
Expression, Purification and Characterization of a Novel Rusticyanin from the Psychrotolerant Acidithiobacillus ferrivorans

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression, and Purification of the Recombinant Type-C Rusticyanin from A. ferrivorans
2.2. Assembly of the Holo-Rusticyanin and Preparations for the Oxidized and Reduced States
2.3. Characterization Methods
2.4. Kinetic Measurement between the Oxidized Rusticyanin and Ferrous ion
2.5. Bioinformatics Analysis
3. Results and Discussion
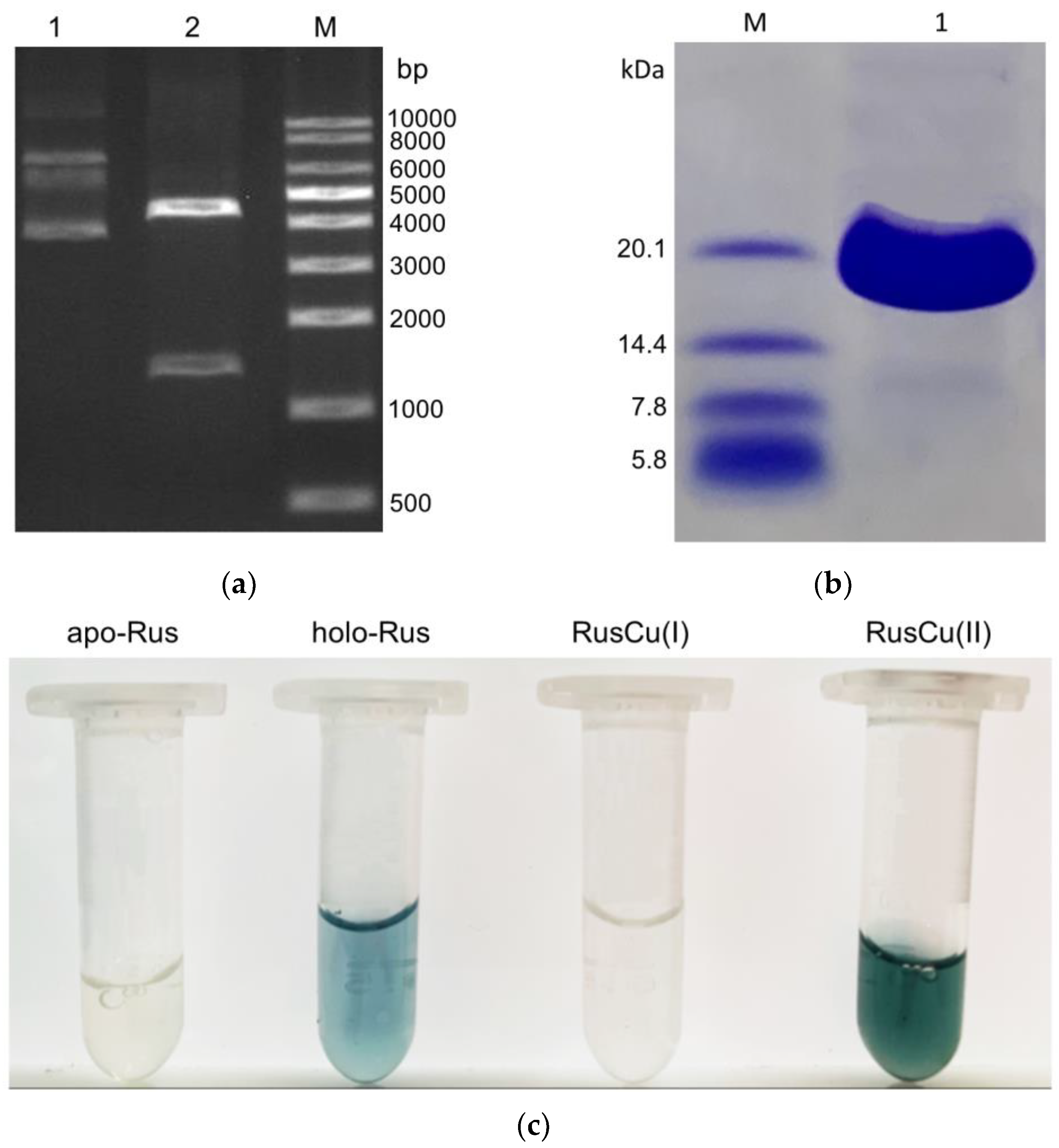
3.1. Gene Cloning, Protein Purification, and Assembly of Type-C Rusticyanin from A. ferrivorans
3.2. MALDI-TOF-MS and DSC of the Type-C Rusticyanin
3.3. UV-Scanning and EPR of the Type-C Rusticyanin
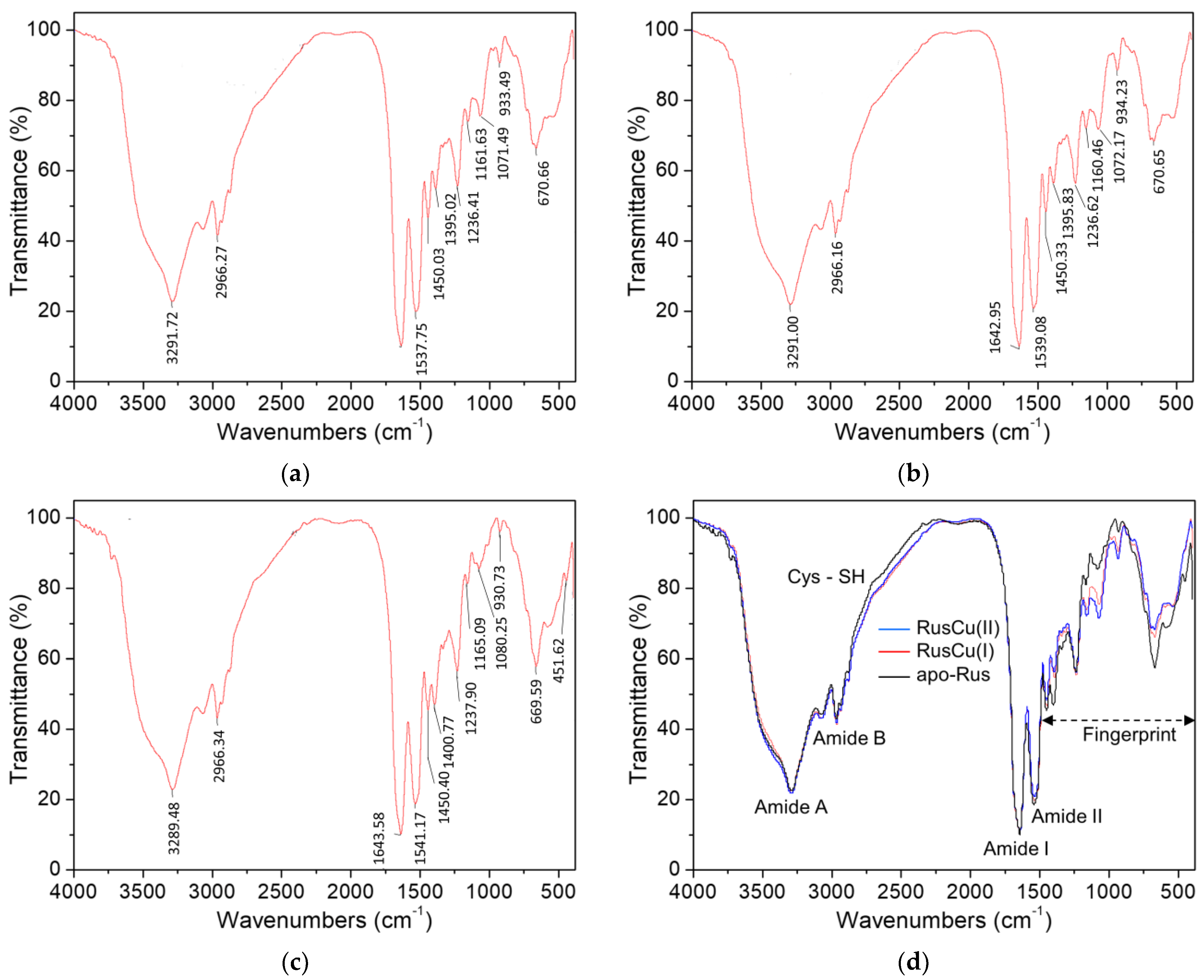
3.4. FTIR for Apo-, Oxidized and Reduced States of the Type-C Rusticyanin
3.5. Electron Transfer from Ferrous Ion to the Type-C
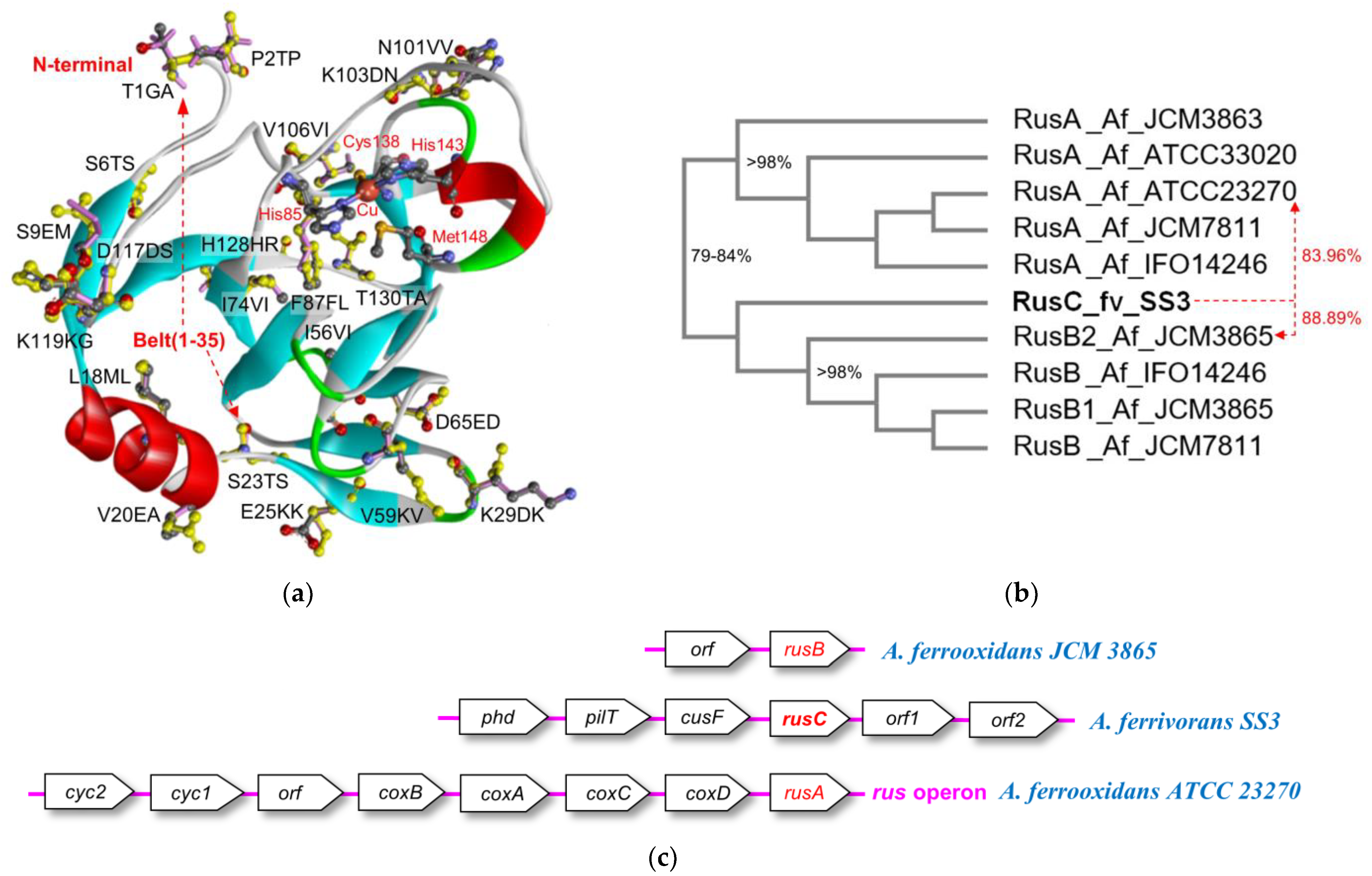
3.6. Bioinformatics Comparison of the Rusticyanins
3.7. Classification of the Type-C Rusticyanin from A. ferrivorans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Botuyan, M.V.; Toy-Palmer, A.; Chung, J.; Blake, R.C., 2nd; Beroza, P.; Case, D.A.; Dyson, H.J. NMR solution structure of Cu(I) rusticyanin from Thiobacillus ferrooxidans: Structural basis for the extreme acid stability and redox potential. J. Mol. Biol. 1996, 263, 752–767. [Google Scholar] [CrossRef] [PubMed]
- Valdes, J.; Pedroso, I.; Quatrini, R.; Dodson, R.J.; Tettelin, H.; Blake, R., II; Eisen, J.A.; Holmes, D.S. Acidithiobacillus ferrooxidans metabolism: From genome sequence to industrial applications. BMC Genom. 2008, 9, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheaton, G.; Counts, J.; Mukherjee, A.; Kruh, J.; Kelly, R. The Confluence of Heavy Metal Biooxidation and Heavy Metal Resistance: Implications for Bioleaching by Extreme Thermoacidophiles. Minerals 2015, 5, 397–451. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.C.; Boxer, D.H. The purification and some properties of rusticyanin, a blue copper protein involved in iron(II) oxidation from Thiobacillus ferrooxidans. Biochem. J. 1978, 174, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Ronk, M.; Shively, J.E.; Shute, E.A.; Blake, R.C., 2nd. Amino acid sequence of the blue copper protein rusticyanin from Thiobacillus ferrooxidans. Biochemistry 1991, 30, 9435–9442. [Google Scholar] [CrossRef]
- Yarzabal, A.; Appia-Ayme, C.; Ratouchniak, J.; Bonnefoy, V. Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes c, a cytochrome oxidase and rusticyanin. Microbiology 2004, 150, 2113–2123. [Google Scholar] [CrossRef] [Green Version]
- Nunzi, F.; Guerlesquin, F.; Shepard, W.; Guigliarelli, B.; Bruschi, M. Active site geometry in the high oxido-reduction potential rusticyanin from Thiobacillus ferrooxidans. Biochem. Biophys. Res. Commun. 1994, 203, 1655–1662. [Google Scholar] [CrossRef]
- Grossmann, J.G. The N-terminal extension of rusticyanin is not responsible for its acid stability. Biochemistry 2002, 41, 3619. [Google Scholar] [CrossRef]
- Barrett, M.L. Atomic resolution crystal structures, EXAFS, and quantum chemical studies of rusticyanin and its two mutants provide insight into its unusual properties. Biochemistry 2006, 45, 2939. [Google Scholar] [CrossRef]
- Yamada, T.; Hiraoka, Y.; Das Gupta, T.K.; Chakrabarty, A.M. Rusticyanin, a bacterial electron transfer protein, causes G(1) arrest in J774 and apoptosis in human cancer cells. Cell Cycle 2004, 3, 1182–1187. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Gallardo, I.; Diaz-Moreno, I.; Diaz-Quintana, A.; Donaire, A.; Velazquez-Campoy, A.; Curd, R.D.; Rangachari, K.; Birdsall, B.; Ramos, A.; Holder, A.A.; et al. Antimalarial Activity of Cupredoxins the Interaction of Plasmodium Merozoite Surface Protein 1(19) (MSP1(19)) and Rusticyanin. J. Biol. Chem. 2013, 288, 