
Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network

 , , , and
, , , and
Abstract
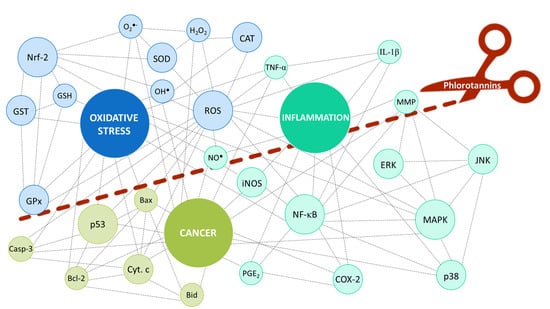
:
1. Introduction
2. Oxidative Stress, Inflammation and Cancer: How Are They Entangled?
3. The Contribution of Phlorotannins
3.1. Role in Oxidative Stress and Inflammation
3.2. Role in Cancer
4. Patent Review
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Stévant, P.; Rebours, C.; Chapman, A. Seaweed aquaculture in Norway: Recent industrial developments and future perspectives. Aquac. Int. 2017, 25, 1373–1390. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Zavaglia, A.; Prieto Lage, M.A.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The potential of seaweeds as a source of functional ingredients of prebiotic and antioxidant value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaza Martínez, J.H.; Torres Castañeda, H.G. Preparation and chromatographic analysis of phlorotannins. J. Chromatogr. Sci. 2013, 51, 825–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal Singh, I.; Bharate, S.B. Phloroglucinol compounds of natural origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef] [PubMed]
- Catarino, D.M.; Silva, M.A.; Cardoso, M.S. Fucaceae: A Source of Bioactive Phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Cancer. Available online: https://www.who.int/health-topics/cancer#tab=tab_1 (accessed on 6 May 2021).
- Ahn, G.-N.; Kim, K.-N.; Cha, S.-H.; Song, C.-B.; Lee, J.; Heo, M.-S.; Yeo, I.-K.; Lee, N.-H.; Jee, Y.-H.; Kim, J.-S.; et al. Antioxidant activities of phlorotannins purified from Ecklonia cava on free radical scavenging using ESR and H2O2-mediated DNA damage. Eur. Food Res. Technol. 2007, 226, 71–79. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.; Cruz, M.T.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Phlorotannins from Fucus vesiculosus: Modulation of inflammatory response by blocking NF-κB signaling pathway. Int. J. Mol. Sci. 2020, 21, 6897. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown seaweed Himanthalia elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Nwosu, F.; Morris, J.; Lund, V.A.; Stewart, D.; Ross, H.A.; McDougall, G.J. Anti-proliferative and potential anti-diabetic effects of phenolic-rich extracts from edible marine algae. Food Chem. 2011, 126, 1006–1012. [Google Scholar] [CrossRef]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Balamurugan, K.; Karthikeyan, J. Evaluation of luteolin in the prevention of N-nitrosodiethylamine-induced hepatocellular carcinoma using animal model system. Indian J. Clin. Biochem. 2012, 27, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruidenier, L.; Verspaget, H.W. Review article: Oxidative stress as a pathogenic factor in inflammatory bowel disease—Radicals or ridiculous? Aliment. Pharmacol. Ther. 2002, 16, 1997–2015. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Alves-Silva, J.M.; Pereira, O.R.; Cardoso, S.M. Antioxidant Capacities of Flavones and Benefits in Oxidative-Stress Related Diseases. Curr. Top. Med. Chem. 2015, 15, 105–119. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, E.H.; Hahm, K.B. Oxidative stress in inflammation-based gastrointestinal tract diseases: Challenges and opportunities. J. Gastroenterol. Hepatol. 2012, 27, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Schoneveld, O.; Georgakilas, A.G.; Panayiotidis, M.I. Oxidative stress, DNA methylation and carcinogenesis. Cancer Lett. 2008, 266, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Morgillo, F.; Tuccillo, C.; Ciardiello, F.; Loguercio, C. Chronic inflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 2007, 121, 2381–2386. [Google Scholar] [CrossRef]
- Corden, J.L.; Tollervey, D. Programmed Cell Death. In Cell Biology; Pollard, T.D., Earnshaw, W.C., Lippincott-Schwartz, J., Johnson, G.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 797–815. ISBN 978-0-323-34126-4. [Google Scholar]
- Green, D.R.; Llambi, F. Cell death signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef] [PubMed]
- Ďuračková, Z. Some current insights into oxidative stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Schwitalla, S.; Ziegler, P.K.; Horst, D.; Becker, V.; Kerle, I.; Begus-Nahrmann, Y.; Lechel, A.; Rudolph, K.L.; Langer, R.; Slotta-Huspenina, J.; et al. Loss of p53 in Enterocytes Generates an Inflammatory Microenvironment Enabling Invasion and Lymph Node Metastasis of Carcinogen-Induced Colorectal Tumors. Cancer Cell 2013, 23, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Olivier, M.; Hollstein, M.; Hainaut, P. TP53 mutations in human cancers: Origins, consequences, and clinical use. Cold Spring Harb. Perspect. Biol. 2010, 2, a001008. [Google Scholar] [CrossRef] [Green Version]
- García-Aranda, M.; Pérez-Ruiz, E.; Redondo, M. Bcl-2 inhibition to overcome resistance to chemo-and immunotherapy. Int. J. Mol. Sci. 2018, 19, 3950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-C.; Cha, S.H.; Wijesinghe, W.A.J.P.; Kang, S.-M.; Lee, S.-H.; Kim, E.-A.; Song, C.B.; Jeon, Y.-J. Protective effect of marine algae phlorotannins against AAPH-induced oxidative stress in zebrafish embryo. Food Chem. 2013, 138, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.R.; Moon, H.E.; Kim, H.R.; Jung, H.A.; Choi, J.S. Neuroprotective effect of edible brown alga Eisenia bicyclis on amyloid beta peptide-induced toxicity in PC12 cells. Arch. Pharm. Res. 2012, 35, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-Q.; Zhang, W.-T.; Shi, C.-H.; Wang, F.-M.; Tian, X.-J.; Ma, L.-L. Phloroglucinol protects the urinary bladder via inhibition of oxidative stress and inflammation in a rat model of cyclophosphamide-induced interstitial cystitis. Chin. Med. J. 2015, 128, 956–962. [Google Scholar] [CrossRef]
- Jun, Y.-J.; Lee, M.; Shin, T.; Yoon, N.; Kim, J.-H.; Kim, H.-R. Eckol enhances heme oxygenase-1 expression through activation of Nrf2/JNK pathway in HepG2 Cells. Molecules 2014, 19, 15638–15652. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Lee, B.; Park, K.E.; Utsuki, T.; Shin, T.; Oh, C.W.; Kim, H.R. Dieckol enhances the expression of antioxidant and detoxifying enzymes by the activation of Nrf2-MAPK signalling pathway in HepG2 cells. Food Chem. 2015, 174, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Wijesinghe, W.A.J.P.; Lee, S.H.; Kang, S.M.; Ko, S.C.; Yang, X.; Kang, N.; Jeon, B.T.; Kim, J.; Lee, D.H.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type II diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Z.-J.; Ryu, B.; Lee, S.-H.; Kim, M.-M.; Kim, S.-K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorgan. Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef]
- Kim, E.-A.; Kang, M.-C.; Lee, J.-H.; Kang, N.; Lee, W.; Oh, J.-Y.; Yang, H.-W.; Lee, J.-S.; Jeon, Y.-J. Protective effect of marine brown algal polyphenols against oxidative stressed zebrafish with high glucose. RSC Adv. 2015, 5, 25738–25746. [Google Scholar] [CrossRef]
