors influencing variability are the temperature, the organic matter composition and load, and the presence of micropollutants such as hormones or pharmaceutical compounds [12]. Organic micropollutants, largely present in wastewater, can exert strong selection pressure on the AS’ microbial community. The presence of pharmaceutical compounds can change the AS’ microbial community composition, leading to the inhibition of its activity [13,14].
In a well-maintained aeration tank, the bacteria, archaea, and fungi are primarily found as flocs, which are mainly aggregations of microorganisms found around the natural and synthetic flocculent materials of activated sludge. Some of them remain free in wastewater, along with the protozoa and metazoa. Flocs have an open porous structure and can withstand water’s movement forces during the aeration of the tanks; their size varies between 10 µm and 1 mm [15]. The formation of flocs is possible thanks to the extracellular polymeric substances (EPSs) produced by several microbial groups. Variations in the composition of EPSs during the biotreatment of the wastewater can affect the sludge floc size, thereby affecting the flocculation and settling properties of activated sludge and the quality of the water rejected in the environment [16].
In the AS ecosystem, some microbial groups are dominant while others are in low abundance. It is usually assumed that the dominant bacterial groups (along with some archaea) play the primary role of degrading chemical compounds during wastewater treatment [17], while bacterial groups of lower abundance act as a seed bank of organisms that grow to transient abundance under specific conditions (for example, the degradation of micropollutants) [18]. Ciliates are the most abundant protozoa in AS and constitute approximately 5% of AS dry mass. They play important roles in the functioning of AS by reducing dispersed bacteria and enhancing flocculation [19,20]. There is a significant positive correlation between the sludge volume index (SVI) and the free-swimming ciliates, so it represents a good indicator of the settlement conditions of sludge [21]. All the ciliate species have an aerobic metabolism. The structure of ciliates is quite sensitive to changes in physico-chemical parameters, mainly to organic loading variation [22]. The rotifers, metazoan organisms naturally present in AS, ingest filamentous bacteria and contribute to improving the settling properties of the sludge [11]. The abundance of protozoa and metazoan populations in WWTPs is not influenced by fluctuations in temperature [22]. Thus, the optimal functioning of AS processes relies on the installation and resilience of a well-balanced microbial ecosystem.
For years and still in WWTPs, the identification and characterization of AS microbial communities has been carried out by microscopy inspections, and quantitative image analysis (QIA). This method has been identified as a quick method for assessing AS content and structure. In view of this, the authors of [23] correlated QIA information and the main parameters of treatment efficiency. In research, however, molecular tools such as high-throughput 16S rDNA sequencing have become a standard for bacterial community characterization. Rouibah et al. [24], through using this method, identified bacteria isolated from hospital effluent.
One of the main drawbacks of wastewater treatment plants is the poor settleability of their sludge, resulting in additional costs to achieve and comply with regulations. It would be interesting to check if the presence of a micropollutant is one of the causes of this drawback. Precisely, the aim of this study was to highlight the evolution of protozoa, metazoa, and bacteria in the presence of an antibiotic while examining its impact on the poor settleability of sludge. The biodegradation of doxycycline by activated sludge is not addressed in this work, as it has already been investigated in a previous study [25]. This study aimed to accomplish two primary goals: (i) to assess the sedimentation characteristics of activated sludge in the presence of doxycycline, a commonly used micropollutant, and (ii) to investigate alterations in the microflora and bacterial community composition of activated sludge through microscopic observations and 16S rDNA analysis. We systematically varied the doxycycline concentration and the duration of culture, establishing correlations with key parameters such as mixed liquor suspended solid (MLSS) content, conductivity, and sludge volume index (SVI). These tests were conducted at elevated concentrations, making them relevant to pharmaceutical effluent treatment or wastewater in post-care centers. To the best of our knowledge, very few studies have simultaneously examined microflora and microorganisms within activated sludge in the presence of antibiotics, rendering this study particularly interesting.
2. Materials and Methods
2.1. Chemicals
Doxycycline hyclate (DC) (C22H24N2O8·HCl·0.5H2O·0.5C2H6O, 99%) was obtained from Sigma-Aldrich (Saint-Quentin-Fallavier, France). All other chemicals used were obtained from VWR (purity grade > 97%).
2.2. Activated Sludge Samples
Mixed liquor (ML) was taken from the aerobic tank of the wastewater treatment plant of Beaurade, Rennes, France, which receives predominantly domestic wastewater. This plant uses an anaerobic–anoxic–oxic (A2/O) process and has a total treatment capacity of 360,000 equivalent inhabitants. Mixed liquor was transported to the laboratory within 20 min and immediately placed in aeration and agitation conditions at room temperature. Some analyses were conducted immediately to ascertain pH, conductivity, mixed liquor suspended solids (MLSS), and sludge volume upon decantation for 30 min (DV30) to calculate the sludge volume index (SVI).
2.3. Experimental Procedures
All tests were conducted in Fed-batch and perfectly shaken media. Experiments were carried out in 2 L beakers at a working volume of 1 L consisting of 990 mL of mixed liquor and 10 mL of nutritive solution in mg L−1: K2HPO4, 208; KH2PO4, 85; Na2HPO4·2H2O, 154.5; MgSO4·7H2O, 22.6; CaCl2, 27.6; FeCl3·6H2O, 260.0; NH4Cl, 74; and 2 g of glucose as carbon source. The composition of the mineral medium was based on our previous study [25]. The essays consisted of four different runs: the addition of 0, 100, 200, and 400 mg L−1 of doxycycline. In order to not starve the microorganisms and change their behavior, it was decided to work without the limitation of a carbon substrate. Every two days, 10 mL of the nutritive solution and 2 g of glucose were added to each beaker. Experiments were conducted at ambient temperature, and the contents of each beaker were magnetically stirred. Aeration was performed by means of an aquarium pump connected to a diffuser, allowing for the production of fine air bubbles to ensure efficient oxygen transfer. The experiments were carried out in, at least, duplicate, and the tests were performed for 23 days.
2.4. Analytical Methods
For the MLSS content, SVI, and the conductivity analysis, the period of investigation was between 0 and 23 days. Sampling for microbial community analysis was performed on days 0, 7, 14, and 23. MLSS and SVI were measured at the beginning and the end of each experiment. For conductivity (WTW COND 3110 SET1), the measurements were taken directly from the mixed liquor. For other measures, samples of 5 mL were periodically taken. Cell growth was monitored by measuring the turbidity (HACH 2100Q) and mixed liquor suspended solids (MLSS) content, which was obtained by centrifuging 5 mL of cell suspension at 1792× g and 4 °C for 15 min, after which the pellet was dried at 105 °C (VWR DRY-Line) for 48 h. The sludge volume index (SVI) was determined based on the sludge volume upon decantation for 30 min (DV30) in a 1 L cylinder and combined with MLLS (SVI = DV30 × MLSS−1). All physico-chemical analyses were performed in duplicate.