20896–20907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Arribas, E.; Falk, M.; Aleksejeva, O.; Bushnev, S.; Sebastian, P.; Feliu, J.M.; Shleev, S. A conventional symmetric biosupercapacitor based on rusticyanin modified gold electrodes. J. Electroanal. Chem. 2018, 816, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K. Respiratory isozyme, two types of rusticyanin of Acidithiobacillus ferrooxidans. Biosci. Biotechnol. Biochem. 2003, 67, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ida, C. Kinetic rate constant for electron transfer between ferrous ions and novel Rusticyanin isoform in Acidithiobacillus ferrooxidans. J. Biosci. Bioeng. 2003, 95, 537. [Google Scholar] [CrossRef]
- Hedrich, S.; Schloemann, M.; Johnson, D.B. The iron-oxidizing proteobacteria. Microbiology 2011, 157, 1551–1564. [Google Scholar] [CrossRef] [Green Version]
- Ccorahua-Santo, R.; Eca, A.; Abanto, M.; Guerra, G.; Ramirez, P. Physiological and comparative genomic analysis of Acidithiobacillus ferrivorans PQ33 provides psychrotolerant fitness evidence for oxidation at low temperature. Res. Microbiol. 2017, 168, 482–492. [Google Scholar] [CrossRef]
- Liljeqvist, M.; Valdes, J.; Holmes, D.S.; Dopson, M. Draft Genome of the Psychrotolerant Acidophile Acidithiobacillus ferrivorans SS3. J. Bacteriol. 2011, 193, 4304–4305. [Google Scholar] [CrossRef] [Green Version]
- Jafarpour, R.; Fatemi, F.; Eidi, A.; Mehrnejad, F. Effect of the Met148Leu mutation on the structure and dynamics of the rusticyanin protein from Acidithiobacillus sp. FJ2. J. Biomol. Struct. Dyn. 2021, 39, 4122–4132. [Google Scholar] [CrossRef]
- Panyushkina, A.E.; Babenko, V.V.; Nikitina, A.S.; Selezneva, O.V.; Tsaplina, I.A.; Letarova, M.A.; Kostryukova, E.S.; Letarov, A.V. Sulfobacillus thermotolerans: New insights into resistance and metabolic capacities of acidophilic chemolithotrophs. Sci. Rep. 2019, 9, 15069. [Google Scholar] [CrossRef] [Green Version]
