
Plant Antimicrobials for Food Quality and Safety: Recent Views and Future Challenges
by
 , , and
, , and
Loris Pinto
1,* ,
,
Melvin R. Tapia-Rodríguez
2,
Federico Baruzzi
1 and
Jesús Fernando Ayala-Zavala
3 1
Institute of Sciences of Food Production, National Research Council of Italy, Via G. Amendola 122/O, 70126 Bari, Italy
2
Departamento de Biotecnología y Ciencias Alimentarias, Instituto Tecnológico de Sonora, 5 de Febrero 818 sur, Col. Centro, Ciudad Obregón, Obregón 85000, Sonora, Mexico
3
Centro de Investigación en Alimentación y Desarrollo, A.C, Carretera Gustavo Enrique Astiazarán Rosas 46, Hermosillo 83304, Sonora, Mexico
*
Author to whom correspondence should be addressed.
Foods 2023, 12(12), 2315; https://doi.org/10.3390/foods12122315
Submission received: 8 May 2023
/
Revised: 3 June 2023
/
Accepted: 5 June 2023
/
Published: 8 June 2023
(This article belongs to the Special Issue Plant Extracts Used to Control Microbial Growth: Efficacy, Stability and Safety Issues for Food Applications)
Abstract
:The increasing demand for natural, safe, and sustainable food preservation methods drove research towards the use of plant antimicrobials as an alternative to synthetic preservatives. This review article comprehensively discussed the potential applications of plant extracts, essential oils, and their compounds as antimicrobial agents in the food industry. The antimicrobial properties of several plant-derived substances against foodborne pathogens and spoilage microorganisms, along with their modes of action, factors affecting their efficacy, and potential negative sensory impacts, were presented. The review highlighted the synergistic or additive effects displayed by combinations of plant antimicrobials, as well as the successful integration of plant extracts with food technologies ensuring an improved hurdle effect, which can enhance food safety and shelf life. The review likewise emphasized the need for further research in fields such as mode of action, optimized formulations, sensory properties, safety assessment, regulatory aspects, eco-friendly production methods, and consumer education. By addressing these gaps, plant antimicrobials can pave the way for more effective, safe, and sustainable food preservation strategies in the future.
1. Introduction
The use of natural antimicrobials in the food industry is gaining attention due to the consumers’ demand for environmentally friendly production systems and products with clean labels, promoting the use of natural antimicrobial preservatives rather than synthetic ones [1,2]. Indeed, synthetic food preservatives such as nitrates, benzoates, sulfites, sorbates, and formaldehyde are known for allergic or carcinogenic effects [3]. Microbial food spoilage is responsible for about 25% of food losses [4]. According to the Food and Agriculture Organization (FAO), wasted food costs approximately 680$ billion in industrialized countries and 310$ billion in developing countries, with a high emission footprint for meat products [5]. Moreover, the growing consumption of fresh, minimally processed, and ready-to-eat foods increases the chance of microbial contamination by spoilage and pathogenic microorganisms [1]. Therefore, natural antimicrobials should be promoted to extend the shelf life of perishable foods, and to ensure the product’s microbial food safety.
Plant antimicrobials represent the main group of natural preservatives, including secondary metabolites targeting microbial cells. Different parts of plants, such as seeds, fruit, peels, leaves, and roots are rich in plant antimicrobials such as phenolic compounds (e.g., simple phenols, phenolic acids, anthocyanins, flavonoids, quinones), tannins, essential oils and terpenoids, glucosinolates derivatives, alkaloids, and thiols [6,7]. Most of the plant extracts are generally recognized as safe (GRAS) and were granted the qualified presumption of safety (QPS) status in the USA and EU, respectively [8]. Plant extracts, such as moso bamboo (Takeguard™) with benzoquinone derivatives and tannin, or an antifungal blend (Biovia™ YM10) with green tea (Camellia sinensis L.) extract and mustard (Brassica nigra W.D.J. Koch) essential oil, are commercially available as alternatives to chemical preservatives [1]. Moreover, the European Food Safety Authority (EFSA) authorized rosemary (Rosmarinus officinalis L.) extract, endowed with antimicrobial activity, as a food additive (E 392) [9,10].
Plant antimicrobials were proposed to control the growth of microbial spoilage populations and foodborne pathogens. As regards the control of spoilage microorganisms, several applications were described in animal-based foods. In fish products, grape (Vitis vinifera L.) seed extract, tea polyphenols, thyme essential oil, and rosemary extract delayed the growth of lactic acid bacteria, Enterobacteriaceae, hydrogen sulfide-producing bacteria (HSPB), and psychrotrophic bacteria, well known to produce off-flavours [11]. In meat products, tannic acid or catechin showed good antimicrobial activity in camel sausages, whereas ethanolic extracts of rosemary and clove (Syzygium aromaticum L.) reduced spoilage bacterial counts in raw chicken meat. In beef sausages, the use of Ziziphus leaf extracts, rich in vanillic and ellagic acids, inhibited the growth of spoilage bacteria during cold storage [12]. Among essential oils, the application of Ziziphora clinopodioides Lam., rich in carvacrol, thymol, p-cymene, and γ-terpinene, showed the best antimicrobial activity against spoilage bacteria in beef patties [13]. In plant-based foods, as reviewed by Patrignani et al. [14], citral, hexanal, and 2-(E)-hexenal showed antimicrobial activity against yeasts responsible for spoilage of fresh-cut fruits, soft drinks, and fruit-based salads, whereas citral-based films or the application of oregano (Origanum vulgare L.) and thyme (Thymus vulgaris L.) oil during the washing step reduced spoilage bacterial populations on salad. The antimicrobial action of plant extracts against foodborne pathogens is well documented [1,2,3,7,15,16,17]. In particular, phenolic extracts and essential oils showed remarkable antibacterial action against Gram-positive and Gram-negative bacteria, including spore-forming bacteria. In addition to the effect against viable cells, plant antimicrobials inhibited the production of microbial toxins [18,19] and biofilm formation [20,21,22].
Despite the antimicrobial action of plant antimicrobials, their use in the food industry is hampered by chemical instability, limited dispersibility in food matrices, limited availability of ready-to-use commercial formulations, or unacceptable flavour profiles [6]. For these reasons, several stabilization techniques, such as nano-emulsions, encapsulation, and inclusion in active packaging, were proposed [6,23,24]. Moreover, these stabilization techniques ensure, in some cases, better antimicrobial activity of the bioactive compounds, and a controlled release during food storage.
However, some challenges remain, including potential negative sensory impacts, variations in antimicrobial effectiveness, and concerns about the possible development of microbial resistance. To address these issues, researchers explored synergistic combinations of plant antimicrobials and the application of hurdle technologies, which involve the simultaneous or successive use of multiple preservation techniques. Although plant extracts showed considerable potential in food preservation, limited information is available concerning their safety. In some instances, these extracts can be contaminated with various hazardous substances, such as heavy metals [25], mycotoxins [26], or crop protection residues [27]. The levels of contamination in plant extracts are affected by several factors, including the cultivation practices employed, the geographical location of the cultivation site, and the application of crop protection products. Further research is needed to establish proper guidelines and regulatory frameworks that can help minimize the risks associated with contaminants in plant extracts, ultimately ensuring the safe application of these natural preservatives in the food industry. Further research is also necessary to understand the modes of action of plant antimicrobials alone or in combination to optimize their formulation and the delivery of bioactive compounds. Addressing these gaps will help the acceptance of plant extracts as food preservatives and their use in different food industries.
This review aims to summarize the applications of plant antimicrobials in the food sector. After that, an overview of different classes of plant antimicrobials, antimicrobial activity against spoilage, and pathogenic microorganisms in different foods is described. Then, the stabilization techniques of plant extracts are presented followed by their use in different food matrices. This review also discusses the additive and synergistic effects of various combinations of plant antimicrobials, as well as the integration of plant extracts into different hurdle technologies, including mild or non-thermal treatments, to enhance food preservation. Finally, safety aspects and regulation related to the use of plant extracts are introduced. By providing a comprehensive overview of the current knowledge, this review aims to contribute to the ongoing development and optimization of food preservation techniques based on plant antimicrobials.
2. Classification and Antimicrobial Activity of Plant Antimicrobials
A great diversity of structures among plant secondary metabolites (PSMs) occurs in nature (e.g., more than 12,000 known alkaloids, more than 10,000 phenolic compounds, and over 25,000 different terpenoids) [1]. From a structural point of view, plant antimicrobials can be divided in two classes: PSMs with one or several nitrogen atoms into their structures, such as alkaloids, glucosinolates, and PSMs without nitrogen, such as terpenoids and phenolic substances. Alkaloids, glucosinolates, and phenolic substances are water-soluble compounds, whereas terpenoids are lipophilic PSMs [28]. The following sections summarize the different classes of plant antimicrobials and their antimicrobial action against main food-related microorganisms.
2.1. Polyphenols
Polyphenols are PSMs produced by higher plants, sharing a common chemical structure characterized by at least one aromatic ring with one or more hydroxyl groups [29]. Polyphenols can be classified as flavonoids and nonflavonoids. The latter includes the phenolic acids (e.g., derivatives of benzoic acid and cinnamic acid), stilbenes (e.g., resveratrol), tannins (e.g., proanthocyanidins, gallotannins, and ellagitannins), and lignins (e.g., secoisolariciresinol). Flavonoids can be divided into six subclasses: flavonols, flavones, flavanones, flavanols, anthocyanins, and isoflavones [30].
2.1.1. Phenolic Acids
Phenolic acids are divided into hydroxybenzoic acids (e.g., vanillic, gallic, salicylic, syringic, and protocatechuic acid) and hydroxycinnamic acids (ferulic, rosmarinic, p-coumaric, chlorogenic, cinnamic, and caffeic acid) [31]. The main phenolic acids showing antimicrobial action are gallic acid, ferulic acid, and p-coumaric acid [31]. A minimum inhibitory concentration (MIC) of 1000–2000 µg mL−1 was found for gallic acid and ferulic acid against Escherichia coli, Staphylococcus aureus, and Listeria monocytogenes [32]. Ferulic acid and p-coumaric acid showed MIC values of 500–1000 µg mL−1 against Salmonella enteritidis [33]. Li et al. [34] recently found that p-coumaric acid controlled the contamination of Alicyclobacillus acidoterrestris in apple juice. However, the antibacterial action of phenolic acids can be enhanced considering their derivatives, as demonstrated for alkyl ferulate and gallate esters against L. monocytogenes and E. coli, respectively [35,36]. Regarding the antifungal action, ferulic acid and p-coumaric acid showed antifungal activity against Botrytis cinerea and Alternaria alternata [37,38]. As reported for the antibacterial action, ester derivatives of phenolic acids showed enhanced antifungal action compared to phenolic acids. In particular, ethyl p-coumarate showed interesting antifungal activity against Alt. alternata [39].
2.1.2. Stilbenes, Tannins, and Lignins
Other polyphenols endowed with antimicrobial activity are stilbenoids, tannins, and lignins. Stilbenes such as resveratrol showed antibacterial action against foodborne pathogens, with MIC values of 100–200 µg mL−1 for S. aureus and Enterococcus faecalis, and >200 µg mL−1 for E. coli and Sal. enterica [40]. Cai et al. [41] found that pterostilbene had higher antifungal activity against ochratoxin A (OTA)-producing Aspergillus carbonarius than piceatannol and resveratrol. As regards tannins, they are classified into hydrolysable and condensed tannins. Hydrolysable tannins such as ellagitannins showed antibacterial action against S. aureus and E. coli. In particular, increased free galloyl groups enhanced antibacterial action against S. aureus, while large molecular size positively affected the antimicrobial effect against E. coli [42]. Condensed tannins such as proanthocyanidins from persimmon [43] or chokeberry [44] showed MIC values of 0.7–5 mg mL−1 against S. aureus. Regarding the antibacterial activity of lignin, different sources and extraction processes can result in different antibacterial performances. However, the ethanol fractionation of bamboo kraft lignin enhanced the antibacterial activity compared to non-fractionated lignin, and the ethanol fraction showed a MIC value of 2 mg mL−1 against Bacillus subtilis and S. aureus [45].
2.1.3. Flavonoids
Flavonoids are the main dietary polyphenols. They show a characteristic phenyl-benzopyrone structure and can be classified into anthocyanidins, flavan-3-ols, flavones, flavanones, flavonols, and isoflavonoids [29]. Among them, flavan-3-ols, flavonols, and flavanones showed the highest antibacterial activity against foodborne pathogens [31]. In particular, flavan-3-ols such as epigallocatechin-3-gallate showed antibiofilm activity against L. monocytogenes [46], and bactericidal effect against E. coli [47]. The main flavonol endowed with antibacterial activity is resveratrol. Resveratrol showed a MIC value lower than 10 mg mL−1 against E. coli O157:H7 and Sal. enteritidis [48] and 300–600 μg mL−1 against methicillin-resistant S. aureus [49]. However, the presence of rhamnose and additional hydroxyl groups in the flavonoids myricetin-3-O-rhamnoside and quercetin-3-O-rhamnoside resulted in reduced antibacterial activity compared to quercetin [31]. As regards the antifungal activity of flavonoids, quercetin at 0.25 mg mL−1 inhibited mycelial growth of Penicillium expansum [50] and showed a MIC value of 505 μg mL−1 against Aspergillus flavus [51]. Flavanones belong to a sub-class of flavonoids. The most interesting antibacterial activity was found for sophoraflavanone G against methicillin-resistant S. aureus, with MIC values ranging from 0.5 to 8 µg mL−1 [52]. Recently, other flavonoids, such as the mono-prenylated isoflavonoids showed high antifungal activity against Zygosaccharomyces parabailii, a spoilage yeast of acidic food products, with a minimum fungicidal concentration (MFC) of 12.5 μg mL−1 [53].
2.2. Terpenes and Essential Oils
Essential oils (EOs) are complex blends of aromatic metabolites extracted from different plant parts, including leaves, bark, flowers, and roots, using solvents, distillation, or microwaves [54]. Volatile compounds represent 90–95% of EOs, including monoterpenes, sesquiterpene hydrocarbons and their oxygenated derivatives, aldehydes, alcohols, and esters. The non-volatile portion (5–10% of the whole EO) comprises hydrocarbons, fatty acids, sterols, carotenoids, waxes, cumarines, and flavonoids. The main antimicrobial compounds present in EOs can be divided into different groups: terpenes (e.g., p-cymene, limonene), terpenoids (e.g., thymol, carvacrol), and phenylpropenes (e.g., eugenol, vanillin) [30].
Rosemary EO, rich in the monoterpenes α-pinene, 1,8-cineol, and camphor, showed antibacterial action against E. coli and S. aureus [55,56]. A recent study [57] showed that the geographic origin of rosemary EOs affected their composition and antimicrobial activity. EOs extracted from Salvia officinalis L., Lavandula dentata L., and Laurus nobilis L., rich in 1,8-cineol, inhibited the growth rate of A. carbonarius and the OTA production [58].
EOs with terpenoids such as thymol and carvacrol as main compounds paid great attention due to their broad spectrum of antimicrobial activity and potential application through direct contact and vapour phase. Oregano and thyme EOs showed antibacterial activity by direct contact against drug-resistant Gram-positive pathogens such as S. aureus and Enterococcus faecium, and Gram-negative pathogens such as E. coli and Sal. thyphimurium [59,60]. These EOs showed antimicrobial activity in vapour phase, with MIC values of 0.16–4.00 μg mL−1 of air against E. coli and Penicillium expansum [61]. Moreover, oregano and thyme EOs vapours showed antifungal activity against different species of the genera Aspergillus, with MIC values of 15.6–62.5 μL L−1 of air [62]. Regarding p-cymene, this monoterpene has low antibacterial activity, high MIC values, and no antifungal action against Rhizopus oryzae and A. niger [63]. Similarly, in B. cinerea, P. italicum, and Alt. alternata, p-cymene showed higher MIC values than other monoterpenes such as thymol and γ-terpinene [64].
Phenylpropanoids such as eugenol and isoeugenol, both present in clove EO, showed antibacterial action against E. coli and L. monocytogenes with MIC values in the range 312.5–625 µg mL−1 [65]. Clove oil, with eugenol as the main compound, inhibited P. italicum growth on citrus fruit when applied at concentrations ranging from 0.05% to 0.8% (v/v) [66]. Other phenylpropanoids, such as vanillin, showed a bacteriostatic effect against foodborne pathogens, but MIC values were higher than that of pure compounds belonging to terpenes or terpenoids [67].
Other bioactive compounds occurring in EOs are the aldehydes citral and cinnamaldehyde, found in lemongrass (Cymbopogon citratus Stapf) EO and cinnamon (Cinnamomum verum Presl) bark EO, respectively. Free citral showed a MIC value of 0.8 mg mL−1 against B. cereus and 2 mg mL−1 against E. coli and S. aureus [68]. However, the main application of citral is its use as an antifungal agent, as demonstrated against different fungal strains [69,70,71]. As regards cinnamaldehyde, it showed higher antibacterial activity than cinnamon oil against Gram-positive bacteria [72]. Cinnamaldehyde at 150 μg mL−1 inhibited the spore production and mycelial growth of A. niger [73] and showed antifungal activity and alternariol reduction at 0.200 μL mL−1 against Alt. alternata [74].
