How Do Indirect Effects of Contaminants Inform Ecotoxicology? A Review

Department of Biological Sciences, Louisiana State University, Baton Rouge, LA 70803, USA
Processes 2020, 8(12), 1659; https://doi.org/10.3390/pr8121659
Submission received: 19 November 2020
/
Revised: 8 December 2020
/
Accepted: 11 December 2020
/
Published: 16 December 2020
(This article belongs to the Special Issue Environmental Risk Assessment Processes and Ecotoxicology)
Abstract
:Indirect effects in ecotoxicology are defined as chemical- or pollutant-induced alterations in the density or behavior of sensitive species that have cascading effects on tolerant species in natural systems. As a result, species interaction networks (e.g., interactions associated with predation or competition) may be altered in such a way as to bring about large changes in populations and/or communities that may further cascade to disrupt ecosystem function and services. Field studies and experimental outcomes as well as models indicate that indirect effects are most likely to occur in communities in which the strength of interactions and the sensitivity to contaminants differ markedly among species, and that indirect effects will vary over space and time as species composition, trophic structure, and environmental factors vary. However, knowledge of indirect effects is essential to improve understanding of the potential for chemical harm in natural systems. For example, indirect effects may confound laboratory-based ecological risk assessment by enhancing, masking, or spuriously indicating the direct effect of chemical contaminants. Progress to better anticipate and interpret the significance of indirect effects will be made as monitoring programs and long-term ecological research are conducted that facilitate critical experimental field and mesocosm investigations, and as chemical transport and fate models, individual-based direct effects models, and ecosystem/food web models continue to be improved and become better integrated.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction to Indirect Effects
Many synthetic chemicals and naturally occurring chemical pollutants (e.g., poly-nuclear aromatic hydrocarbons) impact living things by direct toxic action exerted on an organism’s physiology, behavior, reproduction, or survivorship. Resulting lethal or sublethal (e.g., reproductive failure) effects may alter the abundance and diversity of biota in contaminated habitats to such a degree that severe impacts to ecosystem function and services occur. However, contaminants may also indirectly affect the biota in numerous ways with equivalent consequences. Indirect effects are defined here as chemical- or pollutant-induced alterations in the density or behavior of sensitive species in an exposed system that then have cascading effects on tolerant species [1]. These effects occur at higher levels of biological organization (i.e., populations, communities, and ecosystems) and cannot be identified or assessed in single-species laboratory tests. The purpose of this review is to address indirect effects that are a function of community ecology (i.e., effects that occur when species interaction networks are disrupted by a contaminant), and because changes in populations or communities may cascade to higher levels, indirect contaminant effects associated with ecosystem function and services will also be discussed. Previous reviews on indirect effects have largely focused on specific topics, e.g., how community ecology informs indirect effects [2,3] or indirect effects in aquatic ecosystems [1,4,5,6,7], rather than an overview within the broader context of ecotoxicology as presented here. Although rarely studied compared to traditional laboratory-based investigations of direct effects, the number of publications that consider actual or potential indirect effects of chemicals or pollutants has increased over the last four decades (213 papers published in nine representative environmental toxicology journals from 1988 to 2019 mention indirect effects in a searchable field, as shown in Figure 1).
When relatively sensitive species in a biotic community are affected directly and experience near or complete extirpation, relatively tolerant species experience significant changes in the landscape of species interactions. These interactions include predation, competition, parasitism, and mutualism that occur within and among trophic levels and that govern abundance, distribution, and evolutionary fitness. Behavioral changes from sublethal exposures in relatively sensitive species may also occur that more subtly, but nevertheless, substantially alter species interactions [8]. As a result of these altered interactions, the community may change after exposure to a chemical contaminant or a pollutant, and relatively tolerant species may increase or decrease in abundance. Perhaps the most well-known example of an indirect effect was observed in an experimental whole-lake amendment of an endocrine disrupting chemical that inhibited reproduction and caused the near extirpation of fathead minnows [9]. Declines in this keystone species elicited dramatic cascading top–down and bottom–up indirect effects throughout the broader community; e.g., without minnows, zooplankton populations were released from predation and increased in abundance, and the biomass of an apex predator decreased.
Indirect effects are made possible because contaminants frequently affect exposed organisms differentially. Asymmetrical effects occur when one species is more sensitive and another is more resistant to a contaminant [10]. A simple, straightforward example of indirect effects in a low diversity system is illustrated by a hypothetical food chain in which one or a few producer species are consumed by one or a few primary consumer species, which are in turn consumed by one or a few secondary consumer species (Figure 2). Species in each trophic level in low diversity systems would likely have similar tolerances to herbicides (that are toxic to most primary producers but not most primary consumers) and pesticides (that are toxic to most primary and secondary consumers but not most primary producers). Indirect effects in this community would likely be pronounced and relatively predictable [11,12]. In this scenario, an herbicide addition would have a direct effect on primary producers, and because biomass or primary production would be reduced, primary and secondary consumers would experience reduced energy flow and a negative indirect effect on density (a bottom–up effect). In this same system, pesticide addition would have a direct effect on herbivores, reducing density. This could lead to a positive indirect effect on primary producers as herbivory declines (a top–down effect).
However, given the wide ranges in tolerance among species within ecosystems [13,14], the potential for an asymmetrical response increases with increasing species diversity [4,15]. Interaction networks [16,17] and feedback loops [8] also tend to become more complex as diversity increases. Therefore, as biodiversity increases, indirect effects are likely to be more complex and variable, milder due to possible compensatory responses, and less straightforwardly predictable [3]. In communities with complex food web structures and interaction networks, an herbicide addition would exert a direct effect on relatively sensitive primary producers that would reduce producer biomass and/or primary production (Figure 3). However, a positive indirect effect is possible over time on relatively tolerant primary producer species as they are released from competition, and a resulting increase in biomass or primary production for tolerant species may compensate at the ecosystem level for effects on sensitive primary producers [4]. An indirect effect on primary and secondary consumers may be negative because the overall basal food supply may be reduced, or it may be positive for consumers that are able to exploit tolerant primary producers that increase in biomass (Figure 3). For pesticide exposure in a diverse community, the direct effect on relatively sensitive herbivore species reduces density, and a positive indirect effect is exerted on primary producers as herbivory is reduced (Figure 4). However, a possible positive indirect effect on relatively tolerant herbivores could follow as competition among consumer species is reduced and as the basal food resource is increased [15,18]. Of course, indirect effects will be much more complex and context dependent if herbicide and pesticide exposures occur simultaneously, and it is possible that indirect effects could enhance or mask each other [4,15]. Indirect effects in areas with complex mixtures of contaminants are especially difficult to study and predict for these reasons, adding uncertainty to estimates of chemical risk [4,19,20].
Indirect effects may be of a greater magnitude than the direct effects of contaminants [9], and given the intricacies of ecological interactions, indirect effects may be more complex [11] and difficult to predict [2,3]. As a result, the assessment of chemical contaminants or pollutants that potentially contribute to indirect effects presents a significant challenge for ecotoxicology. For example, pesticides may sublethally diminish immune function, leading to increased parasite loads and heighted parasite-induced negative effects on hosts [21]. Few studies routinely quantify parasite load when examining pesticide effects in field, mesocosm, or laboratory studies, and studies that do consider them require an unusual range in expertise (i.e., zoology, immunochemistry, parasitology, analytical chemistry) among investigators. Furthermore, because many new chemicals are introduced annually and because many pollutants have been growing in use (e.g., neonicotinoids), new challenges to understand indirect effects and their interactions with communities will continue to be presented. For example, the assessment of indirect effects associated with plastics [22], nanoparticles [23], and perfluoroalkyls [24] have only recently been undertaken.
Many trends are becoming apparent in studies that investigate indirect effects. For example, contaminant-induced indirect effects will likely result from exposure to toxicants and chemical contaminants in all chemical classes when species within a community express large differences in toxicological sensitivity to the chemical. Contaminants in many chemical classes have been examined; however, indirect effects caused by pesticides have been most commonly identified, e.g., [25]. Although chemicals in many different classes have been shown to elicit indirect effects, chemical properties may affect the timing and the expression of indirect effects. For example, water-soluble contaminants may express indirect effects quickly, especially in pelagic ecosystems, because they are readily bioavailable [12]. However, highly hydrophobic chemicals may partition to non-aqueous phases (e.g., food, soils, or sediments) before uptake, slowing tissue level buildup within and between trophic levels; indirect effects may be strongest after biomagnification reaches critical levels. Furthermore, indirect effects have been identified in a wide array of ecosystems, although the largest number of field and experimental studies, as well as models, have been developed in freshwater ecosystems [20,26]. Finally, it is perhaps not surprising that indirect effects, particularly those acting via altered behavior, have not yet been broadly studied and quantified; multiple organisms across the trophic spectrum must be studied simultaneously in real time using environmentally realistic mesocosms or field-based studies, and the system often must be studied for longer durations than are typical of toxicological exposures (i.e., months to years).
2. Indirect Effects and Behavior
Direct and indirect contaminant effects on behavior (sometimes called trait-mediated effects) have become the focus of increased scrutiny [8,27,28,29,30]. The mechanisms underlying the subtle direct effects of contaminants on behavior are caused by the disruption of chemical, endocrine, and neural signaling mediated by receptors, enzymes (e.g., cholinesterase activity), and/or transporters [8]. Since direct effects of contaminants on behavior are physiologically complex and associated with receptors [31], species-specific responses to exposure are likely, and responses will undoubtedly vary with developmental stage and with varying environmental conditions. Furthermore, because consumer behavior is affected at low, sublethal concentrations, indirect effects occur in environments with low, chronic contamination, increasing the range of conditions and the frequency of habitats at which indirect effects may occur, e.g., at wastewater or sewage outfalls [29]. As a result, indirect contaminant effects associated with behavioral modifications are probably much more common than those associated with high levels of chemical-induced mortality or reproductive failure [3].
