
Effects of Recirculating Aquaculture System Wastewater on Anammox Performance and Community Structure

 ,
,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. RAS Wastewater Collection and Characteristics
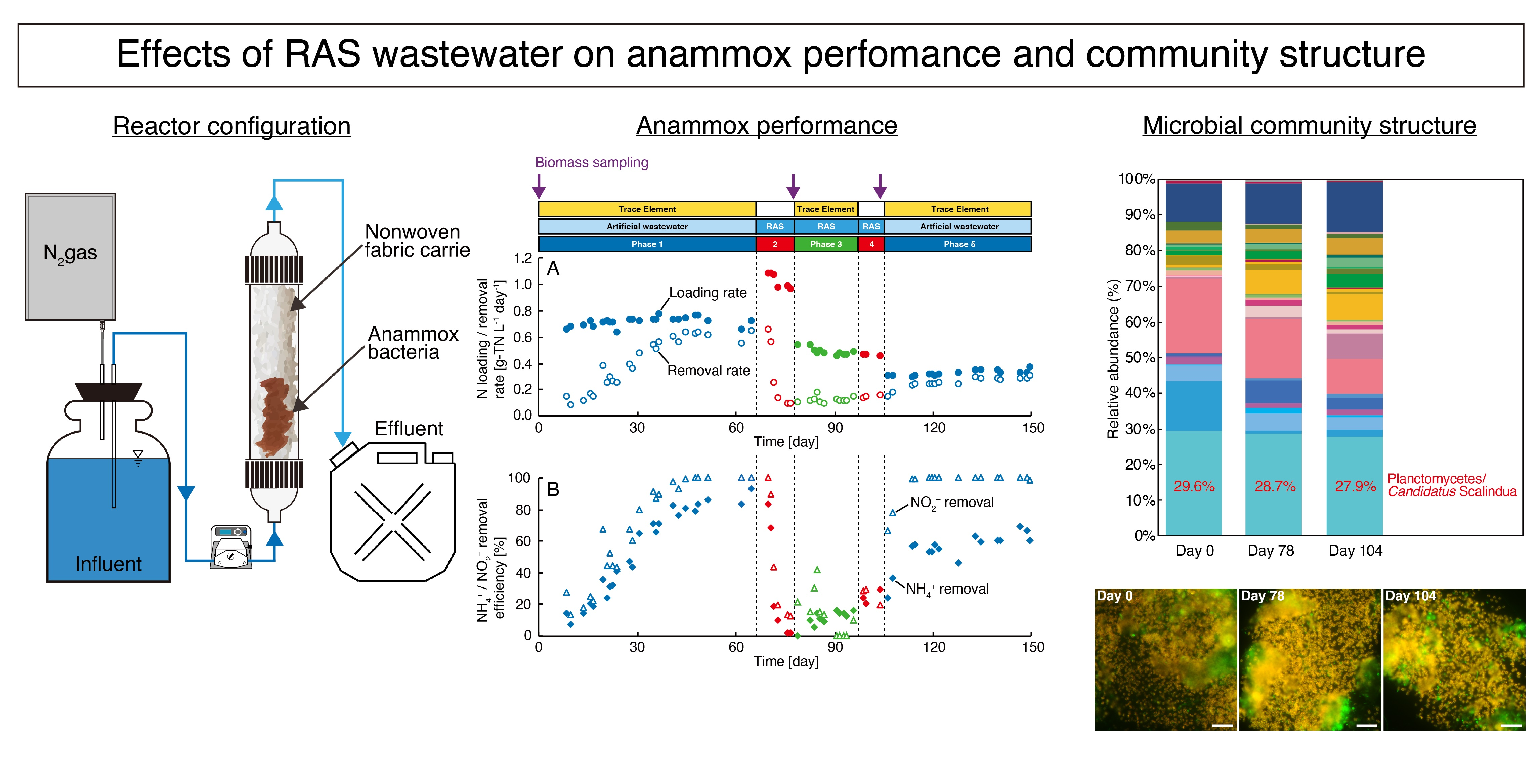
2.2. Reactor Operation
2.3. Analytical Methods
2.4. Microbial Community Analysis
2.5. Fluorescence In Situ Hybridization (FISH)
3. Results
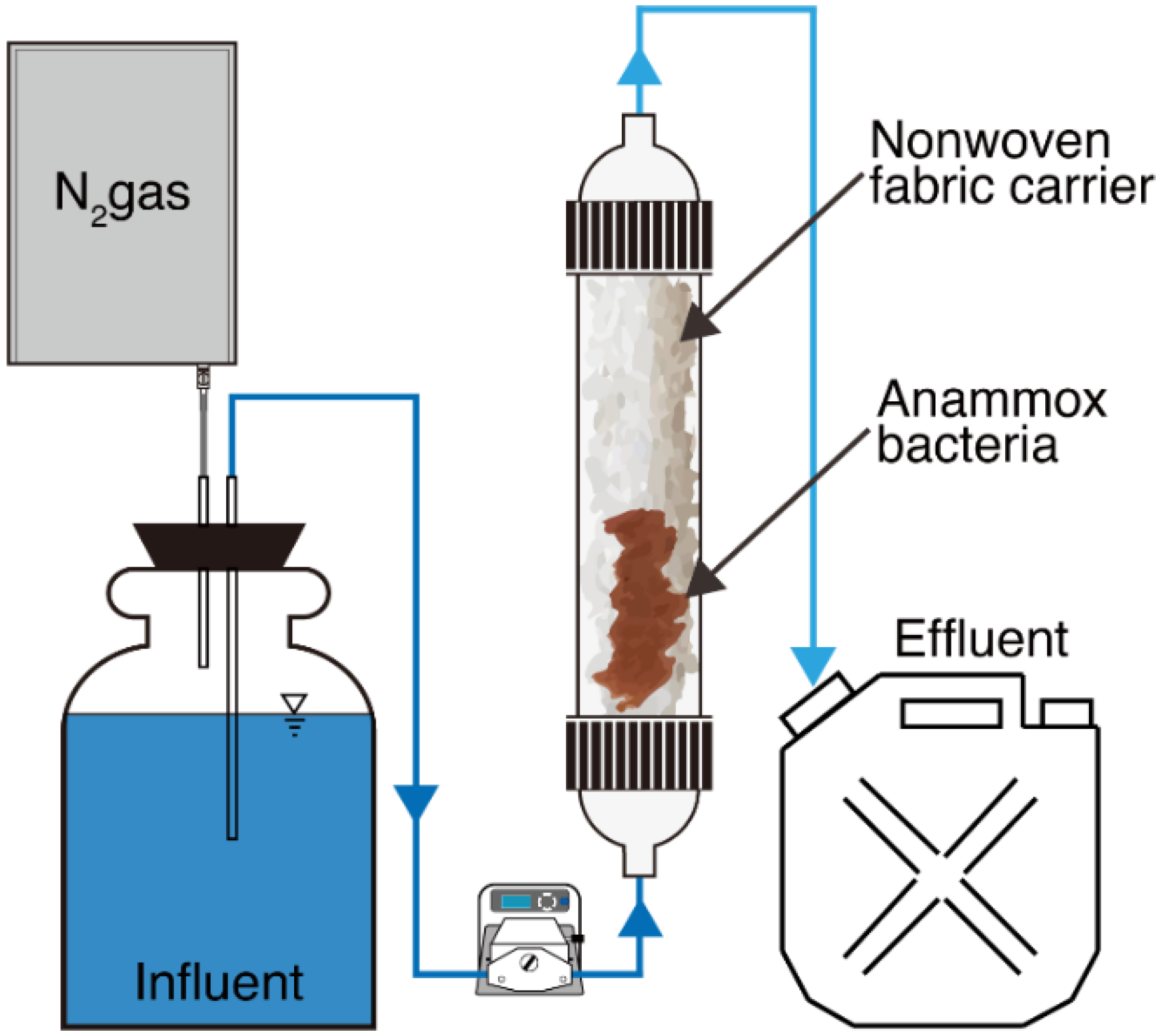
3.1. Reactor Performance
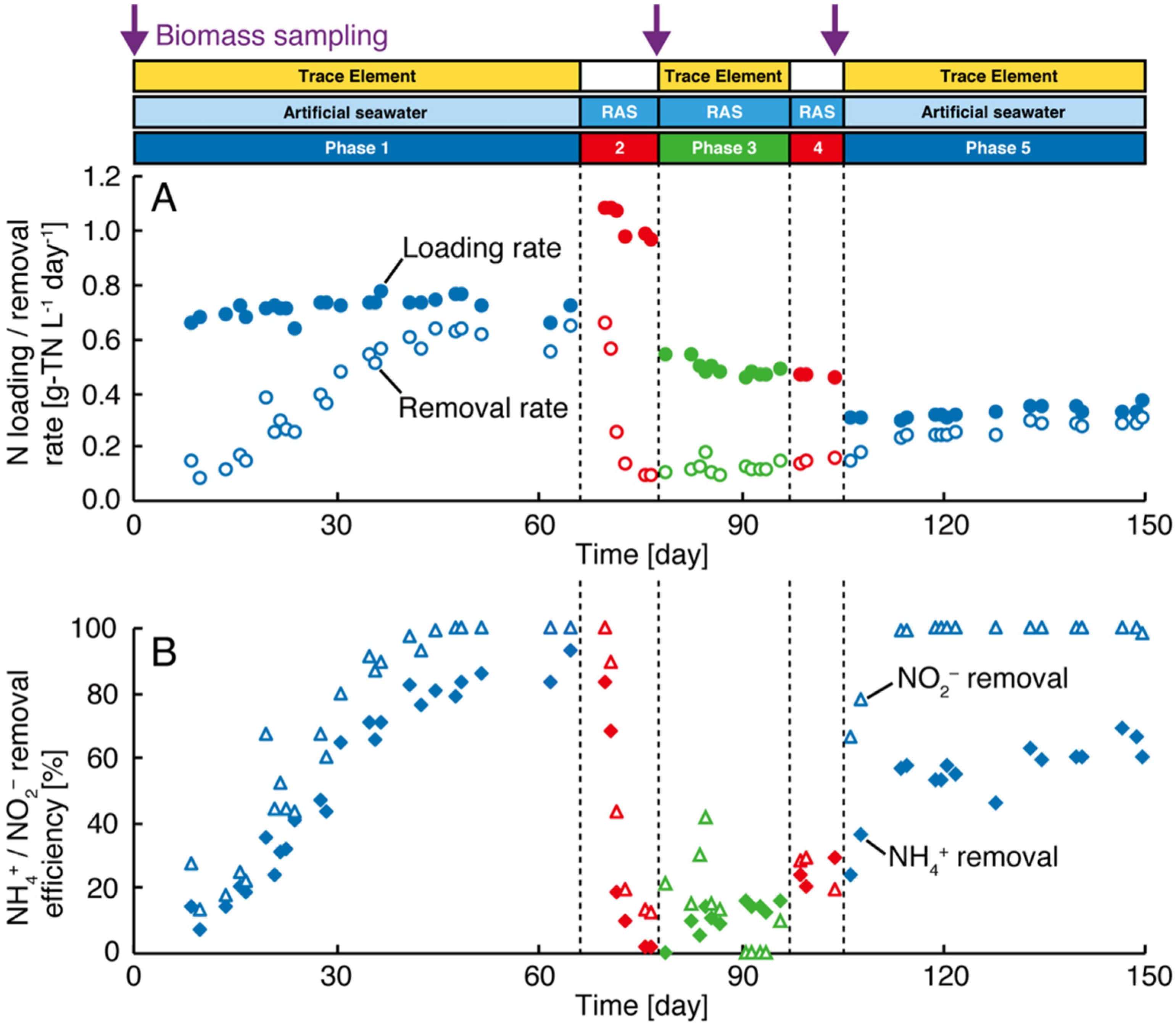
3.2. Microbial Community Analysis and FISH
4. Discussion
4.1. Reactor Performance
4.2. Microbial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects, Online edition; Rev. 1; United Nations, Department of Economic and Social Affairs: New York, NY, USA, 2019. [Google Scholar]
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.-I.; Williams, M. Feeding 9 billion by 2050–Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheries and Aquaculture Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Pahri, S.D.R.; Mohamed, A.F.; Samat, A. LCA for open systems: A review of the influence of natural and anthropogenic factors on aquaculture systems. Int. J. Life Cycle Assess. 2015, 20, 1324–1337. [Google Scholar] [CrossRef]
- Martins, C.; Eding, E.H.; Verdegem, M.C.; Heinsbroek, L.T.; Schneider, O.; Blancheton, J.-P.; d’Orbcastel, E.R.; Verreth, J. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability. Aquacult. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Kolarevic, J.; Baeverfjord, G.; Takle, H.; Ytteborg, E.; Reiten, B.K.M.; Nergård, S.; Terjesen, B.F. Performance and welfare of Atlantic salmon smolt reared in recirculating or flow through aquaculture systems. Aquaculture 2014, 432, 15–25. [Google Scholar] [CrossRef]
- Mota, V.C.; Nilsen, T.O.; Gerwins, J.; Gallo, M.; Ytteborg, E.; Baeverfjord, G.; Kolarevic, J.; Summerfelt, S.T.; Terjesen, B.F. The effects of carbon dioxide on growth performance, welfare, and health of Atlantic salmon post-smolt (Salmo salar) in recirculating aquaculture systems. Aquaculture 2019, 498, 578–586. [Google Scholar] [CrossRef]
- Roques, J.A.C. Aspects of Fish Welfare in Aquaculture Practices. Ph.D. Thesis, Radboud University Nijmegen, Nijmegen, The Netherlands, 2013. [Google Scholar]