- Lee, M.-S.; Shin, T.; Utsuki, T.; Choi, J.-S.; Byun, D.-S.; Kim, H.-R. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and hepatoprotective properties in tacrine-treated HepG2 cells. J. Agric. Food Chem. 2012, 60, 5340–5349. [Google Scholar] [CrossRef] [PubMed]
- Casas, M.P.; Rodríguez-Hermida, V.; Pérez-Larrán, P.; Conde, E.; Liveri, M.T.; Ribeiro, D.; Fernandes, E.; Domínguez, H. In vitro bioactive properties of phlorotannins recovered from hydrothermal treatment of Sargassum muticum. Sep. Purif. Technol. 2016, 167, 117–126. [Google Scholar] [CrossRef]
- Kim, A.-R.; Shin, T.-S.; Lee, M.-S.; Park, J.-Y.; Park, K.-E.; Yoon, N.-Y.; Kim, J.-S.; Choi, J.-S.; Jang, B.-C.; Byun, D.-S.; et al. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yuan, W.; Sharma-shivappa, R.; Zanten, J. Van Antioxidant activity of phlorotannins from brown algae. Int. J. Agric. Biol. Eng. 2017, 10, 184–191. [Google Scholar] [CrossRef]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Kageyama, N.; Nakahara, K.; Miki, W. Phlorotannins as radical scavengers from the extract of Sargassum ringgoldianum. Mar. Biotechnol. 2006, 8, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Ishimaru, K.; Kawaguchi, S.; Yoshikawa, H.; Hama, Y. Antioxidant activities of phlorotannins isolated from Japanese Laminariaceae. J. Appl. Phycol. 2008, 20, 705–711. [Google Scholar] [CrossRef]
- Zaragozá, M.C.; López, D.; Sáiz, M.P.; Poquet, M.; Pérez, J.; Puig-Parellada, P.; Màrmol, F.; Simonetti, P.; Gardana, C.; Lerat, Y.; et al. Toxicity and antioxidant activity in vitro and in vivo of two Fucus vesiculosus extracts. J. Agric. Food Chem. 2008, 56, 7773–7780. [Google Scholar] [CrossRef] [PubMed]
- Dutot, M.; Fagon, R.; Hemon, M.; Rat, P. Antioxidant, anti-inflammatory, and anti-senescence activities of a phlorotannin-rich natural extract from Brown Seaweed Ascophyllum nodosum. Appl. Biochem. Biotechnol. 2012, 167, 2234–2240. [Google Scholar] [CrossRef] [PubMed]
- Quéguineur, B.; Goya, L.; Ramos, S.; Martín, M.A.; Mateos, R.; Guiry, M.D.; Bravo, L. Effect of phlorotannin-rich extracts of Ascophyllum nodosum and Himanthalia elongata (Phaeophyceae) on cellular oxidative markers in human HepG2 cells. J. Appl. Phycol. 2013, 25, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Youn, K.; Kim, D.H.; Ahn, M.R.; Yoon, E.; Kim, O.Y.; Jun, M. Anti-Neuroinflammatory Property of Phlorotannins from Ecklonia cava on Aβ25–35-Induced Damage in PC12 Cells. Mar. Drugs 2019, 17, 7. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Ko, J.Y.; Oh, J.Y.; Kim, E.A.; Kim, C.Y.; Jeon, Y.J. Evaluation of phlorofucofuroeckol-A isolated from Ecklonia cava (Phaeophyta) on anti-lipid peroxidation in vitro and in vivo. Algae 2015, 30, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.C.; Piao, M.J.; Cho, S.J.; Lee, N.H.; Hyun, J.W. Phloroglucinol protects human keratinocytes from ultraviolet B radiation by attenuating oxidative stress. Photodermatol. Photoimmunol. Photomed. 2012, 28, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Ye, B.-R.; Heo, S.-J.; Oh, C.; Kang, D.-H.; Kim, J.H.; Affan, A.; Yoon, K.-T.; Choi, Y.-U.; Park, S.C.; et al. Photo-oxidative stress by ultraviolet-B radiation and antioxidative defense of eckstolonol in human keratinocytes. Environ. Toxicol. Pharmacol. 2012, 34, 926–934. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Hayes, M.; Kerry, J.P.; O’Brien, N.M. The effect of solvents on the antioxidant activity in Caco-2 cells of Irish brown seaweed extracts prepared using accelerated solvent extraction (ASE®). J. Funct. Foods 2013, 5, 940–948. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. In vitro and cellular antioxidant activities of seaweed extracts prepared from five brown seaweeds harvested in spring from the west coast of Ireland. Food Chem. 2011, 126, 1064–1070. [Google Scholar] [CrossRef]
- O’Sullivan, A.M.; O’Callaghan, Y.C.; O’Grady, M.N.; Queguineur, B.; Hanniffy, D.; Troy, D.J.; Kerry, J.P.; O’Brien, N.M. Assessment of the ability of seaweed extracts to protect against hydrogen peroxide and tert-butyl hydroperoxide induced cellular damage in Caco-2 cells. Food Chem. 2012, 134, 1137–1140. [Google Scholar] [CrossRef]
- Corona, G.; Coman, M.M.; Guo, Y.; Hotchkiss, S.; Gill, C.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Effect of simulated gastrointestinal digestion and fermentation on polyphenolic content and bioactivity of brown seaweed phlorotannin-rich extracts. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Kim, M.M.; Kim, S.K. Effect of phloroglucinol on oxidative stress and inflammation. Food Chem. Toxicol. 2010, 48, 2925–2933. [Google Scholar] [CrossRef]
- Kang, S.M.; Heo, S.J.; Kim, K.N.; Lee, S.H.; Jeon, Y.J. Isolation and identification of new compound, 2,7′-phloroglucinol-6,6′-bieckol from brown algae, Ecklonia cava and its antioxidant effect. J. Funct. Foods 2012, 4, 158–166. [Google Scholar] [CrossRef]
- Zou, Y.; Qian, Z.-J.; Li, Y.; Kim, M.-M.; Lee, S.-H.; Kim, S.-K. Antioxidant effects of phlorotannins isolated from Ishige okamurae in free radical mediated oxidative systems. J. Agric. Food Chem. 2008, 56, 7001–7009. [Google Scholar] [CrossRef] [PubMed]
- Guinea, M.; Franco, V.; Araujo-Bazán, L.; Rodríguez-Martín, I.; González, S. In vivo UVB-photoprotective activity of extracts from commercial marine macroalgae. Food Chem. Toxicol. 2012, 50, 1109–1117. [Google Scholar] [CrossRef]
- Kang, M.C.; Kim, K.N.; Kang, S.M.; Yang, X.; Kim, E.A.; Song, C.B.; Nah, J.W.; Jang, M.K.; Lee, J.S.; Jung, W.K.; et al. Protective effect of dieckol isolated from Ecklonia cava against ethanol caused damage in vitro and in zebrafish model. Environ. Toxicol. Pharmacol. 2013, 36, 1217–1226. [Google Scholar] [CrossRef]
- Ko, S.-C.; Cha, S.-H.; Heo, S.-J.; Lee, S.-H.; Kang, S.-M.; Jeon, Y.-J. Protective effect of Ecklonia cava on UVB-induced oxidative stress: In Vitro and in vivo zebrafish model. J. Appl. Phycol. 2011, 23, 697–708. [Google Scholar] [CrossRef]
- Wei, Y.; Li, Z.; Hu, Y.; Xu, Z. Inhibition of mouse liver lipid peroxidation by high molecular weight phlorotannins from Sargassum kjellmanianum. J. Appl. Phycol. 2003, 15, 507–511. [Google Scholar] [CrossRef]
- Sprygin, V.G.; Kushnerova, N.F.; Fomenko, S.E.; Drugova, E.S.; Lesnikova, L.N.; Merzlyakov, V.Y.; Momot, T.V. The Influence of an Extract from the Marine Brown Alga Sargassum pallidum on the Metabolic Reactions in the Liver under Experimental Toxic Hepatitis. Russ. J. Mar. Biol. 2017, 43, 479–484. [Google Scholar] [CrossRef]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.W.; Pourshahidi, L.K.; et al. Impact of a (poly)phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef]
- Schultz Moreira, A.R.; Benedí, J.; González-Torres, L.; Olivero-David, R.; Bastida, S.; Sánchez-Reus, M.I.; González-Muñoz, M.J.; Sánchez-Muniz, F.J. Effects of diet enriched with restructured meats, containing Himanthalia elongata, on hypercholesterolaemic induction, CYP7A1 expression and antioxidant enzyme activity and expression in growing rats. Food Chem. 2011, 129, 1623–1630. [Google Scholar] [CrossRef]