Studies on behavior have identified an exceptionally broad range of direct effects of contaminants belonging to many chemical classes, any of which could potentially lead to indirect effects. A partial listing for fishes indicates pollution-induced alterations of cognition, personality, sociability, memory, learning, appetite, parental care, boldness, and foraging as well as escape and sheltering behaviors [30]. For example, ecologically relevant exposures to metals and pesticides interfere with fish olfaction and behavior with observable effects on survival and reproductive success that could lead to indirect effects [32]. A meta-analysis of studies on amphibians found that exposure to contaminants increased rates of abnormal swimming (an increase of 168% compared to controls across all studies), a reduction in activity levels (−36%) and reproductive behaviors (−20%), and a reduced capacity to exhibit escape responses (−42%) [28]. Contaminants also alter behavior in invertebrates; the combination of predator presence and sublethal pesticide exposure resulted in a significant increase in drift behavior and a higher mortality rate in larval mayflies [33], and malathion caused male crickets to increase periods of non-directional movement, such as twitching and grooming, and to seek shelter less often [34]. These observations suggest that many more direct, yet subtle, alterations in behavior leading to indirect effects are likely to be found associated with a broad range of chemicals and in many taxa.
The potential complexity of behaviorally induced indirect effects is illustrated in a study in which perch (Perca fluviatilis) exposed to a psychiatric drug became more active and bolder than control fish [35]. Changes in behavior led to an increase in feeding rate on zooplankton. A cascading decline in zooplankton abundance could reduce the food supply to competitor fish species that do not express increased boldness. In addition, a reduction in herbivory caused by a declining zooplankton abundance may lead to a positive indirect effect on phytoplankton biomass. Moreover, increased boldness could also expose perch to increased rates of predation [29]. The broad range of direct effects suggests that alterations in the behavior of a sensitive species frequently reduce fitness (leading to reduced density) or cause effects on species interactions via influences on predators, prey, or competitors, further increasing the likelihood of indirect effects.
Behavioral responses that elicit indirect effects will also likely prove to be strongly interactive and context dependent, and many examples have already been observed. For example, environmental factors such as temperature may alter the effects of toxicants on behavior. Jacquin et al. [36] found that high temperatures aggravated the effects of pesticides in goldfish, and Lagesson et al. [37] found that exposure to a hormonal growth promoter increased boldness behavior in mosquitofish, but that this increase depended on temperature. Araujo et al. [38] observed that shrimps balanced exposure to contaminants with a sheltering behavior that protects individuals from predators; when confronted with a choice between a cleaner habitat with no shelter and a contaminated habitat with shelter, shrimps preferred the cleanest area, but when the uncontaminated area contained a predator signal, shrimps balanced the risk of predation and exposure to contamination by selecting a moderately contaminated area further away from predator signals, thereby increasing their exposure to the contaminant. These studies suggest that indirect effects will prove to be nearly ubiquitous, and they will be the subject many additional studies that examine trait-mediated effects.
3. Indirect Effects and Community Ecology
Community ecology provides a theoretical framework that anticipates and may predict the strength of and when and where indirect effects will occur [2,3,4,6,39]. Experimental studies and ecosystem models both indicate that interaction strength among species within communities plays an important role in determining the frequency and prominence of indirect effects [3,4,12]. Although interaction strength is sometimes equated with trophic state by measures such as grazing rate, keystone species, superior competitors, foundation species, and pollinators are also strong interactors, and changes in abundance or behavior that are induced by a chemical contaminant may have a range of indirect effects from subtle to extremely disruptive to ecosystem function [9,40]. In addition to interaction strength, large variations in sensitivity to direct effects among species or functional groups have been shown to influence indirect effects [10]; communities in which relatively sensitive species are ecological dominants increase the potential for indirect effects, and to the contrary, communities in which abundant functional groups with many species that function in the same ecological role that are less sensitive to direct effects will have reduced indirect effects [12]. Modeling also suggests that indirect effects may be greater in eutrophic ecosystems [12,41], although few studies have empirically tested this hypothesis (however, see [42] for an exception). Biodiversity may also affect the frequency and strength of indirect effects. Low diversity systems with few species (and therefore few tolerant species, [4,15]) and with a simple food chain structure (e.g., few trophic levels) may be more prone to pronounced indirect effects than high diversity systems with complex food webs with many functionally redundant species. Therefore, indirect effects in diverse communities with complex trophic relationships may be more difficult to characterize, anticipate, and model.
Although rarely studied explicitly, the magnitude and frequency of indirect effects will undoubtedly differ as biodiversity and species composition vary over space and time within communities. Zhao et al. [15] performed a mesocosm experiment to test for direct and indirect effects of herbicides and pesticides by manipulating the species number within trophic levels and the number of trophic levels within the experimental community. Direct effects of herbicides and pesticides were both reduced in treatments with a higher diversity of primary producers and herbivores, respectively, because more relatively tolerant species were present in high diversity treatments. Herbicide additions indirectly reduced herbivore biovolume, but this indirect effect was reduced when more primary producer species were present but was increased when a predator species was added to the community. Pesticide addition indirectly increased producer biovolume, and increasing the number of producer, herbivore, and predator species increased the indirect effect.
One challenge for future investigation is to evaluate the significance of low levels of chemical contamination on indirect effects [43]. Evidence is growing that commonly used chemicals such as pesticides alter biotic communities at low concentrations [44,45]. Two examples provided here reflect the ecological impact of seemingly subtle changes in community composition caused by pesticides that lead to very different indirect effects. Mensens et al. [46] found that pesticide exposure reduced diatom diet quality by shifting the community structure from dominance by lipid-rich species toward dominance by lipid-poor species, and that energy flow to herbivores was concomitantly reduced. Becker et al. [47] determined that snails that host the human parasite Schistosoma are relatively tolerant of pesticides and become dominant over their less tolerant competitors. This enhancement of snails that host the parasite led to increased schistosomiasis transmission to humans. Undoubtedly, more examples will arise as studies accumulate.
Community theory offers many possible predictions and insights for contaminant effects on species diversity [3]. For example, the general expectation from a high contaminant load is that species diversity will decrease; however, diversity could increase at lower or intermediate levels of contamination if there is a disproportionate effect on competitive dominants [3]. Therefore, community ecology may be a source of testable hypotheses for effects of contaminants as climate change and other perturbations occur. In addition, species diversity and indirect effects have potentially complex and intertwined relationships because of the association between biodiversity and ecosystem function. It is becoming axiomatic that species diversity affects ecosystem function, and that function (and services) decline when biodiversity is lost [48,49]. Since the direct effects of contaminants frequently reduce species diversity, indirect effects on ecosystem processes and services occur as species are reduced in abundance or extirpated. Much of the discussion of community theory and indirect effects centers on keystone species [50] and the loss of a sensitive keystone species may have significant long-term indirect consequences on ecosystem function [51]. The loss of biodiversity may also diminish the ability of communities to persist and be resilient after disturbances including from pollution events [52], or it may increase the possibility that the community will enter an alternative state that changes the ecosystem function for an extended period of time. For example, anthropogenic nutrient pollution contributes to coral-to-macroalgae phase shifts on reefs, and these shifts are enhanced by other anthropogenic factors [53]. Finally, lower diversity may make communities more vulnerable to other stressors including other contaminants, climate change, and invasive species [2].
4. Ecosystem Function/Services and Indirect Effects
Environmental regulations are ultimately intended to preserve ecosystem function (e.g., net primary production, decomposition rates, oxygen dynamics, nutrient cycling) and the ecosystem services (including provisioning, regulating, and supporting services) by which humans benefit. Although the direct and indirect effects of contaminants on ecosystem function have been subject to experimental investigation (for an example of effects on the decomposition rate, see [54]), effects are often inferred from studies on populations and communities. There are many reasons why relatively few studies directly test for contaminant direct and indirect effects on ecosystem function. Ecosystem studies require different and specialized (usually field-based) methods compared to the more assessable laboratory or mesocosm studies that follow contaminant-induced changes in density/survivorship or community structure. Large-scale manipulative experiments examining ecosystem function are especially informative but are expensive, difficult to conduct, and particularly rare [9,55,56]. Furthermore, the results of studies in one ecosystem are difficult to generalize or apply to other ecosystems or other ecosystem functions. In addition, there are two fundamentally different types of indirect contaminant effects that occur associated with ecosystem function that are seldom explicitly distinguished. First, pollutants and contaminants may directly influence important ecosystem functions that secondarily affect biota and wildlife. A common indirect effect at the ecosystem level is a reduction in resource availability to biota; e.g., primary production may be reduced by a pollutant resulting in reductions in the flow of energy up the food web [57] or more subtly, a decrease in the supply of food may increase the severity of direct contaminant effects on consumers [58]. Second, direct contaminant effects may also indirectly alter ecosystem function and services provided by the biota. For example, an ecosystem engineering species may be inhibited by a xenobiotic that slows the engineering process, e.g., bioturbator activity may be affected as a direct effect, altering the cycling or flux of nutrients from sediments [59]. Overall, studies designed to test effects on ecosystem function are seldom included in ecological risk assessment, suggesting a disconnect between ecological processes and the investigatory tools needed to consider chemical risk, although calls for an increased ecosystem perspective to improve the risk characterization of chemicals are common, e.g., [39,58,60]. Unfortunately, indirect effects will grow in importance as climate change continues and as more chemicals are introduced and dispersed globally.
Certainly, pollutants are known to directly and indirectly affect ecosystem function and ecosystem services (e.g., nutrient pollution increases primary production and subsequent oxygen declines associated with decomposition lead to bottom-water hypoxia and dead zones). However, less is known about the indirect effects of many specific chemical contaminants or toxicants on ecosystem function, and relatively few comprehensive reviews have addressed single compounds; however, see [61]. Peters et al. [62] reviewed toxicant effects in freshwater environments, and many instances of indirect effects were identified. The most commonly studied contaminant effect on ecosystem function was on leaf litter breakdown; reductions in the decomposition rate of organic matter in streams were indirectly attributed largely to declines in the feeding activity of macroinvertebrates caused by chemical contaminants. Perhaps the best known instance of an indirect effect of a chemical contaminant on an ecosystem service directly linked to humans is that of population declines of Gyps vultures in India due to diclofenac toxicity, which resulted in an increase in feral dogs scavenging on decaying carcasses and a consequent increase in human rabies infections from dog bites [63]. However, indirect effects on function and services are being explicitly examined in ecosystem models [64,65,66] for selected habitats or chemicals.