- Sousa, P.; Camacho, I.; Câmara, J.S.; Perestrelo, R. Urinary Proteomic/Peptidomic Biosignature of Breast Cancer Patients Using 1D SDS-PAGE Combined with Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry. Separations 2023, 10, 291. [Google Scholar] [CrossRef]
- Zeng, J.; Geng, M.; Liu, Y.; Xia, L.; Liu, J.; Qiu, G. The sulfhydryl group of Cys 138 of rusticyanin from Acidithiobacillus ferrooxidans is crucial for copper binding. Biochim. Biophys. Acta-Proteins Proteom. 2007, 1774, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Geng, M.; Liu, Y.; Zhao, W.; Xia, L.; Liu, J.; Qiu, G. Expression, purification and molecular modelling of the Iro protein from Acidithiobacillus ferrooxidans Fe-1. Protein Expr. Purif. 2007, 52, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.; Markova, N. Value of DSC in Characterization and Optimization of Protein Stability. In Microcalorimetry of Biological Molecules: Methods and Protocols; Ennifar, E., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1964, pp. 33–44. [Google Scholar]
- Wen, J.; Arthur, K.; Chemmalil, L.; Muzammil, S.; Gabrielson, J.; Jiang, Y. Applications of differential scanning calorimetry for thermal stability analysis of proteins: Qualification of DSC. J. Pharm. Sci. 2012, 101, 955–964. [Google Scholar] [CrossRef]
- Feng, J.; Cai, H.; Wang, H.; Li, C.; Liu, S. Improved oxidative stability of fish oil emulsion by grafted ovalbumin-catechin conjugates. Food Chem. 2018, 241, 60–69. [Google Scholar] [CrossRef]
- Brandes, N.; Welzel, P.B.; Werner, C.; Kroh, L.W. Adsorption-induced conformational changes of proteins onto ceramic particles: Differential scanning calorimetry and FTIR analysis. J. Colloid Interface Sci. 2006, 299, 56–69. [Google Scholar] [CrossRef]
- Wu, T.; Jiang, Q.; Wu, D.; Hu, Y.; Chen, S.; Ding, T.; Ye, X.; Liu, D.; Chen, J. What is new in lysozyme research and its application in food industry? A review. Food Chem. 2019, 274, 698–709. [Google Scholar] [CrossRef]