- Jin, D.Q.; Li, G.; Kim, J.S.; Yong, C.S.; Kim, J.A.; Huh, K. Preventive effects of Laminaria japonica aqueous extract on the oxidative stress and xanthine oxidase activity in streptozotocin-induced diabetic rat liver. Biol. Pharm. Bull. 2004, 27, 1037–1040. [Google Scholar] [CrossRef] [Green Version]
- Sadeeshkumar, V.; Duraikannu, A.; Aishwarya, T.; Jayaram, P.; Ravichandran, S.; Ganeshamurthy, R. Radioprotective efficacy of dieckol against gamma radiation-induced cellular damage in hepatocyte cells. Naunyn Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Kang, S.M.; Ahn, G.; Kim, K.N.; Kang, N.; Samarakoon, K.W.; Oh, M.C.; Lee, J.S.; Jeon, Y.J. Protective effect of a marine polyphenol, dieckol against carbon tetrachloride-induced acute liver damage in mouse. Environ. Toxicol. Pharmacol. 2013, 35, 517–523. [Google Scholar] [CrossRef]
- Li, S.; Liu, J.; Zhang, M.; Chen, Y.; Zhu, T.; Wang, J. Protective Effect of Eckol against Acute Hepatic injury Induced by Carbon Tetrachloride in Mice. Mar. Drugs 2018, 16, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.D.; Kang, K.A.; Piao, M.J.; Kim, K.C.; Zheng, J.; Yao, C.W.; Cha, J.W.; Hyun, C.L.; Kang, H.K.; Lee, N.H.; et al. Cytoprotective effect of eckol against oxidative stress-induced mitochondrial dysfunction: Involvement of the FoxO3a/AMPK pathway. J. Cell. Biochem. 2014, 115, 1403–1411. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Lee, K.H.; Chae, S.; Zhang, R.; Jung, M.S.; Ham, Y.M.; Baik, J.S.; Lee, N.H.; Hyun, J.W. Cytoprotective effect of phloroglucinol on oxidative stress induced cell damage via catalase activation. J. Cell. Biochem. 2006, 97, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Lee, K.H.; Chae, S.; Koh, Y.S.; Yoo, B.S.; Kim, J.H.; Ham, Y.M.; Baik, J.S.; Lee, N.H.; Hyun, J.W. Triphlorethol-A from Ecklonia cava protects V79-4 lung fibroblast against hydrogen peroxide induced cell damage. Free Radic. Res. 2005, 39, 883–892. [Google Scholar] [CrossRef]
- Yokogawa, K.; Matsui-Yuasa, I.; Tamura, A.; Terada, M.; Kojima-Yuasa, A. Inhibitory effects of Ecklonia cava extract on high glucose-induced hepatic stellate cell activation. Mar. Drugs 2011, 9, 2783–2808. [Google Scholar] [CrossRef] [Green Version]
- Quéguineur, B.; Goya, L.; Ramos, S.; Martín, M.A.; Mateos, R.; Bravo, L. Phloroglucinol: Antioxidant properties and effects on cellular oxidative markers in human HepG2 cell line. Food Chem. Toxicol. 2012, 50, 2886–2893. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.C.; Kang, K.A.; Zhang, R.; Piao, M.J.; Kim, G.Y.; Kang, M.Y.; Lee, S.J.; Lee, N.H.; Surh, Y.J.; Hyun, J.W. Up-regulation of Nrf2-mediated heme oxygenase-1 expression by eckol, a phlorotannin compound, through activation of Erk and PI3K/Akt. Int. J. Biochem. Cell Biol. 2010, 42, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.-J.; Ahn, S.; Ryu, J.; Choi, M.-S.; Choi, S.; Chong, Y.H.; Hyun, J.-W.; Chang, M.-J.; Kim, H.-S. Phloroglucinol attenuates motor functional deficits in an animal model of Parkinson’s disease by enhancing Nrf2 activity. PLoS ONE 2013, 8, e71178. [Google Scholar] [CrossRef]
- Moon, H.E.; Ahn, B.R.; Jung, H.A.; Choi, J.S. Inhibitory activity of Ecklonia stolonifera and its isolated phlorotannins against Cu2+-induced low-density lipoprotein oxidation. Fish. Sci. 2012, 78, 927–934. [Google Scholar] [CrossRef]
- Shin, H.C.; Hwang, H.J.; Kang, K.J.; Lee, B.H. An antioxidative and antiinflammatory agent for potential treatment of osteoarthritis from Ecklonia cava. Arch. Pharm. Res. 2006, 29, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, J.; Keekan, K.K.; Anil, S.; Bhatnagar, I.; Kim, S.K. Phlorotannins. Encycl. Food Chem. 2018, 3, 515–527. [Google Scholar] [CrossRef]
- Kim, T.H.; Ku, S.-K.; Lee, T.; Bae, J.-S. Vascular barrier protective effects of phlorotannins on HMGB1-mediated proinflammatory responses in vitro and in vivo. Food Chem. Toxicol. 2012, 50, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-M.; Eom, S.-H.; Kim, Y.-M. Protective effect of phlorotannins from Eisenia bicyclis against lipopolysaccharide-stimulated inflammation in HepG2 cells. Environ. Toxicol. Pharmacol. 2013, 35, 395–401. [Google Scholar] [CrossRef]
- Jung, W.-K.; Heo, S.-J.; Jeon, Y.-J.; Lee, C.-M.; Park, Y.-M.; Byun, H.-G.; Choi, Y.H.; Park, S.-G.; Choi, I.-W. Inhibitory effects and molecular mechanism of dieckol isolated from marine brown alga on COX-2 and iNOS in microglial cells. J. Agric. Food Chem. 2009, 57, 4439–4446. [Google Scholar] [CrossRef]
- Paudel, U.; Lee, Y.H.; Kwon, T.H.; Park, N.H.; Yun, B.S.; Hwang, P.H.; Yi, H.K. Eckols reduce dental pulp inflammation through the ERK1/2 pathway independent of COX-2 inhibition. Oral Dis. 2014, 20, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Choi, S.; Kim, G.; Imm, J. Anti-Inflammatory Effect of Ecklonia cava Extract on Porphyromonas gingivalis Lipopolysaccharide-Stimulated Macrophages and a Periodontitis Rat Model. Nutrients 2019, 11, 1143. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.K.; Ahn, Y.W.; Lee, S.H.; Choi, Y.H.; Kim, S.K.; Yea, S.S.; Choi, I.; Park, S.G.; Seo, S.K.; Lee, S.W.; et al. Ecklonia cava ethanolic extracts inhibit lipopolysaccharide-induced cyclooxygenase-2 and inducible nitric oxide synthase expression in BV2 microglia via the MAP kinase and NF-κB pathways. Food Chem. Toxicol. 2008, 47, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Ham, Y.M.; Kim, D.S.; Kim, J.Y.; Hong, J.P.; Kim, M.J.; Moon, J.Y.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Ecklonia stolonifera inhibits lipopolysaccharide-induced production of nitric oxide, prostaglandin E2, and proinflammatory cytokines in RAW264.7 macrophages. Biologia 2010, 65, 362–371. [Google Scholar] [CrossRef]
- Lee, M.-S.; Kwon, M.-S.; Choi, J.-W.; Shin, T.; No, H.-K.; Choi, J.-S.; Byun, D.-S.; Kim, J.-I.; Kim, H.-R. Anti-inflammatory activities of an ethanol extract of Ecklonia stolonifera in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. J. Agric. Food Chem. 2012, 60, 9120–9129. [Google Scholar] [CrossRef]
- Tenorio-Rodríguez, P.A.; Esquivel-Solis, H.; Murillo-Álvarez, J.I.; Ascencio, F.; Campa-Córdova, Á.I.; Angulo, C. Biosprospecting potential of kelp (Laminariales, Phaeophyceae) from Baja California Peninsula: Phenolic content, antioxidant properties, anti-inflammatory, and cell viability. J. Appl. Phycol. 2019, 31, 3115–3129. [Google Scholar] [CrossRef]
- Lee, S.-H.; Eom, S.-H.; Yoon, N.-Y.; Kim, M.-M.; Li, Y.-X.; Ha, S.K.; Kim, S.-K. Fucofuroeckol-A from Eisenia bicyclis Inhibits Inflammation in Lipopolysaccharide-Induced Mouse Macrophages via Downregulation of the MAPK/NF-kappa B Signaling Pathway. J. Chem. 2016, 2016, 6509212. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-I.; Shin, H.-C.; Kim, S.H.; Park, W.-Y.; Lee, K.-T.; Choi, J.-H. 6,6′-Bieckol, isolated from marine alga Ecklonia cava, suppressed LPS-induced nitric oxide and PGE 2 production and inflammatory cytokine expression in macrophages: The inhibition of NFκB. Int. Immunopharmacol. 2012, 12, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Lopes, G.; Ferreres, F.; Andrade, P.B.; Pereira, D.M.; Gil-Izquierdo, Á.; Valentão, P. Phlorotannin extracts from Fucales: Marine polyphenols as bioregulators engaged in inflammation-related mediators and enzymes. Algal Res. 2017, 28, 1–8. [Google Scholar] [CrossRef]