- Van Rijn, J.; Ebeling, J. Recirculating Aquaculture; Ventures: Ithaca, NY, USA, 2010; pp. 387–424. [Google Scholar]
- Chen, S. Recirculating Systems Effluents and Treatments; Aquaculture and the Environment in the United States, World Aquaculture Society: Baton Rouge, LA, USA, 2002; pp. 119–140. [Google Scholar]
- Camargo, J.A.; Alonso, A.; Salamanca, A. Nitrate toxicity to aquatic animals: A review with new data for freshwater invertebrates. Chemosphere 2005, 58, 1255–1267. [Google Scholar] [CrossRef]
- Sindilariu, P.-D.; Brinker, A.; Reiter, R. Waste and particle management in a commercial, partially recirculating trout farm. Aquacult. Eng. 2009, 41, 127–135. [Google Scholar] [CrossRef]
- Stavrakidis-Zachou, O.; Ernst, A.; Steinbach, C.; Wagner, K.; Waller, U. Development of denitrification in semi-automated moving bed biofilm reactors operated in a marine recirculating aquaculture system. Aquac. Int. 2019, 27, 1485–1501. [Google Scholar] [CrossRef]
- Hu, Z.; Lee, J.W.; Chandran, K.; Kim, S.; Khanal, S.K. Nitrous oxide (N2O) emission from aquaculture: A review. Environ. Sci. Technol. 2012, 46, 6470–6480. [Google Scholar] [CrossRef] [PubMed]
- Kuenen, J.G. Anammox bacteria: From discovery to application. Nat. Rev. Microbiol. 2008, 6, 320–326. [Google Scholar] [CrossRef]
- Schmid, M.; Walsh, K.; Webb, R.; Rijpstra, W.I.; van de Pas-Schoonen, K.; Verbruggen, M.J.; Hill, T.; Moffett, B.; Fuerst, J.; Schouten, S. Candidatus “Scalindua brodae”, sp. nov., Candidatus “Scalindua wagneri”, sp. nov., two new species of anaerobic ammonium oxidizing bacteria. Syst. Appl. Microbiol. 2003, 26, 529–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojiri, A.; Ohashi, A.; Ozaki, N.; Aoi, Y.; Kindaichi, T. Integrated anammox-biochar in synthetic wastewater treatment: Performance and optimization by artificial neural network. J. Clean. Prod. 2020, 243, 118638. [Google Scholar] [CrossRef]
- Preena, P.G.; Rejish Kumar, V.J.; Singh, I.S.B. Nitrification and denitrification in recirculating aquaculture systems: The processes and players. Rev. Aqua. 2021. [Google Scholar] [CrossRef]