2.3. Glucosinolate Derivatives
Glucosinolates are the main bioactive compounds of Brassica plants. The breakdown of glucosinolates releases nitriles, thiocyanates, and isothiocyanates. In particular, isothiocyanates, largely occurring in cruciferous vegetables, are the most reactive compounds endowed with antimicrobial activity. Allyl-, benzyl-, and 4-methylsulfinylbutyl isothiocyanates are the main compounds with antimicrobial activity against bacterial pathogens and fungi [7]. Allyl-isothiocyanate at the concentration of 1 µL L−1 reduced of 4 log cfu g−1 the Sal. thyphimurium load on lettuce [75], whereas at 0.1% v/w inhibited L. monocytogenes growth in chickpea puree stored for 10 days at 4 °C [76]. Allyl-isothiocyanate showed antifungal activity against A. flavus in maize and P. verrucosum in barley, reducing the aflatoxin B1 and ochratoxin A accumulation, respectively [77,78]. Benzyl-isothiocyanate showed MIC values ranging from 60 to 160 µM against enterotoxigenic E. coli [79], and 120 µM against L. monocytogenes [80]. Benzyl-isothiocyanate at 25 µg mL−1 inhibited the growth of A. carbonarius and A. ochraceus, whereas A. niger was more resistant to both allyl- and benzyl-isothiocyanates than other aspergilli [81]. Other bioactive isothiocyanates are sulforaphane (4-methylsulfinylbutyl isothiocyanate) and phenethyl isothiocyanate. Both compounds showed MIC values of 40–88 mg mL−1 against S. aureus and E. coli [82]. However, their use for applications in the food sector is limited compared to allyl- and benzyl-isothiocyanates. Other isothiocyanates demonstrated an interesting antifungal activity. In particular, the volatile compound 2-phenylethyl isothiocyanate showed a MIC value of 1.2 mM against Alt. alternata, and reduced the development of the black spot rot on pear [83], whereas 2-(4-methoxyphenyl)ethyl isothiocyanate showed an EC50 value of 4.2 μg mL−1 against A. niger, and inhibited the spore germination by 95% [84].
2.4. Alkaloids and Thiols
Several plant extracts include alkaloids and thiols as antimicrobial compounds. Alkaloids are PSMs classified based on their chemical structure and natural origin. Although more than 18.000 alkaloids are known, mainly represented by plant alkaloids [85], their use in the food sector is limited due to their well-known toxic and neuroactive effects. Recently, berberine, an isoquinoline alkaloid found in roots and stem-bark of Berberis plants, was the most studied alkaloid exploited for its antimicrobial activity against food-related microorganisms and was proposed as a food preservative [86,87,88]. In particular, Berberis vulgaris root and leaf extracts, rich in berberine, showed a MIC value of 150 µg mL−1 against E. coli and S. aureus, and 60–100 µg mL−1 against different Aspergillus species [86]. Berberine at 1.6 mg mL−1 inhibited mycelial growth and spore germination of P. italicum [88].
As regards thiols, the main antimicrobial compounds are allicin and its derivatives [29]. Allicin is a sulphur compound occurring in garlic, effective against spoilage yeasts, Gram-positive and Gram-negative foodborne pathogens, with MIC values lower than 30 µg mL−1 [89]. The main oxidation derivatives of allicin are diallyl disulphide and diallyl trisulfide. Diallyl disulphide showed antibacterial action against B. cereus and a MIC value of 120 µg mL−1 [90], whereas diallyl trisulfide treatment reduced, by 1.5 log cfu g−1, the Campylobacter jejuni count on chicken [91].
2.5. Modes of Action
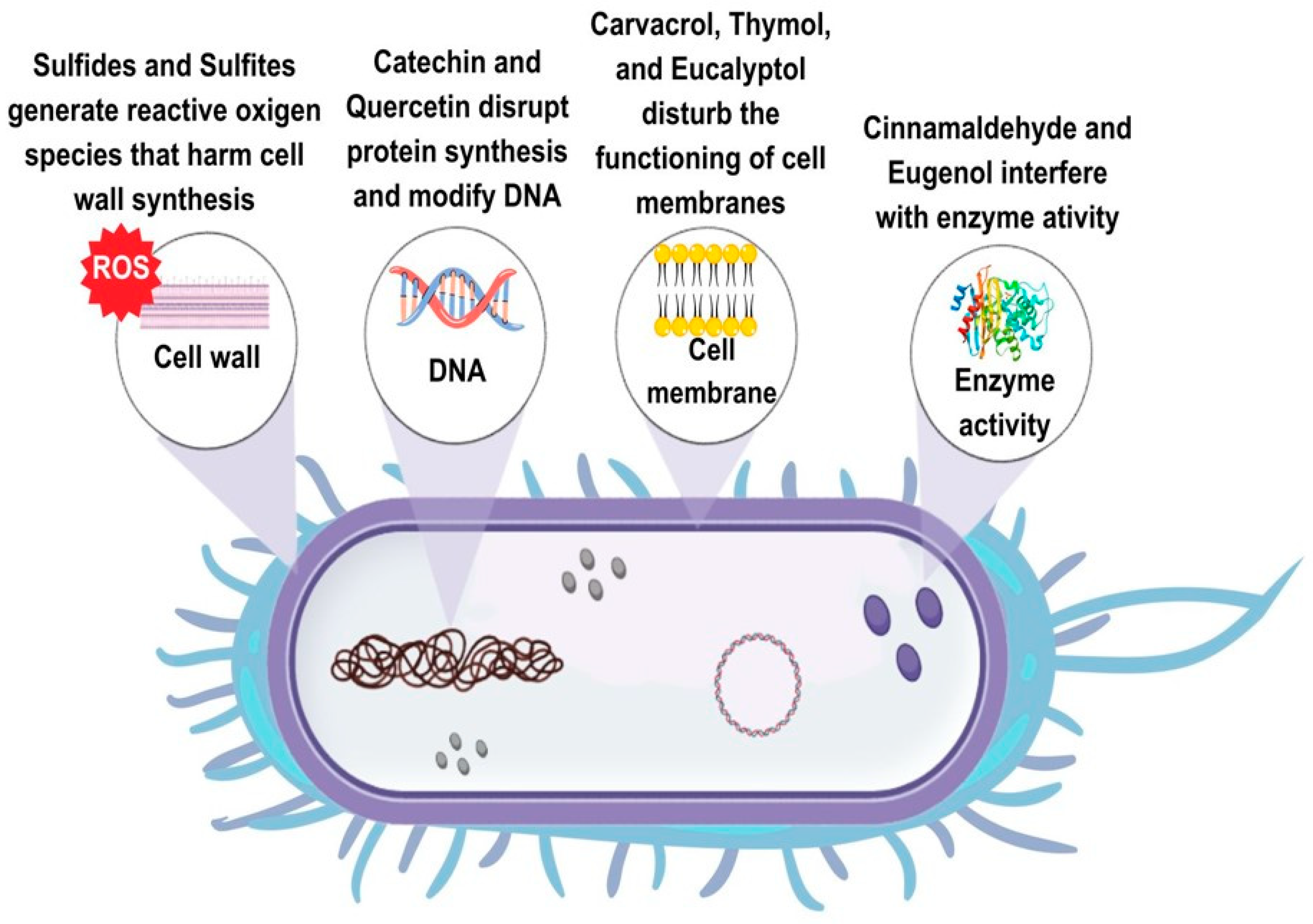
PSMs described in the previous sections have multiple mechanisms of antimicrobial action (Figure 1). In particular, different cell targets are affected by exposure to polyphenolic substances, essential oil compounds, isothiocyanates, alkaloids, and thiols. As regards polyphenols, the three main mechanisms of action are the modification of the membrane permeability, the intracellular enzyme inactivation, and the modification of fungal morphology. Additional mechanisms of antimicrobial action of polyphenols are the modification of intracellular pH, the interference with the ATP-generating system, and the inhibition of DNA synthesis [1].
Different polyphenolic classes have specific mechanisms of action. Phenolic acids mainly interact with the cell membrane intercalating the phospholipid layer, or crossing the membrane, decreasing the intracellular pH, and/or interacting with cellular constituents [92]. The antibacterial action of phenolic acids against L. monocytogenes depends on their dissociated/undissociated form. In particular, chlorogenic acid and gallic acid reduced extracellular pH, caffeic acid, p-hydroxybenzoic acid, protocatechuic acid, and vanillic acid were active in their undissociated form, and p-coumaric acid and ferulic acid showed antibacterial action in both dissociated and undissociated form [92]. In Sal. enteritidis, chlorogenic acid treatment damaged intracellular and outer membranes and inactivated key enzymes of the tricarboxylic acid cycle (TCA) [93]. Phenolic acid esters showed multiple mechanisms of antibacterial action, such as the damage of bacterial membranes, changes in the conformation of protein membranes, formation of complexes with bacterial DNA, and oxidative damage [35,36]. As regards the antifungal mechanism of phenolic acids, it is well known that these compounds produce oxidative stress and disorganization of the wall or membrane of the hyphae [94], but, as in the case of B. cinerea, they can also affect the ATP synthesis and cellular metabolism acting as an uncoupler of oxidative phosphorylation [95]. Resveratrol inhibits ATP synthesis, hydrolysis, and cell division in E. coli [40]. Pterostilbene treatment induces incomplete sporangia, membrane rupture, and downregulation of the biosynthetic genes of the OTA production in A. carbonarius [41]. The disruption of cell membranes and functions is the primary mode of antibacterial action of tannins. However, the inhibition of microbial enzymes, the deprivation of the nutrients required for the microbial growth, and the inhibition of oxidative phosphorylation were also suggested [42]. In P. digitatum, tannins disrupted the cell wall and caused the leakage of intracellular content [96]. The antibacterial modes of action of lignin are the damage of the cell membrane through its phenolic compounds, the decrease in intracellular pH, and the increase in osmotic pressure [97]. Flavonoids have multiple modes of antimicrobial action. Quercetin inhibited DNA gyrase, increased membrane permeability, and prevented ATP synthesis in E. coli [29], whereas in S. aureus, it inhibited key enzymes necessary for the protein synthesis [31]. Flavonols such as quercetin, rutin, morin, rhamnetin, and flavones such as acacetin and apigenin have membrane-disrupting activity. Conversely, flavanones naringenin and sophoraflavanone G reduce the fluidity in regions of both inner and outer cellular membranes [18]. Catechins such as epigallocatechin gallate, at high concentration, generated reactive oxygen species (ROS), causing membrane damage [18]. As regards the antifungal mechanism of flavonoids, a recent study showed that quercetin downregulated genes involved in the conidial and mycelial development, while reducing the production of aflatoxin probably by lowering levels of ROS [51]. Flavonoids from the medicinal–edible plant Sedum aizoon L. damaged the cell membrane and the cell wall, and interfered with the mitochondrial respiratory metabolism, the protein biosynthesis, and the amino acid metabolism in P. italicum [98].
As regards the antibacterial mechanism of action of terpenoids and essential oils, these compounds can disrupt cell walls and cytoplasmic membranes, increasing their permeability. Essential oils can also solidify the cytoplasm, damage lipids and proteins in the cell, and inhibit bacterial enzymes [29]. Specifically, terpenoids such as carvacrol disrupted the cell membranes and inhibited the respiratory activity in L. monocytogenes [99], while it increased the cell permeability and reduced the ATP levels in E. coli [100]. In Sal. enteritidis, the antibacterial action of oregano essential oil was mainly attributed to thymol rather than its isomer carvacrol, with changes in the protein regulation and the DNA synthesis [101]. The antibacterial action of terpenes such as limonene was associated with increased cell permeability, inhibition of the ATP synthesis, dysfunction of the respiratory chain complex, and inhibition of the transcription of nucleic acids [102,103,104]. Phenylpropenes such as eugenol altered the membrane permeability in E. coli [105], whereas they increased reactive oxygen species, depolarized the membrane potential, and decreased the ATP content in Shigella flexneri [106].
The antifungal mechanisms of action of essential oil compounds such as thymol and carvacrol are related to changes in the morphology of hyphae, the increase in membrane permeability, and the reduction in total lipids and ergosterol content [107,108]. As regards p-cymene, the antibacterial mode of action is related to the expansion of the cytoplasmic membrane and a moderate generation of ROS [63]. Other essential oil compounds, such as citral and geraniol, showed distinctively antifungal mechanisms of action. In particular, citral downregulated the sporulation- and growth-related genes in A. flavus and A. ochraceus, whereas geraniol determined intracellular ROS accumulation in A. flavus and increased cell membrane permeability in A. ochraceus [70]. Oxidative stress was partially responsible for the antifungal action of cinnamaldehyde against A. niger, causing cell damage and increasing membrane permeability [73]. Citral, limonene, and eugenol damaged the cell membranes and destroyed the yeast proteins in Zygosaccharomyces rouxii [109]. In A. carbonarius, eugenol determined the leakage of cytoplasmic contents, increased the lipid peroxidation, decreased the ergosterol content, increased the membrane permeability, and induced oxidative stress [110].
Luciano and Holley [111] demonstrated that allyl isothiocyanate inhibited thioredoxin reductase and acetate kinase in E. coli O157:H7. The bacteriostatic/fungistatic effects of benzyl isothiocyanate against E. coli, B. subtilis, Sal. enterica, S. aureus, A. niger, and P. citrinum were associated with interferences with the ATP production, enzymes and coenzymes of the energy metabolism [112]. Conversely, in B. cinerea, benzyl isothiocyanate disrupted the plasma membrane integrity and induced ROS accumulation in the spores, inhibiting their germination [113]. Other glucosinolate derivatives, such as sulforaphane and phenethyl isothiocyanate, are effective against different pathogenic bacteria by inhibiting the synthesis of nucleic acids or disrupting the membrane integrity depending on bacterial species [83].
The alkaloid berberine binds to the FtsZ protein, causing the inhibition of bacterial cell division [114]. In Sal. typhimurium, it reduces the number of type I fimbriae and prevents biofilm formation [115]. In fungi, berberine damages the plasma membrane integrity and reduces the contents of soluble proteins and reducing sugars. In addition, a high H2O2 content was found in berberine-treated P. italicum mycelia [88]. Thiols such as allicin display antimicrobial action due to the rapid reaction of thiosulfinates with thiol groups of key enzymes [89].
3. Plant Antimicrobials for Food Quality and Safety
In the past, the use of plant material during traditional food processing was defined empirically to improve the sensory characteristics of the food and the food safety and quality levels. It should be considered that several spices, obtained from different plant species, often include antimicrobial molecules and are usually supplemented to foods as flavouring agents. For this reason, the use of plant compounds as food preservatives is close to traditional recipes and, therefore, highly accepted by consumers. This section presents the direct application of plant antimicrobials in different foods, highlighting the antimicrobial action against spoilage and pathogenic microorganisms.
3.1. Plant Antimicrobials as Food Preservatives
Plant antimicrobials were exploited as preservatives in several foods to control the microbial growth of food spoilage microorganisms or foodborne pathogens [116]. This section summarizes recent published results, focusing on the direct application of plant extracts or their bioactive compounds as preservatives in food products (Table 1).
3.1.1. Applications in Plant Foods
This section presents the applications of whole plant extracts or their antimicrobial compounds against spoilage microorganisms of fresh fruit and vegetables, ready-to-eat vegetables, and fruit juices.
With regard to the application of plant antimicrobials on fresh fruits and vegetables, pomegranate (Punica granatum L.) peel extract (PPE), rich in polyphenols such as punicalagin and ellagic acid, reduced the growth of post-harvest fungi belonging to the genera Penicillium, Botrytis, Monilinia, and Colletotrichum on various fruits including lemon, strawberry, grape, apple, grapefruit, orange, and capsicum. In addition, the PPE ethanolic or aqueous extracts can preserve foods by dipping treatments or using edible coatings [117]. In this context, the use of ethanolic pomegranate peel extract (PPE) was found to significantly (p ≤ 0.05) reduce the lesion diameter and infection rate in mandarins contaminated with P. italicum and P. digitatum [118]. In addition, other plant-based extracts were also found to be effective in controlling spoilage microorganisms. A mango kernel extract, rich in mangiferin, chlorogenic acid, and myricetin, inhibited anthracnose development caused by Colletotrichum brevisporum on mangoes [119]. A sweet orange (Citrus sinensis L.) peel extract, rich in ferulic acid, showed antifungal activity against M. fructicola and Alt. alternata in a peach-based medium [120].
Other plant antimicrobial extracts with antimicrobial activity against spoilage microorganisms on fresh fruit and vegetables are the essential oils or their main compounds. Mint (Mentha × piperita L.), basil (Ocimum basilicum L.), lavender (Lavandula angustifolia Mill.), and thyme EOs in the vapour phase were used for the post-harvest preservation of strawberry, peach, orange, and lemon [121,122,123,124]. In particular, as recently reported by Pinto et al. [123], the in-package application of red thyme oil vapours reduced the percentage of infected wounds, the mycelium development, and the production of spores by Penicillium strains on oranges during 12 days of cold storage. Dipping in cinnamon essential oil microemulsion at 0.3% v/v eradicated P. fluorescens from iceberg lettuce during 28 days of cold storage [124]. As regards other plant antimicrobials, methyl, allyl, and ethyl isothiocyanate (8–12 µL L−1) completely inhibited citrus sour-rot caused by Geotrichum citriaurantii [125], whereas berberine at 3 mg mL−1 reduced the development of P. italicum and natural decay on citrus fruit [88].