- Kim, M.E.; Jung, Y.C.; Jung, I.; Lee, H.W.; Youn, H.Y.; Lee, J.S. Anti-inflammatory effects of ethanolic extract from Sargassum horneri (Turner) C. Agardh on lipopolysaccharide-stimulated macrophage activation via NF-κB pathway regulation. Immunol. Investig. 2015, 44, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, K.B.W.R.; Jeong, D.H.; Ahn, D.H. Anti-inflammatory activity of ethanolic extract of Sargassum sagamianum in RAW 264.7 cells. Food Sci. Biotechnol. 2013, 22, 1113–1120. [Google Scholar] [CrossRef]
- Kim, K.N.; Kim, J.; Yoon, W.J.; Yang, H.M.; Heo, S.Y.; Ko, J.Y.; Roh, S.W.; Jeon, Y.J.; Kang, S.M.; Heo, S.J.; et al. Inhibitory effect of Sargassum patens on inflammation and melanogenesis. Int. J. Pharmacol. 2013, 9, 524–532. [Google Scholar] [CrossRef]
- Lee, C.; Park, G.H.; Ahn, E.M.; Park, C.I.; Jang, J.H. Sargassum fulvellum protects HaCaT cells and BALB/c mice from UVB-induced proinflammatory responses. Evid. Based Complement. Altern. Med. 2013, 2013, 747846. [Google Scholar] [CrossRef]
- Yayeh, T.; Im, E.J.; Kwon, T.H.; Roh, S.S.; Kim, S.; Kim, J.H.; Hong, S.B.; Cho, J.Y.; Park, N.H.; Rhee, M.H. Hemeoxygenase 1 partly mediates the anti-inflammatory effect of dieckol in lipopolysaccharide stimulated murine macrophages. Int. Immunopharmacol. 2014, 22, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.-R.; Lee, M.-S.; Shin, T.-S.; Hua, H.; Jang, B.-C.; Choi, J.-S.; Byun, D.-S.; Utsuki, T.; Ingram, D.; Kim, H.-R. Phlorofucofuroeckol A inhibits the LPS-stimulated iNOS and COX-2 expressions in macrophages via inhibition of NF-κB, Akt, and p38 MAPK. Toxicol. In Vitro 2011, 25, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Lee, B.; Joung, E.J.; Gwon, W.G.; Utsuki, T.; Kim, N.G.; Kim, H.R. 6,6′-Bieckol suppresses inflammatory responses by down-regulating nuclear factor-κB activation via Akt, JNK, and p38 MAPK in LPS-stimulated microglial cells. Immunopharmacol. Immunotoxicol. 2016, 38, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, A.; Jouini, M.; Bel Haj Amor, H.; Mzoughi, Z.; Dridi, M.; Ben Said, R.; Bouraoui, A. Phytochemical Analysis and Evaluation of the Antioxidant, Anti-Inflammatory, and Antinociceptive Potential of Phlorotannin-Rich Fractions from Three Mediterranean Brown Seaweeds. Mar. Biotechnol. 2018, 20, 60–74. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Lee, H.G.; Herath, K.H.I.N.M.; Jee, Y.; Jeon, Y.J. The protective effect of Sargassum horneri against particulate matter-induced inflammation in lung tissues of an in vivo mouse asthma model. Food Funct. 2019, 10, 7995–8004. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.J.; Lee, M.S.; Choi, J.W.; Kim, J.S.; Shin, T.; Jung, B.M.; Yoon, N.Y.; Lim, C.W.; Kim, J.I.; Kim, H.R. Anti-inflammatory effect of ethanolic extract from Myagropsis myagroides on murine macrophages and mouse ear edema. BMC Complement. Altern. Med. 2012, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.I.; Woo, J.H.; Seo, Y.J.; Lee, K.T.; Lim, Y.; Choi, J.H. Protective Effect of Brown Alga Phlorotannins against Hyper-inflammatory Responses in Lipopolysaccharide-Induced Sepsis Models. J. Agric. Food Chem. 2016, 64, 570–578. [Google Scholar] [CrossRef]
- Corona, G.; Ji, Y.; Anegboonlap, P.; Hotchkiss, S.; Gill, C.; Yaqoob, P.; Spencer, J.P.E.; Rowland, I. Gastrointestinal modifications and bioavailability of brown seaweed phlorotannins and effects on inflammatory markers. Br. J. Nutr. 2016, 15, 1240–1253. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, Y.; Usui, M.; Katsuzaki, H.; Imai, K.; Miyata, M. Anti-inflammatory effects of 6,6′-bieckol and 6,8′-bieckol from Eisenia arborea on mouse ear swelling. Food Sci. Technol. Res. 2017, 23, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, Y.; Usui, M.; Katsuzaki, H.; Imai, K.; Kakinuma, M.; Amano, H.; Miyata, M. Orally Administered Phlorotannins from Eisenia arborea Suppress Chemical Mediator Release and Cyclooxygenase-2 Signaling to Alleviate Mouse Ear Swelling. Mar. Drugs 2018, 16, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, Y.; Tanaka, R.; Katsuzaki, H.; Imai, K.; Matsushita, T. The anti-inflammatory effects of phlorotannins from Eisenia arborea on mouse ear edema by inflammatory inducers. J. Funct. Foods 2013, 5, 2019–2023. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Kim, H.S.; Song, K.S.; Han, H.J.; Park, S.H.; Chang, W.; Lee, M.Y. Protective effect of dieckol against chemical hypoxia-induced cytotoxicity in primary cultured mouse hepatocytes. Drug Chem. Toxicol. 2015, 38, 180–187. [Google Scholar] [CrossRef]
- Cha, S.H.; Ko, C.I.; Kim, D.; Jeon, Y.J. Protective effects of phlorotannins against ultraviolet B radiation in zebrafish (Danio rerio). Vet. Dermatol. 2012, 23, 51–e12. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, J.; Lee, H.; Kim, B.; Lew, J.; Baek, N.; Kim, S.-H. MicroRNA134 Mediated Upregulation of JNK and Downregulation of NFkB Signalings Are Critically Involved in Dieckol Induced Antihepatic Fibrosis. J. Agric. Food Chem. 2016, 64, 5508–5514. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Kim, J.A.; Ahn, B.N.; Kim, S.K. Potential effect of phloroglucinol derivatives from Ecklonia cava on matrix metalloproteinase expression and the inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages. Fish. Sci. 2011, 77, 867–873. [Google Scholar] [CrossRef]
- Lee, S.-H.; Han, J.-S.; Heo, S.-J.; Hwang, J.-Y.; Jeon, Y.-J. Protective effects of dieckol isolated from Ecklonia cava against high glucose-induced oxidative stress in human umbilical vein endothelial cells. Toxicol. In Vitro 2010, 24, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Li, Y.; Qian, Z.J.; Kim, M.M.; Kim, S.K. Differentiation of human osteosarcoma cells by isolated phlorotannins is subtly linked to COX-2, iNOS, MMPs, and MAPK signaling: Implication for chronic articular disease. Chem. Biol. Interact. 2009, 179, 192–201. [Google Scholar] [CrossRef]
- Yang, Y.I.; Jung, S.H.; Lee, K.T.; Choi, J.H. 8,8′-Bieckol, isolated from edible brown algae, exerts its anti-inflammatory effects through inhibition of NF-κB signaling and ROS production in LPS-stimulated macrophages. Int. Immunopharmacol. 2014, 23, 460–468. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, anti-inflammatory and antiproliferative effects of aqueous extracts of three mediterranean brown seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar] [CrossRef]
- Ahn, J.H.; Yang, Y.I.; Lee, K.T.; Choi, J.H. Dieckol, isolated from the edible brown algae Ecklonia cava, induces apoptosis of ovarian cancer cells and inhibits tumor xenograft growth. J. Cancer Res. Clin. Oncol. 2014, 141, 255–268. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, Y.; Chen, Y.; Liu, H.; Yuan, G.; Fan, Y.; Chen, K. Optimization of the microwave-assisted extraction of phlorotannins from Saccharina japonica Aresch and evaluation of the inhibitory effects of phlorotannin-containing extracts on HepG2 cancer cells. Chin. J. Oceanol. Limnol. 2013, 31, 1045–1054. [Google Scholar] [CrossRef]
