
Monitoring Yeast Cultures Grown on Corn Stover Hydrolysate for Lipid Production
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Feedstock
CSH Preparation
2.2. Strains
2.3. Inocula
2.4. Strain Screening Test
2.5. Lab-Scale Bioreactor Cultivation
2.6. Analytical Procedures
2.6.1. Biomass Quantification
2.6.2. Sugar, Short-Chain Organic Acid, and Sugar-Derived Aldehyde Quantification
2.6.3. FC Analysis
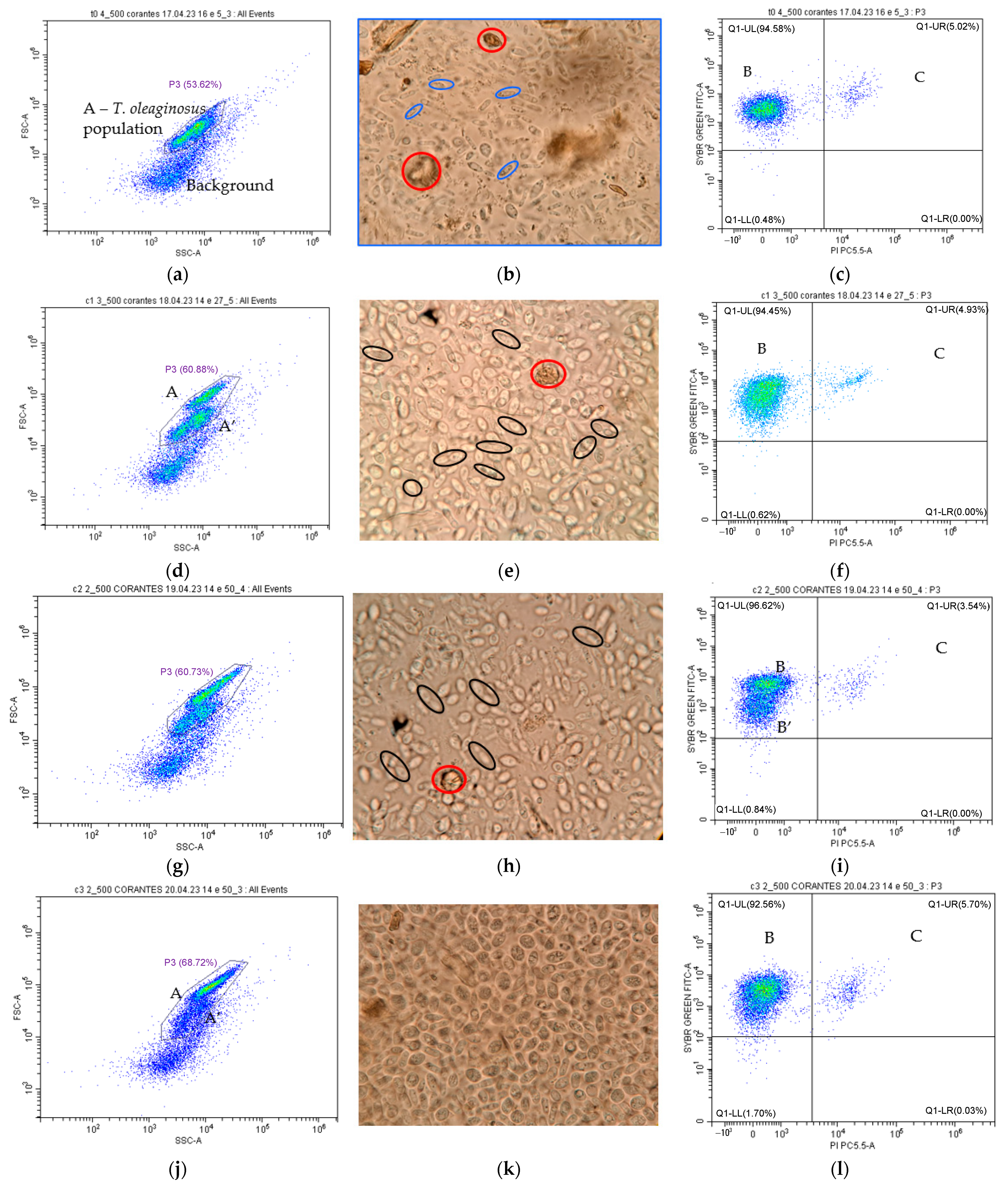
2.6.4. Optical Microscope Observations
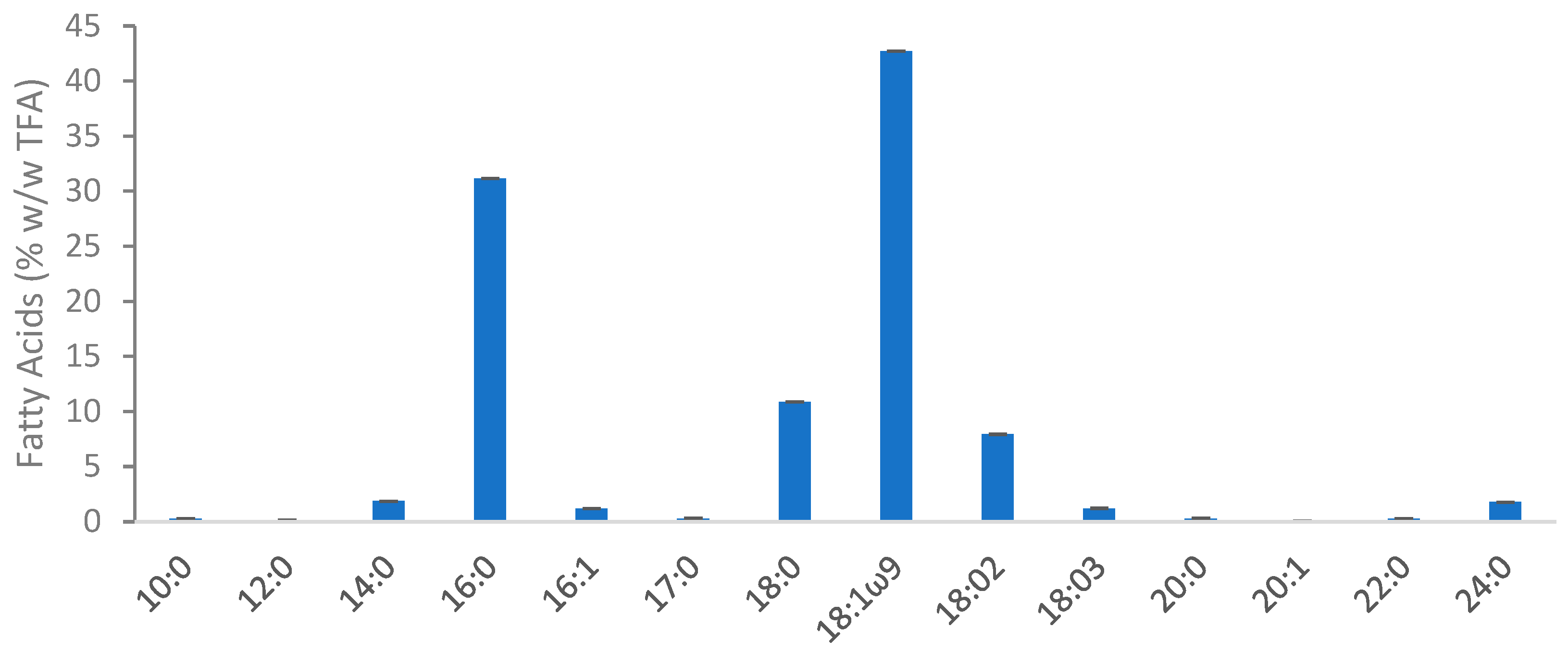