Plant antimicrobials were extensively used to control the spoilage microorganisms on ready-to-eat fruits and vegetables [126]. Dipping of fresh-cut pineapple in Centella asiatica extract, rich in quercetin and kaempferol, reduced the A. niger load during cold storage [127]. In-package application of trans-anethole in ready-to-eat organic lettuce reduced total coliforms during cold storage [128], whereas the addition of β-caryophyllene-rich pepper EOs in salad dressing decreased P. fluorescens development and spoilage activity on fresh-cut lettuce [129]. Pomegranate arils coated with savoury essential oil-loaded chitosan showed a reduction in total mesophilic bacteria and total yeasts and moulds of 1 log CFU g−1 after 18 days of storage [130]. Peppermint and tea tree (Melaleuca alternifolia Cheel) oils controlled the growth of total aerobic bacteria, yeasts, and moulds on fresh-cut green bean pods stored for 9 days at 5 °C [131].
Other applications of plant antimicrobials in plant-based foods concern fruit juices and smoothies [132]. In this context, essential oils and their compounds are the most used antimicrobials. Indeed, Mentha piperita L. EO inclusion (7.50 µL mL−1) in cashew and guava juice caused >5 log reductions in counts of the spoilage yeast Pichia anomala [133]. Thymol in concentrated apple juice showed higher antimicrobial activity than carvacrol and trans-cinnamaldehyde against Z. rouxii [134]. Lee et al. [135] found a synergism between oregano and thyme EOs, at 0.156 μL mL−1, in inhibiting Leuconostoc citreum in tomato juice. As regards the applications of plant antimicrobials in smoothies, the addition of beet (Beta vulgaris L.) leaf extract (30% w/v) in a vegetable smoothie reduced significantly (p ≤ 0.05) total mesophilic bacteria, enterobacteria, and total yeasts and moulds throughout 21 days of cold storage [136].
The use of plant antimicrobials in plant-based food products, specifically fruit juices and fresh and ready-to-eat vegetables, effectively reduces spoilage and increases the shelf life of these products. Essential oils and their bioactive compounds, such as Mentha piperita L. EO, thymol, carvacrol, trans-cinnamaldehyde, oregano and thyme EOs, demonstrated antimicrobial activity against different spoilage microorganisms. These findings suggest that plant antimicrobials have the potential to play a crucial role in preserving the quality of plant-based foods.
3.1.2. Applications in Animal-Based Foods
This section presents some applications of whole plant extracts or their antimicrobial compounds against spoilage microorganisms contaminating animal-based foods (e.g., meat, seafood, and dairy products). In this context, the addition of 200 mg kg−1 of tannic acid or catechin in camel meat decreased total mesophilic and psychrophilic bacterial counts by one order of magnitude after 9 days of refrigeration [137], as well as Nowak et al. [138] demonstrated that sour cherry (Prunus cerasus Scop.) leaf extract, rich in coumaric acid, and blackcurrant (Ribes nigrum L.) leaf extract, rich in gallic acid and quercetin derivatives, delayed the growth of Pseudomonas spp. in pork sausages, but not that of Brochothrix spp. and Enterobacteriaceae. On the contrary, Casaburi et al. [139] reduced the growth of Brochotrix spp. and Enterobacteriaceae, but not that of Pseudomonas spp., of grounded beef meat during cold storage, adding 5% of a freeze-dried myrtle (Myrtus communis L.) extract, rich in phenolic compounds. These results highlight that the effectiveness of phenolics can vary depending on the specific bacterial species, the concentration of phenolics, and other factors such as the food matrix, the presence of other preservatives, and the storage conditions.
The addition of the ethanolic extract of cranberry (Vaccinium oxycoccos L.) pomace, characterized by great amounts of anthocyanins, chlorogenic acid, and myricetin and quercetin derivatives, inhibited the growth of Brochothrix thermospacta and P. putida on pork burgers during the first days of cold storage [140]. As regards the application of essential oils or their compounds on meat products, ethanolic extracts of rosemary and clove (1% v/w) reduced Pseudomonas spp. counts in raw chicken meat during cold storage [141]. The use of Ziziphora clinopodioides essential oil (0.2% v/w), rich in carvacrol and thymol, reduced the Enterobacteriaceae and psychrotrophic bacteria loads of raw beef patties during cold storage by 2–3 log cfu g−1 [142]. Thymol or carvacrol at 0.4% w/w in marinated beef significantly reduced the mesophilic total viable count, lactic acid bacteria, Broch. thermosphacta, Pseudomonas spp., and total coliforms, extending the microbiological shelf life by three days [143]. In this context, only some terpene compounds showed a broad spectrum of activity against various bacterial species, making them effective preservatives for meat products, independently of the source of plant origin, and able to extend the shelf life of some meat products.
Likewise, the reduction in fish spoilage bacteria can be achieved using plant antimicrobials, specifically polyphenolic extracts and essential oils [11]. The use of ethanolic Noni (Morinda citrifolia L.) leaf extract, rich in rutin and kaempferol derivatives, was shown to extend the shelf life of striped catfish slices and maintain the acceptable levels of total viable bacteria and psychrophilic bacteria during storage, with loads remaining below 6 log cfu g−1 [144]. Similarly, the growth of Pseudomonas spp. in Pacific white shrimps was delayed by adding ethanolic guava (Psidium gujava L.) leaf extracts, rich in phenolic compounds such as piceatannol 4′-galloylglucoside, epicatechin, epigallocatechin, procyanidin B2, ellagic acid, quercetin 3′-o-glucuronide, and quercetin 3-galactoside [145]. Grape seed extract, containing high levels of phenolic acids, catechins, and proanthocyanidins, decreased the presence of Aeromonas spp. in snakehead fillets during cold storage. This reduction limited the release of soluble peptides and biogenic amines and increased the shelf life of snakehead fillets by three days [146]. The application of essential oils, such as cinnamon, oregano, and thyme, as marinades was evaluated in salmon and scampi by Van Haute et al. [147]. The immersion of these products in cinnamon essential oil at 1% w/w inhibited the growth of yeasts and moulds. Similarly, cinnamon essential oil at 0.1% w/v effectively inhibited Aeromonas spp. in vacuum-packed carp and extended its shelf life by two days [148]. However, the direct application of essential oils in fish products can cause bitterness, off-flavours, and yellowing of the tissue [11]. The inclusion of essential oils in active packaging or nano-emulsions is recommended to mitigate these effects.
Building on the findings of previous studies on the application of plant antimicrobials in meat and fish products, the use of plant polyphenols, essential oils, and other plant-based compounds in milk and dairy products to control spoilage microorganisms and extend their shelf life is also of interest. For instance, the addition of olive mill wastewater in the governing liquid of “Fior di Latte” cheese (500 µg mL−1 of phenols) resulted in a four-day extension of shelf life due to the increase in the lag phase of P. fluorescens and Enterobacteriaceae [149]. A recent study by Derbassi et al. [150] evaluated the preservative effect of Arbutus unedo L. leaf extracts on the microbiological characteristics of quark cheese during storage. They found that incorporating the dry macerated leaf extract into the cheese resulted in higher efficacy against aerobic mesophiles and yeasts than the use of potassium sorbate after 8 days of storage. Milanović et al. [151] investigated the efficacy of seven essential oils against 74 spoilage yeasts. In a yoghurt model, lemongrass and cinnamon EOs demonstrated the highest antifungal activity in vitro. However, it should be noted that cinnamon EO inhibited lactic acid bacteria, while lemongrass EO displayed species-specific antifungal activity. These findings suggest that further research is needed to fully understand the application of plant antimicrobials in the dairy sector to control spoilage microorganisms.
The direct addition of natural plant antimicrobials in animal-based foods, such as meat, fish, and dairy products, shows the potential to control spoilage microorganisms and extend shelf life. Studies demonstrated the effectiveness of compounds such as phenolic acids, catechins, proanthocyanidins, and EOs in inhibiting the growth of spoilage bacteria. However, more research is necessary to fully understand the mode of action of these natural compounds and optimize their application in animal-based foods. Additionally, it is essential to consider the potential drawbacks, such as the development of off-flavours or bitterness, and address them through alternative delivery methods, such as nano-emulsions or active packaging.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Applications of plant antimicrobials on plant-based and animal-based foods against spoilage microorganisms.
Table 1.
Applications of plant antimicrobials on plant-based and animal-based foods against spoilage microorganisms.
| Food Matrix | Plant Antimicrobial | Concentration/Conditions | Antimicrobial Effect | Data from Ref.* |
|---|---|---|---|---|
| Mandarins | Pomegranate peel extract | Dipping in 25 g L−1 extract for 2 min | Reduction of lesion diameter and infection rate (80–90%) caused by P. italicum and P. digitatum | [118] |
| Fresh-cut lettuce | Pepper EO | 3–5 µL mL−1 addition in salad dressing | Reduction of P. fluorescens biomass by 30–40% | [129] |
| Concentrated apple juice | Thymol, carvacrol | MIC of 0.1–0.16 mM, treatment time 9 days | Reduction of Z. rouxii load by 99% | [134] |
| Pork burgers | Ethanolic extract of cranberry pomace | 2% extract-16 days of storage | Bacteriostatic effect on B. thermospacta and P. putida during cold storage | [140] |
| Snakehead fillets | Grape seed extract | 0.52 mg GAE mL−1 for 20 min | Decrease of Aeromonas spp. abundance by 37% and reduction of 1 log cfu g−1 of total viable counts during cold storage | [146] |
| Quark cheese | Arbutus unedo L. leaf extracts | 0.1 g 100 g−1 cheese, 8 days of cold storage | Reduction of total aerobic mesophilic bacteria and yeasts by 2–3 log cfu g−1 | [150] |
* as cited in the text.
3.2. Use of Plant Antimicrobials for Food Safety
The use of natural compounds derived from plants has numerous benefits, including the potential to provide safer, more sustainable and practical solutions for preserving food safety [152]. These plant-derived compounds showed high antimicrobial activity, making them ideal candidates as natural food preservatives. In particular, the correct use of these natural antimicrobials can fight emerging problems such as the spread of multidrug-resistant pathogens, biofilm-producing strains, and microbial toxins through the food chain.
3.2.1. Effect on Viability of Foodborne Pathogens
Foodborne bacteria are a significant public health concern since they can cause gastrointestinal illness, food poisoning, chronic diseases, economic losses, and the spread of antibiotic-resistant bacteria. Multiple foodborne illnesses were caused by various pathogens such as Sal. enteritidis, L. monocytogenes, E. coli toxigenic strains, Cam. jejuni, Cronobacter sakazakii, and S. aureus. Foodborne outbreaks underline the need for more efficient methods to control foodborne pathogens. Symptoms of foodborne illness can range from mild to severe, including nausea, vomiting, diarrhoea, abdominal cramps, and fever [153]. The outbreak of foodborne illnesses can have significant economic consequences, including loss of income for food producers, increased healthcare costs, and decreased consumer confidence in the food industry. Addressing the issue of foodborne bacteria is crucial to ensure the safety and quality of the food supply, protect public health, and minimize the economic impact of foodborne illnesses.
Several phytochemicals showed antibacterial activity against various foodborne pathogens. For example, studies demonstrated that plant compounds such as carvacrol and thymol, found in essential oils extracted from herbs and spices, have high antibacterial activity against Sal. enteritidis, E. coli, and L. monocytogenes [154]. Similarly, compounds such as cinnamaldehyde and eugenol, present in cinnamon EO and clove EO, respectively, inhibited the growth of foodborne pathogens such as L. monocytogenes and S. aureus [155,156]. These findings provide evidence of the potential of plant-based antimicrobials in controlling foodborne pathogens and improving food safety.
Specific applications of EOs or their compounds were described in plant and animal-based foods to ensure food safety. As regards animal-based foods, cinnamaldehyde inactivated L. monocytogenes at 4 °C in ground pork, reducing its viability by 4 log cfu g−1 in 5 days [155]. Similarly, thymol reduced, by 3 log cfu g−1, the load of S. aureus, E. coli, and C. perfringens on a sausage product during 4 weeks of storage [157]. In dairy products, myrtle EO (31.25 μL mL−1) reduced, by 1–2 log cfu g−1, the load of L. monocytogenes ATCC 679 on sheep cheese during ripening [158], whereas ginger (Zingiber officinale R.) and thyme EOs totally inactivated S. aureus (6 log cfu g−1) on a fresh soft cheese after two weeks of storage [159]. In plant foods, EOs or their compounds were proposed as sanitizers of fresh-cut vegetables and natural preservatives of fruit juices. Rossi et al. [160] treated fresh-cut lettuce contaminated with a cocktail of Salmonella spp. strains, with 5 µL mL−1 of cinnamon EO, reducing the attached cells by 0.6–0.8 log cfu cm−2. Cinnamon EO was also successfully used to control Sal. typhimurium and L. monocytogenes on celery, with a reduction of 2–4 orders of magnitude after 7 days at 4 °C depending on the initial contamination level [161]. As regards the application of EOs in fruit juices, Litsea cubeba Pers. EO reduced 3–4 log cfu mL−1 of the load of E. coli O157:H7 in four vegetable juices after 4 days of storage, and inhibited the respiratory metabolism, the topoisomerase activity, the transcription of virulence genes, and the nucleic acid replication [162]. In watermelon juice, Melissa officinalis L. EO reduced the viability of L. monocytogenes from 2 to 7 days of storage [163]. In some cases, plant antimicrobials can induce tolerance to environmental stresses in bacteria, and cross-resistance to common antibiotics. The use of Melissa officinalis L. EO at subinhibitory levels (0.125 μL mL−1) did not induce high tolerance to stresses (such as high temperature, low pH, osmotic stress, and desiccation) or cross-resistance with antibiotics in L. monocytogenes [163].
Plant phenolic compounds are naturally occurring compounds found in plants used as food preservatives due to their high antimicrobial activity against foodborne pathogenic bacteria [92]. Some of the most commonly used plant phenolic compounds in food include quercetin, and derivatives of cinnamic acid and gallic acid. Grape skin pomace extracts from different cultivars, rich in phenolic acids and flavonoids, showed higher antibacterial activity against Gram-positive strains than Gram-negative ones [164]. The addition of cranberry pomace extracts, rich in quinic and chlorogenic acids, procyanidin B3, myricetin and quercetin derivatives, delayed the growth of L. monocytogenes in cooked ham during cold storage [144]. Yersinia enterocolitica load was reduced by two logarithmic cycles in pork meat containing 5 mg g−1 of gallic acid [165]. Phuong et al. [166] evaluated the antibacterial activity of rambutan (Nephelium lappaceum L.) peel extracts, rich in geraniin, ellagic acid, rutin, quercetin, and corilagin as main phenolic compounds. The phenolic extract inhibited the growth of Sal. Enteritidis in raw chicken and that of Vibrio parahaemolyticus in fish during cold storage. The application of polyphenolic extracts or single polyphenols reduced the growth of foodborne pathogens in fresh-cut fruits, as demonstrated by using pomegranate peel extract or ferulic acid against L. monocytogenes on fresh-cut pear, apple, and melon [167,168]. The dipping of fresh-cut potatoes and fresh-cut lettuce in Centella asiatica L. extract significantly reduced the load of B. cereus and E. coli O157:H7 [131]. The glabridin, a prenylated isoflavonoid, reduced, by at least 1 log cfu g−1, the load of L. monocytogenes on fresh-cut cantaloupe during 4 days of cold storage [169].
As regards the application of the glucosinolate derivatives against food pathogens, the (4-[(4′-O-acetyl-α-l-rhamnosyloxy)benzyl] isothiocyanate) from Moringa oleifera seeds reduced the viable load of Cro. sakazakii and B. cereus in goat milk by three orders of magnitude [170].
In addition to the effect on cell viability, plant antimicrobials improved the thermal sensitivity of foodborne pathogens in the food matrix. In particular, the use of oregano EO in combination with citric acid enhanced the thermal inactivation of L. monocytogenes in sous-vide salmon cooked at 60 °C [171], whereas vanillin and emulsified citral improved the heat-sensitization of E. coli at 58 °C in a blended carrot-orange juice [172]. However, in certain conditions, plant antimicrobials can induce a viable but not culturable (VBNC) state in foodborne pathogens, as demonstrated for the application of citral and trans-cinnamaldehyde in a meat-based broth against S. aureus [173].
In contrast with many of the above-reported papers, the methanolic extract of spices mixtures employed to confer typical pungency and a hot taste to ‘Nduja, a traditional Calabrian sausage produced with about 20% of different spices, showed a limited inhibitory spectrum against ten common foodborne bacteria. Authors concluded that these spice mixtures, rich in hundreds of potentially antimicrobial compounds, can not exert an antimicrobial effect under normal processing conditions, due to the limited release of the bioactive compounds from the plant tissue [174]. In conclusion, the inclusion of plant antimicrobials in real food model systems can control the growth of foodborne pathogens, representing a valuable option to replace synthetic preservatives, even though their efficacy needs to be carefully evaluated under real production conditions.
3.2.2. Effect on Biofilm-Producing Strains
Bacterial biofilms are communities of microorganisms encased in a self-produced extracellular matrix and attached to a surface [22]. Biofilms are prevalent in many natural and artificial environments, including food processing facilities and equipment. Bacterial biofilms can cause serious problems in the food industry by contaminating food products, leading to foodborne illness and decreasing the product’s quality [115]. Biofilms can harbour pathogenic bacteria and provide a protective environment for these microorganisms, making them resistant to cleaning and disinfection procedures. This can result in a persistent contamination and the spread of foodborne illnesses. In addition, biofilm growing on the equipment surfaces can cause clogging, formation of corrosion, and degradation of the equipment surfaces, leading to increased maintenance costs and decreased productivity [175]. Bacterial biofilms are a significant concern in the food industry due to their impact on food safety and quality and the performance and efficiency of food processing equipment. The food industry needs to implement effective strategies to prevent and control the formation of bacterial biofilms to maintain a safe and efficient food processing environment.