2.5. Microbial Community Analysis
2.5.1. Microscopic Examination
The biomass collected from the aeration tank was characterized by microscopic examination using an Optical microscope (ZEISS Primo Star, Carl Zeiss France S.A.S., Ruel-Malmaison, France) equipped with an image acquisition system (BRESSER MikroCamLab). Activated sludge identification was performed according to the methods of [26,27,28]. Twelve microscopic 10 µL observations were performed for each doxycycline concentration at each time of culture.
2.5.2. Bacterial Quantification
Total bacteria was quantified by real-time qPCR targeting the 16S rRNA gene using primers 1055F (5′-ATGGCTGTCGTCAGCT-3′) and 1392r (5′-ACGGGCGGTGTGTAC-3′) [29]. PCR amplifications were carried out in a 25 µL final working volume using the Takyon No ROX SYBR 2X MasterMix blue dTTP (Kaneka Eurogentec S.A., Seraing, Belgium) and the CFX96 thermal cycler (Bio-Rad, Marnes-la-Coquette, France) according to the supplier’s instructions. The amplification reaction was an initial denaturation of 95 °C for 10 min, followed by 45 cycles of 95 °C for 30 s, 60 °C for 50 s, and 72 °C for 30 s. Amplifications were carried out in triplicate on 10- and 100-fold diluted DNA. A melting curve analysis was performed to check the specificity of the amplification and primer dimer formation. The gene copy numbers were calculated by comparison with DNA standards of known concentrations amplified in the same runs and expressed as gene copy number per g of AS suspended solids or mL.
2.5.3. High-Throughput DNA Sequencing
Two mL of mixed liquor was collected from each beaker and centrifuged at 1792× g at 4 °C for 5 min. The supernatants were discarded, while the pellets were stored at −20 °C. Genomic DNA extraction was performed for each frozen pellet using the Macherey-Nagel NucleoSpin® Soil kit and the lysis buffer SL1 + SX (according to the manufacturer recommendations, Macherey-Nagel SAS, Hoerdt cedex, France). Extracted DNA was eluted in a final water volume of 60 µL. The quality of DNA was verified by electrophoresis on agarose gel (TAE 1X, 0.7% agarose w/v, 120 V, 40 min). Its concentration was measured using a biophotometer (Eppendorf BioPhotometer® D30, Eppendorf AG, Hamburg, Germany). The purified DNA was stored at −20 °C until sequencing.
The overall method used for high-throughput DNA sequencing was detailed in [30]. Briefly, the bacterial and archaeal V4-V5 regions of the 16S rRNA genes were amplified by PCR using fusion primers derived from 515F (5′-CTGYCAGCMGCCGCGGTA-3′) and 928R (5′-CCCCGYCAATTCMTTTRAGT-3′) [31]. The resulting amplicons were sequenced by the INRAE PROSE Team (Antony, France) using the Ion PGM Hi-Q View OT2 Kit and Ion PGM Hi-Q View Sequencing kit and an Ion Torrent Personal Genome Machine (Life Technologies, Carlsbad, CA USA). The 1,176,809 sequence reads (of approximately 420 base pairs) were analyzed using the Galaxy portal of the Migale bioinformatics platform of INRAE (Migale bioinformatics Facility, INRAE, Jouy-en-Josas, France) and the FROGS software pipeline (“Find Rapidly OTU with Galaxy Solution”) [32]. The resulting 366,083 correct sequences, obtained from all samples, were clustered in Operational Taxonomic Units (OTUs) based on whether they had more than 97% similarity within an OTU. An analysis of the taxonomic affiliation of the OTUs was carried out by both BLAST (NCBI) and Ribosomal Database Project II database sequence alignment [33,34]. OTU abundance analysis and an analysis of the diversity index of the microbial communities were performed using Easy16S, a shiny web interface [35] of the phyloseq package of R software version 4.3.1 [36].
2.6. Statistical Analysis
In order to verify the relationship between the DC concentration, cultivation time, and the MLSS content, which reflected the microbial community growth during the study, a two-way ANOVA was carried out using R software (Version 4.1.2). A two-way analysis of variance was used since we suspected some kind of interaction between the levels of two experimental factors. The independent variables, also called factors, were DC concentration and cultivation time, and the dependent variable was MLSS.
The null hypothesis (H0) is the claim assumed to be true in a statistical test. Once the hypotheses are formed, sample data are collected, and statistics are calculated according to the parameters detailed in the hypotheses. For every hypothesis test, a significance level of 0.05 was used to validate the result of the test. If the p-value (probability value, used to quantify the amount of evidence against the null hypothesis) is greater than or equal to the significance level, then the null hypothesis was retained.
3. Results and Discussion
3.1. Mixed Liquor Characterization
The characteristics of the mixed liquor at T0 are presented in Table 1. The MLSS values are in accordance with the average value usually found in aeration tanks from communal WWTPs (4 < MLSS < 6 g L−1) [9]. The value of the sludge volume index (SVI) shows that the decantability was sufficient to separate the mixed liquor from the activated sludge. Our microscopic observations of the mixed liquor just after sampling showed numerous flocs, metazoa, and protozoa (Figure 1). These communities are commonly present in activated sludge from communal WWTPs and indicate good-quality treated effluent [26,27].
3.2. Impact of the Doxycycline Concentration on Microbial Growth (MLSS), Conductivity and SVI Parameters after 23 Days of Cultivation
Four concentrations of doxycycline (0, 100, 200, and 400 mg L−1 of doxycycline) were tested in aerobic conditions simulating the activated sludge activity in the wastewater aerobic tank, as explained in the Materials and Methods section. A carbon source and mineral solution were added every two days in mixed liquor to prevent starvation and overcome the impacts of nutrient depletion. Tests were carried out for 23 days.
In all the culture conditions and at different sampling times (0, 7, 14, and 23 days), the presence of Holotriche, Spirotriche, Peritriche, Rotifera, and Nematodes was observed. However, our microscopic observations of the activated sludge were unable to determine the trend of the abundance of the protozoa and metazoa in experimental conditions.