- Alderton, A.L.; Faustman, C.; Liebler, D.C.; Hill, D.W. Induction of redox instability of bovine myoglobin by adduction with 4-hydroxy-2-nonenal. Biochemistry 2003, 42, 4398–4405. [Google Scholar] [CrossRef] [PubMed]
- Sartor, G.; Mayer, E.; Johari, G.P. Calorimetric studies of the kinetic unfreezing of molecular motions in hydrated lysozyme, hemoglobin, and myoglobin. Biophys. J. 1994, 66, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, A.; Zscherp, C. What vibrations tell us about proteins. Q. Rev. Biophys. 2002, 35, 369–430. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Gutfreund, H. Proteins: Structures and molecular properties (second edition). FEBS Lett. 1993, 323, 294. [Google Scholar] [CrossRef] [Green Version]
- Ida, C.; Sasaki, K.; Ohmura, N.; Ando, A.; Saiki, H. Classification of rusticyanin structre genes from five different strains of Thiobacillus ferrooxidans into two groups. Process Metall. 1999, 9, 129–137. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Chen, L.; Shangguan, X.; Ouyang, J.; He, J.; Wang, K.; Tong, Y.; Yu, R.; Zeng, W.; Wu, X.; et al. Expression, Purification and Characterization of a Novel Rusticyanin from the Psychrotolerant Acidithiobacillus ferrivorans. Separations 2023, 10, 448. https://doi.org/10.3390/separations10080448
Liu Y, Chen L, Shangguan X, Ouyang J, He J, Wang K, Tong Y, Yu R, Zeng W, Wu X, et al. Expression, Purification and Characterization of a Novel Rusticyanin from the Psychrotolerant Acidithiobacillus ferrivorans. Separations. 2023; 10(8):448. https://doi.org/10.3390/separations10080448
Chicago/Turabian StyleLiu, Yuandong, Lixiang Chen, Xiangdong Shangguan, Jingying Ouyang, Jiayu He, Kan Wang, Yan Tong, Runlan Yu, Weimin Zeng, Xueling Wu, and et al. 2023. "Expression, Purification and Characterization of a Novel Rusticyanin from the Psychrotolerant Acidithiobacillus ferrivorans" Separations 10, no. 8: 448. https://doi.org/10.3390/separations10080448