The use of plant-derived antimicrobial compounds in food preservation gained attention due to their efficacy against foodborne biofilm-producing strains of bacteria [176]. These compounds act through various mechanisms, such as interference with metabolic processes, oxidative stress, and membrane disruption, and can also exert positive effects in inhibiting the growth and replication of biofilm-producing bacteria. The first anti-biofilm mechanism of action is the inhibition of the bacteria’s attachment to the surfaces. Phenolic compounds, such as phenolic acids, catechins, and quercetin, were found to reduce the adhesion of bacteria affecting flagellum, fimbria, and adhesins, delaying the formation of biofilms [12]. Red Globe and Carignan grape stem extracts, rich in caffeic, ferulic and gallic acids, catechin and rutin, inhibited the adhesion of L. monocytogenes to stainless steel and polypropylene surfaces by inhibiting motility and reducing the adhesion potential [177], as well as quercetin inhibited the early attachment of L. monocytogenes on stainless steel surface by increasing the cell permeability and reducing the superficial cell charge [178,179]. Quercetin also reduced the swimming and swarming motility of Sal. enterica at sub-MIC levels [180].
EOs or their compounds inhibit biofilm formation by different mechanisms. Cinnamon EO inhibited the adhesion of L. monocytogenes on polystyrene, but its efficacy was low on pre-formed biofilm [22]. Additionally, some terpenes, such as eugenol, carvacrol, and thymol, were demonstrated to suppress the production of exopolysaccharides in Salmonella spp., which are key components of bacterial biofilms [175], whereas citral and geraniol decreased the glucan production in E. coli O157:H7 [181]. Eugenol showed similar effectiveness against sessile and planktonic cells of S. aureus, showing a lower resistance coefficient, the ratio of concentrations required to achieve the same log reductions in both populations (Cbiofilm/Cplanktonic), as compared to conventional disinfectants [182]. Carvacrol and oregano EO effectively inhibited biofilm formation by S. aureus on stainless steel surfaces, but the long-term exposure to a sub-MIC concentration of the oregano EO showed an inductive biofilm formation effect [183]. Another mode of action of plant antimicrobials against foodborne biofilm-producing bacteria is destabilizing the biofilm matrix. Compounds such as sulphides, including allicin and diallyl sulfide, and sulfites were shown to penetrate the biofilm and disrupt its stability, causing the release of bacteria from the biofilm [184], as demonstrated in uropathogenic E. coli [185].
On the other hand, plant-derived antimicrobial agents were shown to possess anti-biofilm activity by disrupting the quorum sensing process [186]. Quorum sensing is a communication mechanism that bacteria utilize to coordinate the expression of certain genes, including those involved in biofilm formation. Phytochemicals such as flavonoids (quercetin and kaempferol) and terpenoids (carvacrol and thymol) were demonstrated to interfere with the quorum-sensing by inhibiting the production and the activity of autoinducers (e.g., acyl-homoserine lactone), which play a key role in the quorum-sensing process [187,188]. Lippia origanoides K. EO (thymol-carvacrol chemotype) inhibited the expression of the sdiA, luxS, and luxR genes, which were implicated in the quorum-sensing of Sal. enteritidis. This effect could be related to the inhibition of the biosynthesis of autoinducers or the interference with the reception of acyl-homoserine lactone [189]. Aqueous pomegranate extract showed anti-quorum sensing activity, reducing the violacein production, the quorum-sensing system’s product, in Chromobacterium violaceum [186]. Curcuma longa L. extract, with curcumin and curcumin derivatives as main compounds, showed anti-quorum sensing activity inhibiting the violacein production in C. violaceum, probably disrupting the signal reception or the absorption of the acyl-homoserine lactone. However, this extract showed lower anti-biofilm activity against food pathogens than Camellia sinensis L. extract, rich in epigallocatechin and epicatechin [190].
The use of plant antimicrobials as a strategy to control the biofilm formation in foodborne pathogens gained increasing attention in recent years. Phytochemicals such as phenolic acids, tannins, sulphur compounds, and terpenoids (Figure 2) showed anti-biofilm activity by interfering with the quorum-sensing process of bacteria [191]. Despite these promising results, further research is needed to fully understand how these compounds exhibit antibacterial and anti-biofilm activity and to develop effective strategies for controlling biofilm formation during food processing.
3.2.3. Effect on Microbial Toxins
Microbial toxins (e.g., bacterial toxins and mycotoxins) harm human health. Plant antimicrobials were evaluated to reduce toxin production by foodborne bacteria and mycotoxins by filamentous fungi.
Bacterial exotoxins are proteins that damage host cells and are important for the pathogenesis of many bacterial pathogens, such as Clostridium spp., E. coli, L. monocytogenes, and S. aureus [192]. The use of plant antimicrobials can attenuate the virulence of these foodborne pathogens. In particular, different flavonoids suppressed the toxin production in different foodborne pathogens. Genistein inhibited the exotoxin produced by S. aureus, kaempferol, kaempferol-3-O-rutinoside, quercetin glycoside inhibited the neurotoxin production from Cl. botulinum, and green tea catechins inhibited the release of verotoxin from enterohemorrhagic E. coli [16]. Recent findings showed that the water-soluble fraction of the Eucalyptus camaldulensis Dehnh. leaf extract significantly reduced the listeriolysin O-induced haemolysis in L. monocytogenes at sub-inhibitory concentrations [193]. A witch-hazel extract, with hamamelitannin as the main phenolic compound, inhibited the production of the staphylococcal enterotoxin A in S. aureus at non-inhibitory concentrations for microbial cells [194]. As regards the EO and their compounds, sub-inhibitory concentrations of tea tree EO downregulated the transcription of genes encoding α-hemolysin, staphylococcal enterotoxin A, and staphylococcal enterotoxin B in S. aureus, inhibited their production, and the hemolytic activity [195]. Zhang et al. demonstrated that citronellal significantly reduced the production of enterotoxins in S. aureus-contaminated pork meat without reducing the viable cell load [196]. Other EO compounds, such as carvacrol and trans-cinnamaldehyde, reduced the production of TcdA and TcdB toxins produced by Cl. difficile in in vitro conditions [197]. Organic sulphur compounds such as the diallyl disulphide, at sub-inhibitory concentrations, reduced the production of the B. cereus enterotoxins Nhe and Hbl [91].
Plant antimicrobials also showed the ability to control the mycotoxin production by filamentous fungi. The mechanisms of action are the inhibition of the fungal growth and the induction of xenobiotic detoxification and/or the activation of biotransformation pathways [19]. In the first case, turmeric, rosemary and clove EOs demonstrated great efficacy in controlling the growth of mycotoxigenic A. flavus through the inhibition of ergosterol biosynthesis, the disruption of the fungal cell membrane, and the production of reactive oxygen species (ROS). In some cases, essential oils showed anti-aflatoxigenic activity at concentrations inhibiting or completely suppressing fungal growth. In contrast, in other cases, the anti-aflatoxigenic activity was detected at non-inhibiting concentrations. However, in a few cases, plant antimicrobials stimulated the production of secondary metabolites, including mycotoxins, in Aspergillus species [19]. Natural flavonoids such as baicalein, flavone, hispidulin, kaempferol, and liquiritigenin reduced the aflatoxin production in maize kernels contaminated with A. flavus by 50–67% [198], whereas a ternary mixture of naringin, neohesperidin, and quercetin reduced the aflatoxin accumulation in maize contaminated with A. parasiticus by more than 85% [199]. In sausages, the combined application of Salvia farinacea Benth. and Azadirachta indica A.Juss. extract at 2 mg mL−1 suppressed the production of ochratoxin A and aflatoxin B1 produced by A. ochraceous and A. parasiticus, respectively [200]. The degradation of aflatoxin B1 treated with the leaf extract from rosemary reached 60% after 48 h of incubation. Araçá (Psidium cattleianum S.) and oregano extracts produce less degradation than rosemary extract. Substances such as alkaloids and enzymes occurring in the plant extract might be involved in the structural modification of aflatoxin B1 [201]. Although the effect of plant antimicrobials on mycotoxin accumulation in food products is promising, more in-depth information regarding the toxicity of the resulting compounds from the degradation activity is required.
In conclusion, plant antimicrobials can reduce or suppress the production of bacterial and fungal toxins by reducing microbial growth or downregulating toxin gene expression. Further research is necessary to understand the modes of action of different plant extracts and their bioactive compounds on toxin production to exploit their potential to improve food safety under real contamination conditions.
4. Stabilization Techniques
Plant antimicrobials can have limited stability under processing or storage conditions of foods. The efficacy of plant antimicrobials is affected by several factors such as pH, the temperature, and the concentration. Caffeic, chlorogenic, and gallic acids are not stable at high pH values, whereas chlorogenic acid is stable at low pH values and heat [202]. Some phenolic compounds and EOs, and their compounds, are thermolabile. Achillea sp., rosemary, sage (Salvia officinalis L.), and thyme EOs were more effective at low pH and low temperature against pathogenic bacteria [203], whereas carvacrol and cymene showed higher antibacterial activity in carrot juice at 25 °C than at 4 °C and 15 °C [204]. Several plant antimicrobials show a dose-dependent effect against spoilage and pathogenic microorganisms. The stabilization techniques described in this section can help to protect plant antimicrobials and, in some cases, reduce the concentration necessary to exert their antimicrobial activity. The direct addition of plant extracts or their bioactive compounds in foods is the most common method of food preservation. However, the direct addition of plant extracts is often responsible for changes in sensory properties such as flavour and texture. In addition, the bioavailability of these compounds and their effectiveness in improving food safety can be affected by the interaction with the macronutrients and ingredients. For these reasons, several stabilization techniques were proposed to enhance stability, drive the release of bioactive compounds during storage, and reduce the negative effects of plant extracts on the sensory characteristics of foods.
4.1. Nano-Emulsions
The encapsulation of plant antimicrobials into edible colloidal delivery systems is a promising method to enhance the efficacy of these substances and reduce the negative effects due to the interaction with food ingredients. In particular, encapsulation in small particles increases water dispersibility and resistance to environmental conditions enhancing plant antimicrobials’ efficacy [6]. Oil-in-water nano-emulsions containing lipid nanoparticles dispersed in water are currently the most common delivery system for plant antimicrobials. These nano-emulsions can be manufactured from food-grade ingredients, such as plant-based emulsifiers and different stabilizers, using common processing methods, such as mixing (low-energy emulsification), sonication, and homogenization (high-energy emulsification) [6].
Different studies investigated the efficacy of nano-emulsions against foodborne pathogens. The plant antimicrobials most used to prepare nano-emulsions are the EOs and their compounds. Lemongrass, clove, thyme, or palmarosa (Cymbopogon martini Will. Watson)-loaded EOs nano-emulsions, prepared after micro fluidization of the primary emulsion, inactivated E. coli by 3–4 log cfu mL−1. The use of alginate in the aqueous phase is useful for applying these nano-emulsions in the coating material of fruits and vegetables [205]. Anise (Pimpinella anisum L.) oil nano-emulsions showed the same MIC (1% v/v) of the bulk EO and coarse emulsion against L. monocytogenes and E. coli O157:H7. However, the anise oil nano-emulsion displayed the highest physical stability and antibacterial efficacy [206]. More recently, other plant antimicrobials were used to prepare nano-emulsions to control the growth of bacterial pathogens. Anise seed extract, with anethole, naringenin, and taxifolin as main compounds, was used to develop an antibacterial nano-emulsion using the ultrasound emulsification method. The nano-emulsion was active against E. coli and Sal. thyphimurium, whose growth was not affected by the bulk extract [207]. Ghazy et al. [208] evaluated the antimicrobial action of henna (Lawsonia inermis L.) extract as a nano-emulsion against seven pathogenic bacteria. The nano-emulsion, rich in catechin, methyl gallate, ellagic acid, and coumaric acid, displayed higher antimicrobial activity against E. coli, and B. cereus, than the course emulsion. Regarding the application of nano-emulsions, including plant essential oils, to control pathogens in plant foods, oregano oil nano-emulsion at 0.1% reduced the load of L. monocytogenes, Sal. typhimurium, and E. coli O157:H7 on lettuce by 3 log cfu g−1 [209]. Cinnamon oil nano-emulsion at 0.5% determined more than five log reductions in L. monocytogenes and Salmonella spp. on melon [210]. Lemongrass and mandarin (Citrus reticulata Blanco) EO nano-emulsions inactivated E. coli in apple juice, but when the nano-emulsions were prepared directly in the apple medium as a continuous phase, the antibacterial efficacy was reduced in comparison to the use of water [211]. Citral nano-emulsions at 0.15 μL mL−1 inactivated L. monocytogenes (5 log cfu g−1 reduction) on fresh-cut melon and papaya during cold storage [212].
Regarding the efficacy of nano-emulsions including plant antimicrobials against spoilage microorganisms, thyme EO nano-emulsion showed lower efficacy than bulk EO against fish spoilage bacteria, except for Serratia liquefascens [213]. For this spoilage bacteria, laurel (Laurus nobilis L.) and grapefruit (Citrus paradisi Macfad.) EO nano-emulsions showed lower MIC values than the corresponding EOs [214,215]. Ginger EO nano-emulsion, prepared with zein and sodium caseinate as co-emulsifiers, showed higher bactericidal activity against total viable counts of chicken breasts than the bulk EO, extending the shelf life of the product by 6 days [216]. As regards the antifungal activity of plant antimicrobial nano-emulsions, cinnamaldehyde, eugenol, and carvacrol nano-emulsion showed a dose-dependent effect against the spore germination and mycelial growth of P. digitatum, with a MIC value of 0.125 mg mL−1 [217]. Gundewadi et al. [218] found that basil EO nano-emulsion displayed lower lethal concentration values (LC50) than course emulsion against P. chrysogenum and A. flavus during 8 days of incubation. Oregano and clove EOs nano-emulsions, at 1.95 mg g−1, showed fungicidal activity against Z. bailii in a salad dressing after 4 days of storage [219]. EOs nano-emulsions also showed anti-mycotoxigenic activity. Indeed, lemongrass EO nano-emulsion reduced by 99.5% the deoxynivalenol content in rice contaminated with F. graminearum. The lemongrass EO nano-emulsion showed better anti-mycotoxigenic activity than the bulk EO, but the efficacy was strain-specific [220]. Oregano EO encapsulated into chitosan nano-emulsion suppressed the production of aflatoxin B1 by A. flavus in maize [221].
Given these results, the antimicrobial action of plant antimicrobial nano-emulsions depends on the chemical composition of the plant extract, the emulsion droplet size, and the target microbial species. In addition, many studies demonstrated higher efficacy of nano-emulsion than course emulsion and bulk plant extract.
4.2. Spray-Drying and Encapsulation
Spray-drying and encapsulation are techniques commonly used to improve the stability and functionality of plant antimicrobials in food products. Spray-drying is a process in which a liquid solution or suspension is atomized into a hot air stream, causing the rapid evaporation of droplets, resulting in a dry powder. This process can produce dry powders of plant antimicrobials that are more stable and easier to handle than the liquid form. Spray-drying can also encapsulate the plant antimicrobials in a protective matrix, improving their stability and functionality. Encapsulation is a process in which a natural antimicrobial is surrounded by a protective matrix, such as a polymer or lipid, to improve its stability and functionality. Encapsulation can enhance the natural antimicrobials’ shelf life and protect them from degradation induced by light, heat, or moisture. Additionally, encapsulation can improve the solubility and dispersibility of natural antimicrobials, making them easier to incorporate into foods. These techniques can help to preserve the antimicrobial activity of the natural antimicrobials and improve their effectiveness in controlling the growth of spoilage and pathogenic microorganisms.
4.2.1. Spray-Drying Process
Spray-drying and encapsulation techniques provides numerous benefits in handling, storage, and transportation of plant antimicrobials. Powdered antimicrobials are more suitable for various applications within the food industry [222]. Powdered antimicrobials minimize the risk of spillage and waste during handling and processing, as they can be easily measured and transferred without causing mess or loss of material. This ensures a more efficient use of resources and reduced operational costs. In some cases, the spray-drying process conditions can have pros and cons related to the stability and functionality of plant extracts, as briefly pointed out in Table 2.
In the study of Chen et al. [223], eugenol and thymol were co-encapsulated into zein-casein nano-capsules through spray-drying. The resulting powders showed good water hydration, stability during storage, controlled release during 24 h, and bactericidal and bacteriostatic effects against E. coli O157:H7 and L. monocytogenes in milk whey, respectively. Thyme EO encapsulated by spray-drying, with casein and maltodextrin as wall materials, showed antibacterial action against thermotolerant coliforms and E. coli in meat burgers [228]. The wall material employed to protect plant antimicrobials can affect their antibacterial action. Indeed, the inclusion of chitosan in a whey protein/maltodextrin blend reduced the antibacterial action of eugenol against E. coli and L. innocua. A low inlet temperature used in the spray-drying of pectin/sodium alginate capsules including carvacrol, increased the antibacterial activity against E. coli K12 (Table 2). The use of nano spray-drying, a novel process to produce plant antimicrobial powders, was evaluated to obtain whey protein/maltodextrin capsules, including oregano EO. The capsules showed enhanced antibacterial action against E. coli and S. aureus compared to pure EO. However, this process has drawbacks such as high production costs, high processing time, and reduced spraying effectiveness of viscous solutions [226]. These examples illustrate the potential benefits and drawbacks of the spray-drying technique to produce plant antimicrobial powders. The specific advantages and disadvantages observed depend on the antimicrobial compound, the spray-drying conditions, and the choice of the protective matrix.