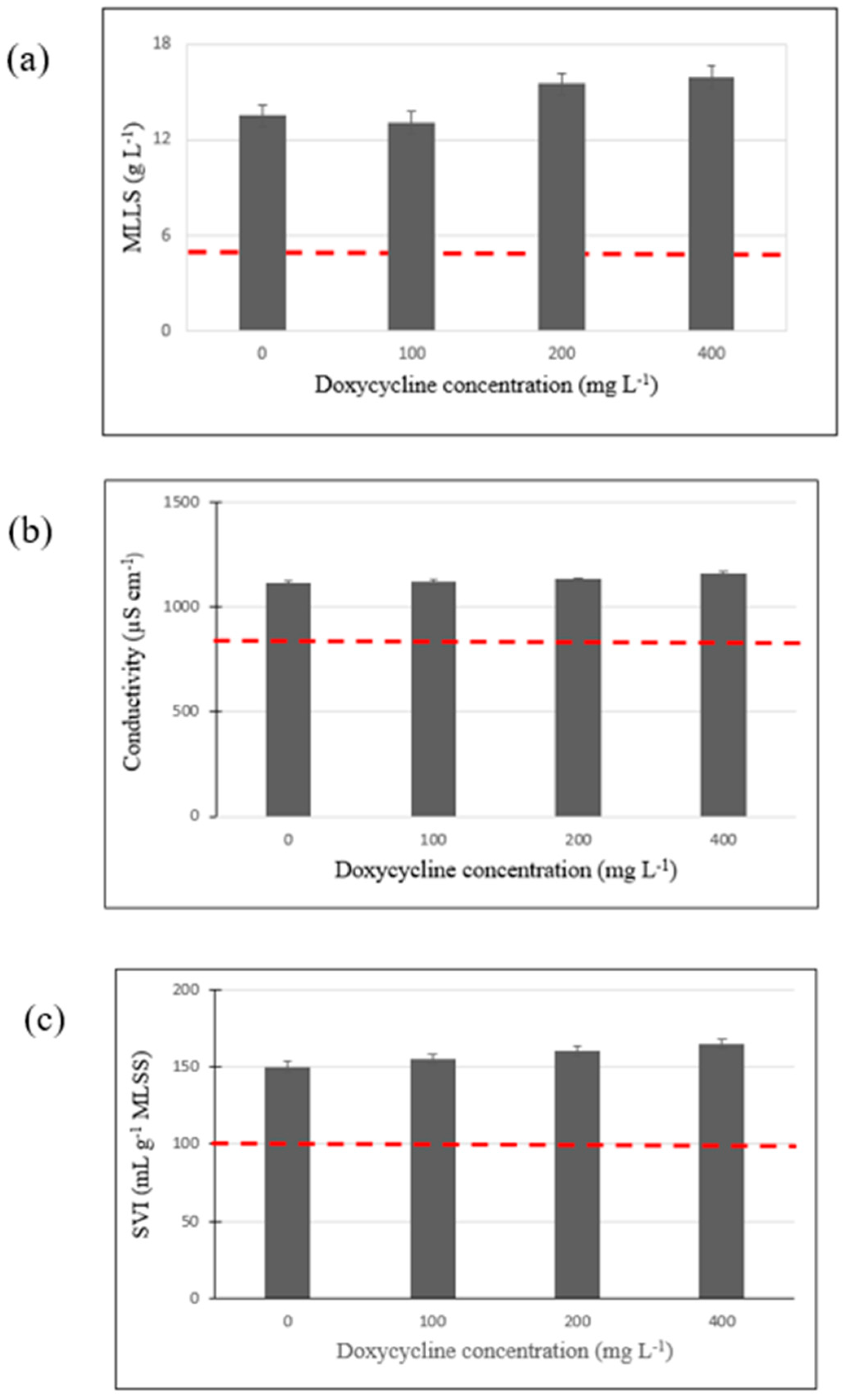
The mixed liquor suspended solids (MLSS) content, measured to quantify the growth of the AS biomass after 23 days, increased by 1.75, 1.67, 2.16, and 2.24, respectively compared to the initial mixed liquor (Figure 2a). These increases are probably due to the addition of glucose during the experiment. These results show that doxycycline did not completely inhibit the growth of all of the microbial populations in the activated sludge, as previously observed by Djelal et al. in 2020 [25]. The sludge supplemented with 200 and 400 mg L−1 of doxycycline showed the highest sludge growth.
An increase of about 31% in the conductivity was observed for all conditions, certainly caused by the accumulation of mineral nutrients (Figure 2b) which did not affect the growth of microbial community. On the 23rd day, the pH values for the various tests ranged from 7.1 to 7.6, closely resembling the initial pH. Consequently, it can be concluded that pH did not exert any significant influence on the experiments.
The SVI value increased by about 50% for all conditions, indicating the deterioration of the settling performance of the activated sludge (Figure 2c). An SVI < 100 mL g−1 indicates well-flocculated sludge. Poor settling is generally due to the disintegration of the flocs or/and the presence of filamentous bacteria. A high hydraulic retention time (inhere 23 days) favors the growth of certain filamentous bacteria, thereby changing the bacterial community and affecting sludge decantation [37].
Figure 3 shows the settling ability and our microscopic observations of the mixed liquor on day 0 and after 23 days of culture in the absence of doxycycline addition. An abundant active flora of free-swimming ciliates had developed, and the flocs appeared to be disintegrated (deflocculation), which explains the poor settling of the sludge and the bulking of the mixed liquor. It has been observed that in the presence of micropollutants, bacteria produce fewer EPSs, resulting in the disintegration of flocs [38]. However, it is not easy to explain the cellular mechanisms underlying this [39]. For Dos Santos et al. [21], an increase in SVI is in accordance with the rise of free-swimming ciliates, influencing the settleability of sludge and the global performance of the WWTP. In our experiment, the addition of 2 g L−1 of glucose every two days to simulate high-organic-matter loads induced excessive microflora and microbial community growth, as well an increase in SVI. These results agree with those of [40]. Optimizing the correlation between SVI and HRT is important for maintaining efficient wastewater treatment. Operators often adjust biological treatment parameters such as HRT to achieve the desired sludge settling characteristics indicated by SVI values. Monitoring both parameters can help in identifying and addressing issues related to biological activity and sludge settling in wastewater treatment plants. For high HRT, the contact time between microorganisms and the organic matter in the wastewater are important, allowing for better microbial activity. However, extended residence times can lead to the accumulation of degradation compounds that may inhibit the growth or activity of biodegradation.