2.6.5. Fatty Acid Profile and Quantification
2.6.6. Total Lipid Quantification
2.6.7. Moisture Content
2.7. Kinetic Parameters
3. Results and Discussion
3.1. Yeast Strain Screening Test in CSH + CSL Complex Medium
3.2. Bioreactor Experiment
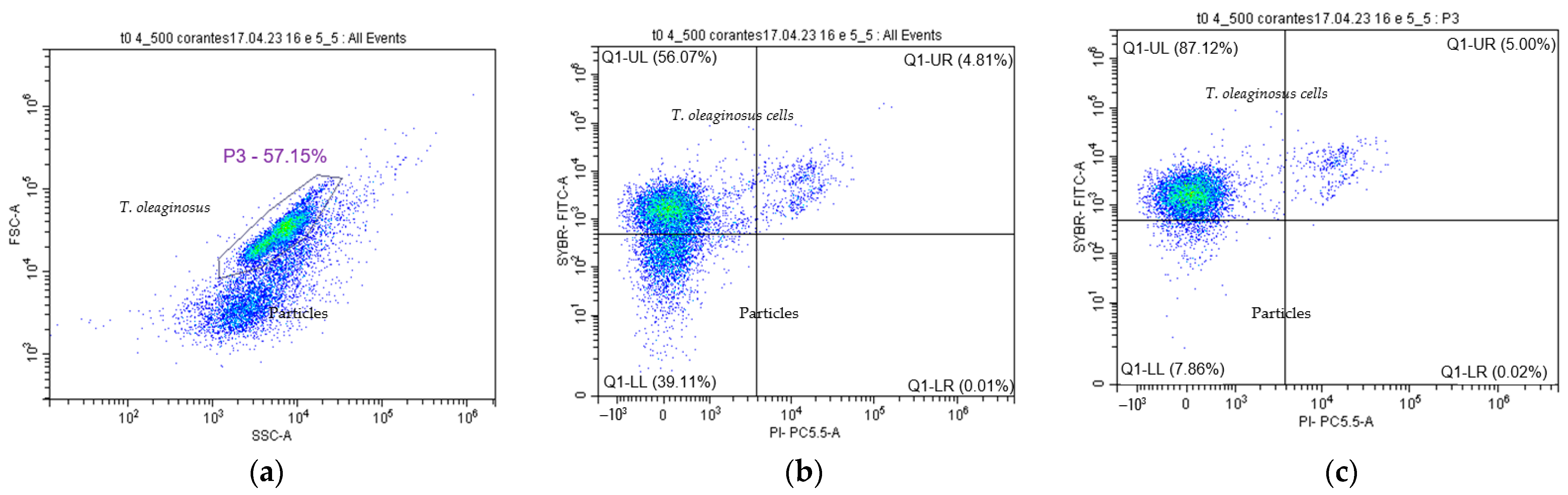
3.2.1. Flow Cytometry Preliminary Controls for T. oleaginous
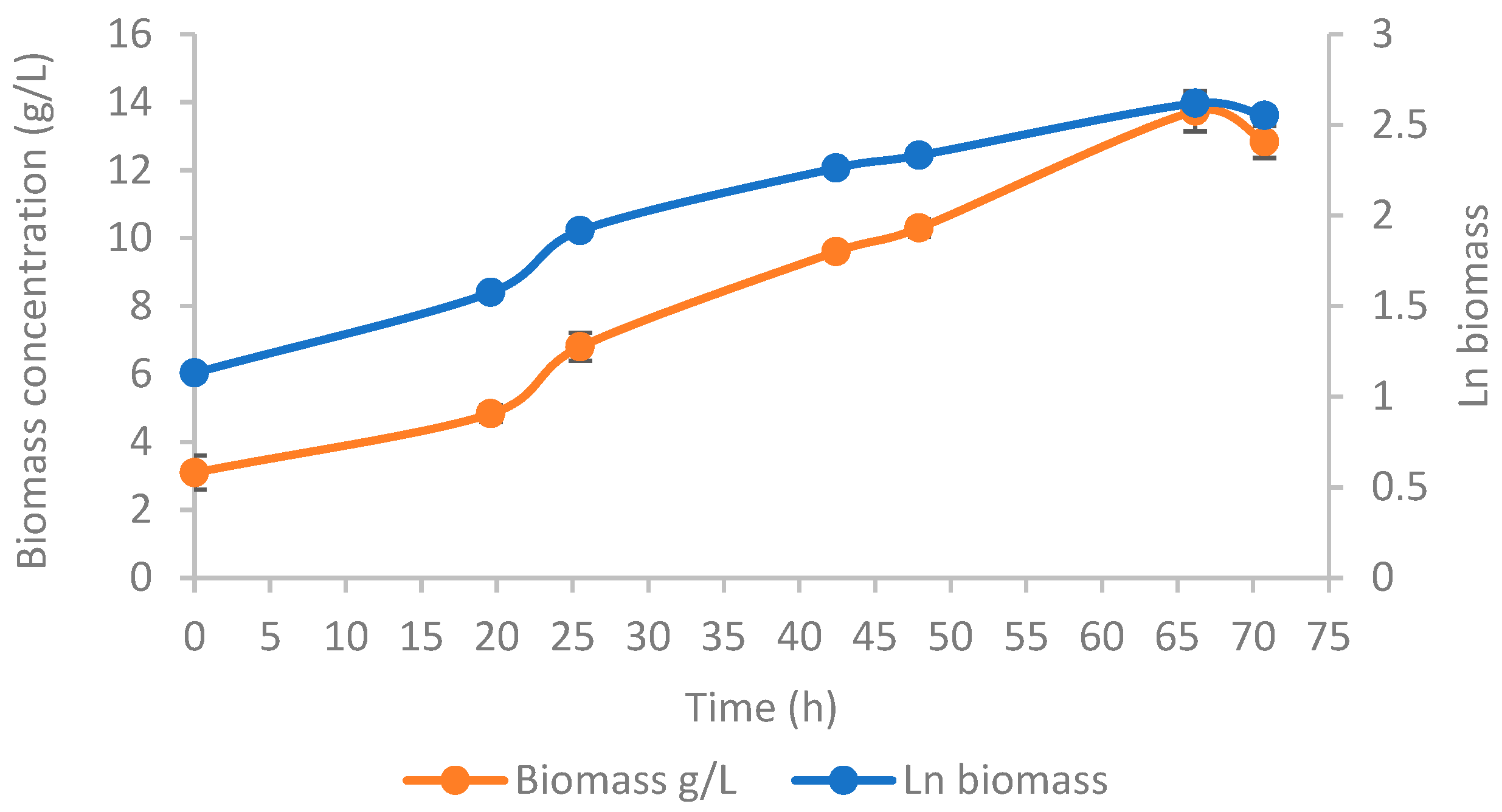
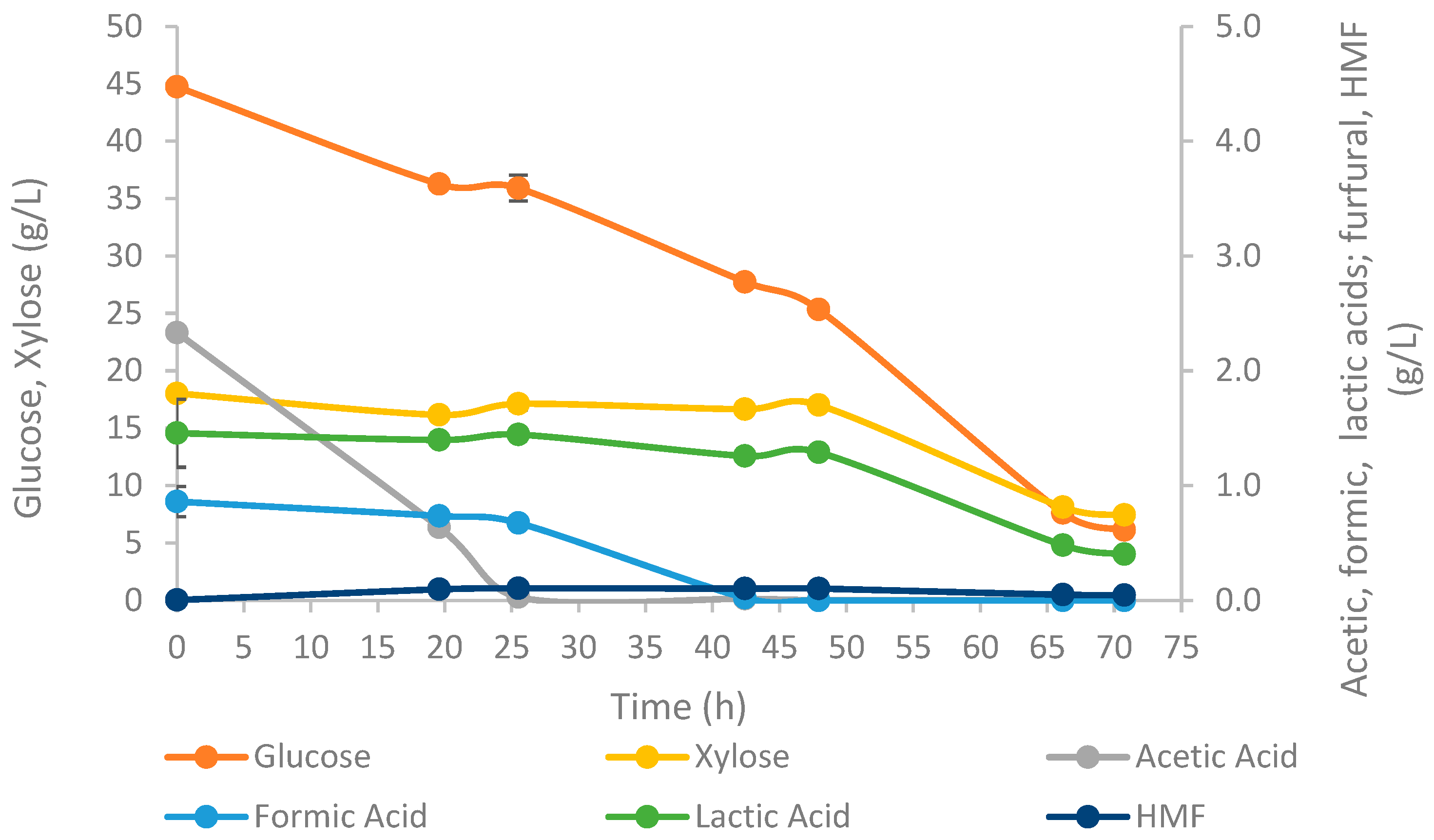
3.2.2. T. oleaginosus Growth
3.2.3. T. oleaginosus Morphological and Physiological Status Monitoring throughout the Bioreactor Cultivation
3.2.4. Lipid Production by T. oleaginosus Yeast Strains Grown on Synthetic Media and Lignocellulosic Hydrolysates
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Paraskar, P.M.; Prabhudesai, M.S.; Hatkar, V.; Kulkarni, R. Vegetable oil-based polyurethane coatings—A sustainable approach: A review. Progr. Org. Coat. 2021, 156, 106267. [Google Scholar] [CrossRef]
- Karmakar, G.; Ghosh, P.; Sharma, B. Chemically Modifying vegetable oils to prepare green lubricants. Lubricants 2017, 5, 44. [Google Scholar] [CrossRef]
- Jez, S.; Spinelli, D.; Fierro, A.; Dibenedetto, A.; Aresta, M.; Busi, E.; Basosi, R. Comparative life cycle assessment study on environmental impact oil production from microalgae and terrestrial oilseed crops. Bioresour. Technol. 2017, 239, 266–275. [Google Scholar] [CrossRef]
- 174th Session of the FAO Council. Item 6: The Impact of the War in Ukraine on Global Food Security and Related Matters under the Mandate of the FOOD and Agriculture Organization of the United Nations (FAO). 2022. Available online: https://www.fao.org/3/no015en/no015en.pdf (accessed on 12 January 2024).