The increased stability of powdered antimicrobials extends their shelf life, as demonstrated for the encapsulated peanut (Arachis hypogaea L.) skin extracts [229]. It maintains the efficacy of plant antimicrobials throughout storage, reducing the need for frequent replacements and ensuring consistent antimicrobial action.
Powdered antimicrobials generally have lower storage requirements than their liquid counterparts, as they do not require refrigeration or specific storage conditions to maintain their stability. This reduces energy consumption and storage costs for food manufacturers. Additionally, the nature, weight, and form of powdered antimicrobials facilitates more efficient transportation and shipping, as they occupy less space and require less protective packaging than liquid antimicrobials [222]. Powdered antimicrobials can be more easily integrated into various food matrices, as their fine and uniform particles allow a more homogeneous distribution throughout the product. This ensures consistent antimicrobial protection across the whole food matrix, enhancing food safety and quality.
4.2.2. Other Encapsulation Techniques of Plant Antimicrobials
Encapsulation of plant antimicrobials, a method of entrapment of a core material within another solid or liquid immiscible substance, allows the production of capsules or spheres in micrometre to millimetre in size [230]. Encapsulation can involve various types of protective matrices that impact the stability and functionality of the antimicrobial agents. Some common matrices for encapsulation include polysaccharides, lipids, and proteins. Polysaccharides such as alginate, chitosan, and maltodextrin are widely used as encapsulating agents due to their biocompatibility, non-toxicity, and excellent film-forming properties. A study by de Araújo et al. [231] demonstrated that using maltodextrin/gelatine mixtures as a protective matrix for encapsulating the sweet orange EO positively affected the thermo-oxidative stability of bioactive compounds and maintained its antibacterial properties. The encapsulation of plant antimicrobials can be obtained through the formation of inclusion complexes using the β-cyclodextrins, cyclic oligosaccharides with amphipathic properties. These complexes can stabilize the guest molecule against the degradation, mask off-flavours, and control the release of the encapsulated compounds [232]. Thyme EO microcapsules exerted a bacteriostatic effect over Enterobacteriaceae, mesophilic bacteria, and psychrotrophic bacteria on lettuce [232], whereas inclusion complexes with rosemary EO showed better antimicrobial activity against Saccharomyces pastorianus than free EO in pasteurised tomato juice [233]. Coriander (Coriandrum sativum L.) EO encapsulated in β-cyclodextrin nano-sponge showed bactericidal activity against L. monocytogenes, Y. enterocolitica, and Cam. jejuni in aqueous media [234]. Black pepper (Piper nigrum L.) oleoresin was stabilized in β-cyclodextrins using the kneading method, a method in which the β-cyclodextrins and the guest compound are mixed with small amounts of ethanol or water using a kneader for a specific time, showing antimicrobial activity against L. monocytogenes and improved thermal stability [235].
Lipid-based encapsulation systems, such as solid lipid nanoparticles, the use of nanostructured lipid carriers, and liposomes, are also employed for encapsulating plant-derived antimicrobials. These systems can improve the stability of the encapsulated compounds, their bioavailability, and ensure a controlled release. The study by Lin et al. [236] reported that encapsulating chrysanthemum (Chrysanthemum flosculosum L.) EO in triple-layer liposomes led to long-term antimicrobial activity against Cam. jejuni in chicken.
Protein-based matrices, such as gelatine, soy protein, and whey protein, can also encapsulate plant-derived antimicrobials. These matrices offer advantages in biodegradability, biocompatibility, and the ability to form stable complexes with antimicrobial agents. Recently, the microencapsulation of cinnamon EO using chitosan and whey protein isolate showed enhanced thermal stability and long-term antimicrobial effect against S. aureus, E. coli, P. fragi, and Shewanella putrefaciens [237]. The selection of the most suitable matrix depends on the specific antimicrobial compound, the target application, and the desired release characteristics.
4.2.3. Challenges Associated with Spray-Drying and Encapsulation Techniques
Although spray-drying and encapsulation techniques offer various advantages for the stabilization and incorporation of plant-derived antimicrobials into food systems, there are challenges associated with these processes that require further research. One of the issues associated with spray-drying is the potential degradation or loss of activity of heat-sensitive plant compounds during the drying process, as high temperatures are often involved [222]. This can lead to reduced antimicrobial efficacy or the modification of sensory properties. Additional and specific research studies need to be carried out to explore alternative drying techniques, such as freeze-drying or nano spray-drying, that might better preserve heat-sensitive compounds. Another challenge is the selection of the most appropriate encapsulation matrix to ensure optimal protection, release, and stability of the encapsulated antimicrobial compound. The choice of the encapsulation material can greatly affect the effectiveness and shelf life of the antimicrobial agent for food applications [238]. Further research is needed to understand the interactions between various wall materials and plant-derived antimicrobials, and to optimize the encapsulation processes for specific food systems. Additionally, scaling up from lab-scale to industrial-scale production of encapsulated plant-derived antimicrobials poses challenges related to the encapsulation efficiency, the product stability, and the process economics [239]. More research is also required to develop cost-effective and efficient methods for large-scale production, maintaining the quality and functionality of the encapsulated antimicrobials.
4.3. Active Packaging
Active packaging involves the deliberate inclusion of subsidiary constituents in or on either the packaging material or the package headspace to enhance the performance of the package system. Active packaging can preserve the food quality and can extend the product’s shelf life through the direct interaction between the food and bioactive substances intentionally incorporated into the package [240]. Antimicrobial packaging is one type of active packaging, in which the antimicrobial activity strongly depends on the migration rate of the biologically active molecule incorporated into the polymer matrix [240]. Recently, the market was oriented to replace packaging produced with fossil fuels with more sustainable materials such as biopolymers. In this context, the use of biopolymers, including plant antimicrobials, is very attractive to develop active films or coatings, to overcome the thermal/oxidative instability of these compounds during the manufacturing of the polymers or storage of the final product, and to mask undesirable sensorial aspects of some plant extracts. In addition, biopolymers have a lower environmental impact compared to standard food packaging polymers such as polyethylene or polypropylene. Plant antimicrobials can be added directly into the biopolymer or loaded into clays or nanocarriers, as demonstrated for EO compounds [241,242] and polyphenols [243,244]. This strategy ensures, in most cases, their controlled release and, in some cases, an improvement in the mechanical and physical properties of the film.
The most used biopolymers for including plant antimicrobials are chitosan, starch, carrageenan, cellulose, and alginate. However, other polymers used for this purpose are polyvinyl alcohol (PVA), poly lactic acid (PLA), poly butylene-succinate-co-adipate (PBSA), poly butylene-adipate-co-terephthalate (PBAT), poly(hydroxybutyrate)s (PHBs), and poly (ε-caprolactone) (PCL).
Regarding active chitosan films, the inclusion of apple peel polyphenols (1%) into chitosan film enhanced the antibacterial activity against B. cereus, E. coli, Sal. typhimurium, and S. aureus [245]. A composite film based on grapefruit seed extract-loaded poly(ε-caprolactone)/chitosan reduced the E. coli population on salmon by more than 2 log cfu g−1 after 6 days at 4 °C compared to the packaging into polyethylene or poly(ε-caprolactone)/chitosan films, and suppressed mould development on bread stored for 7 days at 24 °C [246]. Surendhiran et al. [247] developed active nanofibers based on chitosan/Poly (ethylene oxide) loaded with pomegranate peel extract. The nanofibers reduced by 3 log cfu g−1 the E. coli O157:H7 population in raw beef stored at 4 °C for 10 days. The coating of fresh cucumber with chitosan loaded with oregano EO reduced the viability of total mesophilic bacteria and total yeasts and moulds during storage at 10 °C for 15 days [248]. Starch films were also enriched with plant antimicrobials as demonstrated by Saberi et al. [249] that developed pea starch-guar gum films including epigallocatechin-3-gallate and blueberry ash fruit (Elaeocarpus reticulatus Sm.) and macadamia (Macadamia tetraphylla) skin extracts. Active films showed antimicrobial activity against spoilage bacteria and fungi, and pathogenic bacteria, with a reduction in microbial load in the range of 40–80% for the films loaded with epigallocatechin-3-gallate and blueberry ash fruit skin extracts at the MIC level (ranging from 93 to 1500 μg mL−1). A bio-composite film made with cassava starch and whey protein loaded with rambutan peel extract and clove oil slightly inhibited B. cereus, E. coli, and S. aureus in in vitro conditions, and reduced total viable count of salami stored for 10 days [250]. A sweet potato starch-based film activated with montmorillonite nano-clay and thyme EO reduced the load of E. coli and S. thypimurium on fresh spinach leaves during 8 days of cold storage [251]. Other biopolymers used to manufacture active films/coatings with plant antimicrobials are alginate and carrageenan. A sodium alginate film loaded with the gallnut extract (Quercus infectoria Oliv.), rich in gallotannins, ellagic acid, and gallic acid, showed antibacterial activity against S. aureus and E. coli [252], whereas the coating of apples and pears with alginate loaded with cinnamon EO at 0.9% v/v inhibited the A. carbonarius growth and the OTA production [253]. Compared with the control film, a carrageenan film containing 3% rosemary extract displayed >99% inhibition against B. cereus, E. coli, P. aeruginosa, and S. aureus, reducing by 2–4 orders of magnitude the microbial load [254]. He and Wang [255] recently demonstrated that a K-carrageenan coating enriched with cinnamon EO delayed the growth of total viable count, lactic acid bacteria, and H2S-producing bacteria in pork meat. Active films incorporating plant antimicrobials can be also produced using proteins. Indeed, zein nanofibers loaded with 1,8-cineol rich extracts reduced the load of L. monocytogenes and S. aureus on cheese slices by two orders of magnitude during 28 days of cold storage [256].
Other active films, including plant antimicrobials, can be manufactured using PLA and PHBs. A PLA/PBAT composite film including 7% w/w of grapefruit seed extract showed bactericidal activity against L. monocytogenes and a bacteriostatic effect against E. coli [257]. Additionally, a PLA/PBSA blend including 6% w/w thymol delayed the mould development on bread compared to polypropylene or neat PLA [258]. PHBV films loaded with eugenol and carvacrol showed antibacterial action against E. coli in cheese and pumpkin but not in melon, where the highest release of the active compounds from the films was observed [259]. This result highlights that the interaction of antimicrobial compounds with the food components and their diffusion within the food matrix play an important role in the antimicrobial activity of active films including plant antimicrobials.
Plant antimicrobials can be incorporated into active packaging using encapsulated extracts or nano-emulsions. Pabast et al. [260] developed a chitosan film in which the EO extracted from Satureja khuzestanica J. was encapsulated into nanoliposomes. The film delayed the growth of total mesophilic bacteria, Pseudomonas spp., and lactic acid bacteria of lamb meat stored for 20 days at 4 °C. Interestingly, the antimicrobial effect was higher than that displayed by the film including free Satureja khuzestanica J. EO [260]. Chitosan film loaded with microcapsules, including basil EO, slightly reduces total mesophilic bacteria, enterobacteria, and lactic acid bacteria on cooked ham during storage [261]. A PVA loaded with cinnamon EO encapsulated in β-cyclodextrin showed a bacteriostatic effect against S. aureus and E. coli [262]. In addition, a PVA/starch film including β-cyclodextrin inclusion complex embedding lemongrass EO showed antibacterial action against She. putrefaciens [263]. Nano-emulsions of essential oils were used for the inclusion in active packaging. Lee et al. [264] developed hydroxypropyl methylcellulose-based films incorporating oregano EO nano-emulsions. The active film showed inhibition zones against Sal. thyphimurium, E. coli, L. monocytogenes, B. cereus, and S. aureus. Chitosan- Ferulago angulata essential oil nano-emulsion showed lower MIC and MBC values against the fish-spoilage bacteria P. fluorescens and She. putrefaciens than the corresponding coating emulsions. Moreover, the coatings, including the nano-emulsion, reduced the total viable and psychrotrophic counts of rainbow trout fillets by 3 log cfu g−1 after 16 days of storage at 4 °C [265]. The incorporation of Zataria multiflora Boiss. EO and cinnamaldehyde in the form of nano-emulsions into starch coatings reduced the growth of L. monocytogenes, psychrotrophic bacteria, and Enterobacteriaceae in chicken during cold storage [266].
5. Combining Effects and Hurdle Technologies
In this section, the more recent studies demonstrating the possible improvement in food safety and shelf life by combination of bioactive compounds from different plant-based extracts or by their combination with non-thermal or mild food technologies are briefly reported.
5.1. Additive or Synergistic Effects
The use of combinations of antimicrobial plant extracts and their compounds showed additive or synergistic effects against spoilage and pathogenic microorganisms. This approach is cost-efficient for the food industry and adheres to the hurdle technology in inhibiting the proliferation of undesirable microorganisms, improving the preservative effects of plant antimicrobials and reducing the negative sensory effects of single plant extracts [2].
Additive or synergistic effects are found in combinations of plant extracts and their compounds, reducing the MIC of the plant antimicrobials. As regards the additive effects, cinnamon EO with clove EO showed an additive effect against L. monocytogenes [67], whereas the combination of cinnamaldehyde with 2-hydroxycinnamic acid showed additive effects against L. monocytogenes and Sal. enteritidis under in vitro conditions, but it was not effective in contaminated cooked ham [267]. Regarding the synergism among plant antimicrobials, thyme EO with cinnamon EO (0.312 g L−1), cinnamon EO (0.156 g L−1) with rosemary EO (0.625 g L−1) and thyme EO (0.078 g L−1) showed synergistic effects in inhibiting Alt. alternata and P. expansum on jujube fruit [268]. Cinnamon EO with clove EO showed synergistic antibacterial activity against S. aureus, L. monocytogenes, and Sal. typhimurium [269], and against L. monocytogenes when vanillin was combined with both EOs [67]. Synergistic effects were found using combinations of EOs compounds or isothiocyanates with phenolic acids against bacterial pathogens. The combination of thymol with gallic acid determined a synergistic effect at sub-inhibitory concentrations against E. coli O157:H7 and S. aureus on fresh-cut tomatoes [270]. Allyl isothiocyanate with o-coumaric acid showed synergism, obtaining 2 log reduction in E. coli O157:H7 in a dry-fermented sausage when the antimicrobial compounds were added at the concentration of 6.25 μL and 750 mg per 100 g fresh weight, respectively [271].
As regards the antifungal interactive effect of plant antimicrobials, a triple combination of thyme EO, cinnamon EO, and rosemary EO showed a synergistic antifungal effect against B. cinerea and P. expansum, reducing their development on pear [272]. Pinto et al. [64] demonstrated a synergistic effect in the vapour phase between thymol and γ-terpinene in binary combinations and between p-cymene, γ-terpinene, and thymol in ternary combinations against the strain P. digitatum ITEM 9569, which is resistant to single thyme EO exposure. The use of combinations of plant antimicrobials to control the development of spoilage and pathogenic microorganisms in foods is a research area showing rapid development. The use of combinations of plant antimicrobials can reduce the concentration of plant antimicrobials added in foods, minimizing the negative impact of these compounds on the sensory properties of foods as previously demonstrated [269,270]. Studies related to this topic are expected to increase in the future, paying attention to the effect of the combination of plant antimicrobial compounds in real food matrices, and the elucidation of the modes of action.
5.2. Hurdle Technologies
The “hurdle approach” in the food sector refers to the successive or simultaneous application of two or more food preservation techniques for enhancing food safety and quality using lower individual treatment intensities and for achieving multi-target, mild, and reliable preservation effects [273]. This approach was followed to reduce the dose of chemical preservatives used to control the development of spoilage microorganisms in foods [274]. Recently, several studies investigated the application of mild or non-thermal technologies in combination with the use of plant antimicrobials to control the growth of spoilage and pathogenic bacteria in food and to extend the shelf life.
The combined application of high-pressure homogenization (HPH) and nano-emulsions of hexanal and trans-2-hexanal inactivated S. cerevisiae in apple juice up to 22 days of storage, with better performance as compared to individual treatments [275]. Citral (1% w/w) combined with high-pressure processing reduced the viability of a cocktail of E. coli STEC in ground beef by 4–7 log cfu g−1, depending on the pressure level applied [276]. The treatment of ground chicken meat with 320 MPa for 23 min at 4 °C with allyl isothiocyanate and acetic acid at ca. 0.2% w/w achieved a 5-log reduction in E. coli O157:H7, with a better inactivation compared to single treatments [277]. As regards the application of cold plasma technology with plant antimicrobials, González-González et al. [278] found that the combined application of cold plasma and linalool nano-emulsion reduced by 3 log cfu g−1 the load of E. coli O157:H7 and Sal. enterica in chicken meat, while individual treatments showed limited efficacy. The effect of plant antimicrobials combined with food technologies against E. coli O157:H7 on chicken meat is depicted in Figure 3.