3.3. Impact of Doxycycline on the Bacterial Communities of Activated Sludge Samples
The bacterial composition of the activated sludge was assessed by 16S rDNA targeted qPCR and high-throughput sequencing on days 0, 7, 14, and 23 for all doxycycline concentrations. The analysis was replicated for the initial sludge on day 0 and for the sludge incubated with 400 mg L−1 of doxycycline. The qPCR showed a slight increase in bacterial concentration after day 7, but this was not statistically significant (Figure S1). This suggests that there was no real variation in the bacterial concentration that remained at about 2.5 ± 1.3 × 1011 copies of 16S rRNA gene per mL of activated sludge, regardless of the sampling time or the doxycycline concentration. The 366,083 sequences obtained by high-throughput 16S rDNA sequencing were distributed at a mean of 21,534 ± 5593 sequences per sample. Sequence clustering resulted in a mean of 440 ± 173 Operational Taxonomy Units (OTUs) per sludge sample. The coverage of sequencing ranged from 90% of the expected diversity for the sludge on day 0 (i.e., relatively good coverage) to as low as about 60% for some sludges, but there were no evident relations between sequence coverage and sampling time or antibiotic concentration.
The characterization of the microbial populations during all incubations showed a high diversity of 27 microbial phyla but a constant dominance of the Bacteroidetes and the Proteobacteria. Whether doxycycline is present or not, the residence time has an impact on the profile of the microbial population, leading to a loss of biodiversity (Figure 4). The addition of glucose alone sustained the enrichment of these two phyla up to 53% and 42%, respectively, of the total sequences on day 23. The addition of both glucose and doxycycline, regardless of concentration, promoted a strong enrichment in Proteobacteria for the first 14 days of incubation for up to 70 to 90% of the total sequences. However, the relative abundance of Bacteroidetes in the sludge increased again at the end of the experiment for all doxycycline concentrations except 100 mg L−1.
Our comparison of the sludge bacterial community structures shows a strong reduction in bacterial diversity after the addition of glucose (Figure 5). The number of OTUs and the Shannon diversity index (SDI) of the initial sludge on day 0 dropped from 560 OTUs and a SDI of 5.45 to as low as 193 OTUs and a SDI of 2.51 after 23 days of incubation without doxycycline. This reduction is much stronger than the analytical difference observed for the initial sludge on day 0 (S-d0 and S′-d0). This drop in diversity probably results from the death of several bacterial species in the new environment represented by the laboratory device compared to the wastewater treatment plant. However, since the experiments were performed in batch mode and the total bacterial concentration remained similar in all conditions (as measured by qPCR), this bacterial death was probably balanced by an enrichment in heterotrophic species because of the addition of glucose. Interestingly, a different behavior was observed upon doxycycline addition, suggesting that the antibiotic had a stronger effect than glucose addition on the sludge’s bacterial community. When added at 400 mg L−1, the impact of doxycycline on bacterial diversity was visible as early as day 7 due to a reduction in the bacterial richness to 200 to 300 OTUs and a drop in the SDI to about 2.4. Thereafter, the bacterial richness remained similar until the end of the experiment, while at the same time, the SDI rose to about 4. The Shannon diversity index is the product of the richness (in log) by the regularity. This suggests that the antibiotic affected all the community without the specific enrichment of a few species, as can be expected from a broad-spectrum antibiotic. When doxycycline was added at lower concentrations (100 and 200 mg L−1), a strong impact was observed only after 14 days of incubation.
Proteobacteria and Bacteroidetes phyla have been reported as the prevailing classes in activated sludge from communal WWTPs [13,41,42,43]. Proteobacteria play a significant and broad role in organic and nutrient removal [37]. As reviewed by Cydzik-Kwiatkowska and Zielinska [44], the amount and kind of antibiotics in the wastewater decides the bacterial composition of the WWTP effluent. Liu et al. [45] observed that aerobic starvation had important effects on the succession of microbial community, and the Proteobacteria and Bacteroidetes phyla declined while the Firmicutes increased. In this study, we worked in abundance conditions and observed the opposite.
Analyzing the sludge community structures at the family level and in an in-depth manner revealed that all the sludges had different behaviors; nevertheless, there were two constants (Figure S2): the strong decrease in sub-dominant populations (i.e., the ones containing less than 1% of the total sequences), and the enrichment (sometimes transient) of two to three families of microorganisms that differ from one incubation to another. To better target the microbial populations that affected the sludge dynamics, a principal component analysis (PCA) was performed on the relative abundance of all the species of all sludges (Figure 6). PCA allows for a visualization of the variability between all samples (here, the sludge samples) and of the vectors (here, the microbial populations) that drive this variability. As shown in Figure 6, the PCA explained only 43% of the community structures differences. However, there is a trend along the horizontal axis—shown below via the communities that submitted to increasing concentrations of doxycycline ranging from the left to the upper right of the graph—and, possibly, a downward vertical trend of increasing time and/or glucose addition.
The microbial populations underlined by the PCA and targeting the initial sludge on day 0 were identified (Table S1) as an uncultured Saprospiraceae (C-26), Nitrospira defluvii (C-20), and Tolumonas auensis (C-14). The first two microorganisms are common in activated sludge [44]. The latter is common in sediments [46]. The three populations targeting down the figure were identified as Gluconacetobacter (C-3) and two Xanthomonadales, either Dyella sp. or Frateuria aurantia (C-9), and an uncultured Xanthomonadaceae (C-2). Several species of Gluconacetobacter can grow under very high glucose concentrations (up to 30% glucose) [47]. These probably resulted from the repeated addition of glucose during the experiment. The Xanthomonadales are common microbial groups in WWTPs and soil [44]. Finally, nine populations target the communities submitted to high doxycycline concentrations. They all correspond to widespread species found in soil, sediments, water, etc. The three most important were identified as Brevundimonas sp. (C-5), Luteibacter anthropi (C-6), and Chryseobacterium massoliae (C-4). Brevundimonas is a heterotrophic bacterium able to grow on several sugars. Chryseobacterium contains several species known to be resistant to a wide spectrum of antibiotics, including tetracyclines [48]. Along with Luteibacter (herein C-6), Dyella (C-9), several members of the Burkholderiaceae (C-11), and several other genera isolated from soil, it possesses tet genes involved in tetracycline resistance [49]. Furthermore, some species of Chryseobacterium have been isolated as amoebae-resisting bacteria by Greub et al. [50]. Finally, members of the Enterobacteriacea, such as Enterobacter and Citrobacter (identified here as C-16), often exhibit antibiotic resistance. The properties of the species underlined by PCA are thus in agreement with the hypothesis of a selection of heterotrophic microorganisms able to resist antibiotics. The addition of doxycycline promoted a strong reduction in bacterial diversity with the enrichment in the Proteobacteria Brevundimonas sp. and Luteibacter anthropi and the Bacteroidetes Chryseobacterium massoliae. These species are known to be resistant to a wide spectrum of antibiotics, including tetracyclines.