- Jetten, M.S.; Niftrik, L.v.; Strous, M.; Kartal, B.; Keltjens, J.T.; Op den Camp, H.J. Biochemistry and molecular biology of anammox bacteria. Crit. Rev. Biochem. Mol. Biol. 2009, 44, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Lahav, O.; Massada, I.B.; Yackoubov, D.; Zelikson, R.; Mozes, N.; Tal, Y.; Tarre, S. Quantification of anammox activity in a denitrification reactor for a recirculating aquaculture system. Aquaculture 2009, 288, 76–82. [Google Scholar] [CrossRef]
- Tal, Y.; Watts, J.E.; Schreier, H.J. Anaerobic ammonium-oxidizing (anammox) bacteria and associated activity in fixed-film biofilters of a marine recirculating aquaculture system. Appl. Environ. Microbiol. 2006, 72, 2896–2904. [Google Scholar] [CrossRef] [Green Version]
- van Kessel, M.A.; Harhangi, H.R.; van de Pas-Schoonen, K.; van de Vossenberg, J.; Flik, G.; Jetten, M.S.; Klaren, P.H.; den Camp, H.J.O. Biodiversity of N-cycle bacteria in nitrogen removing moving bed biofilters for freshwater recirculating aquaculture systems. Aquaculture 2010, 306, 177–184. [Google Scholar] [CrossRef]
- Schreier, H.J.; Mirzoyan, N.; Saito, K. Microbial diversity of biological filters in recirculating aquaculture systems. Curr. Opin. Biotechnol. 2010, 21, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Tal, Y.; Schreier, H.J.; Sowers, K.R.; Stubblefield, J.D.; Place, A.R.; Zohar, Y. Environmentally sustainable land-based marine aquaculture. Aquaculture 2009, 286, 28–35. [Google Scholar] [CrossRef]
- Awata, T.; Oshiki, M.; Kindaichi, T.; Ozaki, N.; Ohashi, A.; Okabe, S. Physiological characterization of an anaerobic ammonium-oxidizing bacterium belonging to the “Candidatus Scalindua” group. Appl. Environ. Microbiol. 2013, 79, 4145–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kindaichi, T.; Awata, T.; Tanabe, K.; Ozaki, N.; Ohashi, A. Enrichment of marine anammox bacteria in Hiroshima Bay sediments. Water Sci. Technol. 2011, 63, 964–969. [Google Scholar] [CrossRef] [Green Version]