Sea bass slices packed under modified atmosphere packaging and pre-treated with cold plasma and a liposomal ethanolic coconut husk extract showed the lowest increase in Pseudomonas spp. and Enterobacteriaceae during 18 days at 4 °C in comparison to cold plasma treatment alone or the application of the liposomal ethanolic coconut husk extract [279]. On the contrary, the combined use of cinnamon EO and modified atmosphere packaging showed limited efficacy against spoilage bacteria of lean pork meat or salmon during cold storage [280]. The pulsed electric field pre-treatment of Pacific white shrimp, followed by the soaking in 1% of Chamuang (Garcinia cowa Roxb.) leaf extract, showed a lower increase in mesophilic, psychrophilic, Pseudomonas spp., Enterobacteriaceae, and H2S producing bacterial counts in comparison to individual treatments and the application of sodium metabisulfite during cold storage [281]. A synergistic effect in reducing the microbial load of E. coli O157:H7 was found between thyme EO nano-emulsion treatment and the sonoporation induced by ultrasounds [282]. Microwave heating at 915 MHz with carvacrol showed a synergistic effect against E. coli O157:H7, Sal. typhimurium and L. monocytogenes in buffered peptone water but not in hot chilli sauce [283].
Light technologies were also combined with the use of plant antimicrobials to control the contamination by foodborne pathogens. In reconstituted powdered infant formula, the load of Cro. sakazakii was reduced by 6.5 log cfu mL−1 following the combined 405 nm light-emitting diode and 9 µL mL−1 citral treatment for 90 min compared with untreated samples [284]. Silva-Espinosa et al. [285] found that the combined application of UV-C light and clove EO on stainless steel surface achieved a complete bacterial reduction (6.8 log cm−2) on biofilms of Sal. thyphimurium.
The use of chemical compounds was associated with plant extracts and their compounds. The nano-emulsion of thyme EO enhanced the antimicrobial effect of slightly acidic electrolyzed water against foodborne pathogens, suggesting the formation of complexes probably through hydrophobic interactions [286]. Based on total viable counts, a shelf-life extension of 7 days was observed in fresh fish fillets treated with gaseous ozone and coated with alginate, including different EOs and citrus extract, as compared to single treatments [287].
The combination of mild or non-thermal technologies with plant antimicrobials is a recent research trend. Many of these studies showed synergistic effects in reducing the microbial load of spoilage and pathogenic microorganisms on foods. These effects can be explained by the exposure of target microorganisms to multiple hurdles and stresses. Even though more studies are necessary to understand the modes of action of these combined approaches, the enhanced antimicrobial activity of plant extracts and their compounds, when coupled with innovative technologies, allows to reduce their concentration in different foods, mitigating some of their drawbacks such as the modification of sensory characteristics.
6. Regulation and Safety Issues of Plant Extracts
Plant extracts can be contaminated by different dangerous compounds, such as heavy metals, crop-protection residues, and mycotoxins. The concentration of these compounds depends on the cultivation practices employed, the geographical location of the cultivation site, the application of crop protection products, and the extraction method. In this section, data related to the contamination levels of plant extracts employed for the production of plant antimicrobials for food purposes are reported.
6.1. Heavy Metals and Crop-Protection Residues
Heavy metals can contaminate plant extracts. In plants that produce EOs, the uptake of metals is associated with soil contamination, and their transfer to EOs depends on the extraction technology. Moreover, the storage of EOs in metallic containers can promote the transfer of metals into the oil. As, Cd, Pb, and Hg cause toxic effects at relatively low levels. For Cd, Hg, and Pb, recommendations for safety limits in medicinal plants are imposed by European Pharmacopoeia, FAO, and the World Health Organization (WHO). Iordache et al. [25] evaluated the heavy metal content of EOs from different sites. High levels of Hg, Cr, Pb, Cu contamination were found in Mentha × pipperita L. EO. However, the authors concluded that the analyzed EOs could be safely consumed in the doses recommended by the manufacturers, and the content of heavy metals does not pose a significant risk to the consumer’s health. High levels of Cr, Cd, and Pb in thyme and oregano plant samples were found by Reinholds et al. [288].
Regarding the contamination levels of crop-protection residues in EOs, they depended on the agricultural practices. Conventional orange EOs contained 17 pesticides and a total concentration of 5.1 mg L−1, whereas organic orange EOs contained only 4 pesticides and a total concentration of 0.087 mg L−1 [27]. Organophosphorus and organochlorine pesticide residues were found in citrus EOs, especially those produced by cold-pressing and conventional agriculture practice, albeit with a concentration lower than 1 mg L−1. Tebuconazole and propiconazole co-distillated in peppermint EO with a different degree depending on the vapour pressure [289]. Cymoxanyl, dimethoate, and tebuconazole residues exceeded the maximum residue level set by the European Union in thyme samples from Poland [288]. In conventional grape skin extracts, Boscalid, Fludioxinil, Mycobutanil, and Pyraclostrobin levels exceeded the maximum residue level. However, the concentration of these crop-protection products was lower than the detection limit or the maximum residue level in organic grape skin extracts [290].
6.2. Mycotoxins
Mycotoxins can contaminate plant extracts. Different studies evaluated the mycotoxin contamination of plants such as Mentha sp. and Zingiber officinale R., which are generally used to produce EOs. Although the plant material showed levels of mycotoxins within the EU regulation limits, in some cases, the aflatoxin concentration exceeded acceptable standards [26]. In addition, the fungal contamination and mycotoxin accumulation in Z. officinale R. decreased the bioactive compounds of ginger [291]. Dried thyme herbs from Lebanon showed 75% of samples exceeding the limit of aflatoxin B1 for spices according to the European regulation. Similarly, the OTA level exceeded the maximum limits for Lebanese thyme and thyme mixes in 13% of the samples [292]. Zearalenone and deoxynivalenol contamination (range 10–209 µg kg−1) was detected in thyme samples from Poland [288]. Additional data are required to monitor the mycotoxin contamination of plant extracts (e.g., essential oils, phenolic extracts) and to establish regulatory limits.
6.3. Regulation
Comprehensive toxicological studies are necessary to obtain the approval of plant antimicrobials as food preservatives by the European Food Safety Authority (EFSA), the Food and Drug Administration (FDA), and the China Food Additives Association (CFAA). Many plant antimicrobials have the GRAS status for specific food applications, but their use in other food applications is not expressly approved. Indeed, plant phenolics are actually absent in the positive list of food preservatives. Regulatory authorities’ approval of plant extracts as food additives is essential to ensure consumer safety and confidence. Such authorization must be based on comprehensive safety assessments, including toxicological studies, exposure assessments, and evaluations of factors such as purity, stability, and potential allergenicity. Regulatory approval helps to guarantee that these plant extracts are safe for human consumption and meet specific quality standards. However, there can be challenges and barriers in the regulatory approval process. The complexity of plant antimicrobial mixtures and the need for extensive safety data may lead to time-consuming and costly approval processes. Global policy inequities significantly impact the approval of plant extracts as food additives. Addressing these inequities requires international cooperation, knowledge sharing, and capacity building to develop robust regulatory frameworks, protect and promote traditional knowledge, enhance research capacity in developing countries, and ensure fair and equitable access to the global market for plant-derived products. In order to improve and expedite the approval of plant extracts as food additives by authorities, several actions could be taken: encourage international collaboration and communication to harmonize regulations, establishing consistent guidelines and standards, increase investment in research and development to generate robust scientific data on plant extracts’ safety, efficacy, and potential applications, strengthen developing countries’ research and regulatory capacities through training, resources, and technical assistance, facilitate knowledge sharing and collaboration among various stakeholders, including researchers, industry professionals, and regulatory authorities, and employ advanced technologies such as data science for data-driven decision-making in the approval process. In turn, this will ensure consumer safety, promote the sustainable use of plant resources, and contribute to the growth of the global market for plant-derived products.
7. Conclusions and Future Directions
Plant antimicrobials gained considerable attention as promising alternatives to synthetic preservatives in the food industry, offering numerous benefits such as enhanced safety, extended shelf life, and increased consumer acceptance. The observed additive or synergistic effects between plant extracts, essential oils, and their compounds, and the successful integration of hurdle technologies, contributed to the scouting of novel and mild food preservation methods. Despite the significant progress made in the field, several research areas warrant further investigation. A deeper understanding of modes of action of plant antimicrobials and their combinations, including their effects at molecular and cellular levels on target microorganisms, is crucial for optimizing their application and enhancing their efficacy. The development of optimized formulations and delivery systems, such as nano-emulsions, encapsulation, or edible coatings, will improve stability, bioavailability, and targeted delivery of plant antimicrobials and, as a consequence, will increase their use in various food systems. A thorough examination of the impact of plant antimicrobials on the sensory properties of food is important to ensure consumer acceptance, focusing on minimizing adverse effects on taste, aroma, and texture while maintaining antimicrobial effectiveness.
Conducting comprehensive toxicological studies on plant antimicrobials, their derivatives, and combinations is essential to establish safe consumption levels and guarantee consumer safety. Additionally, addressing the challenges and barriers to regulatory approval for plant antimicrobials as food additives will accelerate their broader application in the food industry. The development of sustainable and environmentally friendly methods for extracting and producing plant antimicrobials aligns with the overall sustainability goals of the food industry. Given the knowledge on the chemical composition of plant extracts and the antimicrobial activity of plant compounds, plant antimicrobials could be recovered from agri-food by-products or from food waste, creating new value chains. Moreover, since non-edible wild plant species are often a good source of plant antimicrobial compounds, these species could be cultivated in marginal areas, promoting new cultivation practices in depressed rural territories, and also increasing the sustainable production of plant additives for the food industry. Increasing consumer awareness of plant antimicrobials’ benefits and their role in food safety and preservation will enhance market acceptance and drive demand for such products.
The description of the results allows us to decontextualize results reported in single studies for a wider comprehension. The analysis of studies suggests that the application of plant antimicrobials, also when combined with other technologies, still needs to overcome some critical aspects. As with all reviews, this study reported a selection of case studies among many others and some of the drawbacks highlighted herein are actually under examination.
In conclusion, we believe that this review provided useful updated information promoting a scientific evidence-based approach for researchers, aimed to understand if the use of plant antimicrobials can be scaled up from laboratory trials towards industrial applications. Further research and development efforts in these areas will help to overcome the current challenges and pave the way towards a widespread adoption of plant antimicrobials in the food industry, contributing to safer, more sustainable, and consumer-friendly foods.
Author Contributions
Conceptualization, L.P. and J.F.A.-Z.; writing—original draft preparation, L.P., M.R.T.-R. and J.F.A.-Z.; writing—review and editing, L.P., F.B. and J.F.A.-Z.; visualization, L.P. and M.R.T.-R.; supervision, J.F.A.-Z. All authors have read and agreed to the published version of the manuscript.
Funding
The authors greatly acknowledge the Project “Eco-sustainable flexible plastic films with high breathability endowed with antibacterial activity for vegetable foods—ECOATTIVE (Ministerial decree n. 0002441/28-07-2022, CUP: B29J21032720008—COR: 9200716)” and Carmela Magno (Vibac S.p.a, Ticineto (AL), Italy) for the stimulating discussion and useful suggestions.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bouarab Chibane, L.; Degraeve, P.; Ferhout, H.; Bouajila, J.; Oulahal, N. Plant antimicrobial polyphenols as potential natural food preservatives. J. Sci. Food Agric. 2019, 99, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Olszewska, M.A.; Gędas, A.; Simões, M. Antimicrobial polyphenol-rich extracts: Applications and limitations in the food industry. Food Res. Int. 2020, 134, 109214. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A.; Georgescu, C.; Turcuş, V.; Olah, N.K.; Mathe, E. An overview of natural antimicrobials role in food. Eur. J. Med. Chem. 2018, 143, 922–935. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Microbial spoilage of foods: Fundamentals. In The Microbiological Quality of Food; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Sawston, UK, 2017; pp. 1–21. [Google Scholar] [CrossRef]
- Joardder, M.U.; Masud, M.H. Food Preservation in Developing Countries: Challenges and Solutions; Springer: Berlin, Germany, 2019; pp. 1–239. [Google Scholar]
- McClements, D.J.; Das, A.K.; Dhar, P.; Nanda, P.K.; Chatterjee, N. Nanoemulsion-based technologies for delivering natural plant-based antimicrobials in foods. Front. Sustain. Food Syst. 2021, 5, 643208. [Google Scholar] [CrossRef]
- Favela-González, K.M.; Hernández-Almanza, A.Y.; De la Fuente-Salcido, N.M. The value of bioactive compounds of cruciferous vegetables (Brassica) as antimicrobials and antioxidants: A review. J. Food Biochem. 2020, 44, e13414. [Google Scholar] [CrossRef] [PubMed]
- Saeed, F.; Afzaal, M.; Tufail, T.; Ahmad, A. Use of natural antimicrobial agents: A safe preservation approach. In Active Antimicrobial Food Packaging; Var, I., Uzunlu, S., Eds.; IntechOpen: London, UK, 2019; Volume 18, pp. 7–24. [Google Scholar] [CrossRef]
- EFSA Panel on Food Additives and Nutrient Sources Added to Food (EFSA ANS Panel); Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; et al. Refined Exposure Assessment of Extracts of Rosemary (E 392) from Its Use as Food Additive. EFSA J. 2018, 16, e05373. [Google Scholar]
- Christopoulou, S.D.; Androutsopoulou, C.; Hahalis, P.; Kotsalou, C.; Vantarakis, A.; Lamari, F.N. Rosemary extract and essential oil as drink ingredients: An evaluation of their chemical composition, genotoxicity, antimicrobial, antiviral, and antioxidant properties. Foods 2021, 10, 3143. [Google Scholar] [CrossRef]