High concentrations of antibiotics can apply a high amount of pressure on microbial communities and allow for the emergence and dissemination of antibiotic resistance genes (ARGs) [1,51]. WWTPs have been found to be one of the major sources of ARGs being released into environment [52]. More specifically, tetracycline-resistant bacteria (TRB) and tetracycline-resistant genes (TRGs) have been frequently detected in activated sludge [53]. A study conducted in batch conditions on the impact of chloroxylenol, an antibacterial agent, on the granular sludge structure showed an increase in antibiotic resistance genes (ARGs) [38]. The same response was observed with triclosan, a commonly used antimicrobial agent [39]. The bacterial communities in treated wastewater could have an increased capacity for facilitating the spread of doxycycline resistance genes (ARGs) and, likely, the biodegradation of doxycycline, being better adapted to cope with the effects of antibiotics [54]. Our results are in agreement with those of Djelal et al. [25], who studied the doxycycline removal efficiency (60%) of activated sludge after 14 days in batch conditions. Dai et al. suggest using triclosan-degrading bacteria identified as bioaugmentation strains in practical engineering [55]. Abou Dalle et al. successfully implemented this approach by isolating two metronidazole-resistant bacteria from activated sludge and then adding them to the activated sludge (bioaugmentation process). This led to a significant reduction in treatment times [56].
3.4. ANOVA of Microbial Community Growth
The aim of this section is to describe how we used the ANOVA database in Table 2 to conduct a statistical test (ANOVA) to verify the effect of cultivation time and doxycycline concentration on microbial community growth (MLSS).
Before performing an ANOVA, we must check its assumptions. An ANOVA assumes that data are normally distributed. An ANOVA also assumes homogeneity of variance, which means that the variance among the groups should be approximately equal. According to the Shapiro–Wilk normality test at 5% of significance, residuals can be considered normal, but Bartlett’s test indicated the existence of heteroscedasticity of variance. We then applied the Box–Cox transformation to the response variable as a corrective measure, which resulted in a better model.
The next step was to test for interaction effects between the following factors: DC concentration and cultivation time. Interaction effects represent the combined effects of factors on the dependent measure (MLSS). When interaction effects are present, it means that interpretation of the main effects is incomplete or misleading. A two-way ANOVA test indicated that there was no interaction between the above factors at 5% significance. In fact, if we inspect Table 3 (see DC concentration x day (mg L−1·day)), we notice a p-value of 0.600 (greater than 0.05), meaning that the following null hypothesis was verified: there is no interaction between factors. So, it was appropriate to carry out further tests concerning the presence of the main effects. The results are shown in Table 3.
The first null hypothesis stated that the DC concentration factor has no effect on the means of MLSS. The F-test showed a p-value of 0.607 (see DC concentration (mg L−1) in Table 3), so we failed to reject this null hypothesis. The second null hypothesis stated that the cultivation time factor has no effect on the means of MLSS. The p-value for this test was 0.078 (see Time of cultivation (day) in Table 3), so, once again, we failed to reject this null hypothesis.
4. Conclusions
This study evaluated the impact of the antibiotic doxycycline on activated sludge settleability and microbial community composition in batch conditions using abundant mineral and organic materials. Doxycycline concentrations up to 400 mg L−1 did not affect the apparent global biomass growth. However, the SVI increased after 23 days of culture regardless of the doxycycline concentration, which led to a bad decantation of the sludge. This could be explained by the deterioration of the flocs due to the high HTR. Based on our two-way analysis of variance (ANOVA), HTR and doxycycline concentration had no effect on microbial community growth. For a better understanding of the floc destabilization mechanisms in the presence of doxycycline, it would be advisable to measure the EPS levels in the various conditions. The results obtained indicate that microorganisms can adapt to an environment rich in doxycycline. This has interesting implications for the potential use of activated sludge in the treatment of micropollutants. However, to prevent the disintegration of flocs and thus maintain optimal settling, it would be necessary to reduce hydraulic retention times and organic load. The tests were conducted in batch culture, and for high HTR, the accumulation of degradation compounds may inhibit the activity of activated sludge. So, it would be interesting to conduct experiments under the actual operating conditions of wastewater treatment plants, namely in continuous culture with sludge recirculation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pr12020350/s1.
Author Contributions
H.D.: conceptualization, methodology, supervision, writing; P.D. and M.L.: worked on bacterial community characterization, methodology, analysis, writing; V.B. and C.V.: worked on the ANOVA analysis, methodology, writing; D.H.: essays and analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors would like to extend their deep gratitude to the staff of the WWTP of Rennes, France, for allowing us to collect mixed liquor samples. The authors are grateful to the INRAE MIGALE bioinformatics facility (MIGALE, INRAE, 2018) for providing computing and storage resources.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhao, J.; Zhang, J.; Zhang, D.; Hu, Z.; Sun, Y. Effect of Emerging Pollutant Fluoxetine on the Excess Sludge Anaerobic Digestion. Sci. Total Environ. 2021, 752, 141932. [Google Scholar] [CrossRef] [PubMed]
- Holmes, N.E.; Charles, G.P. Safety and efficacy review of doxycycline. Clin. Med. Ther. 2009, 1, CMT-S2035. [Google Scholar] [CrossRef]
- Ali, A.S.; ASattar, M.A.; Karim, S.; Kutbi, D.; Aljohani, H.; Bakhshwin, D.; Alsieni, M.; Alkreathy, H.M. Pharmacological Basis for the Potential Role of Azithromycin and Doxycycline in Management of COVID-19. Arab. J. Chem. 2021, 14, 102983. [Google Scholar] [CrossRef]
- Malek, A.E.; Granwehr, B.P.; Kontoyiannis, D.P. Doxycycline as a Potential Partner of COVID-19 Therapies. IDCases 2020, 21, e00864. [Google Scholar] [CrossRef] [PubMed]