Indirect contaminant effects are being shown to affect many environmental issues and to foster unexpected or potentially undesirable outcomes. Indirect effects on ecosystem function have been found to affect biodiversity [4], be caused by invasive species [67,68], to occur during conservation efforts [69] and during habitat restoration [70,71], and after large-scale disturbance events such as oil spills [72,73], eutrophication [74], and climate change [51,75]. One concerning possibility is that pre-existing levels of contamination may affect the rate of recovery of biological functions after habitat restoration (e.g., wetland construction in areas in which soil contamination was not mitigated prior to construction); if the restored habitat is attractive to colonists, a so-called ecological trap may occur in which an existing contaminant load may reduce the survivorship or reproductive potential of colonists, slowing increases in biodiversity and the recovery of ecological function [76]. Mitigation efforts and recovery from the 2010 Deepwater Horizon oil spill are being influenced indirectly. Powers et al. [77] report that releasing Mississippi River water in coastal wetlands intended to reduce the transport of oil residues into productive salt marshes negatively affected oyster populations. In heavily oiled salt marshes after the spill, a slow recovery of vegetation led to decreased plant cover over marsh soils that in turn allowed more light to reach the marsh surface [78]. This increased light stimulated benthic microalgae, increasing the food supply for benthic animals, which unexpectedly led to higher densities in heavily oiled compared to unoiled marshes [78,79]. Finally, interactions between chemical contaminants and climate change will likely reveal many indirect effects on contaminant dispersion and bioavailability as well as ecosystem function [80,81,82,83,84].
5. Indirect Effects and Ecological Risk Assessment
Single-species toxicity tests have long formed the backbone of ecological risk assessment, and the results of such tests have routinely been used to establish no-effects standards for contaminants on populations and ecosystems and across landscapes [73]. Assessing risk based on effect levels determined for model organisms and applying safety margins ideally account for variation in sensitivity among species that would be sufficient to protect biota and ecosystem function. However, reliance on single-species tests in controlled settings fails to recognize that qualitatively distinctive impacts emerge when species interact and when contaminants affect communities and ecosystem processes. Indirect effects have frequently been shown to alter the outcome of a pollutant or a chemical in nature, making the effects different than those predicted in an ecological risk assessment [7,11,58,85,86]. For example, Rohr et al. [21] showed that a toxicant at a concentration that was been found to be safe in laboratory tests can affect amphibians when parasitic infection is present. Fodrie et al. [87] point out that studies of fish gene expression, physiology, and development exposed to oiling after the Deepwater Horizon oil spill uniformly indicate strong impacts at environmentally realistic exposure levels indicative of effects expected to reduce reproduction or survival, e.g., [88]. Yet, there was no indication of declines in population densities of fishes in oiled salt marshes and seagrass beds after the spill, and density increases were observed in nearshore species [89]. The behavioral avoidance of oil residues and the patchiness of oiling may have contributed to these observations, but indirect effects also contributed. Food web alterations occurred due to the direct effects of oil on predators, and commercial and recreation fishing closures were mandated; thereby, mortality rates were reduced, balancing potential declines from exposure to oil [89].
Indirect effects add uncertainty to ecological risk assessment in several ways. They may lessen apparent contaminant effects as relatively tolerant species compensate for contaminant-induced declines in ecological function delivered by relatively sensitive species [15,90]. For example, conversely indirect effects may magnify a direct impact if a keystone species or ecosystem engineering species is intolerant [4]. Indirect effects may also increase over time, making a determination of risk based on short-term tests more problematic. For example, a pesticide and a competitor (Daphnia magna) acted in concert over several generations to cause reductions in reproduction or survivorship that led toward a smaller population size facilitating extirpation in Culex pipiens [91], and Kattwinkel and Liess [92] modeled the population recovery time and found slower recovery times for species when in the presence of a competitor. These observations demonstrate that the ultimate goal of ecological risk assessment of understanding the effects of stressors/contaminants on individuals and populations at different spatial scales cannot be reached without an appreciation of and exploration of indirect effects.
6. Indirect Effects and Multiple Contaminants/Stressors
One of the most impenetrable problems facing ecotoxicology is the assessment of risk of multiple stressors that include chemical contaminants [20,93,94]. Groundwater, wastewater, and many soils and sediments are contaminated with complex mixtures of chemicals including metals, hydrocarbons, pesticides, and so-called down-the-drain chemicals [86,95]. The direct toxicological effects of chemicals in the presence of other chemicals are potentially complex and difficult to characterize because they may interact (e.g., synergisms are possible), and interactions are likely context dependent as environmental factors affect outcomes [96]. Indirect effects associated with chemical mixtures are largely unexplored but potentially common. For example, Wei et al. [74] found that nutrients indirectly intensified toxicity risk of metals by ≈14% in sediments. Additional complexities may occur because contaminants in different classes may have opposing direct effects that could lead to contrasting indirect outcomes [58]. Metals and organic contaminants such as poly-nuclear aromatic hydrocarbons and pesticides may slow predator feeding rates, suggesting reduced effects on prey populations [8]. To the contrary, some down-the-drain chemicals reduce predator latency time and increase boldness, suggesting increased effects on prey populations. In addition, contaminants that may co-occur in mixtures may cause changes in sheltering and boldness behaviors in ways that make individuals less or, for other individuals, more susceptible to predation [97]. Exposure to toxicants may also increase metabolic rates associated with biochemical detoxification mechanisms, suggesting increased food consumption and greater effects on prey populations. New techniques, including genomics and gene expression profiles [93], will likely lead to a better understanding of direct contaminant effects in chemical mixtures; however, novel experimental or modeling studies will be necessary to evaluate the significance of possible contrasting influences on indirect effects.
7. Indirect Effects and Experimental Studies
Indirect effects may be inferred from a combination of field and laboratory studies [89]. However, because indirect effects are amenable to experimental investigation, empirical tests provide direct insight into the potential for ecological disruption associated with exposure to potentially harmful chemicals [98]. Therefore, the direct observation of community or ecosystem responses to contaminants is becoming an important avenue to improve estimates of chemical risk [4,99,100], and experimental procedures are included in regulatory protocols in the European Union [11,85]. Experiments that address contaminant effects at the community and ecosystem level can be successfully conducted in the field and with laboratory-based methodologies that are well suited for some, but not all, taxa, habitats, or communities of interest. Some experiments manipulate indigenous (e.g., a soil with its native microbial or faunal assemblage) or naturally colonized communities [98], while others utilize prescribed experimental communities in field settings [101,102] or in microcosms or mesocosms [103]. Experimental approaches offer many advantages; experimental units are usually replicable, exposure concentrations may be controlled, species composition can be manipulated, and experimental designs allow specific hypotheses to be tested. One meaningful way to better predict indirect effects in a given habitat is to utilize data from long-term monitoring programs or intensive field studies to generate hypotheses and to then test them experimentally [104]. Ideally, more long-term field studies will be combined with mesocosm studies, as they are powerful tools to understand community dynamics in the presence of pollutants [105].
Mesocosm experiments are becoming more common and are being conducted on a wide array of taxa, including microbes [106] and in terrestrial and soil communities [107], as well as with newly created chemicals of potential concern [108]. Even mesocosm studies that are based on simplistic, tri-trophic communities, e.g., in freshwater pelagic systems [4,103], have proven successful to elucidate the mechanisms and strength of indirect effects [60]. More complex experimental ecosystems have been implemented over time that, for example, include aquatic plants and benthic invertebrates [100], increasing the ecological realism of experimental tests.
However, mesocosm studies have limitations. Large predators are typically excluded. Many predators are keystone species or strong interactors, and their presence or absence may change the behavior of prey in ways that may affect exposure to contaminants [38]. Mesocosms usually do not permit colonization, reducing biological realism because colonization is a principal mechanism that promotes resiliency after a disturbance [109]. Dispersal is also essential to study indirect effects associated with spatially structured populations because the so-called action-at-a-distance pollution effects expressed in meta-communities are a function of the rate and frequency of dispersal [110]. Many experiments are not long enough to take into account factors such as the relatively slow uptake of highly hydrophobic contaminants into food or soils and sediments before entering the food chain to biomagnify, and contaminant effects on sensitive early life stages cannot be detected in short-term experiments that do not support reproduction [105]. In addition, the environment in experimental systems cannot be perfectly mimicked, and some species may experience altered behavior or reproduction e.g., feeding rates may change in the absence of a complex environment or reproductive output can be affected [111]. Given the importance of behavior to species interactions that lead to indirect effects, this “mesocosm effect” may alter the outcome of interactions in mesocosms compared to natural systems, altering indirect effects.
8. Indirect Effects and Models
Mechanistic effects models, including organism-level, population-level, and multispecies models (i.e., food web and ecosystem models) may provide great insight into and important predictions of indirect effects [86]. Various models have been developed, many as an aid to ecological risk assessment, and they may be used individually or in concert to extrapolate the impacts of chemicals across levels of biological organization and spatial scales [26,112,113,114]. However, overall, relatively few mechanistic models have addressed higher levels of biological organization. For example, Galic et al. [26] identified 18 different models that could be used to assess indirect effects, although the number of models, case studies, and ecological scenarios investigated is growing [112,113]. Models are often intended to reduce unexpected outcomes (i.e., ecological surprises) when chemicals enter the environment, and they are often used to examine worst-case scenarios of chemical effects [11]. They will likely grow in prominence and frequency as methods and approaches are improved [90], and as models are developed for particular chemicals, habitats or taxa, e.g., [115,116].
Models used to predict indirect effects require a sophisticated mechanistic understanding of the ecosystem under study, and therefore must include working knowledge of the functioning and underlying processes of the system as a whole to make appropriate assumptions and to parameterize models. Thus, the principal limitation of models is an inadequate knowledge of key processes and species interactions, which remain understudied in most ecosystems. Furthermore, in order to confirm the suitability for a specific application, model outputs should be compared with empirical data to determine if patterns observed in the model generate patterns observed in the field [113]. Since large amounts of data are required to parameterize effects models that are able to encompass the complexity of ecosystems, models may be best applied to well-studied, targeted ecosystems and/or focused on questions about vulnerable species. Some factors have been slow to be included in models; e.g., contaminant effects on behavior are not considered in most ecosystem models [12].
Simulations may also be used to determine the factors that contribute to indirect effects. De Laender et al. [12] combined a chemical fate model with an ecosystem/trophic model to explore indirect effects in different ecological scenarios using hypothetical communities. The goal was to simulate factors such as species interaction effects size, trophic state, contaminant chemical properties, and species sensitivities. The results suggest that indirect effects will likely vary with natural variation in communities over space and time and that various ecological scenarios should be tested when using models to achieve a better understanding of ecological risk. The choice of and number of scenarios tested by mechanistic models should be extensive enough to ensure ecological realism associated with indirect effects [11]. Finally, an integration of mechanistic models and experimental studies have a great potential for insight into indirect effects because experimental studies, especially with well-studied toxicants such as pesticides or in habitats in which there is a history of successful experimentation, may provide important data needed to validate models, and models can be used to interpret effects observed in experimental studies [26,109].