- Rathod, N.B.; Ranveer, R.C.; Benjakul, S.; Kim, S.K.; Pagarkar, A.U.; Patange, S.; Ozogul, F. Recent developments of natural antimicrobials and antioxidants on fish and fishery food products. Compr. Rev. Food Sci. Food Saf. 2021, 20, 4182–4210. [Google Scholar] [CrossRef]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant polyphenols as antioxidant and antibacterial agents for shelf-life extension of meat and meat products: Classification, structures, sources, and action mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef]
- Pateiro, M.; Munekata, P.E.; Sant’Ana, A.S.; Domínguez, R.; Rodríguez-Lázaro, D.; Lorenzo, J.M. Application of essential oils as antimicrobial agents against spoilage and pathogenic microorganisms in meat products. Int. J. Food Microbiol. 2021, 337, 108966. [Google Scholar] [CrossRef]
- Patrignani, F.; Siroli, L.; Serrazanetti, D.I.; Gardini, F.; Lanciotti, R. Innovative strategies based on the use of essential oils and their components to improve safety, shelf-life and quality of minimally processed fruits and vegetables. Trends Food Sci. Technol. 2015, 46, 311–319. [Google Scholar] [CrossRef]
- Ceruso, M.; Clement, J.A.; Todd, M.J.; Zhang, F.; Huang, Z.; Anastasio, A.; Pepe, T.; Liu, Y. The inhibitory effect of plant extracts on growth of the foodborne pathogen, Listeria monocytogenes. Antibiotics 2020, 9, 319. [Google Scholar] [CrossRef] [PubMed]
- Kalogianni, A.I.; Lazou, T.; Bossis, I.; Gelasakis, A.I. Natural phenolic compounds for the control of oxidation, bacterial spoilage, and foodborne pathogens in meat. Foods 2020, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Xie, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. The inhibitory effect of plant essential oils on foodborne pathogenic bacteria in food. Crit. Rev. Food Sci. Nutr. 2019, 59, 3281–3292. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Makhuvele, R.; Naidu, K.; Gbashi, S.; Thipe, V.C.; Adebo, O.A.; Njobeh, P.B. The use of plant extracts and their phytochemicals for control of toxigenic fungi and mycotoxins. Heliyon 2020, 6, e05291. [Google Scholar] [CrossRef] [PubMed]
- Sakarikou, C.; Kostoglou, D.; Simões, M.; Giaouris, E. Exploitation of plant extracts and phytochemicals against resistant Salmonella spp. in biofilms. Food Res. Int. 2020, 128, 108806. [Google Scholar] [CrossRef]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathogen. 2020, 142, 104056. [Google Scholar] [CrossRef]
- Somrani, M.; Inglés, M.-C.; Debbabi, H.; Abidi, F.; Palop, A. Garlic, onion, and cinnamon essential oil anti-biofilms’ effect against Listeria monocytogenes. Foods 2020, 9, 567. [Google Scholar] [CrossRef]
- Castro-Rosas, J.; Ferreira-Grosso, C.R.; Gómez-Aldapa, C.A.; Rangel-Vargas, E.; Rodríguez-Marín, M.L.; Guzmán-Ortiz, F.A.; Falfan-Cortes, R.N. Recent advances in microencapsulation of natural sources of antimicrobial compounds used in food—A review. Food Res. Int. 2017, 102, 575–587. [Google Scholar] [CrossRef]
- Pinto, L.; Bonifacio, M.A.; De Giglio, E.; Santovito, E.; Cometa, S.; Bevilacqua, A.; Baruzzi, F. Biopolymer hybrid materials: Development, characterization, and food packaging applications. Food Packag. Shelf Life 2021, 28, 100676. [Google Scholar] [CrossRef]
- Iordache, A.M.; Nechita, C.; Voica, C.; Roba, C.; Botoran, O.R.; Ionete, R.E. Assessing the health risk and the metal content of thirty-four plant essential oils using the ICP-MS technique. Nutrients 2022, 14, 2363. [Google Scholar] [CrossRef] [PubMed]
- Ałtyn, I.; Twarużek, M. Mycotoxin contamination concerns of herbs and medicinal plants. Toxins 2020, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Fillâtre, Y.; Gray, F.X.; Roy, C. Pesticides in essential oils: Occurrence and concentration in organic and conventional orange essential oils from eleven geographical origins. Anal. Chim. Acta 2017, 992, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Current understanding of modes of action of multicomponent bioactive phytochemicals: Potential for nutraceuticals and antimicrobials. Annu. Rev. Food Sci. Technol. 2022, 13, 337–359. [Google Scholar] [CrossRef]
- Gutiérrez-del-Río, I.; Fernández, J.; Lombó, F. Plant nutraceuticals as antimicrobial agents in food preservation: Terpenoids, polyphenols and thiols. Int. J. Antimicrob. Agents 2018, 52, 309–315. [Google Scholar] [CrossRef]
- Villalobos-Delgado, L.H.; Nevárez-Moorillon, G.V.; Caro, I.; Quinto, E.J.; Mateo, J. Natural antimicrobial agents to improve foods shelf-life. In Food Quality and Shelf-Life; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 125–157. [Google Scholar] [CrossRef]
- Bae, J.Y.; Seo, Y.H.; Oh, S.W. Antibacterial activities of polyphenols against foodborne pathogens and their application as antibacterial agents. Food Sci. Biotechnol. 2022, 31, 985–997. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, H.X.; Chen, J.Y.; Xue, Y.S.; Kodirkhonov, B.; Han, B.Z. Comparative study on inhibitory effects of ferulic acid and p-coumaric acid on Salmonella Enteritidis biofilm formation. World J. Microbiol. Biotechnol. 2022, 38, 136. [Google Scholar] [CrossRef]
- Li, J.; Zhao, N.; Xu, R.; Li, G.; Dong, H.; Wang, B.; Li, Z.; Fan, M.; Wei, X. Deciphering the antibacterial activity and mechanism of p-coumaric acid against Alicyclobacillus acidoterrestris and its application in apple juice. Int. J. Food Microbiol. 2022, 378, 109822. [Google Scholar] [CrossRef]
- Shi, Y.G.; Zhu, Y.J.; Shao, S.Y.; Zhang, R.R.; Wu, Y.; Zhu, C.M.; Liang, X.R.; Cai, W.Q. Alkyl ferulate esters as multifunctional food additives: Antibacterial activity and mode of action against Escherichia coli in vitro. J. Agric. Food Chem. 2018, 66, 12088–12101. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.G.; Zhang, R.R.; Zhu, C.M.; Liang, X.R.; Ettelaie, R.; Jiang, L.; Lin, S. On the mechanism behind enhanced antibacterial activity of alkyl gallate esters against foodborne pathogens and its application in Chinese icefish preservation. Food Microbiol. 2021, 99, 103817. [Google Scholar] [CrossRef] [PubMed]
- Patzke, H.; Schieber, A. Growth-inhibitory activity of phenolic compounds applied in an emulsifiable concentrate-ferulic acid as a natural pesticide against Botrytis cinerea. Food Res. Int. 2018, 113, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, W.; Li, Q.; Wang, L.; Cao, J.; Jiang, W. Defense responses, induced by p-coumaric acid and methyl p-coumarate, of jujube (Ziziphus jujuba Mill.) fruit against black spot rot caused by Alternaria alternata. J. Agric. Food Chem. 2019, 67, 2801–2810. [Google Scholar] [CrossRef]
- Li, W.; Yuan, S.; Sun, J.; Li, Q.; Jiang, W.; Cao, J. Ethyl p-coumarate exerts antifungal activity in vitro and in vivo against fruit Alternaria alternata via membrane-targeted mechanism. Int. J. Food Microbiol. 2018, 278, 26–35. [Google Scholar] [CrossRef]
- Vestergaard, M.; Ingmer, H. Antibacterial and antifungal properties of resveratrol. Int. J. Antimicrob. Agents 2019, 53, 716–723. [Google Scholar] [CrossRef]
- Cai, X.; Qi, J.; Xu, Z.; Huang, L.; Li, Y.; Ren, X.; Kong, Q. Three stilbenes make difference to the antifungal effects on ochratoxin A and its precursor production of Aspergillus carbonarius. Food Microbiol. 2022, 103, 103967. [Google Scholar] [CrossRef]
- Puljula, E.; Walton, G.; Woodward, M.J.; Karonen, M. Antimicrobial activities of ellagitannins against Clostridiales perfringens, Escherichia coli, Lactobacillus plantarum and Staphylococcus aureus. Molecules 2020, 25, 3714. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Y.; Jia, Y.; Zhang, M.; Huang, Y.; Li, C.; Li, K. Persimmon oligomeric proanthocyanidins exert antibacterial activity through damaging the cell membrane and disrupting the energy metabolism of Staphylococcus aureus. ACS Food Sci. Technol. 2020, 1, 35–44. [Google Scholar] [CrossRef]
- Peng, M.; Jiang, C.; Jing, H.; Du, X.; Fan, X.; Zhang, Y.; Wang, H. Comparison of different extraction methods on yield, purity, antioxidant, and antibacterial activities of proanthocyanidins from chokeberry (Aronia melanocarpa). J. Food Meas. Charact. 2022, 16, 2049–2059. [Google Scholar] [CrossRef]
- Wang, G.; Pang, T.; Xia, Y.; Liu, X.; Li, S.; Parvez, A.M.; Kong, F.; Si, C. Subdivision of bamboo kraft lignin by one-step ethanol fractionation to enhance its water-solubility and antibacterial performance. Int. J. Biol. Macromol. 2019, 133, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; Zhou, M.; Liu, Z.; Chen, Y.; Li, R. Inhibition effects of low concentrations of epigallocatechin gallate on the biofilm formation and hemolytic activity of Listeria monocytogenes. Food Control 2018, 85, 119–126. [Google Scholar] [CrossRef]
- Xiong, L.G.; Chen, Y.J.; Tong, J.W.; Huang, J.A.; Li, J.; Gong, Y.S.; Liu, Z.H. Tea polyphenol epigallocatechin gallate inhibits Escherichia coli by increasing endogenous oxidative stress. Food Chem. 2017, 217, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial efficacy of plant phenolic compounds against Salmonella and Escherichia coli. Food Biosci. 2015, 11, 8–16. [Google Scholar] [CrossRef]
- Amin, M.U.; Khurram, M.; Khattak, B.; Khan, J. Antibiotic additive and synergistic action of rutin, morin and quercetin against methicillin resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2015, 15, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, L.; Zhang, L.; Guo, Y.; Qi, X.; He, L. Effects of quercetin on postharvest blue mold control in kiwifruit. Sci. Hortic. 2018, 228, 18–25. [Google Scholar] [CrossRef]
- Li, X.M.; Li, Z.Y.; Wang, Y.D.; Wang, J.Q.; Yang, P.L. Quercetin inhibits the proliferation and aflatoxins biosynthesis of Aspergillus flavus. Toxins 2019, 11, 154. [Google Scholar] [CrossRef]
- Cha, J.D.; Moon, S.E.; Kim, J.Y.; Jung, E.K.; Lee, Y.S. Antibacterial activity of sophoraflavanone G isolated from the roots of Sophora flavescens against methicillin-resistant Staphylococcus aureus. Phytother. Res. 2009, 23, 1326–1331. [Google Scholar] [CrossRef]
- Kalli, S.; Araya-Cloutier, C.; Chapman, J.; Sanders, J.W.; Vincken, J.P. Prenylated (iso) flavonoids as antifungal agents against the food spoiler Zygosaccharomyces parabailii. Food Control 2022, 132, 108434. [Google Scholar] [CrossRef]
- Batiha, G.E.S.; Hussein, D.E.; Algammal, A.M.; George, T.T.; Jeandet, P.; Al-Snafi, A.E.; Tiwari, A.; Pagnossa, G.P.; Lima, C.M.; Thorat, N.D.; et al. Application of natural antimicrobials in food preservation: Recent views. Food Control 2021, 126, 108066. [Google Scholar] [CrossRef]
- Bajalan, I.; Rouzbahani, R.; Pirbalouti, A.G.; Maggi, F. Antioxidant and antibacterial activities of the essential oils obtained from seven Iranian populations of Rosmarinus officinalis. Ind. Crops Prod. 2017, 107, 305–311. [Google Scholar] [CrossRef]
- Rathore, S.; Mukhia, S.; Kapoor, S.; Bhatt, V.; Kumar, R.; Kumar, R. Seasonal variability in essential oil composition and biological activity of Rosmarinus officinalis L. accessions in the western Himalaya. Sci. Rep. 2022, 12, 3305. [Google Scholar] [CrossRef] [PubMed]
- Micić, D.; Đurović, S.; Riabov, P.; Tomić, A.; Šovljanski, O.; Filip, S.; Tosti, T.; Dojčinović, B.; Božović, R.; Jovanović, D.; et al. Rosemary essential oils as a promising source of bioactive compounds: Chemical composition, thermal properties, biological activity, and gastronomical perspectives. Foods 2021, 10, 2734. [Google Scholar] [CrossRef] [PubMed]
- Dammak, I.; Hamdi, Z.; El Euch, S.K.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia officinalis, Lavandula dentata and Laurus nobilis essential oils and a major monoterpene constituent 1, 8-cineole against Aspergillus carbonarius. Ind. Crops Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Sakkas, H.; Economou, V.; Gousia, P.; Bozidis, P.; Sakkas, V.A.; Petsios, S.; Mpekoulis, G.; Ilia, A.; Papadopoulou, C. Antibacterial efficacy of commercially available essential oils tested against drug-resistant Gram-positive pathogens. Appl. Sci. 2018, 8, 2201. [Google Scholar] [CrossRef]
- Valdivieso-Ugarte, M.; Plaza-Diaz, J.; Gomez-Llorente, C.; Gómez, E.L.; Sabés-Alsina, M.; Gil, Á. In vitro examination of antibacterial and immunomodulatory activities of cinnamon, white thyme, and clove essential oils. J. Funct. Foods 2021, 81, 104436. [Google Scholar] [CrossRef]
- Reyes-Jurado, F.; Cervantes-Rincón, T.; Bach, H.; López-Malo, A.; Palou, E. Antimicrobial activity of Mexican oregano (Lippia berlandieri), thyme (Thymus vulgaris), and mustard (Brassica nigra) essential oils in gaseous phase. Ind. Crops Prod. 2019, 131, 90–95. [Google Scholar] [CrossRef]
- Císarová, M.; Hleba, L.; Medo, J.; Tančinová, D.; Mašková, Z.; Čuboň, J.; Kováčik, A.; Foltinová, D.; Božike, M.; Klouček, P. The in vitro and in situ effect of selected essential oils in vapour phase against bread spoilage toxicogenic aspergilli. Food Control 2020, 110, 107007. [Google Scholar] [CrossRef]
- Marchese, A.; Arciola, C.R.; Barbieri, R.; Silva, A.S.; Nabavi, S.F.; Tsetegho Sokeng, A.J.; Izadi, M.; Jafari, N.J.; Suntar, I.; Daglia, M.; et al. Update on monoterpenes as antimicrobial agents: A particular focus on p-cymene. Materials 2017, 10, 947. [Google Scholar] [CrossRef]
- Pinto, L.; Bonifacio, M.A.; De Giglio, E.; Cometa, S.; Logrieco, A.F.; Baruzzi, F. Unravelling the antifungal effect of red thyme oil (Thymus vulgaris L.) compounds in vapor phase. Molecules 2020, 25, 4761. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, L.F.; Xu, J.G.; Hu, Q.P. Comparison study on antioxidant, DNA damage protective and antibacterial activities of eugenol and isoeugenol against several foodborne pathogens. Food Nutr. Res. 2017, 61, 1353356. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove essential oil as an alternative approach to control postharvest blue mold caused by Penicillium italicum in citrus fruit. Biomolecules 2019, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Cava-Roda, R.; Taboada-Rodríguez, A.; López-Gómez, A.; Martínez-Hernández, G.B.; Marín-Iniesta, F. Synergistic antimicrobial activities of combinations of vanillin and essential oils of cinnamon bark, cinnamon leaves, and cloves. Foods 2021, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- Yoplac, I.; Vargas, L.; Robert, P.; Hidalgo, A. Characterization and antimicrobial activity of microencapsulated citral with dextrin by spray drying. Heliyon 2021, 7, e06737. [Google Scholar] [CrossRef]
- Zheng, S.; Jing, G.; Wang, X.; Ouyang, Q.; Jia, L.; Tao, N. Citral exerts its antifungal activity against Penicillium digitatum by affecting the mitochondrial morphology and function. Food Chem. 2015, 178, 76–81. [Google Scholar] [CrossRef]
- Tang, X.; Shao, Y.L.; Tang, Y.J.; Zhou, W.W. Antifungal activity of essential oil compounds (geraniol and citral) and inhibitory mechanisms on grain pathogens (Aspergillus flavus and Aspergillus ochraceus). Molecules 2018, 23, 2108. [Google Scholar] [CrossRef]
- Quintieri, L.; Fancello, F.; Caputo, L.; Sorrentino, A.; Zara, S.; Lippolis, V.; Cervellieri, S.; Fanelli, F.; Corvino, A.; Pace, B.; et al. Effect of gaseous citral on table grapes contaminated by Rhizopus oryzae ITEM 18876. Foods 2022, 11, 2478. [Google Scholar] [CrossRef]
- Siddiqua, S.; Anusha, B.A.; Ashwini, L.S.; Negi, P.S. Antibacterial activity of cinnamaldehyde and clove oil: Effect on selected foodborne pathogens in model food systems and watermelon juice. J. Food Sci. Technol. 2015, 52, 5834–5841. [Google Scholar] [CrossRef]
- Sun, Q.; Li, J.; Sun, Y.; Chen, Q.; Zhang, L.; Le, T. The antifungal effects of cinnamaldehyde against Aspergillus niger and its application in bread preservation. Food Chem. 2020, 317, 126405. [Google Scholar] [CrossRef]
- Xu, L.; Tao, N.; Yang, W.; Jing, G. Cinnamaldehyde damaged the cell membrane of Alternaria alternata and induced the degradation of mycotoxins in vivo. Ind. Crops Prod. 2018, 112, 427–433. [Google Scholar] [CrossRef]
- Song, H.J.; Ku, K.M. Optimization of allyl isothiocyanate sanitizing concentration for inactivation of Salmonella typhimurium on lettuce based on its phenotypic and metabolome changes. Food Chem. 2021, 364, 130438. [Google Scholar] [CrossRef] [PubMed]
- Olaimat, A.N.; Al-Holy, M.A.; Ghoush, M.A.; Al-Nabulsi, A.A.; Holley, R.A. Control of Salmonella enterica and Listeria monocytogenes in hummus using allyl isothiocyanate. Int. J. Food Microbiol. 2018, 278, 73–80. [Google Scholar] [CrossRef]
- de Melo Nazareth, T.; Alonso-Garrido, M.; Stanciu, O.; Mañes, J.; Manyes, L.; Meca, G. Effect of allyl isothiocyanate on transcriptional profile, aflatoxin synthesis, and Aspergillus flavus growth. Food Res. Int. 2020, 128, 108786. [Google Scholar] [CrossRef] [PubMed]