- Rok, J.; Rzepka, Z.; Banach, K.; Kowalska, J.; Wrześniok, D. The Assessment of the Phototoxic Action of Chlortetracycline and Doxycycline as a Potential Treatment of Melanotic Melanoma—Biochemical and Molecular Studies on COLO 829 and G-361 Cell Lines. Int. J. Mol. Sci. 2023, 24, 2353. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Wang, Y.; Liu, L.; Zou, Y.; Liao, X.; Liang, J.; Wu, Y. The Excretion and Environmental Effects of Amoxicillin, Ciprofloxacin, and Doxycycline Residues in Layer Chicken Manure. Poult. Sci. 2016, 95, 1033–1041. [Google Scholar] [CrossRef]
- Aniagor, C.O.; Igwegbe, C.A.; Ighalo, J.O.; Oba, S.N. Adsorption of Doxycycline from Aqueous Media: A Review. J. Mol. Liq. 2021, 334, 116124. [Google Scholar] [CrossRef]
- Wei, Y.; Van Houten, R.T.; Borger, A.R.; Eikelboom, D.H.; Fan, Y. Minimization of Excess Sludge Production for Biological Wastewater Treatment. Water Res. 2003, 37, 4453–4467. [Google Scholar] [CrossRef]
- Sadr, S.M.K.; Saroj, D.P. 14—Membrane Technologies for Municipal Wastewater Treatment. In Advances in Membrane Technologies for Water Treatment; Basile, A., Cassano, A., Rastogi, N.K., Eds.; Woodhead Publishing Series in Energy; Woodhead Publishing: Oxford, UK, 2015; pp. 443–463. ISBN 978-1-78242-121-4. [Google Scholar]
- Maza-Márquez, P.; Vílchez-Vargas, R.; González-Martínez, A.; González-López, J.; Rodelas, B. Assessing the Abundance of Fungal Populations in a Full-Scale Membrane Bioreactor (MBR) Treating Urban Wastewater by Using Quantitative PCR (qPCR). J. Environ. Manag. 2018, 223, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fiałkowska, E.; Pajdak-Stós, A. The Role of Lecane Rotifers in Activated Sludge Bulking Control. Water Res. 2008, 42, 2483–2490. [Google Scholar] [CrossRef] [PubMed]
- Leiviskä, T.; Risteelä, S. Analysis of Pharmaceuticals, Hormones and Bacterial Communities in a Municipal Wastewater Treatment Plant—Comparison of Parallel Full-Scale Membrane Bioreactor and Activated Sludge Systems. Environ. Pollut. 2022, 292, 118433. [Google Scholar] [CrossRef]
- Davids, M.; Gudra, D.; Radovica-Spalvina, I.; Fridmanis, D.; Bartkevics, V.; Muter, O. The Effects of Ibuprofen on Activated Sludge: Shift in Bacterial Community Structure and Resistance to Ciprofloxacin. J. Hazard. Mater. 2017, 340, 291–299. [Google Scholar] [CrossRef]
- Mesquita, D.P.; Amaral, A.L.; Ferreira, E.C. Activated Sludge Characterization through Microscopy: A Review on Quantitative Image Analysis and Chemometric Techniques. Anal. Chim. Acta 2013, 802, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Davies, P. The Biological Basis of Wastewater Treatment; Strathkelvin Instruments Ltd.: Glasgow, UK, 2006. [Google Scholar]
- Li, B.; Qiu, Y.; Zhang, J.; Liang, P.; Huang, X. Conjugative Potential of Antibiotic Resistance Plasmids to Activated Sludge Bacteria from Wastewater Treatment Plants. Int. Biodeterior. Biodegrad. 2019, 138, 33–40. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, H.; Tian, Y.; Yu, X.; Li, X. Stochasticity versus Determinism: Microbial Community Assembly Patterns under Specific Conditions in Petrochemical Activated Sludge. J. Hazard. Mater. 2021, 407, 124372. [Google Scholar] [CrossRef] [PubMed]
- Saunders, A.M.; Albertsen, M.; Vollertsen, J.; Nielsen, P.H. The Activated Sludge Ecosystem Contains a Core Community of Abundant Organisms. ISME J. 2016, 10, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Pajdak-Stós, A.; Sobczyk, M.; Fiałkowska, E.; Kocerba-Soroka, W.; Fyda, J. The Effect of Three Different Predatory Ciliate Species on Activated Sludge Microfauna. Eur. J. Protistol. 2017, 58, 87–93. [Google Scholar] [CrossRef]
- Pérez-Uz, B.; Franco, C.; Martín-Cereceda, M.; Arregui, L.; Campos, I.; Serrano, S.; Guinea, A.; Fernández-Galiano, D. Biofilm Characterization of Several Wastewater Treatment Plants with Rotating Biological Contactors in Madrid (Spain). Water Sci. Technol. 1998, 37, 215–218. [Google Scholar] [CrossRef]
- Araújo Dos Santos, L.; Ferreira, V.; Pereira, M.O.; Nicolau, A. Relationship between Protozoan and Metazoan Communities and Operation and Performance Parameters in a Textile Sewage Activated Sludge System. Eur. J. Protistol. 2014, 50, 319–328. [Google Scholar] [CrossRef]
- Martín-Cereceda, M.; Pérez-Uz, B.; Serrano, S.; Guinea, A. Dynamics of Protozoan and Metazoan Communities in a Full Scale Wastewater Treatment Plant by Rotating Biological Contactors. Microbiol. Res. 2001, 156, 225–238. [Google Scholar] [CrossRef]
- Mesquita, D.P.; Amaral, A.L.; Ferreira, E.C. Estimation of Effluent Quality Parameters from an Activated Sludge System Using Quantitative Image Analysis. Chem. Eng. J. 2016, 285, 349–357. [Google Scholar] [CrossRef]
- Rouibah, I.; Hassen, W.; Sallem, O.F.; Khellaf, N.; Hassen, A.; Mansour, H.B. Photocatalytic and Biodegradation Treatments of Paracetamol: Investigation of the in Vivo Toxicity. Environ. Sci. Pollut. Res. 2021, 28, 14530–14545. [Google Scholar] [CrossRef] [PubMed]
- Djelal, H.; Martinez, P.E.; Haddouche, D.; Chabani, M. Assessment of the Biodegradation of Doxycycline by Biostimulation with Addition of Glucose, Phenol or/and Copper. Ecocycles 2020, 6, 25–31. [Google Scholar] [CrossRef]
- Canler, J.P.; Perret, J.M.; Duchène, P.; Cotteux, E. Aide au Diagnostic des Stations D’épuration par L’observation Microscopique des Boues Activées; Editions Cemagref; Antony: Chippenham, UK, 1999. [Google Scholar]
- Foissner, W. Protists as Bioindicators in Activated Sludge: Identification, Ecology and Future Needs. Eur. J. Protistol. 2016, 55, 75–94. [Google Scholar] [CrossRef]
- Pérez-Uz, B.; Arregui, L.; Calvo, P.; Salvadó, H.; Fernández, N.; Rodríguez, E.; Zornoza, A.; Serrano, S. Assessment of Plausible Bioindicators for Plant Performance in Advanced Wastewater Treatment Systems. Water Res. 2010, 44, 5059–5069. [Google Scholar] [CrossRef]
- Ferris, M.J.; Muyzer, G.; Ward, D.M. Denaturing Gradient Gel Electrophoresis Profiles of 16S rRNA-Defined Populations Inhabiting a Hot Spring Microbial Mat Community. Appl. Environ. Microbiol. 1996, 62, 340–346. [Google Scholar] [CrossRef]
- Fisgativa, H.; Tremier, A.; Le Roux, S.; Bureau, C.; Dabert, P. Understanding the Anaerobic Biodegradability of Food Waste: Relationship between the Typological, Biochemical and Microbial Characteristics. J. Environ. Manag. 2017, 188, 95–107. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, P.-Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Haddad, S.M.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287. [Google Scholar] [CrossRef] [PubMed]
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 9 January 2024).