9. Final Comments on Indirect Effects
Indirect effects of chemical contaminants are difficult to anticipate and predict, because biotic communities and ecosystems are complex and difficult to study. As a result, species interaction networks are poorly known, and the factors that drive ecosystem function in the presence and/or absence of anthropogenic influences remain understudied in most ecosystems. Populations, communities, and ecosystems may display nonlinear dynamics that can change over space and time. Functional redundancy among species means that species may be lost without obvious impacts on ecosystem function or services. Alternatively, impacts to species that strongly interact with other species (e.g., keystone species or superior competitors) can lead to disproportionately large changes in ecosystem properties if they are relatively intolerant of a chemical. Ecosystems may also exhibit threshold behavior where incremental change or disturbance pushes the system beyond a threshold and result in dramatic changes that are not easily reversible. Thus, indirect effects are also difficult to quantify, and they will likely change with variation in species composition, trophic structure, ecological function, and environmental factors over space and time.
Indirect effects of contaminants inform ecotoxicology in ways that help avoid ecological surprises and better estimate chemical and pollutant risk. However, ecologists, ecotoxicologists, and risk analysts must continue to foster research on several fronts to further integrate indirect effects into the fabric of ecotoxicology and strengthen our understanding of contaminant effects on populations, communities, and ecosystems. Most research in ecotoxicology has been at the individual-effects level [117], and methods to improve toxicity models continue to be developed. For example, models based on species-level traits and bioenergetics have the potential to improve predictions of species-level responses to various chemicals [114,118], and databases associated with toxic effects that include traditional toxicological information and the results of toxicogenomic or other molecular methods that identify the mode of toxic action are being developed, in some instances with computational modeling [119,120]. Information on direct effects on individuals will improve population models that estimate safe exposure limits for species, representing an important starting place to predict indirect effects. Nevertheless, research on higher levels of organization has been much slower to take place, and it is needed to quantify species interactions and ecosystem functions necessary to assess the potential for indirect effects. Monitoring programs and long-term research in areas of concern (e.g., areas sensitive to oiling) to provide background data on communities would contribute to a better anticipation of indirect effects and stimulate experimental investigation to test hypotheses. Continued refinements of field and mesocosm experimental procedures will also be helpful to detect indirect effects in specific ecosystems. Ecosystem models have provided important contributions to conceptualize the nature of indirect effects but have limitations. For example, Grechi et al. [64] report that better information associated with interaction networks, i.e., feeding preferences and abundance over time of representative species within functional groups, would be of great help to refine the parameterization and calibration of an Aquatox model for contaminant direct and indirect effects. Species interaction networks (also called network ecology) are often poorly known in many natural communities. Interaction networks provide insight into niche-based community assembly by reflecting the complexity of species interactions and underlying ecosystem processes, and they have been used as the basis for risk assessment in the biological control of pest species [121]. More emphasis on quantifying network ecology should serve as a goal for basic research. Finally, models of the physical/chemical environment, individual-based effects models, and ecosystem/food web models should be tested with many ecological contingencies and integrated to achieve better predictions [122].
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Fleeger, J.W.; Carman, K.R.; Nisbet, R.M. Indirect effects of contaminants on aquatic ecosystems. Sci. Total Environ. 2003, 317, 207–233. [Google Scholar] [CrossRef]
- Clements, W.H.; Rohr, J.R. Community responses to contaminants: Using basic ecological principles to predict ecotoxicological effects. Environ. Toxicol. Chem. 2009, 28, 1789–1800. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Kerby, J.L.; Sih, A. Community ecology as a framework for predicting contaminant effects. Trends Ecol. Evol. 2006, 21, 606–613. [Google Scholar] [CrossRef]
- Halstead, N.T.; McMahon, T.A.; Johnson, S.A.; Raffel, T.R.; Romansic, J.M.; Crumrine, P.W.; Rohr, J.R. Community ecology theory predicts the effects of agrochemical mixtures on aquatic biodiversity and ecosystem properties. Ecol. Lett. 2014, 17, 932–941. [Google Scholar] [CrossRef]
- Relyea, R.; Hoverman, J. Assessing the ecology in ecotoxicology: A review and synthesis in freshwater systems. Ecol. Lett. 2006, 9, 1157–1171. [Google Scholar] [CrossRef]
- Schmitt-Jansen, M.; Veit, U.; Dudel, G.; Altenburger, R. An ecological perspective in aquatic ecotoxicology: Approaches and challenges. Basic Appl. Ecol. 2008, 9, 337–345. [Google Scholar] [CrossRef]
- Preston, B.L. Indirect effects in aquatic ecotoxicology: Implications for ecological risk assessment. Environ. Manag. 2002, 29, 311–323. [Google Scholar] [CrossRef]
- Saaristo, M.; Brodin, T.; Balshine, S.; Bertram, M.G.; Brooks, B.W.; Ehlman, S.M.; McCallum, E.S.; Sih, A.; Sundin, J.; Wong, B.B.M.; et al. Direct and indirect effects of chemical contaminants on the behaviour, ecology and evolution of wildlife. Proc. R. Soc. B-Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Kidd, K.A.; Paterson, M.J.; Rennie, M.D.; Podemski, C.L.; Findlay, D.L.; Blanchfield, P.J.; Liber, K. Direct and indirect responses of a freshwater food web to a potent synthetic oestrogen. Philos. Trans. R. Soc. B-Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [Green Version]
- Baert, J.M.; De Laender, F.; Janssen, C.R. The Consequences of nonrandomness in species-sensitivity in relation to functional traits for ecosystem-level effects of chemicals. Environ. Sci. Technol. 2017, 51, 7228–7235. [Google Scholar] [CrossRef]
- Rico, A.; Van den Brink, P.J.; Gylstra, R.; Focks, A.; Brock, T.C.M. Developing ecological scenarios for the prospective aquatic risk assessment of pesticides. Integr. Environ. Assess. Manag. 2016, 12, 510–521. [Google Scholar] [CrossRef]
- De Laender, F.; Morselli, M.; Baveco, H.; Van den Brink, P.J.; Di Guardo, A. Theoretically exploring direct and indirect chemical effects across ecological and exposure scenarios using mechanistic fate and effects modelling. Environ. Int. 2015, 74, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zang, W.; Yan, Z.; Hong, Y.; Liu, Z.; Yi, X.; Wang, X.; Liu, T.; Zhou, L. Species sensitivity analysis of heavy metals to freshwater organisms. Ecotoxicology 2015, 24, 1621–1631. [Google Scholar] [CrossRef]
- Long, E.R. Ranges in chemical concentrations in sediments associated with adverse biological effects. Mar. Poll. Bull. 1992, 24, 38–45. [Google Scholar] [CrossRef]
- Zhao, Q.H.; De Laender, F.; Van den Brink, P.J. Community composition modifies direct and indirect effects of pesticides in freshwater food webs. Sci. Total Environ. 2020, 739. [Google Scholar] [CrossRef]
- Delmas, E.; Besson, M.; Brice, M.H.; Burkle, L.A.; Dalla Riva, G.V.; Fortin, M.J.; Gravel, D.; Guimaraes, P.R.; Hembry, D.H.; Newman, E.A.; et al. Analysing ecological networks of species interactions. Biol. Rev. 2019, 94, 16–36. [Google Scholar] [CrossRef] [Green Version]
- Baert, J.M.; De Laender, F.; Sabbe, K.; Janssen, C.R. Biodiversity increases functional and compositional resistance, but decreases resilience in phytoplankton communities. Ecology 2016, 97, 3433–3440. [Google Scholar] [CrossRef]
- Carman, K.R.; Fleeger, J.W.; Pomarico, S. Response of a benthic food web to hydrocarbon contamination. Limnol. Oceanogr. 1997, 42, 561–571. [Google Scholar] [CrossRef]
- Van den Brink, P.J.; Bracewell, S.A.; Bush, A.; Chariton, A.; Choung, C.B.; Compson, Z.G.; Dafforn, K.A.; Korbel, K.; Lapen, D.R.; Mayer-Pinto, M.; et al. Towards a general framework for the assessment of interactive effects of multiple stressors on aquatic ecosystems: Results from the Making Aquatic Ecosystems Great Again (MAEGA) workshop. Sci. Total Environ. 2019, 684, 722–726. [Google Scholar] [CrossRef]