- Abdelhamid, A.; Lajili, S.; Elkaibi, M.A.; Ben Salem, Y.; Abdelhamid, A.; Muller, C.D.; Majdoub, H.; Kraiem, J.; Bouraoui, A. Optimized Extraction, Preliminary Characterization and Evaluation of the In Vitro Anticancer Activity of Phlorotannin-Rich Fraction from the Brown Seaweed, Cystoseira sedoides. J. Aquat. Food Prod. Technol. 2019, 28, 892–909. [Google Scholar] [CrossRef]
- Imbs, T.; Krasovskaya, N.; Ermakova, S.; Makarieva, T.; Shevchenko, N.; Zvyagintseva, T. Comparative study of chemical composition and antitumor activity of aqueous-ethanol extracts of brown algae Laminaria cichorioides, Costaria costata, and Fucus evanescens. Russ. J. Mar. Biol. 2009, 35, 164–170. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Walsh, N.A. Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem. Toxicol. 2006, 44, 1144–1150. [Google Scholar] [CrossRef]
- Montero, L.; Sánchez-Camargo, A.P.; García-Cañas, V.; Tanniou, A.; Stiger-Pouvreau, V.; Russo, M.; Rastrelli, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Anti-proliferative activity and chemical characterization by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry of phlorotannins from the brown macroalga Sargassum muticum collected on North-Atlantic coasts. J. Chromatogr. A 2016, 1428, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Geisen, U.; Zenthoefer, M.; Peipp, M.; Kerber, J.; Plenge, J.; Managò, A.; Fuhrmann, M.; Geyer, R.; Hennig, S.; Adam, D.; et al. Molecular mechanisms by which a Fucus vesiculosus extract mediates cell cycle inhibition and cell death in pancreatic cancer cells. Mar. Drugs 2015, 13, 4470–4491. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Kang, C.; Jung, E.S.; Kim, J.S.; Kim, E. Antimetastatic activity of polyphenol-rich extract of Ecklonia cava through the inhibition of the Akt pathway in A549 human lung cancer cells. Food Chem. 2011, 127, 1229–1236. [Google Scholar] [CrossRef]
- Kim, M.M.; Van Ta, Q.; Mendis, E.; Rajapakse, N.; Jung, W.K.; Byun, H.G.; Jeon, Y.J.; Kim, S.K. Phlorotannins in Ecklonia cava extract inhibit matrix metalloproteinase activity. Life Sci. 2006, 79, 1436–1443. [Google Scholar] [CrossRef]
- Karadeniz, F.; Lee, S.G.; Oh, J.H.; Yu, G.H.; Jang, M.S.; Seo, Y.; Kong, C.S. Solvent-partitioned fractions from Ishige okamurae extract inhibit MMP-2 and MMP-9 activities in human fibrosarcoma cells in vitro. J. Appl. Phycol. 2018, 30, 121–127. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Shi, X.; Kim, S.K. Inhibition of the expression on MMP-2, 9 and morphological changes via human fibrosarcoma cell line by 6,6′-bieckol from marine alga Ecklonia cava. BMB Rep. 2010, 43, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.H.; Li, X.F.; Jin, L.F.; Zhao, Y.; Zhu, G.J.; Shen, W.Z. Dieckol inhibits non-small-cell lung cancer cell proliferation and migration by regulating the PI3K/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeeshkumar, V.; Duraikannu, A.; Ravichandran, S. Modulatory efficacy of dieckol on xenobiotic-metabolizing enzymes, cell proliferation, apoptosis, invasion and angiogenesis during NDEA-induced rat hepatocarcinogenesis. Mol. Cell. Biochem. 2017, 433, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Tang, Y.; Kim, Y.S.; Hwang, J.W.; Choi, E.J.; Lee, J.H.; Lee, S.H.; Jeon, Y.U.J.; Park, P.J. First evidence that Ecklonia cava-derived dieckol attenuates MCF-7 human breast carcinoma cell migration. Mar. Drugs 2015, 13, 1785–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.-M.; Park, C.G.; Kang, J.H.; Kim, E.-J.; Chee, H.Y.; Lee, B.H.; Lee, K.B. Dieckol inhibits 12-O-tetradecanoylphorbol-13-acetate-induced SK-Hep1 human hepatoma cell motility through suppression of matrix metalloproteinase-9 activity. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 376–381. [Google Scholar] [CrossRef]
- Lee, Y.J.; Park, J.H.; Park, S.A.; Joo, N.R.; Lee, B.H.; Lee, K.B.; Oh, S.M. Dieckol or phlorofucofuroeckol extracted from Ecklonia cava suppresses lipopolysaccharide-mediated human breast cancer cell migration and invasion. J. Appl. Phycol. 2020, 32, 631–640. [Google Scholar] [CrossRef]
- Yoon, J.S.; Kasin Yadunandam, A.; Kim, S.J.; Woo, H.C.; Kim, H.R.; Kim, G.D. Dieckol, isolated from Ecklonia stolonifera, induces apoptosis in human hepatocellular carcinoma Hep3B cells. J. Nat. Med. 2013, 67, 519–527. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Guo, J.; Hu, X.M.; Zhao, S.Q.; Li, S.L.; Wang, J. An in vivo anti-tumor effect of eckol from marine brown algae by improving the immune response. Food Funct. 2019, 10, 4361–4371. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef]
- Kang, M.H.; Kim, I.H.; Nam, T.J. Phloroglucinol induces apoptosis via apoptotic signaling pathways in HT-29 colon cancer cells. Oncol. Rep. 2014, 32, 1341–1346. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Kim, I.H.; Nam, T.J. Phloroglucinol induces apoptosis through the regulation of insulin-like growth factor 1 receptor signaling pathways in human colon cancer HT-29 cells. Int. J. Oncol. 2014, 45, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Eo, H.J.; Kwon, T.H.; Park, G.H.; Song, H.M.; Lee, S.J.; Park, N.H.; Jeong, J.B. In Vitro Anticancer Activity of Phlorofucofuroeckol A via Upregulation of Activating Transcription Factor 3 against Human Colorectal Cancer Cells. Mar. Drugs 2016, 14, 69. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Avila, G.; Sommer, B.; Mendoza-Posada, D.A.; Ramos, C.; Garcia-Hernandez, A.A.; Falfan-Valencia, R. Matrix metalloproteinases participation in the metastatic process and their diagnostic and therapeutic applications in cancer. Crit. Rev. Oncol. Hematol. 2019, 137, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.Y.; Ham, Y.M.; Jeong, D.J.; Kim, H.B.; Kim, J.H.; Song, Y.K.; Yoo, B.S. Ecklonia cava Extract with High Antioxidant Activity and Process for Preparing the Same Using Supercritical Carbon Dioxide Extraction. KR101217472B1, 2 January 2013. [Google Scholar]
- Garcia, M.J.N.; Guerrero, M.S.; Torres, J.S. Antioxidant Extract from Brown Macroalgae and Method for Obtaining Same. US20160074317A1, 17 March 2016. [Google Scholar]
- Choi, J.S.; Jung, H.Y.; Jung, J.H.; Kang, H.S.; Son, B.H. Composition Comprising Phlorotannins Isolated from the Extract of Ecklonia stolonifera Okamura Having Anti-Oxidative Activity. KR100512482B1, 5 September 2005. [Google Scholar]
- Kim, M.M.; Kim, S.K.; Lee, S.H.; Yanping, Z. An Anti-Oxidant Active Composition Containing Compounds from Ishige okamurae. KR20100039104A, 15 April 2010. [Google Scholar]
- Choi, J.S.; Kim, A.R.; Kim, H.R.; Kwon, M.S.; Shin, T.S.; Woo, H.C. Anti-Inflammatory Composition Containing Phlorotannins from Ecklonia stolonifera and Ecklonia cava Extract as an Effective Component. KR20110035127A, 6 April 2011. [Google Scholar]