- RDP—Database Commons. Available online: https://ngdc.cncb.ac.cn/databasecommons/database/id/237 (accessed on 9 January 2024).
- MIGALE Bioinformatics Facility (MIGALE, INRAE, 2020). Available online: https://migale.inrae.fr (accessed on 7 July 2022). [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, L.; Xiang, F.; Zhao, L.; Qiao, Z. Activated Sludge Microbial Community and Treatment Performance of Wastewater Treatment Plants in Industrial and Municipal Zones. Int. J. Environ. Res. Public Health 2020, 17, 436. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, J.; Wang, Z.; Cui, Y.; Li, Z.; Wu, Z.; Zhao, Y.; Li, D.; Dai, H. The fate and behavior mechanism of antibiotic resistance genes and microbial communities in flocs, aerobic granular and biofilm sludge under chloroxylenol pressure. J. Hazard. Mater. 2022, 438, 129465. [Google Scholar] [CrossRef]
- Carey, D.E.; McNamara, P.J. The impact of triclosan on the spread of antibiotic resistance in the environment. Front. Microbiol. 2015, 5, 780. [Google Scholar] [CrossRef] [PubMed]
- Fan, N.; Wang, R.; Qi, R.; Gao, Y.; Rossetti, S.; Tandoi, V.; Yang, M. Control Strategy for Filamentous Sludge Bulking: Bench-Scale Test and Full-Scale Application. Chemosphere 2018, 210, 709–716. [Google Scholar] [CrossRef]
- Yu, N.; Zhao, C.; Ma, B.; Li, S.; She, Z.; Guo, L.; Zhang, Q.; Zhao, Y.; Jin, C.; Gao, M. Impact of Ampicillin on the Nitrogen Removal, Microbial Community and Enzymatic Activity of Activated Sludge. Bioresour. Technol. 2019, 272, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.-T.; Yuan, Q.-B.; Yang, J. Insights into the Amplification of Bacterial Resistance to Erythromycin in Activated Sludge. Chemosphere 2015, 136, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Munz, G.; Gualtiero, M.; Salvadori, L.; Claudia, B.; Claudio, L. Process Efficiency and Microbial Monitoring in MBR (Membrane Bioreactor) and CASP (Conventional Activated Sludge Process) Treatment of Tannery Wastewater. Bioresour. Technol. 2008, 99, 8559–8564. [Google Scholar] [CrossRef] [PubMed]
- Cydzik-Kwiatkowska, A.; Zielińska, M. Bacterial Communities in Full-Scale Wastewater Treatment Systems. World J. Microbiol. Biotechnol. 2016, 32, 66. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Peng, Y.; Ma, B.; Ma, L.; Jia, F.; Li, X. Dynamics of Microbial Activities and Community Structures in Activated Sludge under Aerobic Starvation. Bioresour. Technol. 2017, 244, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Romero, C.; Tindall, B.J.; Juttner, F. Tolumonas Auensis Gen. Nov., Sp. Nov., a Toluene-Producing Bacterium from Anoxic Sediments of a Freshwater Lake. Int. J. Syst. Bacteriol. 1996, 46, 183–188. [Google Scholar] [CrossRef]
- Franke, I.H.; Fegan, M.; Hayward, C.; Leonard, G.; Stackebrandt, E.; Sly, L.I. Description of Gluconacetobacter sacchari Sp. Nov., a New Species of Acetic Acid Bacterium Isolated from the Leaf Sheath of Sugar Cane and from the Pink Sugar-Cane Mealy Bug. Int. J. Syst. Evol. Microbiol. 1999, 49, 1681–1693. [Google Scholar] [CrossRef]
- Loch, T.P.; Faisal, M. Emerging Flavobacterial Infections in Fish: A Review. J. Adv. Res. 2015, 6, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, Y.; Hasebe, A.; Nishio, M.; Uchiyama, H. Diversity of Tetracycline Resistance Genes in Bacteria Isolated from Various Agricultural Environments. Microb. Environ. 2007, 22, 44–51. [Google Scholar] [CrossRef]
- Greub, G.; Scola, B.L.; Raoult, D. Amoebae-Resisting Bacteria Isolated from Human Nasal Swabs by Amoebal Coculture. Emerg. Infect. Dis. 2004, 10, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewska, E.; Harnisz, M. Relationship between Modification of Activated Sludge Wastewater Treatment and Changes in Antibiotic Resistance of Bacteria. Sci. Total Environ. 2018, 639, 304–315. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Shen, L.; Chen, Z.; Zhang, Y.; Zhang, C.; Li, Q.; Wang, Y. Production of Polyhydroxyalkanoates by Activated Sludge: Correlation with Extracellular Polymeric Substances and Characteristics of Activated Sludge. J. Chem. Eng. 2019, 361, 219–226. [Google Scholar] [CrossRef]
- Fan, Z.; Yang, S.; Zhu, Q.; Zhu, X. Effects of Different Oxygen Conditions on Pollutants Removal and the Abundances of Tetracycline Resistance Genes in Activated Sludge Systems. Chemosphere 2022, 291, 132681. [Google Scholar] [CrossRef]
- Losada-Carrillo, M.P.; Sevilla, M.; Casado, M.; Piña, B.; Pastor Lópeza, E.; Matamoros, V.; Vila-Costa, M.; Barata, C. Impact of the Antibiotic Doxycycline on the D. Magna Reproduction, Associated Microbiome and Antibiotic Resistance Genes in Treated Wastewater Conditions. Environ. Pollut. 2023, 334, 122188. [Google Scholar] [CrossRef]
- Dai, H.; Gao, J.; Li, D.; Wang, Z.; Cui, Y.; Zhao, Y. Family Sphingomonadaceae as the key executor of triclosan degradation in both nitrification and denitrification systems. J. Chem. Eng. 2022, 442, 136202. [Google Scholar] [CrossRef]
- Abou Dalle, A.; Djelal, H.; Domergue, L.; Fourcade, F.; Amrane, A. A novel system coupling an electr-Fenton process and an advanced biological process to remove pharmaceutical coumpound, metronidazole. J. Hazard. Mater. 2021, 415, 125705. [Google Scholar] [CrossRef]
Figure 1.