- Orr, J.A.; Vinebrooke, R.D.; Jackson, M.C.; Kroeker, K.J.; Kordas, R.L.; Mantyka-Pringle, C.; Van den Brink, P.J.; De Laender, F.; Stoks, R.; Holmstrup, M.; et al. Towards a unified study of multiple stressors: Divisions and common goals across research disciplines. Proc. R. Soc. B-Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Rohr, J.R.; Schotthoefer, A.M.; Raffel, T.R.; Carrick, H.J.; Halstead, N.; Hoverman, J.T.; Johnson, C.M.; Johnson, L.B.; Lieske, C.; Piwoni, M.D.; et al. Agrochemicals increase trematode infections in a declining amphibian species. Nature 2008, 455, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Man, Y.B.; Mo, W.Y.; Man, K.Y.; Wong, M.H. Direct and indirect effects of microplastics on bivalves, with a focus on edible species: A mini-review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2109–2143. [Google Scholar] [CrossRef]
- Zhai, Y.; Brun, N.R.; Bundschuh, M.; Schrama, M.; Hin, E.; Vijver, M.G.; Hunting, E.R. Microbially-mediated indirect effects of silver nanoparticles on aquatic invertebrates. Aquat. Sci. 2018, 80. [Google Scholar] [CrossRef] [Green Version]
- Gredelj, A.; Barausse, A.; Grechi, L.; Palmeri, L. Deriving predicted no-effect concentrations (PNECs) for emerging contaminants in the river Po, Italy, using three approaches: Assessment factor, species sensitivity distribution and AQUATOX ecosystem modelling. Environ. Int. 2018, 119, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Relyea, R. Chemical cocktails in aquatic systems: Pesticide effects on the response and recovery of >20 animal taxa. Environ. Poll. 2014, 189, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Galic, N.; Hommen, U.; Baveco, J.; van den Brink, P.J. Potential application of population models in the European ecological risk assessment of chemicals II: Review of models and their potential to address environmental protection aims. Integr. Environ. Assess. Manag. 2010, 6, 338–360. [Google Scholar] [CrossRef] [PubMed]
- Agerstrand, M.; Arnold, K.; Balshine, S.; Brodin, T.; Brooks, B.W.; Maack, G.; McCallum, E.S.; Pyle, G.; Saaristo, M.; Ford, A.T. Emerging investigator series: Use of behavioural endpoints in the regulation of chemicals. Environ. Sci. Process. Impacts 2020, 22, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Sievers, M.; Hale, R.; Parris, K.M.; Melvin, S.D.; Lanctot, C.M.; Swearer, S.E. Contaminant-induced behavioural changes in amphibians: A meta-analysis. Sci. Total Environ. 2019, 693. [Google Scholar] [CrossRef]
- Brodin, T.; Piovano, S.; Fick, J.; Klaminder, J.; Heynen, M.; Jonsson, M. Ecological effects of pharmaceuticals in aquatic systems-impacts through behavioural alterations. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130580. [Google Scholar] [CrossRef] [Green Version]
- Jacquin, L.; Petitjean, Q.; Cote, J.; Laffaille, P.; Jean, S. Effects of pollution on fish behavior, personality, and cognition: Some research perspectives. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Sousa, B.; Nunes, B. Reliability of behavioral test with fish: How neurotransmitters may exert neuromodulatory effects and alter the biological responses to neuroactive agents. Sci. Total Environ. 2020, 734, 139372. [Google Scholar] [CrossRef] [PubMed]
- Tierney, K.B.; Baldwin, D.H.; Hara, T.J.; Ross, P.S.; Scholz, N.L.; Kennedy, C.J. Olfactory toxicity in fishes. Aquat. Toxicol. 2010, 96, 2–26. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.; Dabrowski, J.M. Combined effects of predatory fish and sublethal pesticide contamination on the behavior and mortality of mayfly nymphs. Environ. Toxicol. Chem. 2001, 20, 2537–2543. [Google Scholar] [CrossRef] [PubMed]
- Neuman-Lee, L.A.; Hopkins, G.R.; Brodie, E.D.; French, S.S. Sublethal contaminant exposure alters behavior in a common insect: Important implications for trophic transfer. J. Environ. Sci. Health Part B 2013, 48, 442–448. [Google Scholar] [CrossRef]
- Brodin, T.; Fick, J.; Jonsson, M.; Klaminder, J. Dilute concentrations of a psychiatric drug alter behavior of fish from natural populations. Science 2013, 339, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Jacquin, L.; Gandar, A.; Aguirre-Smith, M.; Perrault, A.; Le Henaff, M.; De Jong, L.; Paris-Palacios, S.; Laffaille, P.; Jean, S. High temperature aggravates the effects of pesticides in goldfish. Ecotoxicol. Environ. Saf. 2019, 172, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Lagesson, A.; Saaristo, M.; Brodin, T.; Fick, J.; Kiaminder, J.; Martin, J.M.; Wong, B.B.M. Fish on steroids: Temperature-dependent effects of 17 beta-trenbolone on predator escape, boldness, and exploratory behaviors. Environ. Poll. 2019, 245, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Araujo, C.V.M.; Pereira, K.C.; Sparaventi, E.; Gonzalez-Ortegon, E.; Blasco, J. Contamination may induce behavioural plasticity in the habitat selection by shrimps: A cost-benefits balance involving contamination, shelter and predation. Environ. Poll. 2020, 263. [Google Scholar] [CrossRef]
- Gessner, M.O.; Tlili, A. Fostering integration of freshwater ecology with ecotoxicology. Freshwat. Biol. 2016, 61, 1991–2001. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.N.; Llanos, J.E.M.; Kunin, W.E.; Evison, S.E.F. Indirect effects of agricultural pesticide use on parasite prevalence in wild pollinators. Agric. Ecosyst. Environ. 2018, 258, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Bontje, D.; Kooi, B.W.; van Hattum, B. Sublethal toxic effects in a generic aquatic ecosystem. Ecotoxicol. Environ. Saf. 2011, 74, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Schuler, M.S.; Hintz, W.D.; Stoler, A.B.; Jones, D.K.; Mattes, B.M.; Relyea, R.A. Salty fertile lakes: How salinization and eutrophication alter the structure of freshwater communities. Ecosphere 2018, 9, 19. [Google Scholar] [CrossRef]
- Thrupp, T.J.; Runnalls, T.J.; Scholze, M.; Kugathas, S.; Kortenkamp, A.; Sumpter, J.P. The consequences of exposure to mixtures of chemicals: Something from ‘nothing’ and ‘a lot from a little’ when fish are exposed to steroid hormones. Sci. Total Environ. 2018, 619-620, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Graymore, M.; Stagnitti, F.; Allinson, G. Impacts of atrazine in aquatic ecosystems. Environ. Int. 2001, 26, 483–495. [Google Scholar] [CrossRef]
- Groner, M.L.; Relyea, R.A. A tale of two pesticides: How common insecticides affect aquatic communities. Freshwat. Biol. 2011, 56, 2391–2404. [Google Scholar] [CrossRef]
- Mensens, C.; De Laender, F.; Janssen, C.R.; Rivera, F.C.; Sabbe, K.; De Troch, M. Selective and context-dependent effects of chemical stress across trophic levels at the basis of marine food webs. Ecol. Appl. 2018, 28, 1342–1353. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.M.; Ganatra, A.A.; Kandie, F.; Muhlbauer, L.; Ahlheim, J.; Brack, W.; Torto, B.; Agola, E.L.; McOdimba, F.; Hollert, H.; et al. Pesticide pollution in freshwater paves the way for schistosomiasis transmission. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Ann. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Jordan, F.; Gjata, N.; Mei, S.; Yule, C.M. Simulating food web dynamics along a gradient: Quantifying human influence. PLoS ONE 2012, 7, e40280. [Google Scholar] [CrossRef] [Green Version]
- Rasher, D.B.; Steneck, R.S.; Halfar, J.; Kroeker, K.J.; Ries, J.B.; Tinker, M.T.; Chan, P.T.W.; Fietzke, J.; Kamenos, N.A.; Konar, B.H.; et al. Keystone predators govern the pathway and pace of climate impacts in a subarctic marine ecosystem. Science 2020, 369, 1351–1354. [Google Scholar] [CrossRef] [PubMed]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, D.L.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, T.C.; Burkepile, D.E.; Holbrook, S.J.; Carpenter, R.C.; Claudet, J.; Loiseau, C.; Thiault, L.; Brooks, A.J.; Washburn, L.; Schmitt, R.J. Landscape-scale patterns of nutrient enrichment in a coral reef ecosystem: Implications for coral to algae phase shifts. Ecol. Appl. 2020, e2227. [Google Scholar] [CrossRef]
- Pimentao, A.R.; Pascoal, C.; Castro, B.B.; Cassio, F. Fungistatic effect of agrochemical and pharmaceutical fungicides on non-target aquatic decomposers does not translate into decreased fungi- or invertebrate-mediated decomposition. Sci. Total Environ. 2020, 712, 10. [Google Scholar] [CrossRef] [PubMed]
- Pereda, O.; Solagaistua, L.; Atristain, M.; de Guzman, L.; Larranaga, A.; von Schiller, D.; Elosegi, A. Impact of wastewater effluent pollution on stream functioning: A whole-ecosystem manipulation experiment. Environ. Poll. 2020, 258. [Google Scholar] [CrossRef] [PubMed]
- Deegan, L.A.; Johnson, D.S.; Warren, R.S.; Peterson, B.J.; Fleeger, J.W.; Fagherazzi, S.; Wollheim, W.M. Coastal eutrophication as a driver of salt marsh loss. Nature 2012, 490, 388–394. [Google Scholar] [CrossRef]
- Johnston, E.L.; Mayer-Pinto, M.; Crowe, T.P. Chemical contaminant effects on marine ecosystem functioning. J. Appl. Ecol. 2015, 52, 140–149. [Google Scholar] [CrossRef] [Green Version]
- De Laender, F.; Janssen, C.R. Brief communication: The ecosystem perspective in ecotoxicology as a way forward for the ecological risk assessment of chemicals. Integr. Environ. Assess. Manag. 2013, 9, E34–E38. [Google Scholar] [CrossRef]