- Besnard, M.; Grandieu, P.; Inisan, C.; Lerat, Y.; Megard, D.; Mitjavila, M.T.; Rousseau, I.; Simonetti, P. Algae Extract Containing Polyphenols. EP1977756A1, 8 October 2008. [Google Scholar]
- Kim, M.M.; Kim, S.K. Composition Containing Phlorotannin for Inhibition of Matrix Metalloproteinase Activities. KR100858847B1, 17 September 2008. [Google Scholar]
- Hyun, J.W.; Hyun, K.H.; Kim, M.J.; Lee, N.H.; Lee, S.J. A Composition Containing Eckol for Inhibiting a Growth of Cancer Stem Cells. KR20120053265A, 23 September 2013. [Google Scholar]
- Hyiun, J.W.; Kim, R.K.; Lee, S.J.; Yoon, C.H. An Anti-Cancer Composition Containing Eckol for Cancer Expressing k-Ras. KR101596989B1, 23 February 2016. [Google Scholar]

{kind=link}
| Extract/Compound | Model | Conditions | Effect | Ref. |
|---|---|---|---|---|
| Sargassum kjellmanianum | ||||
| EtOH 85%→Fract. CHCl3→PE→cellulose column→HMW phlorotannins | CCl4 or FeSO4-VC-induced liver homogenate in Kumming mice | Oral administration with 5.0 g/kg before obtaining liver homogenate | ↓ MDA, lipid peroxidation and swelling | [60] |
| Sargassum pallidum | ||||
| Ac 70%→Fract. DCM→EtOAc | Wistar rats with CCl4-induced hepatic injury | Oral administration of 100 mg/kg for 1 week after CCl4 injection | ↓ MDA; ↑ GSH and SOD | [61] |
| Fucus vesiculosus | ||||
| Ext. 1: EtOH 30%–35% Ext. 2: EtOH 50%–70% | Sprague-dawley rats | Oral administration of 200 mg/kg/day for 4 weeks | ↑ antioxidant defenses, PON-1 and ↓ MDA (in plasm) and ↑ SOD (in rats’ erythrocytes) | [43] |
| Lessonia vadosa | ||||
| Ac→Ac 70% | UVB-induced zebrafish | 40 µg/mL PGE prior to irradiation (0.15 mW/cm2) | ↓ ROS and malformations | [57] |
| Macrocystis pyrifera | ||||
| Ac→Ac 70% | UVB-induced zebrafish | 40 µg/mL PGE prior to irradiation (0.15 mW/cm2) | ↓ ROS and malformations | [57] |
| Ascophyllum nodosum | ||||
| EtOH 60%→Fract. tangential flow ultrafiltration | Human trials | Oral administration of a food-grade capsule containing 100 mg of polyphenols for 8 weeks | ↓ DNA damage in obese subjects | [62] |
| Himanthalia enlogata | ||||
| Restructured pork with 5% seaweed powder | Hypercholesterolaemia-induced Wistar rats | Feeding ad libitum over 5 weeks | ↑ CYP7A1, GR and SOD levels in liver homegenates | [63] |
| Laminaria japonica | ||||
| Aqueous extract | STZ-induced diabetes in Sprague-Dawley rats | Oral gavage of 100 mg/kg/d for 5 days prior to STZ injection (55 mg/kg) | ↓ lipid peroxidation, XO; ↑ GSH, GPx and GR | [64] |
| Isolated compounds | ||||
| Dieckol from Ecklonia stolonifera | ICR mice | Oral administration of 100 mg/kg/d for 4 days | ↑ HO-1 | [32] |
| 6,6-bieckol | High glucose-induced zebrafish | 20 µM 16 prior to glucose stimulation | ↓ ROS | [35] |
| Dieckol from Ecklonia cava | γ-radiation-induced hepatocyte cells isolated from Wistar rats | 5–20 µM 1 h prior to irradiation (8 Gy/min) | ↓ lipid peroxidation, DNA damage; ↑ SOD, CAT, GSH and GPx | [65] |
| C57BL/KsJ-db/db mice | 10 and 20 10 mg/kg/d over 14 days | ↓ lipid peroxidation; ↑ SOD, GPx and CAT | [33] | |
| CCl4-induced hepatic injury in ICR mice | Oral administration of 5 mg/kg/d 6 days prior to CCl4 administration (0.5 mg/kg) | ↓ MDA; ↑ SOD, GPx and CAT | [66] | |
| AAPH-induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS, lipid peroxidation and cell death | [28] | |
| High glucose-induced zebrafish | 20 µM 16 h prior to glucose exposure (50–300 mM) | ↓ ROS cell death and lipid peroxidation | [35] | |
| EtOH-induced zebrafish | 10 and 20 µM 1 h prior to ethanol exposure (0.5%) | ↓ ROS, lipid peroxidation and cell death | [58] | |
| UVB-induced zebrafish | 50 µM 1 h prior to irradiation (50 mJ/cm) | ↓ ROS and cell death | [59] | |
| Eckol from Ecklonia cava | AAPH-induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS, lipid peroxidation and cell death | [28] |
| EtOH-induced zebrafish | 10 and 20 µM 1 h prior to ethanol exposure (0.5%) | ↓ ROS, lipid peroxidation and cell death | [58] | |
| Eckol commercial | CCl4-induced hepatic injury in Kumming mice | Oral administration of 0.5 and 1 mg/kg/d 7 days prior to CCl4 injection (0.5%) | ↓ MDA; ↑ SOD, GPx and GSH | [67] |
| Eckstolonol from Ecklonia cava | AAPH-induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS, lipid peroxidation and cell death | [28] |
| Phloroeckol from Ecklonia cava | High glucose-induced zebrafish | 20 µM 16 h prior to glucose exposure (50–300 mM) | ↓ ROS | [35] |
| Phlorofucofuroeckol A from Ecklonia cava | High glucose-induced zebrafish | 20 µM 16 h prior to glucose exposure (50–300 mM) | ↓ ROS | [35] |
| AAPH- induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS and cell death | [47] | |
| Phloroglucinol from Ecklonia cava | AAPH-induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS, lipid peroxidation and cell death | [28] |
| EtOH-induced zebrafish | 10 and 20 µM 1 h prior to ethanol exposure (0.5%) | ↓ ROS, lipid peroxidation and cell death | [58] | |
| Phloroglucinol commercial | CYP-induced in Sprague-Dawley rat | 15 and 30 mg/kg/d i.p. over 7 + 75 mg/kg of CYP i.p. at day 1, 4 and 7 | ↓ MPO and ↑ CAT | [30] |
| Triphloroethol A from Ecklonia cava | AAPH-induced zebrafish | 50 µM 1 h prior to AAPH exposure (25 mM) | ↓ ROS, lipid peroxidation and cell death | [28] |
| Extract/Compound | Model | Conditions | Effect | Ref. |
|---|---|---|---|---|
| Cystoseira sedoides | ||||
| EtOH 50% → Fract. PE → DCM → EtOAc | Model 1—XYL-induced mice ear edema Models 2—Carrageenan-induced paw edema in Wistar rats | Model 1—50 and 100 mg EtOAc/kg s.c. 30 min prior to XYL application Model 2—50 and 100 mg EtOAc/kg i.p. 30 min prior to carrageenan application | Model 1—↓ swelling up to 83% Model 2—↓ swelling up to 81% | [98] |
| Cladostephus spongiosis | ||||
| EtOH 50% → Fract. PE → DCM → EtOAc | Model 1—XYL-induced mice ear edema Models 2—Carrageenan-induced paw edema in Wistar rats | Model 1—50 and 100 mg EtOAc/kg s.c. 30 min prior to XYL Model 2—50 and 100 mg EtOAc/kg i.p. 30 min prior to carrageenan | Model 1—↓ swelling up to 69% Model 2—↓ swelling up to 71% | [98] |
| Padina pavonica | ||||
| EtOH 50% → Fract. PE → DCM → EtOAc | Carrageenan-induced paw edema in Wistar rats | 50 and 100 mg EtOAc/kg i.p. 30 min prior to carrageenan | ↓ swelling up to 58% | [98] |
| Sargassum horneri | ||||
| EtOH 70% | OVA + PM-induced lung inflammation in BALB/c mice | 200 and 400 mg extract/kg orally administered before PM | ↓ iNOS and COX-2, IL-1β, IL-6, TNF-α, pERK 1/2, p-p38-MAPK and p-JNK; ↑ HO-1 and Nrf-2 | [99] |
| Sargassum sagamianum | ||||
| EtOH 80% | Ear edema in ICR mice | 0–100 µg/mL of extract + 1 μg/mL LPS | ↓ swelling | [92] |
| Sargassum fulvellum | ||||
| EtOH 95% → fract H2O → EtOAc → n-But | UVB-induced BALB/c mice | 3 and 10 μg topically applied in the back 30 min prior to irradiation | EtOAc fraction ↓ NO●, PGE2, TNF-α, COX-2 and iNOS | [94] |
| Myagropsis myagroides | ||||
| EtOH 96% | PMA-induced ear edema in mice | 90 µg topically administered 1 h prior to PMA injection | ↓ swelling in 67% | [100] |