Microscopic observations (×100) of the communal WWTP mixed liquor just after sampling (a) its main species—Holotriche (b), Spirotriche (c), Acinetiens (d), Peritriche (e), Rotifera (f), Nematodes (g)—regular floc (h), and desintegrated floc (i).
Figure 1.
Microscopic observations (×100) of the communal WWTP mixed liquor just after sampling (a) its main species—Holotriche (b), Spirotriche (c), Acinetiens (d), Peritriche (e), Rotifera (f), Nematodes (g)—regular floc (h), and desintegrated floc (i).

Figure 2.
Mixed liquor suspended solids content (MLSS) (a), conductivity (b), and SVI (c) of the mixed liquor with 0, 100, 200, and 400 mg L−1 of doxycycline after 23 days of culture at ambient temperature and under fed-batch conditions (--- initial concentration).
Figure 2.
Mixed liquor suspended solids content (MLSS) (a), conductivity (b), and SVI (c) of the mixed liquor with 0, 100, 200, and 400 mg L−1 of doxycycline after 23 days of culture at ambient temperature and under fed-batch conditions (--- initial concentration).

Figure 3.
Comparison of sludge settleability and microscopic images (×100) of mixed liquor at day 0 (a) and after 23 days of culture (b) without doxycycline addition.
Figure 3.
Comparison of sludge settleability and microscopic images (×100) of mixed liquor at day 0 (a) and after 23 days of culture (b) without doxycycline addition.


Figure 4.
Relative abundance of the different bacterial phyla according to incubation time and doxycycline concentration. S and S′ are replicates of the sludge on day 0. D0, D100, D200, and D400 represent sludges incubated with the addition of glucose (2 g L−1) and increasing concentrations of doxycycline (0, 100, 200, and 400 mg L−1, respectively) on days 7, 14, and 23. D400′ is a replicate of D400.
Figure 4.
Relative abundance of the different bacterial phyla according to incubation time and doxycycline concentration. S and S′ are replicates of the sludge on day 0. D0, D100, D200, and D400 represent sludges incubated with the addition of glucose (2 g L−1) and increasing concentrations of doxycycline (0, 100, 200, and 400 mg L−1, respectively) on days 7, 14, and 23. D400′ is a replicate of D400.

Figure 5.
Richness (bars) and Shannon diversity index (open squares) of the activated sludge microbial communities. The legend is the same as in Figure 4.
Figure 5.
Richness (bars) and Shannon diversity index (open squares) of the activated sludge microbial communities. The legend is the same as in Figure 4.

Figure 6.
Principal component analysis of the relative abundance of all species of all microbial communities. The dots are the microbial communities named in Figure 4. The colors correspond to the doxycycline concentrations in (mg L−1): red (0), green (100), blue (200), and purple (400). The arrows are the 15 most dominant vectors identifying the microbial species driving the structural differences between the microbial communities. They are identified in the text and Table S1.
Figure 6.
Principal component analysis of the relative abundance of all species of all microbial communities. The dots are the microbial communities named in Figure 4. The colors correspond to the doxycycline concentrations in (mg L−1): red (0), green (100), blue (200), and purple (400). The arrows are the 15 most dominant vectors identifying the microbial species driving the structural differences between the microbial communities. They are identified in the text and Table S1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mixed liquor composition (n = 3).
| Parameters | Values |
|---|---|
| pH | 7.3 ± 0.28 |
| Turbidity (NTU) | 5.8 ± 4.20 |
| Conductivity (µS cm−1) | 870 ± 20 |
| MLSS (g L−1) | 4.9 ± 0.11 |
| SVI (mL g−1 MLSS) | 100 ± 4.20 |
Table 2.
Database used for our ANOVA.
| Sample N° | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [DC] (mg L−1) | 0 | 0 | 0 | 0 | 100 | 100 | 100 | 100 | 200 | 200 | 200 | 200 | 400 | 400 | 400 | 400 |
| Cultivation time (day) | 0 | 7 | 14 | 23 | 0 | 7 | 14 | 23 | 0 | 7 | 14 | 23 | 0 | 7 | 14 | 23 |
| MLSS (g·L−1) | 4.9 | 7.0 | 5.6 | 13.5 | 4.9 | 6.2 | 4.4 | 13.1 | 4.9 | 6.5 | 6.2 | 15.5 | 4.9 | 6.6 | 6.2 | 15.9 |
Table 3.
Analysis of variance (ANOVA) results.
| Degree of Freedom (DF) | Sum of Squares (SS) | Mean Square (MS) | f-Statistical | p-Value | |
|---|---|---|---|---|---|
| DC concentration (mg L−1) | 1 | 0.0003339 | 0.000334 | 0.2796 | 0.607 |
| Time of cultivation (day) | 1 | 0.0044208 | 0.004421 | 3.7022 | 0.078 |
| DC concentration x days (mg L−1·day) | 1 | 0.0003468 | 0.000357 | 0.2904 | 0.600 |
| Residuals | 12 | 0.0143292 | 0.001194 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Djelal, H.; Haddouche, D.; Lebreton, M.; Barros, V.; Villegas, C.; Dabert, P. Impact of Doxycycline Addition on Activated Sludge Microflora and Microbial Communities. Processes 2024, 12, 350. https://doi.org/10.3390/pr12020350
AMA Style
Djelal H, Haddouche D, Lebreton M, Barros V, Villegas C, Dabert P. Impact of Doxycycline Addition on Activated Sludge Microflora and Microbial Communities. Processes. 2024; 12(2):350. https://doi.org/10.3390/pr12020350
Chicago/Turabian StyleDjelal, Hayet, Djouza Haddouche, Megane Lebreton, Vanessa Barros, Cristian Villegas, and Patrick Dabert. 2024. "Impact of Doxycycline Addition on Activated Sludge Microflora and Microbial Communities" Processes 12, no. 2: 350. https://doi.org/10.3390/pr12020350
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.