- Asare, M.; Souza, F.; Gupta, R. Waste to resource: Synthesis of polyurethanes from waste cooking oil. Ind. Eng. Chem. Res. 2022, 61, 18400–18411. [Google Scholar] [CrossRef]
- Sajish, S.; Singh, S.; Nain, L. Yeasts for Single Cell Oil Production from Non-Conventional Bioresources. In Microbial Biotechnology for Renewable and Sustainable Energy; Saini, J.K., Sani, R.K., Eds.; Clean Energy Production Technologies; Springer Nature: Singapore, 2022; pp. 337–364. [Google Scholar]
- Caporusso, A.; Capece, A.; De Bari, I. Oleaginous yeasts as cell factories for the sustainable production of microbial lipids by the valorization of agri-food wastes. Fermentation 2021, 7, 50. [Google Scholar] [CrossRef]
- Worldwide Production of Grain in 2023/24, by Type. Available online: https://www.statista.com/statistics/263977/world-grain-production-by-type/ (accessed on 22 January 2024).
- Ruan, Z.; Wang, W.; Liu, Y.; Liao, W. Chapter 3: Corn. In Integrated Processing Technologies for Food and Agricultural By-Products; Pan, Z., Zhang, R., Zicar, S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 59–72. [Google Scholar]
- Amândio, M.S.T.; Rocha, J.M.S.; Xavier, A.M.R.B. Enzymatic hydrolysis strategies for cellulosic sugars production to obtain bioethanol from eucalyptus globulus bark. Fermentation 2023, 9, 241. [Google Scholar] [CrossRef]
- Li, X.; Shi, Y.; Kong, W.; Wei, J.; Song, W.; Wang, S. Improving enzymatic hydrolysis of lignocellulosic biomass by bio-coordinated physicochemical pretreatment—A review. Energy Rep. 2022, 8, 696–709. [Google Scholar] [CrossRef]
- Freitas, C.; Neves, E.; Reis, A.; Passarinho, P.; Lopes da Silva, T. Effect of acetic acid n Saccharomyces carlsbergensis ATCC 6269 batch ethanol production monitored by flow cytometry. Appl. Biochem. Biotechnol. 2012, 168, 1501–1515. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C. Improved recovery of fatty acid through direct transesterification without prior extraction and purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Di Fidio, N.; Minonne, F.; Antonetti, C.; Raspolli Galletti, A.M. Cutaneotrichosporon oleaginosus: A versatile whole-cell biocatalyst for production of single-cell oil from agro-industrial wastes. Catalyst 2021, 11, 1291. [Google Scholar] [CrossRef]
- Wang, J.; Gao, Q.; Zhang, H.; Bao, J. Inhibitor degradation and lipid accumulation potentials of oleaginous yeast Trichosporom cutaneum using lignocellulose feedstock. Bioresour. Technol. 2016, 281, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, J.; Blomqvist, J.; Shapaval, V.; Kohler, A.; Sampels, S.; Sandgren, M.; Volkmar, P. Oleaginous yeasts respond differently to carbon sources present in lignocellulose hydrolysate. Biotechnol. Biofuels 2021, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Thabet, S.; Weiss-Gayet, M.; Dappozze, F.; Cotton, P.; Guillard, C. Photocatalysis on yeast cells: Towards targets and mechanisms. Appl. Catal. B Environ. 2013, 140–141, 169–178. [Google Scholar] [CrossRef]