- Lenihan, H.S.; Peterson, C.H.; Miller, R.J.; Kayal, M.; Potoski, M. Biotic disturbance mitigates effects of multiple stressors in a marine benthic community. Ecosphere 2018, 9. [Google Scholar] [CrossRef]
- Beketov, M.A.; Liess, M. Ecotoxicology and macroecology—Time for integration. Environ. Poll. 2012, 162, 247–254. [Google Scholar] [CrossRef]
- Chagnon, M.; Kreutzweiser, D.; Mitchell, E.A.; Mitchell, E.A.; Morrissey, C.A.; Noome, D.A.; Noome, D.A.; Van der Sluijs, J.P.; Van der Sluijs, J.P. Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ. Sci. Pollut. Res. 2015, 22, 119–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, K.; Bundschuh, M.; Schäfer, R.B. Review on the effects of toxicants on freshwater ecosystem functions. Environ. Poll. 2013, 180, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Markandya, A.; Taylor, T.; Longo, A.; Murty, M.N.; Murty, S.; Dhavala, K. Counting the cost of vulture decline - An appraisal of the human health and other benefits of vultures in India. Ecol. Econ. 2008, 67, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Grechi, L.; Franco, A.; Palmeri, L.; Pivato, A.; Barausse, A. An ecosystem model of the lower Po river for use in ecological risk assessment of xenobiotics. Ecol. Modell. 2016, 332, 42–58. [Google Scholar] [CrossRef]
- Lombardo, A.; Franco, A.; Pivato, A.; Barausse, A. Food web modeling of a river ecosystem for risk assessment of down-the-drain chemicals: A case study with AQUATOX. Sci. Total Environ. 2015, 508, 214–227. [Google Scholar] [CrossRef]
- Zhang, L.L.; Liu, J.L. AQUATOX coupled foodweb model for ecosystem risk assessment of Polybrominated diphenyl ethers (PBDEs) in lake ecosystems. Environ. Poll. 2014, 191, 80–92. [Google Scholar] [CrossRef]
- Rodrigues, A.C.M.; Machado, A.L.; Bordalo, M.D.; Saro, L.; Simao, F.C.P.; Rocha, R.J.M.; Golovko, O.; Zlabek, V.; Barata, C.; Soares, A.; et al. Invasive species mediate insecticide effects on community and ecosystem functioning. Environ. Sci. Technol. 2018, 52, 4889–4900. [Google Scholar] [CrossRef]
- Chagaris, D.D.; Patterson, W.F.; Allen, M.S. Relative effects of multiple stressors on reef food webs in the northern Gulf of Mexico revealed via ecosystem modeling. Front. Mar. Sci. 2020, 7, 17. [Google Scholar] [CrossRef]
- Macneale, K.H.; Kiffney, P.M.; Scholz, N.L. Pesticides, aquatic food webs, and the conservation of Pacific salmon. Front. Ecol. Environ. 2010, 8, 475–482. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Sivakoff, F.S.; Cayton, H.L.; Morris, W.F.; Haddad, N.M. Habitat restoration affects immature stages of a wetland butterfly through indirect effects on predation. Ecology 2015, 96, 1761–1767. [Google Scholar] [CrossRef]
- Watts, C.; Thornburrow, D.; Cave, V. Responses of invertebrates to herbicide in Salix cinerea invaded wetlands: Restoration implications. Ecol. Manag. Restor. 2016, 17, 243–249. [Google Scholar] [CrossRef]
- Peterson, C.H.; Rice, S.D.; Short, J.W.; Esler, D.; Bodkin, J.L.; Ballachey, B.E.; Irons, D.B. Long-term ecosystem response to the Exxon Valdez oil spill. Science 2003, 302, 2082–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hook, S.E. Beyond thresholds: A holistic approach to impact assessment is needed to enable accurate predictions of environmental risk from oil spills. Integr. Environ. Assess. Manag. 2020, 16, 813–830. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhang, H.; Yuan, Y.; Zhao, Y.; Li, G.; Zhang, F. Indirect effect of nutrient accumulation intensified toxicity risk of metals in sediments from urban river network. Environ. Sci. Pollut. Res. 2020, 27, 6193–6204. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.J.; De Schamphelaere, K.; Clements, W.H.; Sorensen, M.T.; Van den Brink, P.J.; Liess, M. Combined and interactive effects of global climate change and toxicants on populations and communities. Environ. Toxicol. Chem. 2013, 32, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, M.; Hale, R.; Parris, K.M.; Swearer, S.E. Impacts of human-induced environmental change in wetlands on aquatic animals. Biol. Rev. Camb. Philos. Soc. 2018, 93, 529–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, S.P.; Grabowski, J.H.; Roman, H.; Geggel, A.; Rouhani, S.; Oehrig, J.; Baker, M. Consequences of large-scale salinity alteration during the Deepwater Horizon oil spill on subtidal oyster populations. Mar. Ecol. Prog. Ser. 2017, 576, 175–187. [Google Scholar] [CrossRef]
- Deis, D.R.; Fleeger, J.W.; Johnson, D.S.; Mendelssohn, I.A.; Lin, Q.; Graham, S.A.; Zengel, S.; Hou, A. Recovery of the salt marsh periwinkle (Littoraria irrorata) 9 years after the Deepwater Horizon oil spill: Size matters. Mar. Poll. Bull. 2020, 160, 111581. [Google Scholar] [CrossRef]
- Fleeger, J.W.; Johnson, D.S.; Zengel, S.; Mendelssohn, I.A.; Deis, D.R.; Graham, S.A.; Lin, Q.; Christman, M.C.; Riggio, M.R.; Pant, M. Macroinfauna responses and recovery trajectories after an oil spill differ from those following saltmarsh restoration. Mar. Environ. Res. 2020, 155. [Google Scholar] [CrossRef]
- Nadal, M.; Marques, M.; Mari, M.; Domingo, J.L. Climate change and environmental concentrations of POPs: A review. Environ. Res. 2015, 143, 177–185. [Google Scholar] [CrossRef]
- Pincebourde, S.; van Baaren, J.; Rasmann, S.; Rasmont, P.; Rodet, G.; Martinet, B.; Calatayud, P.A. Plant-insect interactions in a changing world. In Insect-Plant Interactions in a Crop Protection Perspective; Sauvion, N., Thiery, D., Calatayud, P.A., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 81, pp. 289–332. [Google Scholar]
- Sumner, A.W.; Johnston, T.A.; Lescord, G.L.; Branfireun, B.A.; Gunn, J.M. Mercury Bioaccumulation in lacustrine fish populations along a climatic gradient in northern Ontario, Canada. Ecosystems 2019. [Google Scholar] [CrossRef]
- Bates, M.L.; Nash, S.M.B.; Hawker, D.W.; Shaw, E.C.; Cropp, R.A. The distribution of persistent organic pollutants in a trophically complex Antarctic ecosystem model. J. Mar. Syst. 2017, 170, 103–114. [Google Scholar] [CrossRef]
- Cambronero, M.C.; Marshall, H.; De Meester, L.; Davidson, T.A.; Beckerman, A.P.; Orsini, L. Predictability of the impact of multiple stressors on the keystone species Daphnia. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Van den Brink, P.J. Ecological risk assessment: From book-keeping to chemical stress ecology. Environ. Sci. Technol. 2008, 42, 8999–9004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, A.; Price, O.R.; Marshall, S.; Jolliet, O.; Van den Brink, P.J.; Rico, A.; Focks, A.; De Laender, F.; Ashauer, R. Toward refined environmental scenarios for ecological risk assessment of down-the-drain chemicals in freshwater environments. Integr. Environ. Assess. Manag. 2017, 13, 233–248. [Google Scholar] [CrossRef] [Green Version]
- Fodrie, F.J.; Able, K.W.; Galvez, F.; Heck, K.L.; Jensen, O.P.; Lopez-Duarte, P.C.; Martin, C.W.; Turner, R.E.; Whitehead, A. Integrating organismal and population responses of estuarine fishes in Macondo spill research. Bioscience 2014, 64, 778–788. [Google Scholar] [CrossRef]
- Whitehead, A.; Dubansky, B.; Bodinier, C.; Garcia, T.I.; Miles, S.; Pilley, C.; Raghunathan, V.; Roach, J.L.; Walker, N.; Walter, R.B.; et al. Genomic and physiological footprint of the Deepwater Horizon oil spill on resident marsh fishes. Proc. Nat. Acad. Sci. USA 2012, 109, 20298–20302. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.W.; Lewis, K.A.; McDonald, A.M.; Spearman, T.P.; Alford, S.B.; Christian, R.C.; Valentine, J.F. Disturbance-driven changes to northern Gulf of Mexico nekton communities following the Deepwater Horizon oil spill. Mar. Poll. Bull. 2020, 155, 111098. [Google Scholar] [CrossRef]
- De Laender, F. Community- and ecosystem-level effects of multiple environmental change drivers: Beyond null model testing. Glob. Chang. Biol. 2018, 24, 5021–5030. [Google Scholar] [CrossRef] [Green Version]
- Liess, M.; Foit, K.; Becker, A.; Hassold, E.; Dolciotti, I.; Kattwinkel, M.; Duquesne, S. Culmination of low-dose pesticide effects. Environ. Sci. Technol. 2013, 47, 8862–8868. [Google Scholar] [CrossRef]
- Kattwinkel, M.; Liess, M. Competition matters: Species interactions prolong the long-term effects of pulsed toxicant stress on populations. Environ. Toxicol. Chem. 2014, 33, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Van den Brink, P.J.; Choung, C.B.; Landis, W.; Mayer-Pinto, M.; Pettigrove, V.; Scanes, P.; Smith, R.; Stauber, J. New approaches to the ecological risk assessment of multiple stressors. Mar. Freshwat. Res. 2016, 67, 429–439. [Google Scholar] [CrossRef]
- Galic, N.; Sullivan, L.L.; Grimm, V.; Forbes, V.E. When things don’t add up: Quantifying impacts of multiple stressors from individual metabolism to ecosystem processing. Ecol. Lett. 2018, 21, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Gobel, P.; Dierkes, C.; Coldewey, W.C. Storm water runoff concentration matrix for urban areas. J. Contam. Hydrol. 2007, 91, 26–42. [Google Scholar] [CrossRef]