- Oshiki, M.; Awata, T.; Kindaichi, T.; Satoh, H.; Okabe, S. Cultivation of planktonic anaerobic ammonium oxidation (anammox) bacteria using membrane bioreactor. Microbes Environ. 2013, 28, 436–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awata, T.; Kindaichi, T.; Ozaki, N.; Ohashi, A. Biomass yield efficiency of the marine anammox bacterium, “Candidatus Scalindua sp.,” is affected by salinity. Microbes Environ. 2015, 30, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brijs, J.; Hjelmstedt, P.; Berg, C.; Johansen, I.; Sundh, H.; Roques, J.; Ekström, A.; Sandblom, E.; Sundell, K.; Olsson, C. Prevalence and severity of cardiac abnormalities and arteriosclerosis in farmed rainbow trout (Oncorhynchus mykiss). Aquaculture 2020, 526, 735417. [Google Scholar] [CrossRef]
- Van de Graaf, A.A.; de Bruijn, P.; Robertson, L.A.; Jetten, M.S.; Kuenen, J.G. Autotrophic growth of anaerobic ammonium-oxidizing micro-organisms in a fluidized bed reactor. Microbiology 1996, 142, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Kindaichi, T.; Awata, T.; Suzuki, Y.; Tanabe, K.; Hatamoto, M.; Ozaki, N.; Ohashi, A. Enrichment Using an Up-flow Column Reactor and Community Structure of Marine Anammox Bacteria from Coastal Sediment. Microbes Environ. 2011, 26, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojiri, A.; Nishimoto, K.; Awata, T.; Aoi, Y.; Ozaki, N.; Ohashi, A.; Kindaichi, T. Effects of salts on the activity and growth of “Candidatus Scalindua sp.”, a marine Anammox bacterium. Microbes Environ. 2018, 33, 336–339. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Shoiful, A.; Kambara, H.; Cao, L.T.T.; Matsushita, S.; Kindaichi, T.; Aoi, Y.; Ozaki, N.; Ohashi, A. Mn (II) oxidation and manganese-oxide reduction on the decolorization of an azo dye. Int. Biodeter. Biodeg. 2020, 146, 104820. [Google Scholar] [CrossRef]
- Awata, T.; Goto, Y.; Kuratsuka, H.; Aoi, Y.; Ozaki, N.; Ohashi, A.; Kindaichi, T. Reactor performance and microbial community structure of single-stage partial nitritation anammox membrane bioreactors inoculated with Brocadia and Scalindua enrichment cultures. Biochem. Eng. J. 2021, 170, 107991. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. QIIME 2: Reproducible, Interactive, Scalable, and Extensible Microbiome Data Science. PeerJ 2018, 2167–9843. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.