| Ecklonia cava | ||||
| EtOH 70% | Periodontitis-induced Sprague-Dawley rats | 100–400 µg extract/kg/day for 8 weeks | ↓ gene expression IL-1β, MMP-2 and -9 and RANKL/OPG ratio | [83] |
| EtOH 30% → EtOH 95% | LPS-induced septic shock in C57BL/6 mice | 10–100 mg/kg/d orally administered over 7 days before i.p. of LPS (25 mg/kg) | ↓ NO●, PGE2, iNOS, COX-2, IL-6, TNF-α, HMGB-1, and p-NIK, p-TAK1, p-IKK, p-IκB α, NF-κB-p50 and -p65; ↑ HO-1 and Nrf-2 | [101] |
| Ascophyllum nodosum | ||||
| EtOH 60% → fract → food-grade capsule (101·89 mg TPC) | LPS-induced Human blood ex vivo | Oral administration of a single capsule prior to blood collection | ↓ IL-6 | [102] |
| Isolated compounds | ||||
| 6,6′-Bieckol from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (15 nmol/µL) with each sensitizer | ↓ swelling in 62%, 36% and 59%, respectively | [103] |
| AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 42%, 34% and 18%, respectively | [104] | |
| 6,6′-bieckol from Myagropsis myagroides | PMA-induced ear edema in mice | 30 µg topically administered, 1 h prior to PMA injection | ↓ swelling in 64% | [100] |
| 6,8′-bieckol from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (15 nmol/µL) with each sensitizer | ↓ swelling in 56%, 66% and 78%, respectively | [103] |
| AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 40%, 49% and 78%, respectively | [104] | |
| 8,8′-Bieckol from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (0.01–0.1 mg/mL) with each sensitizer | ↓ swelling up to 80%, 49% and 71%, respectively | [105] |
| AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 21%, 32% and 32%, respectively | [104] | |
| Dieckol from Eisenia bicyclis | Acetic acid-induced ICR mice | 20 µM, 6 h prior to i.p. 0.7% acetic acid | ↓ CAMs and adhesion and migration of leucocytes | [79] |
| Dieckol from Ecklonia cava | γ-radiation induced hepatocytes from Wistar rats | 5–20 µM 1 h prior to irradiation (8 Gy/min) | ↓ NF-kB and COX-2 | [65] |
| CoCl2-induced hepatocytes from ICR mice | 100 µM 30 min prior to CoCl2 stimulation (500 µM) | ↓ COX-2 and p-p38-MAPK | [106] | |
| High glucose-induced zebrafish | 20 µM 16 h prior to glucose | ↓ NO●, iNOS and COX-2 | [35] | |
| UVB-irradiated zebrafish embryos | 50 µM 1 h prior to irradiation (50 mJ/cm2) | ↓ NO● | [107] | |
| Eckol comercial | CCl4-injured livers of Kunming mice | 0.5 and 1.0 mg compound/kg/day orally administered over 7 days before i.p. 0.5% CCl4 | ↓ IL-1β, IL-6 and TNF-α, ↑ IL-10 | [67] |
| Eckol from Ecklonia cava | UVB-irradiated zebrafish embryos | 50 µM 1 h prior to irradiation (50 mJ/cm2) | ↓ NO● | [107] |
| Eckol from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (0.01–0.1 mg/mL) with each sensitizer | ↓ swelling up to 49%, 38% and 65%, respectively | [105] |
| AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 13%, 40% and 19%, respectively | [104] | |
| Eckol from Eisenia bicyclis | Acetic acid-induced ICR mice | 20 µM, 6 h prior to i.p. 0.7% acetic acid | ↓ CAMs and adhesion and migration of leucocytes | [79] |
| Eckstolonol from Ecklonia cava | UVB-irradiated zebrafish embryos | 50 µM 1 h prior to irradiation (50 mJ/cm2) | ↓ NO● | [107] |
| Phlorofucofuroeckol-A from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (0.01–0.1 mg/mL) with each sensitizer | ↓ swelling up to 68%, 44% and 77%, respectively | [105] |
| Pholorofucofuroeckol-A from Eisenia bicyclis | AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 31%, 32% and 23%, respectively | [104] |
| Phlorofucofuroeckol-B from Eisenia arborea | AA, TPA, OXA-induced ear edema in ICR mice | Topical application (0.01–0.1 mg/mL) with each sensitizer | ↓ swelling up to 78%, 56% and 72%, respectively | [105] |
| AA, TPA, OXA-induced ear edema | Oral gavage of 75 nmol, 18 h prior to each sensitizer | ↓ swelling 42%, 38% and 41%, respectively | [104] | |
| Phloroglucinol commercial | Acetic acid-induced ICR mice | 20 µM, 6 h prior to i.p. 0.7% acetic acid | ↓ CAMs and adhesion and migration of leucocytes | [79] |
| Phloroglucinol from Ecklonia cava | UVB-irradiated zebrafish embryos | 50 µM 1 h prior to irradiation (50 mJ/cm2) | ↓ NO● | [107] |
| Triphlorethol A from Ecklonia cava | UVB-irradiated zebrafish embryos | 50 µM, 1 h prior to irradiation (50 mJ/cm2) | ↓ NO● | [107] |
| Extract/Compound | Model | Conditions | Effect | Ref. |
|---|---|---|---|---|
| Saccharina japonica | ||||
| MAE with EtOH 55% | HepG2 cells | Incubation with 0.2–2 mg/mL for 24 h | ↓ cell prolif. | [115] |
| Ascophyllum nodosum | ||||
| ACN:0.2% CH2O2 (1:1) → purification in SPE columns | Caco-2 cells | Incubation with 0–50 µg/mL for 72 h | ↓ cell prolif. (IC50 = 33 μg/mL) | [12] |
| Alaria esculenta | ||||
| ACN:0.2% CH2O2 (1:1) → purification in SPE columns | Caco-2 cells | Incubation with 0–50 µg/mL fot 72 h | ↓ cell prolif. (IC50 = 7 μg/mL) | [12] |
| Cystoria sedoides | ||||
| MAE with EtOH 50% | MCF-7 cells | Incubation with 10–200 µg/mL for 24 h | ↓ cell prolif. | [116] |
| Cystoseira spp. | ||||
| H2O extract | Tumor cells: A549, HCT15 and MCF-7 cells Normal cells: MDCK cells and rat fibroblasts | Incubation with 25–250 µg/mL for 48 h | Tumor cells: ↓ cell prolif. (IC50 = 17.9–90.3 μg/mL) Normal cells: ↓ toxicity than cisplatin | [113] |
| Fucus evanescens | ||||
| EtOH 60% | DLD-1 and HT-29 cells | Incubation with 50 µg/mL for 30 d | ↓ cell prolif. by 67% and 63%, respectively | [117] |
| Laminaria cichorioides | ||||
| EtOH 60% | DLD-1 and HT-29 cells | Incubation with 50 µg/mL for 30 d | ↓ cell prolif. by 64% and 56%, respectively (July specimen) ↓ cell prolif. by 50% and 52%, respectively (September specimen) | [117] |
| Costaria costata | ||||
| EtOH 60% | DLD-1 and HT-29 cells | Incubation with 50 µg/mL for 30 d | ↓ cell prolif. by 38% and 31%, respectively (May’s specimen) ↓ cell prolif. by 50% and 44%, respectively (July specimen) | [117] |
| Macrocystis integrifolia | ||||
| MeOH → Fract. with n-hex, H2O:EtOAc (2:3) and 1-But → subfraction of 1-But | HeLa cells | Incubation with 0.5–5 mg/mL for 72 h | ↓ cell prolif. (EC50 = 4.11 mg/mL) | [118] |
| Nereocystis leutkeana | ||||
| MeOH → Fract. with n-hex → H2O + EtOAc (2:3) → 1-But | HeLa cells | Incubation with 0.5–5 mg/mL for 72 h | ↓ cell prolif. (EC50 = 4.10 mg/mL) | [118] |
| Laminaria setchellii | ||||
| MeOH → Fract. with n-hex → H2O + EtOAc (2:3) → 1-But | HeLa cells | Incubation with 0.5–5 mg/mL for 72 h | ↓ cell prolif. (EC50 = 4.53 mg/mL) | [118] |
| Sargassum muticum | ||||
| PLE with EtOH 95% → Fract. with DCM → Ac → EtOH → EtOAC | HT-29 cells | Incubation with 12.5–100 µg/mL for 24, 48 and 72 h | ↓ cell prolif. by 50% after 24 h with 50 μg/mL of extract (Norway specimen) | [119] |
| Fucus vesiculosus | ||||
| Ac 99.5% → purification by HPLC | PancTu1, Panc89, Panc1 and Colo357 cells | Incubation with 0.01–100 µg/mL for 24 h | ↓ cell prolif., ↑ cell cycle inhibitors (EC50 = 17.35, 17.5, 19.23 and 28.9 μg/mL, for each cell line, respectively) | [120] |
| Ecklonia cava | ||||