- Holdsworth, J.; Ratledge, C. Lipid turnover in oleaginous yeasts. J. Gen. Microbiol. 1988, 134, 339–346. [Google Scholar] [CrossRef]
- Yen, H.-W.; Zhang, Z. Effects of dissolved oxygen level on cell growth and total lipid accumulation on the cultivation of Rhodotorula glutinis. J. Biosci. Bioeng. 2011, 112, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Caporusso, A.; Bari, I.; Valerio, V.; Albergo, R.; Liuzzi, F. Conversion of cardoon crop residues into single cell oils by Lipomyces tetrasporus and Cutaneaotrichosporum curvatus; process optimizations to overcome the microbial inhibition of lignocellulosic hydrolysates. Ind. Crops Prod. 2021, 159, 113030. [Google Scholar] [CrossRef]
- Bouchedja, D.N.; Danthine, A.; Kar, T.; Fickers, P.; Boudjellal, A.; Delvigne, F. Online flow cytometry, and interesting investigation process for monitoring lipid accumulation, dimorphism and cell’s growth on the oleaginous yeast Yarrowia lipolytica JMY 775. Bioresour. Bioprocess. 2017, 4, 3. [Google Scholar] [CrossRef]
- Santek, M.; Miskulin, A.; Petrovic, A.; Beluhan, S.; Santek, B. Effect of carbon and nitrogen source concentrations on the growth and lipid accumulation of yeast Trichosporon oleaginosus in continuous and batch culture. J. Chem. Technol. Biotechnol. 2017, 92, 1620–1629. [Google Scholar] [CrossRef]
- Capusoni, C.; Rodighiero, V.; Cucchetti, D.; Galafassi, S.; Bianchi, D.; Franzosi, G.; Compagno, C. Characterization of lipid accumulation and lipidome analysis in the oleaginous yeasts Rhodosporidium azoricum and Trichosporon oleaginosus. Bioresour. Technol. 2017, 238, 281–289. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, R.; Tang, L.; Zhu, L.; Zhu, D.; Bai, F. Dimorphim of Trichosporon cutaneum and impact on its lipid production. Biotechnol. Biofuels 2019, 12, 203. [Google Scholar] [CrossRef]
- Meo, A.; Priebe, X.; Weuster-Botz, D. Lipid production with Trichosporon oleaginosus in a membrane bioreactor using microalgae hydrolysate. J. Biotechnol. 2017, 241, 1–10. [Google Scholar] [CrossRef]
- Liu, Q.; Lu, M.; Jin, C.; Hou, W.; Zhao, L.; Bao, J. Ultra-centrifugation force in adaptive changes the cells structure of oleaginous yeast Trichosporon cutaneum into favorable space for lipid accumulation. Biotechnol. Bioeng. 2022, 119, 1509–1521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bao, J. Tolerance of Trichosporon cutaneum to lignin derived phenolic aldehydes facilitate the cell growth and cellulosic lipid accumulation. J. Biotechnol. 2022, 343, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xu, Z.; Chen, S.; Jin, M. Microbial lipid production from dilute acid alkali pretreated corn stover via Trichosporon dermatis. Bioresour. Technol. 2020, 295, 122523. [Google Scholar] [CrossRef]