-H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Schmid, M.C.; Maas, B.; Dapena, A.; van de Pas-Schoonen, K.; van de Vossenberg, J.; Kartal, B.; van Niftrik, L.; Schmidt, I.; Cirpus, I.; Kuenen, J.G. Biomarkers for in situ detection of anaerobic ammonium-oxidizing (anammox) bacteria. Appl. Environ. Microbiol. 2005, 71, 1677–1684. [Google Scholar] [CrossRef] [Green Version]
- Tsushima, I.; Ogasawara, Y.; Kindaichi, T.; Satoh, H.; Okabe, S. Development of high-rate anaerobic ammonium-oxidizing (anammox) biofilm reactors. Water Res. 2007, 41, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Takaki, K.; Koyama, T.; Furukawa, K. Long-term stability of partial nitritation of swine wastewater digester liquor and its subsequent treatment by Anammox. Bioresour. Technol. 2008, 99, 6419–6425. [Google Scholar] [CrossRef]
- Okabe, S.; Oshiki, M.; Takahashi, Y.; Satoh, H. Development of long-term stable partial nitrification and subsequent anammox process. Bioresour. Technol. 2011, 102, 6801–6807. [Google Scholar] [CrossRef]
- Kimura, Y.; Isaka, K. Evaluation of inhibitory effects of heavy metals on anaerobic ammonium oxidation (anammox) by continuous feeding tests. Appl. Microbiol. Biot. 2014, 98, 6965–6972. [Google Scholar] [CrossRef]
- Li, H.; Yao, H.; Zhang, D.; Zuo, L.; Ren, J.; Ma, J.; Pei, J.; Xu, Y.; Yang, C. Short-and long-term effects of manganese, zinc and copper ions on nitrogen removal in nitritation-anammox process. Chemosphere 2018, 193, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aklujkar, M.; Coppi, M.V.; Leang, C.; Kim, B.C.; Chavan, M.; Perpetua, L.; Giloteaux, L.; Liu, A.; Holmes, D. Proteins involved in electron transfer to Fe (III) and Mn (IV) oxides by Geobacter sulfurreducens and Geobacter uraniireducens. Microbiology 2013, 159, 515–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Gao, D.; Peng, S.; Tao, Y. Effects of ferrous and manganese ions on anammox process in sequencing batch biofilm reactors. J. Environ. Sci. 2014, 26, 1034–1039. [Google Scholar] [CrossRef]
- Lotti, T.; Cordola, M.; Kleerebezem, R.; Caffaz, S.; Lubello, C.; Van Loosdrecht, M. Inhibition effect of swine wastewater heavy metals and antibiotics on anammox activity. Water Sci. Technol. 2012, 66, 1519–1526. [Google Scholar] [CrossRef]