| MeOH 70% → Fract. n-hex → DCM → EtOAc | A549 cells | Incubation with 0–200 µg/mL for 24 or 48 h | ↓ exp. of MMP-2 | [121] |
| EtOH 95% | HDFs and HT1080 cells | Incubation with 10–1000 µg/mL for 3 d | ↓ exp. of MMP-2 and -9 | [122] |
| EtOH extract | A2780 and SKOV3 cells | Incubation with serial dilutions for 24 h | ↓ cell prolif. (IC50 = 84.3 and 99.6 μg/mL, respectively) | [114] |
| Ishige okamurae | ||||
| EtOH → Fract. H2O → n-But → MeOH 85% → n-hex | HT1080 cells | Incubation with 5 or 50 µg/mL for 24 h | ↓ exp. of MMP-2 and -9; ↑ exp. TIMP-1 and -2 | [123] |
| Isolated compounds | ||||
| 6,6’-Bieckol from Ecklonia cava | HT1080 cells | Incubation with 5–100 µg/mL for 48 h | ↓ exp. of MMP-2 and -9 and exp. of NF-κB p50 and p65 | [124] |
| Dieckol commercial | In vitro: SKOV3 cells In vivo: SKOV3-xenograft in BALB/c mice | In vitro: Incubation with 80–120 µM for 24 h In vivo: oral administration of 50 and 100 mg/mL 3 d a week for 28 days | In vitro: ↑ exp. of casp-3, -8 and -9; ↓ exp. of the XIAP, FLIP and Bcl-2 In vivo: ↓ cell prolif. and lower toxicity than cisplatin | [114] |
| A549 cells | Incubation with 25–50 µg/mL for 24 h | ↑ exp. of casp-3, -8 and -9, of tumor suppressor protein E-cadherin | [125] | |
| Dieckol from Ecklonia cava | NDEA-induced Wistar rats | Oral administration of 40 mg/kg/d for 15 weeks | ↓ exp. of Bcl-2, VEGF, MMP-2 and -9, ↑ exp. of Bax, cytochrome c and casp-3 | [126] |
| MCF-7 cells | Incubation with 1–100 µM for 48 h | ↓ cell migration, exp. of MMP-9 and VEGF, ↑ exp. of TIMP-1 and -2 | [127] | |
| TPA-induced SK-Hep1 cells | Incubation with 1–50 µM for 1.5 h | ↓ exp. of MMP-9 and exp. of p-ERK 1/2, p-MEK 1/2 and p-JNK 1/2 | [128] | |
| LPS-induced MDA-MB-231 cells | Incubation with 50 µM for 48 h | ↓ cell invasion, exp. of TLR-4, NF-κB and MMP-2 and -9 | [129] | |
| Dieckol from Ecklonia stolonifera | Hep3B cells | Incubation with 70–110 µM for 24 h | ↑ exp. of casp -3, -7, -8 and -9, Bid and Bim | [130] |
| Eckol commercial | S180-xenograft in Kumming mice | Oral administration of 0.25–1 mg/kg/d for 7 d before and 10 d after xenograft | ↑ exp. of casp-3 and -9, ↓ exp. of Bcl-2, Bax, EGFR and p-EGFR | [131] |
| Eckstolonol from Ecklonia cava | MCF-7 cells | Incubation with 5–110 µM for 48 h | ↓ exp. of Bcl-2, ↑ exp. of casp-3 and -9, Bax and tumor suppressor p53 | [132] |
| Phloroglucinol commercial | HT-29 cells | Incubation with 12.5–50 µg/mL for 24 h | ↑ exp. of casp-3 and -8, cytochrome c, Apaf-1, Bad and Bax, ↓ exp. of Bcl-2, Bcl-XL | [133] |
| HT-29 cells | Incubation with 0–50 µg/mL for 24 h | ↓ exp. of IGFR-1 | [134] | |
| Phlorofucofuroeckol A from Eisinia bicyclis | HCT116, SW480, LoVo and HT-29 cells | Incubation with 100 µM for 24 h | ↑ exp. of ATF3 | [135] |
| Phlorofucofuroeckol A from Ecklonia cava | LPS-induced MDA-MB-231 cells | Incubation with 50 µM for 48 h | ↓ cell invasion, exp. of TLR-4, NF-κB and MMP-2 and -9 | [129] |
| Product | Seaweed Species | Claims | Active Ingredients | Patent/Patent Application nr |
|---|---|---|---|---|
| PhExt with antioxidant activity | E. cava | Method for production of a phlorotannin extract with high antioxidant activity for cosmetic application using supercritical extraction | E. cava extract with 10%–15% phlorotannins | KR20080004758A [137] |
| PhExt with antioxidant activity | Bifurcaria bifurcata + A. nodosum | Method for production of a phlorotannin extract with antioxidant activity for cosmetic and food application using ultrasound assisted extraction | B. bifurcate extract containing 62.4 mg eq. of phloroglucinol/g and A. nodosum extract containing 44 mg eq. of phloroglucinol/g. | US2016074317A1 [138] |
| Antioxidant composition | E. stolonifera | Production of a composition of several phlorotannins isolated from E. stolonifera with high antioxidant activity for application in functional foods to prevent oxidative spoilage or pharmaceuticals for preventing aging | Eckstolonol, eckstolonol pentaacetate, eckol, phlorofucofuroeckol A and dieckol | KR20040095125A [139] |
| Antioxidant composition | I. okamurae | Production of an antioxidant composition containing phlorotannins isolated from I. okamurae for application in pharmaceuticals, food supplements and cosmetics aiming the prevention of ROS-related diseases | Phloroglucinol, 6,6’-bieckol or diphlorethohydroxycarmalol | KR20100039104A [140] |
| PhExt and anti-inflammatory composition | F. vesiculosus + A. nodosum | Method for production extracts with high content of phlorotannins and further production of an anti-inflammatory composition using these phlorotannin-rich extracts as active ingredient for application in medicaments for treating allergies and arthritis | Algal extract containing 11%–19% phlorotannins | FR2914190A1 [142] |
| Anti-inflammatory composition | E. stolonifera and E. cava | Production of a composition containing E. stolonifera and E. cava phlorotannin extracts or isolated phlorotannins for application in farmaceutical formulations aiming the inhibition of iNOS and COX-2 on inflammatory-related diseases | Phlorofucofuroeckol A | KR20120054577A [141] |
| Anti-MMP composition | Brown seaweeds | Production of a composition containing phlorotannins from brown seaweeds with inhibitory effects against matrix metalloproteinases to be used in form of powders, tablets, capsules, ointment compositions, solutions, gels, pastes, patches, granules for the treatment of wound, cancer metastasis, rheumatoid arthritis, inflammation, hyperparathyroidism, diabetes, corneal ulcer, osteoporosis, stomach ulcer, acne, burn, periodontal diseases, arteriosclerosis and fracture | Dieckol and bieckol | KR20080005711A [143] |
| Anti-cancer composition | E. cava | Production of a composition containing eckol isolated from E. cava capable of suppressing tumorigenesis, cancer growth, invasion, metastasis and malignance in k-Ras expressing cancers via inhibition of the PI3K-Akt signaling and Ras signaling through Ras-Raf-ERK signal transduction system | Eckol | KR20130104543A [145] |
| Anti-cancer composition | E. cava | Production of a composition containing eckol isolated from E. cava for prevention of cancer stem cell growth via suppression of the Ras signaling through the Ras-Raf-ERK signal transduction system | Eckol | KR20120053265A [145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catarino, M.D.; Amarante, S.J.; Mateus, N.; Silva, A.M.S.; Cardoso, S.M. Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network. Foods 2021, 10, 1478. https://doi.org/10.3390/foods10071478
Catarino MD, Amarante SJ, Mateus N, Silva AMS, Cardoso SM. Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network. Foods. 2021; 10(7):1478. https://doi.org/10.3390/foods10071478
Chicago/Turabian StyleCatarino, Marcelo D., Sónia J. Amarante, Nuno Mateus, Artur M. S. Silva, and Susana M. Cardoso. 2021. "Brown Algae Phlorotannins: A Marine Alternative to Break the Oxidative Stress, Inflammation and Cancer Network" Foods 10, no. 7: 1478. https://doi.org/10.3390/foods10071478