- Hu, C.; Wu, S.; Wang, Q.; Jin, Q.; Shen, H.; Zhao, Z. Simultaneous utilization of glucose and xylose for lipid production by Trichosporon cutaneum. Biotechnol. Biofuels 2011, 4, 25. [Google Scholar] [CrossRef]
- Santek, M.; Lisicar, J.; Musak, L.; Spoljaric, I.; Beluhan, S.; Santek, B. Lipid production by yeast Trichosporon oleaginosus on the enzymatic hydrolysate of alkaline pretreated corn cobs for biodiesel production. Energy Fuels 2018, 32, 12501–12513. [Google Scholar] [CrossRef]
- Ujor, V.C.; Okonkwo, C. Microbial detoxification of lignocellulosic biomass hydrolysates: Biochemical and molecular aspects, challenges, exploits and future perspectives. Front. Bioeng. Biotechnol. 2022, 10, 1061667. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
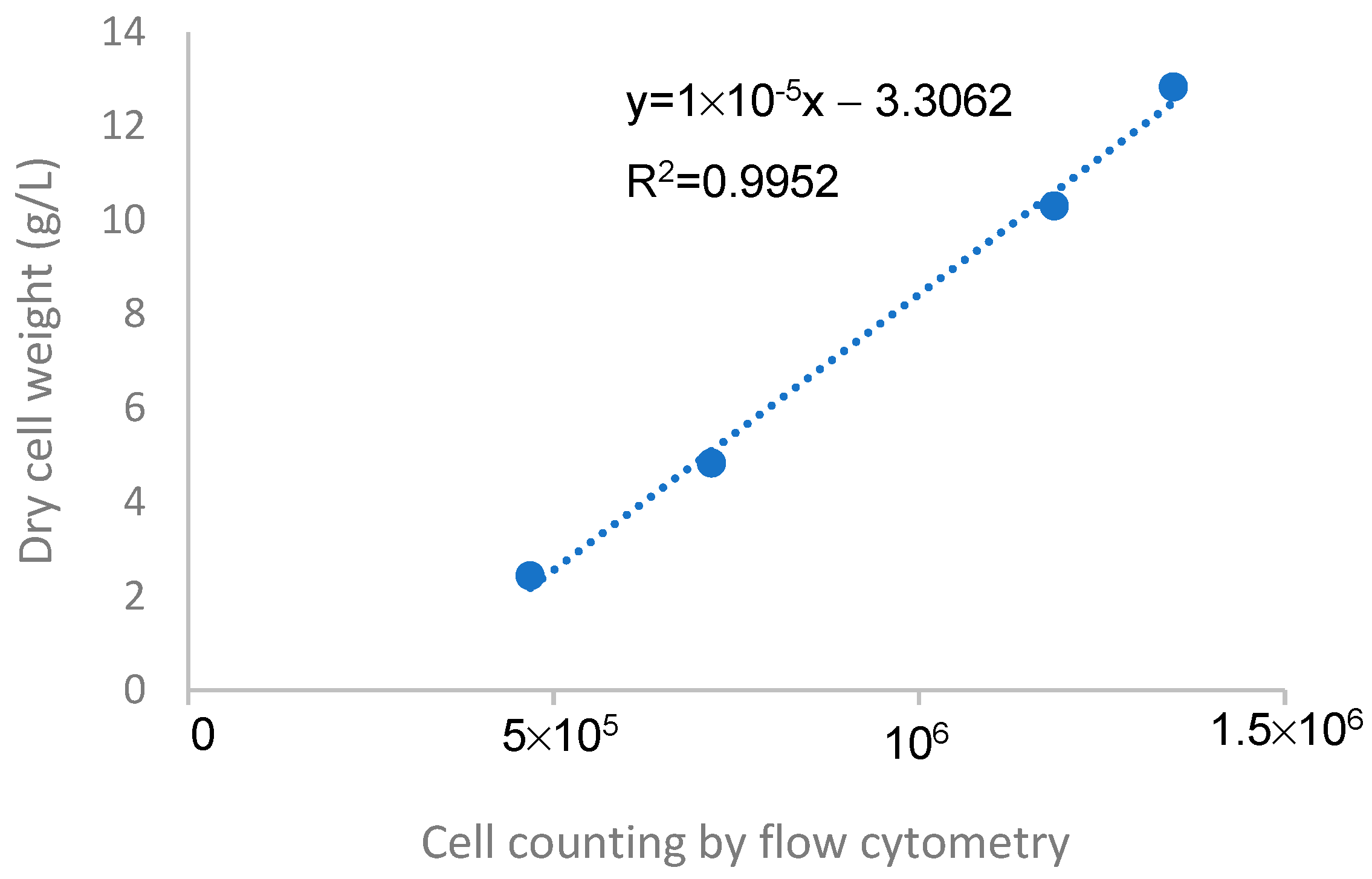{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | T. oleaginosus | R. babjevae |
|---|---|---|
| Cultivation Time (h) | 47.1 | 88.9 |
| Growth Kinetic Parameters: | ||
| Xmax (g/L) | 27.48 ± 0.58 (47.1 h) | 12.95 ± 0.86 (66.1 h) |
| Xfinal (g/L) | 27.48 ± 0.58 | 11.54 ± 1.46 |
| PX (g/Lh) | 0.51 ± 0.01 | 0.14 ± 0.00 |
| µmax (/h) | 0.05 ± 0.00 | 0.06 ± 0.00 |
| YX/S (gbiomass/gsubstrate) | 0.39 ± 0.00 | 0.27 ± 0.06 |
| rglucose (g/Lh) | 0.85 ± 0.03 | 0.27 ± 0.01 |
| rxylose (g/Lh) | 0.41 ± 0.02 | 0.03 ± 0.01 |
| racetic acid (g/Lh) | 0.06 ± 0.00 | 0.03 ± 0.00 |
| rformic acid (g/Lh) | 0.02 ± 0.00 | - |
| rlactic acid (g/Lh) | 0.02 ± 0.00 | 0.01 ± 0.00 |
| Parameter | T. oleaginosus | R. babjevae |
|---|---|---|
| Lipid content (w/w%) | 24.6 ± 0.1 | 18.5 ± 0.3 |
| PL (g/Lh) | 0.14 ± 0.00 | 0.04 ± 0.00 |
| YLipids/S (g/g) | 0.11± 0.00 | 0.08 ± 0.00 |
| Parameter | T. oleaginosus |
|---|---|
| Xmax (g/L) | 13.73 ± 0.50 (t = 66.2 h) |
| Xfinal (g/L) | 12.83 ± 0.50 (t = 70.8 h) |
| ProdX (g/Lh) | 0.14 ± 0.01 |
| µ (/h) | 0.02 ± 0.06 |
| YX/S (gbiomass/gglucos+xylose) | 0.20 ± 0.02 |
| rglucose (g/Lh) | 0.54 ± 0.01 |
| rxylose (g/Lh) | 0.15 ± 0.01 |
| racetic acid (g/Lh) | 0.11 ± 0.00 |
| rformic acid (g/Lh) | 0.02 ± 0.00 |
| rlactic acid (g/Lh) | 0.01 ± 0.00 |
| Lipid content (% w/w) | 42.28 ± 0.01 |
| PL (g/Lh) | 0.06 ± 0.01 |
| YLipids/S | 0.11± 0.00 |
| Yeast | Genetically Modified | Feedstock | Pretreatment Hydrolysis | Cultivation System/Regime | % Lipids (w/w) | CL g/L | Lipid Productivity (g/L/h) | Reference |
|---|---|---|---|---|---|---|---|---|