- Laskowski, R.; Bednarska, A.J.; Kramarz, P.E.; Loureiro, S.; Scheil, V.; Kudłek, J.; Holmstrup, M. Interactions between toxic chemicals and natural environmental factors—A meta-analysis and case studies. Sci. Total Environ. 2010, 408, 3763–3774. [Google Scholar] [CrossRef]
- Sievers, M.; Hale, R.; Swearer, S.E.; Parris, K.M. Contaminant mixtures interact to impair predator-avoidance behaviours and survival in a larval amphibian. Ecotoxicol. Environ. Saf. 2018, 161, 482–488. [Google Scholar] [CrossRef]
- Kotalik, C.J.; Cadmus, P.; Clements, W.H. Indirect effects of iron oxide on stream benthic communities: Capturing ecological complexity with controlled mesocosm experiments. Environ. Sci. Technol. 2019, 53, 11532–11540. [Google Scholar] [CrossRef]
- Boyle, T.P.; Fairchild, J.F. The role of mesocosm studies in ecological risk analysis. Ecol. Appl. 1997, 7, 1099–1102. [Google Scholar] [CrossRef]
- Clemow, Y.H.; Manning, G.E.; Breton, R.L.; Winchell, M.F.; Padilla, L.; Rodney, S.I.; Hanzas, J.P.; Estes, T.L.; Budreski, K.; Toth, B.N.; et al. A Refined ecological risk assessment for California red-legged frog, delta smelt, and California tiger salamander exposed to malathion. Integr. Environ. Assess. Manag. 2018, 14, 224–239. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, C.A.; Crawley, M.J.; Wright, D.J.; Kuczynski, J.; Robinson, L.; Knight, R.; Abu Al-Soud, W.; Sorensen, S.J.; Deng, Y.; Zhou, J.Z.; et al. Identifying qualitative effects of different grazing types on below-ground communities and function in a long-term field experiment. Environ. Microbiol. 2015, 17, 841–854. [Google Scholar] [CrossRef]
- Gutierrez, Y.; Ott, D.; Scherber, C. Direct and indirect effects of plant diversity and phenoxy herbicide application on the development and reproduction of a polyphagous herbivore. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Riedl, V.; Agatz, A.; Benstead, R.; Ashauer, R. A standardized tritrophic small-scale system (tricosm) for the assessment of stressor-induced effects on aquatic community dynamics. Environ. Toxicol. Chem. 2018, 37, 1051–1060. [Google Scholar] [CrossRef]
- Duggan, S.B.; Kotalik, C.J.; Clements, W.H. Integrating results of field biomonitoring and mesocosm experiments to validate postspill impacts of petroleum hydrocarbons on stream benthic communities. Environ. Sci. Technol. 2018, 52, 13584–13590. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, Y.; Schmidt, T.S.; Clements, W.H. Quantifying differences in responses of aquatic insects to trace metal exposure in field studies and short-term stream mesocosm experiments. Environ. Sci. Technol. 2018, 52, 4378–4384. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Dos Santos, S.P.; Gustavsen, J.A.; Imfeld, G.; Lamy, F.; Mitchell, E.A.D.; Mota, M.; Noll, D.; Planchamp, C.; Heger, T.J. Impact of a synthetic fungicide (fosetyl-Al and propamocarb-hydrochloride) and a biopesticide (Clonostachys rosea) on soil bacterial, fungal, and protist communities. Sci. Total Environ. 2020, 738. [Google Scholar] [CrossRef]
- Helander, M.; Pauna, A.; Saikkonen, K.; Saloniemi, I. Glyphosate residues in soil affect crop plant germination and growth. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Auffan, M.; Masion, A.; Mouneyrac, C.; de Garidel-Thoron, C.; Hendren, C.O.; Thiery, A.; Santaella, C.; Giamberini, L.; Bottero, J.-Y.; Wiesner, M.R.; et al. Contribution of mesocosm testing to a single-step and exposure-driven environmental risk assessment of engineered nanomaterials. Nanoimpact 2019, 13, 66–69. [Google Scholar] [CrossRef]
- Traas, T.P.; Janse, J.H.; van den Brink, P.J.; Brock, T.C.M.; Aldenberg, T. A freshwater food web model for the combined effects of nutrients and insecticide stress and subsequent recovery. Environ. Toxicol. Chem. 2004, 23, 521–529. [Google Scholar] [CrossRef]
- Spromberg, J.A.; John, B.M.; Landis, W.G. Metapopulation dynamics: Indirect effects and multiple distinct outcomes in ecological risk assessment. Environ. Toxicol. Chem. 1998, 17, 1640–1649. [Google Scholar] [CrossRef]
- Howick, G.L.; Giddings, J.M.; Denoyelles, F.; Ferrington, L.C.; Kettle, W.D.; Baker, D. Rapid establishment of test conditions and trophic-level interactions in 0.04-hectare earthen pond mesocosms. Environ. Toxicol. Chem. 1992, 11, 107–114. [Google Scholar] [CrossRef]
- Forbes, V.E.; Salice, C.J.; Birnir, B.; Bruins, R.J.F.; Calow, P.; Ducrot, V.; Galic, N.; Garber, K.; Harvey, B.C.; Jager, H.; et al. A framework for predicting impacts on ecosystem services from (sub)organismal responses to chemicals. Environ. Toxicol. Chem. 2017, 36, 845–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galic, N.; Schmolke, A.; Forbes, V.; Baveco, H.; van den Brink, P.J. The role of ecological models in linking ecological risk assessment to ecosystem services in agroecosystems. Sci. Total Environ. 2012, 415, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Van den Brink, P.J.; Baird, D.J.; Baveco, H.; Focks, A. The use of traits-based approaches and eco(toxico)logical models to advance the ecological risk assessment framework for chemicals. Integr. Environ. Assess. Manag. 2013, 9, E47–E57. [Google Scholar] [CrossRef]
- Strona, G.; Fattorini, S.; Fiasca, B.; Di Lorenzo, T.; Di Cicco, M.; Lorenzetti, W.; Boccacci, F.; Galassi, D.M.P. AQUALIFE software: A new tool for a standardized ecological assessment of groundwater dependent ecosystems. Water 2019, 11, 2574. [Google Scholar] [CrossRef] [Green Version]
- Etterson, M.; Garber, K.; Odenkirchen, E. Mechanistic modeling of insecticide risks to breeding birds in North American agroecosystems. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.C.; Jin, X.; Nakada, N.; Sumpter, J.P. Learning from the past and considering the future of chemicals in the environment. Science 2020, 367, 384–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, A.; Van den Brink, P.J. Evaluating aquatic invertebrate vulnerability to insecticides based on intrinsic sensitivity, biological traits, and toxic mode of action. Environ. Toxicol. Chem. 2015, 34, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Xia, P.; Wang, P.P.; Yang, J.H.; Baird, D.J. Omics advances in ecotoxicology. Environ. Sci. Technol. 2018, 52, 3842–3851. [Google Scholar] [CrossRef]
- Ciallella, H.L.; Russo, D.P.; Aleksunes, L.M.; Grimm, F.A.; Zhu, H. Predictive modeling of estrogen receptor agonism, antagonism, and binding activities using machine- and deep-learning approaches. Lab. Investig. 2020. [Google Scholar] [CrossRef]
- Ollivier, M.; Lesieur, V.; Raghu, S.; Martin, J.F. Characterizing ecological interaction networks to support risk assessment in classical biological control of weeds. Curr. Opin. Insect Sci. 2020, 38, 40–47. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Kong, X.Z.; Qin, N.; He, Q.S.; Liu, W.X.; Bai, Z.L.; Wang, Y.; Xu, F.L. Combining species sensitivity distribution (SSD) model and thermodynamic index (exergy) for system-level ecological risk assessment of contaminates in aquatic ecosystems. Environ. Int. 2019, 133. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The number of publications that consider indirect effects published in nine representative environmental toxicology journals from 1988 (the first year the search term returned a publication) to 2019. The journals are Environmental Toxicology and Chemistry, Ecotoxicology, Marine Pollution Bulletin, Archives of Environmental Contamination and Toxicology, Environmental Pollution, Bulletin of Environmental Contamination and Toxicology, Environmental Toxicology, Ecotoxicology and Environmental Safety, and Aquatic Toxicology. A Web of Science advanced search was preformed using search terms “indirect effects” AND the various “source titles” in September 2020. Publications that use the term “indirect effects” but were unrelated to effects at the population, community, or ecosystem level were excluded from the count.
Figure 1.
The number of publications that consider indirect effects published in nine representative environmental toxicology journals from 1988 (the first year the search term returned a publication) to 2019. The journals are Environmental Toxicology and Chemistry, Ecotoxicology, Marine Pollution Bulletin, Archives of Environmental Contamination and Toxicology, Environmental Pollution, Bulletin of Environmental Contamination and Toxicology, Environmental Toxicology, Ecotoxicology and Environmental Safety, and Aquatic Toxicology. A Web of Science advanced search was preformed using search terms “indirect effects” AND the various “source titles” in September 2020. Publications that use the term “indirect effects” but were unrelated to effects at the population, community, or ecosystem level were excluded from the count.
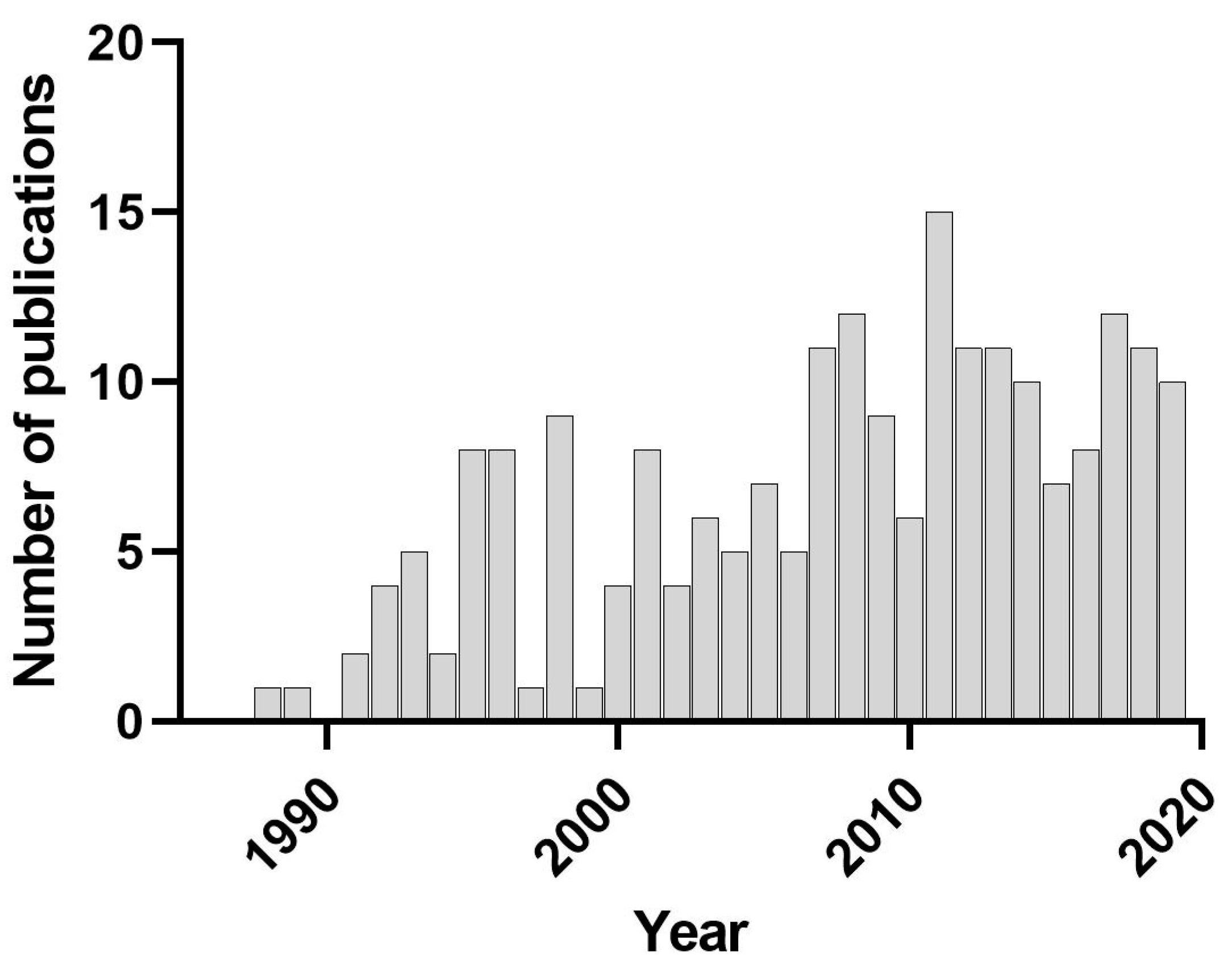
Figure 2.