- Yang, G.-F.; Ni, W.-M.; Wu, K.; Wang, H.; Yang, B.-E.; Jia, X.-Y.; Jin, R.-C. The effect of Cu (II) stress on the activity, performance and recovery on the anaerobic ammonium-oxidizing (Anammox) process. Chem. Eng. J. 2013, 226, 39–45. [Google Scholar] [CrossRef]
- Li, J.; Feng, L.; Qiang, Z.; Dong, H.; Wang, D. Enhanced performance and kinetics of marine anammox bacteria (MAB) treating nitrogen-rich saline wastewater with Mn (II) and Ni (II) addition. Bioresour. Technol. 2018, 249, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Molinuevo, B.; García, M.C.; Karakashev, D.; Angelidaki, I. Anammox for ammonia removal from pig manure effluents: Effect of organic matter content on process performance. Bioresour. Technol. 2009, 100, 2171–2175. [Google Scholar] [CrossRef]
- Jin, R.-C.; Yang, G.-F.; Yu, J.-J.; Zheng, P. The inhibition of the Anammox process: A review. Chem. Eng. J. 2012, 197, 67–79. [Google Scholar] [CrossRef]
- Chamchoi, N.; Nitisoravut, S.; Schmidt, J.E. Inactivation of ANAMMOX communities under concurrent operation of anaerobic ammonium oxidation (ANAMMOX) and denitrification. Bioresour. Technol. 2008, 99, 3331–3336. [Google Scholar] [CrossRef] [PubMed]
- Kindaichi, T.; Awata, T.; Mugimoto, Y.; Rathnayake, R.M.; Kasahara, S.; Satoh, H. Effects of organic matter in livestock manure digester liquid on microbial community structure and in situ activity of anammox granules. Chemosphere 2016, 159, 300–307. [Google Scholar] [CrossRef]
- Mizuno, C.M.; Rodriguez-Valera, F.; Ghai, R. Genomes of planktonic acidimicrobiales: Widening horizons for marine actinobacteria by metagenomics. MBio 2015, 6, e02083-14. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Oomaru, K.; Nakamura, S.; Sato, T.; Nakatani, Y. Reducing the switching current with a Gilbert damping constant in nanomagnets with perpendicular anisotropy. Appl. Phys. Lett. 2015, 106, 042402. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.K. Encyclopedia of Food Microbiology; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Nedashkovskaya, O.I.; Kim, S.B.; Han, S.K.; Rhee, M.S.; Lysenko, A.M.; Falsen, E.; Frolova, G.M.; Mikhailov, V.V.; Bae, K.S. Ulvibacter litoralis gen. nov., sp. nov., a novel member of the family Flavobacteriaceae isolated from the green alga Ulva fenestrata. Int. J. Syst. Evol. Microbiol. 2004, 54, 119–123. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J. Planctomycetes and macroalgae, a striking association. Front. Microbiol. 2014, 5, 267. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Salinity (‰) | NH4+ (mg-N L−1) | NO2− (mg-N L−1) | NO3− (mg-N L−1) | pH | TSS (mg L−1) | TOC (mg-C L−1) |
|---|---|---|---|---|---|---|---|
| Value | 14.5 | 0.25 | 0.017 | 29.22 | 7.5 | 70.5 | 6.8 |
| Phase | Period | AS/RAS 1 | TEs 2 | HRT | NLR | pH * | Salinity (‰) * | ||
|---|---|---|---|---|---|---|---|---|---|
| (d) | (h) 3,* | (g-TN L−1 day−1) 4,* | Influent | Effluent | Influent | Effluent | |||
| 1 | 0–65 | AS | + | 2.3 | 0.72 ± 0.03 | 7.62 ± 0.16 | 7.82 ± 0.14 | 25.5 ± 2.9 | 25.6 ± 3.1 |
| 2 | 65–77 | RAS | − | 2.4 | 1.02 ± 0.06 | 7.80 ± 0.24 | 7.95 ± 0.20 | 26.6 ± 0.1 | 26.2 ± 1.0 |
| 3 | 77–96 | RAS | + | 4.8 | 0.48 ± 0.03 | 7.57 ± 0.16 | 7.78 ± 0.09 | 26.1 ± 0.6 | 26.2 ± 0.8 |
| 4 | 96–104 | RAS | − | 5.1 | 0.45 ± 0.01 | 7.47 ± 0.06 | 7.67 ± 0.15 | 26.2 ± 0.7 | 26.3 ± 0.4 |
| 5 | 104–150 | AS | + | 5.1 | 0.33 ± 0.02 | 7.66 ± 0.19 | 7.91 ± 0.12 | 27.2 ± 0.6 | 27.2 ± 0.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roques, J.A.C.; Micolucci, F.; Hosokawa, S.; Sundell, K.; Kindaichi, T. Effects of Recirculating Aquaculture System Wastewater on Anammox Performance and Community Structure. Processes 2021, 9, 1183. https://doi.org/10.3390/pr9071183
Roques JAC, Micolucci F, Hosokawa S, Sundell K, Kindaichi T. Effects of Recirculating Aquaculture System Wastewater on Anammox Performance and Community Structure. Processes. 2021; 9(7):1183. https://doi.org/10.3390/pr9071183
Chicago/Turabian StyleRoques, Jonathan A. C., Federico Micolucci, Suguru Hosokawa, Kristina Sundell, and Tomonori Kindaichi. 2021. "Effects of Recirculating Aquaculture System Wastewater on Anammox Performance and Community Structure" Processes 9, no. 7: 1183. https://doi.org/10.3390/pr9071183