| T. cutaneum AS 2.571 | No | Corn stover | Dilute sulfuric acid pretreatment; enzymatic hydrolysis; hydrolysate detoxified with an excess of lime | 250 mL unbaffled conical flasks/batch | 39.20 | 7.6 | 0.08 | Hu et al., 2011 [29] |
| T. oleaginosus ATCC 20509 | No | Synthetic media | n.a. | 20 L bioreactor with 6 L medium/fed-batch | 54.00 | 71.28 | 0.51 | Capusoni et al., 2017 [23] |
| T. oleaginosus DSM 11815 | No | Microalgae hydrolysate | High-impact homogenizer/enzymatic hydrolysis | Membrane bioreactor with total cell retention under phosphate limitation/batch | 53.00 | 30.6 | 0.33 | Meo et al., 2017 [25] |
| T. oleaginosus DSM 11815 | No | Synthetic culture medium | n.a. | 0.5 L shake flasks/batch | 45.33 | 6.7 | 0.67 | Šantek et al., 2017 [22] |
| T. oleaginosus DSM 11815 | No | Corn cobs | Alkaline pretreatment/enzymatic hydrolysis | Bioreactor with 25 L working volume/fed-batch | 22.82 | 0.07 | Šantek et al., 2018 [30] | |
| T. cutaneum B3 | Yes | Synthetic medium | n.a. | 5 L bioreactor with 2 L working volume | 46.20 | 12.52 | 0.17 | Wang et al., 2019 [24] |
| T. dermatis 32903 | No | Corn stover | Diluted alkali pretreatment/enzymatic hydrolysis | 250 mL baffled shake flasks | 55.97 | 20.36 | 0.21 | Yu et al., 2020 [28] |
| T. cutaneum MP11 | Yes | Wheat straw | Dry acid pretreatment, followed by detoxification with Ca(OH)2; enzymatic hydrolysis | 3 L bioreactor containing 1 L wheat straw hydrolysate; simultaneous saccharification and co-fermentation (SSCF) | 67.80 | 34.4 | 0.36 | Liu et al., 2022 [26] |
| T. cutaneum MP11 | Yes | Wheat straw | Acid pretreatment followed by biodetoxification using Paecylomyces variotii FN89; enzymatic hydrolysis | 3 L bioreactor (SSCF)/batch | 40.87 | 0.28 | Zhang et al., 2022 [27] | |
| T. oleaginosus DSM 11815 | No | Corn stover | Steam explosion; enzymatic hydrolysis | 7 L bioreactor with 2 L medium/batch | 42.28 | 5.38 | 0.06 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontes, A.; Francisco, R.; Ferreira, F.C.; Faria, N.T.; Marques, S.; Reis, A.; Moura, P.; Lukasik, R.; Santos, J.; Lopes da Silva, T. Monitoring Yeast Cultures Grown on Corn Stover Hydrolysate for Lipid Production. Processes 2024, 12, 558. https://doi.org/10.3390/pr12030558
Fontes A, Francisco R, Ferreira FC, Faria NT, Marques S, Reis A, Moura P, Lukasik R, Santos J, Lopes da Silva T. Monitoring Yeast Cultures Grown on Corn Stover Hydrolysate for Lipid Production. Processes. 2024; 12(3):558. https://doi.org/10.3390/pr12030558
Chicago/Turabian StyleFontes, Afonso, Ricardo Francisco, Frederico Castelo Ferreira, Nuno Torres Faria, Susana Marques, Alberto Reis, Patrícia Moura, Rafal Lukasik, José Santos, and Teresa Lopes da Silva. 2024. "Monitoring Yeast Cultures Grown on Corn Stover Hydrolysate for Lipid Production" Processes 12, no. 3: 558. https://doi.org/10.3390/pr12030558