Hypothetical indirect effects of separate exposures of a pesticide and an herbicide in a low-diversity ecosystem. In each case, exposure will lead to strong and relatively predictable indirect effects with straightforward causative mechanisms. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes.
Figure 2.
Hypothetical indirect effects of separate exposures of a pesticide and an herbicide in a low-diversity ecosystem. In each case, exposure will lead to strong and relatively predictable indirect effects with straightforward causative mechanisms. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes.
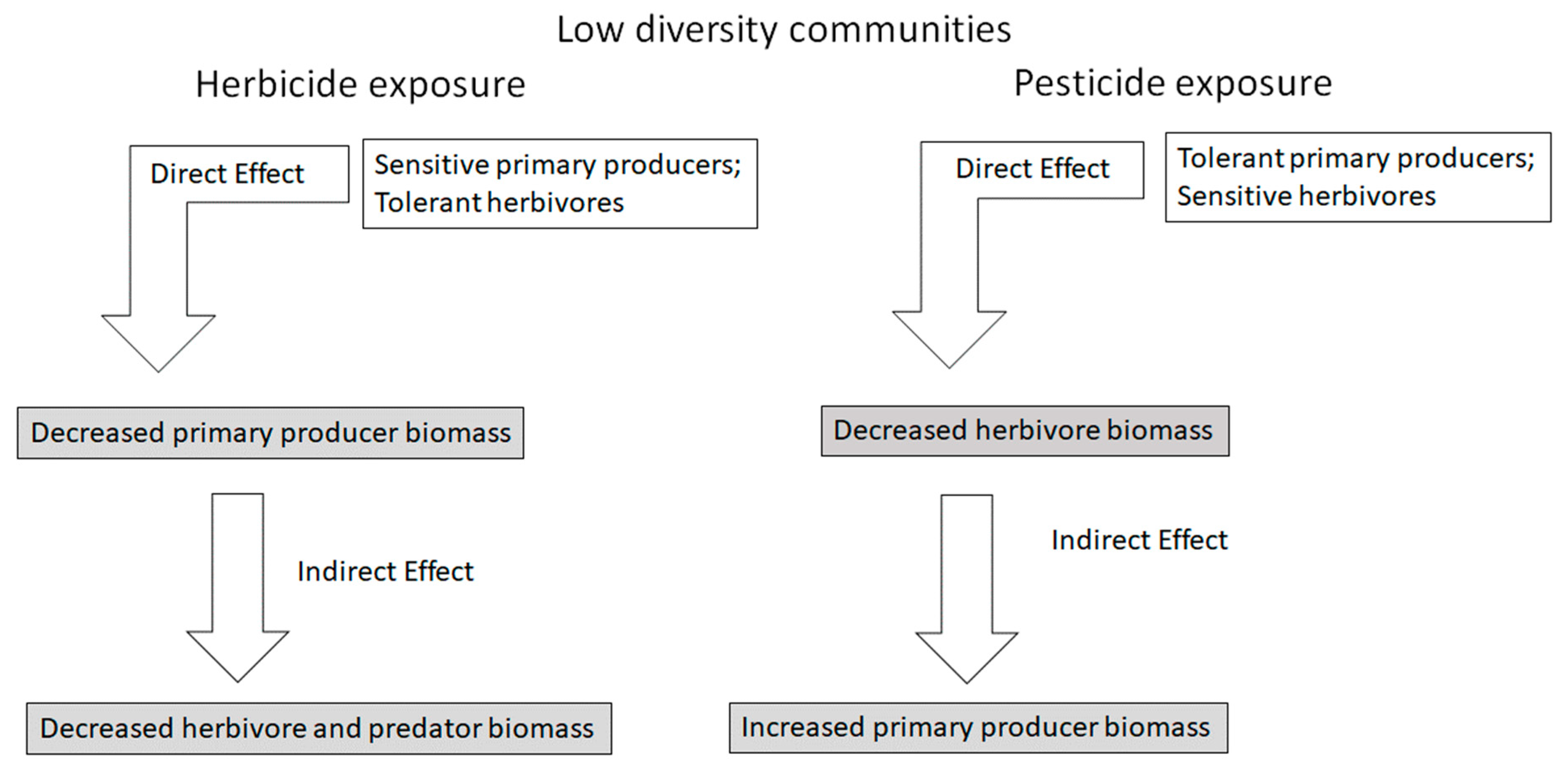
Figure 3.
One scenario for indirect effects in a diverse community with relatively sensitive and relatively tolerant primary producer species exposed to an herbicide. In this case, indirect effects may be mild but will be difficult to predict, and they will vary depending on species composition and with changes in environmental conditions. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes.
Figure 3.
One scenario for indirect effects in a diverse community with relatively sensitive and relatively tolerant primary producer species exposed to an herbicide. In this case, indirect effects may be mild but will be difficult to predict, and they will vary depending on species composition and with changes in environmental conditions. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes.
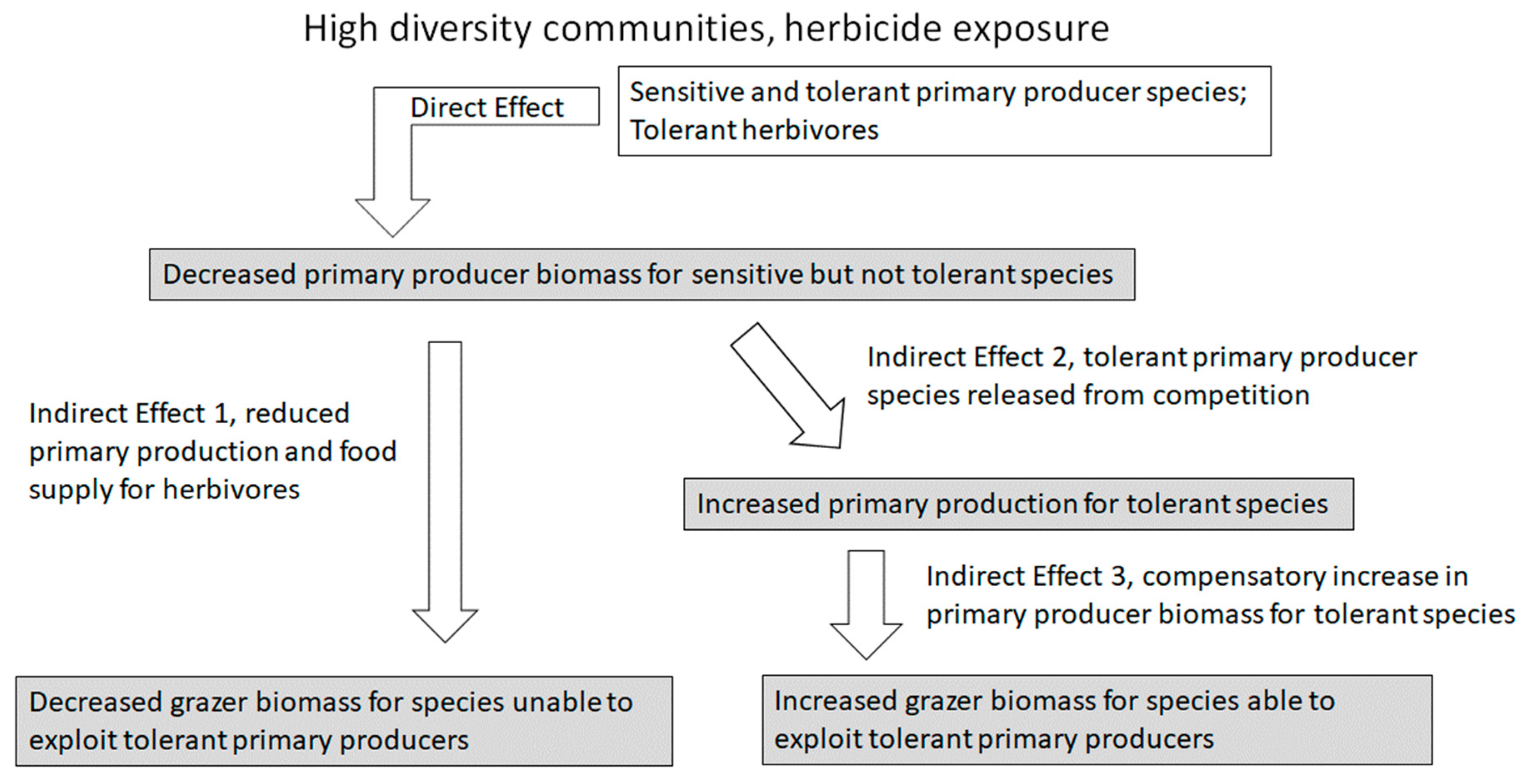
Figure 4.
One scenario for indirect effects in a diverse community with relatively sensitive and relatively tolerant herbivores species exposed to a pesticide. In this example, indirect effects may be mild but will be difficult to predict, and they will vary depending on species composition and with changes in environmental conditions. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes. This figure was based on a figure published in [1], page number 217, Copyright Elsevier.
Figure 4.
One scenario for indirect effects in a diverse community with relatively sensitive and relatively tolerant herbivores species exposed to a pesticide. In this example, indirect effects may be mild but will be difficult to predict, and they will vary depending on species composition and with changes in environmental conditions. Ecosystem state changes caused by direct and indirect effects are described in shaded boxes. This figure was based on a figure published in [1], page number 217, Copyright Elsevier.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fleeger, J.W. How Do Indirect Effects of Contaminants Inform Ecotoxicology? A Review. Processes 2020, 8, 1659. https://doi.org/10.3390/pr8121659
AMA Style
Fleeger JW. How Do Indirect Effects of Contaminants Inform Ecotoxicology? A Review. Processes. 2020; 8(12):1659. https://doi.org/10.3390/pr8121659
Chicago/Turabian StyleFleeger, John W. 2020. "How Do Indirect Effects of Contaminants Inform Ecotoxicology? A Review" Processes 8, no. 12: 1659. https://doi.org/10.3390/pr8121659
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.