- de Melo Nazareth, T.; Quiles, J.M.; Torrijos, R.; Luciano, F.B.; Manes, J.; Meca, G. Antifungal and antimycotoxigenic activity of allyl isothiocyanate on barley under different storage conditions. LWT 2019, 112, 108237. [Google Scholar] [CrossRef]
- Yang, C.X.; Wu, H.T.; Li, X.X.; Wu, H.Y.; Niu, T.X.; Wang, X.N.; Lian, R.; Zhang, G.L.; Hou, H.M. Comparison of the inhibitory potential of benzyl isothiocyanate and phenethyl isothiocyanate on Shiga toxin-producing and enterotoxigenic Escherichia coli. LWT 2020, 118, 108806. [Google Scholar] [CrossRef]
- Wu, H.Y.; Xu, Y.H.; Wei, L.N.; Bi, J.R.; Hou, H.M.; Hao, H.S.; Zhang, G.L. Inhibitory effects of 3-(methylthio) propyl isothiocyanate in comparison with benzyl isothiocyanate on Listeria monocytogenes. J. Food Meas. Charact. 2022, 16, 1768–1775. [Google Scholar] [CrossRef]
- Yang, B.; Li, L.; Geng, H.; Zhang, C.; Wang, G.; Yang, S.; Gao, S.; Zhao, Y.; Xing, F. Inhibitory effect of allyl and benzyl isothiocyanates on ochratoxin a producing fungi in grape and maize. Food Microbiol. 2021, 100, 103865. [Google Scholar] [CrossRef]
- Nowicki, D.; Maciąg-Dorszyńska, M.; Bogucka, K.; Szalewska-Pałasz, A.; Herman-Antosiewicz, A. Various modes of action of dietary phytochemicals, sulforaphane and phenethyl isothiocyanate, on pathogenic bacteria. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Bi, Y.; Wang, T.; Dong, Y.; Yang, Q.; Zhang, T. 2-Phenylethyl isothiocyanate exerts antifungal activity against Alternaria alternata by affecting membrane integrity and mycotoxin production. Toxins 2020, 12, 124. [Google Scholar] [CrossRef]
- Wu, T.L.; Hu, Y.M.; Sun, Y.; Zhang, Z.J.; Wu, Z.R.; Zhao, W.B.; Tang, C.; Du, S.S.; He, Y.H.; Ma, Y.; et al. Insights into the mode of action of 2-(4-methoxyphenyl) ethyl isothiocyanate on Aspergillus niger. Food Control 2022, 136, 108871. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- El-Zahar, K.M.; Al-Jamaan, M.E.; Al-Mutairi, F.R.; Al-Hudiab, A.M.; Al-Einzi, M.S.; Mohamed, A.A.Z. Antioxidant, antibacterial, and antifungal activities of the ethanolic extract obtained from Berberis vulgaris roots and leaves. Molecules 2022, 27, 6114. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Zhe, T.; Li, F.; Zhang, Y.; Yu, M.; Li, R.; Wang, L. Sustainable films containing AIE-active berberine-based nanoparticles: A promising antibacterial food packaging. Food Hydrocoll. 2022, 123, 107147. [Google Scholar] [CrossRef]
- He, M.; Wang, Y.; Hong, M.; Li, T. Berberine as a promising natural compound to control Penicillium italicum causing blue mold of citrus fruit. Sci. Hortic. 2022, 305, 111370. [Google Scholar] [CrossRef]
- Marchese, A.; Barbieri, R.; Sanches-Silva, A.; Daglia, M.; Nabavi, S.F.; Jafari, N.J.; Izadi, M.; Ajami, M.; Nabavi, S.M. Antifungal and antibacterial activities of allicin: A review. Trends Food Sci. Technol. 2016, 52, 49–56. [Google Scholar] [CrossRef]
- Jin, Z.; Li, L.; Zheng, Y.; An, P. Diallyl disulfide, the antibacterial component of garlic essential oil, inhibits the toxicity of Bacillus cereus ATCC 14579 at sub-inhibitory concentrations. Food Control 2021, 126, 108090. [Google Scholar] [CrossRef]
- Tang, Y.; Li, F.; Gu, D.; Wang, W.; Huang, J.; Jiao, X. Antimicrobial effect and the mechanism of diallyl trisulfide against Campylobacter jejuni. Antibiotics 2021, 10, 246. [Google Scholar] [CrossRef]
- Pernin, A.; Guillier, L.; Dubois-Brissonnet, F. Inhibitory activity of phenolic acids against Listeria monocytogenes: Deciphering the mechanisms of action using three different models. Food Microbiol. 2019, 80, 18–24. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, X.; Wu, H.; Wang, H.; Bian, H.; Zhu, Y.; Xu, W.; Liu, F.; Wang, D.; Fu, L. Antibacterial activity and action mode of chlorogenic acid against Salmonella enteritidis, a foodborne pathogen in chilled fresh chicken. World J. Microbiol. Biotechnol. 2020, 36, 24. [Google Scholar] [CrossRef]
- Ansari, M.A.; Anurag, A.; Fatima, Z.; Hameed, S. Natural phenolic compounds: A potential antifungal agent. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education, 1st ed.; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 1189–1195. [Google Scholar]
- Morales, J.; Mendoza, L.; Cotoras, M. Alteration of oxidative phosphorylation as a possible mechanism of the antifungal action of p-coumaric acid against Botrytis cinerea. J. Appl. Microbiol. 2017, 123, 969–976. [Google Scholar] [CrossRef]
- Zhu, C.; Lei, M.; Andargie, M.; Zeng, J.; Li, J. Antifungal activity and mechanism of action of tannic acid against Penicillium digitatum. Physiol. Mol. Plant Pathol. 2019, 107, 46–50. [Google Scholar] [CrossRef]
- You, S.; Xie, Y.; Zhuang, X.; Chen, H.; Qin, Y.; Cao, J.; Lan, T. Effect of high antioxidant activity on bacteriostasis of lignin from sugarcane bagasse. Biochem. Eng. J. 2022, 180, 108335. [Google Scholar] [CrossRef]
- Luo, J.; Xu, F.; Zhang, X.; Shao, X.; Wei, Y.; Wang, H. Transcriptome analysis of Penicillium italicum in response to the flavonoids from Sedum aizoon L. World J. Microbiol. Biotechnol. 2020, 36, 62. [Google Scholar] [CrossRef] [PubMed]
- Churklam, W.; Chaturongakul, S.; Ngamwongsatit, B.; Aunpad, R. The mechanisms of action of carvacrol and its synergism with nisin against Listeria monocytogenes on sliced bologna sausage. Food Control 2020, 108, 106864. [Google Scholar] [CrossRef]
- Stratakos, A.C.; Sima, F.; Ward, P.; Linton, M.; Kelly, C.; Pinkerton, L.; Stef, L.; Pet, I.; Corcionivoschi, N. The in vitro effect of carvacrol, a food additive, on the pathogenicity of O157 and non-O157 Shiga-toxin producing Escherichia coli. Food Control 2018, 84, 290–296. [Google Scholar] [CrossRef]
- Barbosa, L.N.; Alves, F.C.B.; Andrade, B.F.M.T.; Albano, M.; Rall, V.L.M.; Fernandes, A.A.H.; Buzalaf, M.A.R.; de Lima Leite, A.; de Pontes, L.G.; dos Santos, L.D.; et al. Proteomic analysis and antibacterial resistance mechanisms of Salmonella enteritidis submitted to the inhibitory effect of Origanum vulgare essential oil, thymol and carvacrol. J. Proteomics 2020, 214, 103625. [Google Scholar] [CrossRef]
- Han, Y.; Sun, Z.; Chen, W. Antimicrobial susceptibility and antibacterial mechanism of limonene against Listeria monocytogenes. Molecules 2019, 25, 33. [Google Scholar] [CrossRef]
- Gupta, A.; Jeyakumar, E.; Lawrence, R. Strategic approach of multifaceted antibacterial mechanism of limonene traced in Escherichia coli. Sci. Rep. 2021, 11, 13816. [Google Scholar] [CrossRef]
- Han, Y.; Chen, W.; Sun, Z. Antimicrobial activity and mechanism of limonene against Staphylococcus aureus. J. Food Saf. 2021, 41, e12918. [Google Scholar] [CrossRef]
- Jeyakumar, G.E.; Lawrence, R. Mechanisms of bactericidal action of eugenol against Escherichia coli. J. Herb. Med. 2021, 26, 100406. [Google Scholar] [CrossRef]
- Bai, X.; Li, X.; Liu, X.; Xing, Z.; Su, R.; Wang, Y.; Xia, X.; Shi, C. Antibacterial effect of eugenol on Shigella flexneri and its mechanism. Foods 2022, 11, 2565. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, S.; Du, S.; Chen, S.; Sun, H. Antifungal activity of thymol and carvacrol against postharvest pathogens Botrytis cinerea. J. Food Sci. Technol. 2019, 56, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Li, Z.; Wang, X. UHPLC-HRMS-based untargeted lipidomics reveal mechanism of antifungal activity of carvacrol against Aspergillus flavus. Foods 2022, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Hu, M.; Zhang, Y.; Niu, C.; Yue, T.; Yuan, Y.; Wang, Z. Antifungal activity and mechanism of citral, limonene and eugenol against Zygosaccharomyces rouxii. LWT 2019, 106, 50–56. [Google Scholar] [CrossRef]
- Jiang, N.; Wang, L.; Jiang, D.; Wang, M.; Liu, H.; Yu, H.; Yao, W. Transcriptomic analysis of inhibition by eugenol of ochratoxin A biosynthesis and growth of Aspergillus carbonarius. Food Control 2022, 135, 108788. [Google Scholar] [CrossRef]
- Luciano, F.B.; Holley, R.A. Enzymatic inhibition by allyl isothiocyanate and factors affecting its antimicrobial action against Escherichia coli O157: H7. Int. J. Food Microbiol. 2009, 131, 240–245. [Google Scholar] [CrossRef]
- Li, P.; Zhao, Y.M.; Wang, C.; Zhu, H.P. Antibacterial activity and main action pathway of benzyl isothiocyanate extracted from papaya seeds. J. Food Sci. 2021, 86, 169–176. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Xu, Y.; Chen, T.; Li, B.; Zhang, Z.; Tian, S. Application and mechanism of benzyl-isothiocyanate, a natural antimicrobial agent from cruciferous vegetables, in controlling postharvest decay of strawberry. Postharvest Biol. Technol. 2021, 180, 111604. [Google Scholar] [CrossRef]
- Boberek, J.M.; Stach, J.; Good, L. Genetic evidence for inhibition of bacterial division protein FtsZ by berberine. PLoS ONE 2010, 5, e13745. [Google Scholar] [CrossRef]
- Xu, C.; Wang, F.; Huang, F.; Yang, M.; He, D.; Deng, L. Targeting effect of berberine on type I fimbriae of Salmonella typhimurium and its effective inhibition of biofilm. Appl. Microbiol. Biotechnol. 2021, 105, 1563–1573. [Google Scholar] [CrossRef]
- Arshad, M.S.; Batool, S.A. Natural antimicrobials, their sources and food safety. In Food Additives; Karunaratne, D.N., Pamunuwa, G., Eds.; IntechOpen: London, UK, 2017; Volume 87, pp. 87–104. [Google Scholar] [CrossRef]
- Belgacem, I.; Li Destri Nicosia, M.G.; Pangallo, S.; Abdelfattah, A.; Benuzzi, M.; Agosteo, G.E.; Schena, L. Pomegranate peel extracts as safe natural treatments to control plant diseases and increase the shelf-life and safety of fresh fruits and vegetables. Plants 2021, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Givi, F.; Gholami, M.; Massah, A. Application of pomegranate peel extract and essential oil as a safe botanical preservative for the control of postharvest decay caused by Penicillium italicum and Penicillium digitatum on “Satsuma” mandarin. J. Food Saf. 2019, 39, e12639. [Google Scholar] [CrossRef]
- Gómez-Maldonado, D.; Lobato-Calleros, C.; Aguirre-Mandujano, E.; Leyva-Mir, S.G.; Robles-Yerena, L.; Vernon-Carter, E.J. Antifungal activity of mango kernel polyphenols on mango fruit infected by anthracnose. LWT 2020, 126, 109337. [Google Scholar] [CrossRef]
- Hernández, A.; Ruiz-Moyano, S.; Galván, A.I.; Merchán, A.V.; Nevado, F.P.; Aranda, E.; Serradilla, M.J.; de Guía Córdoba, M.; Martín, A. Anti-fungal activity of phenolic sweet orange peel extract for controlling fungi responsible for post-harvest fruit decay. Fungal Biol. 2021, 125, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Wu, H.; Shi, F.; Wang, H.; Chen, K.; Feng, J.; Jia, W. Antifungal activity screening for mint and thyme essential oils against Rhizopus stolonifer and their application in postharvest preservation of strawberry and peach fruits. J. Appl. Microbiol. 2021, 130, 1993–2007. [Google Scholar] [CrossRef] [PubMed]
- Sumalan, R.M.; Kuganov, R.; Obistioiu, D.; Popescu, I.; Radulov, I.; Alexa, E.; Negrea, M.; Salimzoda, A.F.; Sumalan, R.L.; Cocan, I. Assessment of mint, basil, and lavender essential oil vapor-phase in antifungal protection and lemon fruit quality. Molecules 2020, 25, 1831. [Google Scholar] [CrossRef]
- Pinto, L.; Cefola, M.; Bonifacio, M.A.; Cometa, S.; Bocchino, C.; Pace, B.; De Giglio, E.; Palumbo, M.; Sada, A.; Logrieco, A.F.; et al. Effect of red thyme oil (Thymus vulgaris L.) vapours on fungal decay, quality parameters and shelf-life of oranges during cold storage. Food Chem. 2021, 336, 127590. [Google Scholar] [CrossRef]
- Arellano, S.; Zhu, L.; Dev Kumar, G.; Law, B.; Friedman, M.; Ravishankar, S. Essential oil microemulsions inactivate antibiotic-resistant bacteria on iceberg lettuce during 28-day storage at 4 °C. Molecules 2022, 27, 6699. [Google Scholar] [CrossRef]
- Kara, M.; Soylu, E.M. Assessment of glucosinolate-derived isothiocyanates as potential natural antifungal compounds against citrus sour rot disease agent Geotrichum citri-aurantii. J. Phytopathol. 2020, 168, 279–289. [Google Scholar] [CrossRef]
- Santos, M.I.S.; Marques, C.; Mota, J.; Pedroso, L.; Lima, A. Applications of essential oils as antibacterial agents in minimally processed fruits and vegetables—A review. Microorganisms 2022, 10, 760. [Google Scholar] [CrossRef]
- Wong, J.X.; Ramli, S.; Desa, S.; Chen, S.N. Use of Centella asiatica extract in reducing microbial contamination and browning effect in fresh cut fruits and vegetables during storage: A potential alternative of synthetic preservatives. LWT 2021, 151, 112229. [Google Scholar] [CrossRef]
- Wieczyńska, J.; Cavoski, I. Antimicrobial, antioxidant and sensory features of eugenol, carvacrol and trans-anethole in active packaging for organic ready-to-eat iceberg lettuce. Food Chem. 2018, 259, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Myszka, K.; Schmidt, M.T.; Majcher, M.; Juzwa, W.; Czaczyk, K. β-Caryophyllene-rich pepper essential oils suppress spoilage activity of Pseudomonas fluorescens KM06 in fresh-cut lettuce. LWT Food Sci. Technol. 2017, 83, 118–126. [Google Scholar] [CrossRef]
- Amiri, A.; Ramezanian, A.; Mortazavi, S.M.H.; Hosseini, S.M.H.; Yahia, E. Shelf-life extension of pomegranate arils using chitosan nanoparticles loaded with Satureja hortensis essential oil. J. Sci. Food Agric. 2021, 101, 3778–3786. [Google Scholar] [CrossRef]
- Awad, A.H.R.; Parmar, A.; Ali, M.R.; El-Mogy, M.M.; Abdelgawad, K.F. Extending the shelf-life of fresh-cut green bean pods by ethanol, ascorbic Acid, and essential oils. Foods 2021, 10, 1103. [Google Scholar] [CrossRef] [PubMed]
- Maleš, I.; Pedisić, S.; Zorić, Z.; Elez-Garofulić, I.; Repajić, M.; You, L.; Vladimir-Knežević, S.; Butorac, D.; Dragović-Uzelac, V. The medicinal and aromatic plants as ingredients in functional beverage production. J. Funct. Foods 2022, 96, 105210. [Google Scholar] [CrossRef]
- da Cruz Almeida, E.T.; de Souza, G.T.; de Sousa Guedes, J.P.; Barbosa, I.M.; de Sousa, C.P.; Castellano, L.R.C.; Magnani, M.; de Souza, E.L. Mentha piperita L. essential oil inactivates spoilage yeasts in fruit juices through the perturbation of different physiological functions in yeast cells. Food Microbiol. 2019, 82, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, H. Assessment of different antimicrobials to inhibit the growth of Zygosaccharomyces rouxii cocktail in concentrated apple juice. Food Microbiol. 2020, 91, 103549. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.; Beuchat, L.R.; Kim, Y.; Ryu, J.H. Synergistic antimicrobial activity of oregano and thyme thymol essential oils against Leuconostoc citreum in a laboratory medium and tomato juice. Food Microbiol. 2020, 90, 103489. [Google Scholar] [CrossRef]
- Fernandez, M.V.; Bengardino, M.; Jagus, R.J.; Agüero, M.V. Enrichment and preservation of a vegetable smoothie with an antioxidant and antimicrobial extract obtained from beet by-products. LWT 2020, 117, 108622. [Google Scholar] [CrossRef]
- Maqsood, S.; Abushelaibi, A.; Manheem, K.; Al Rashedi, A.; Kadim, I.T. Lipid oxidation, protein degradation, microbial and sensorial quality of camel meat as influenced by phenolic compounds. LWT Food Sci. Technol. 2015, 63, 953–959. [Google Scholar] [CrossRef]
- Nowak, A.; Czyzowska, A.; Efenberger, M.; Krala, L. Polyphenolic extracts of cherry (Prunus cerasus L.) and blackcurrant (Ribes nigrum L.) leaves as natural preservatives in meat products. Food Microbiol. 2016, 59, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, A.; Di Martino, V.; Ercolini, D.; Parente, E.; Villani, F. Antimicrobial activity of Myrtus communis L. water-ethanol extract against meat spoilage strains of Brochothrix thermosphacta and Pseudomonas fragi in vitro and in meat. Ann. Microbiol. 2015, 65, 841–850. [Google Scholar] [CrossRef]
- Tamkutė, L.; Gil, B.M.; Carballido, J.R.; Pukalskienė, M.; Venskutonis, P.R. Effect of cranberry pomace extracts isolated by pressurized ethanol and water on the inhibition of food pathogenic/spoilage bacteria and the quality of pork products. Food Res. Int. 2019, 120, 38–51. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J.; Guo, X. Effects of antimicrobial and antioxidant activities of spice extracts on raw chicken meat quality. Food Sci. Hum. Wellness 2016, 5, 39–48. [Google Scholar] [CrossRef]
- Shahbazi, Y.; Shavisi, N.; Mohebi, E. Effects of Ziziphora clinopodioides essential oil and nisin, both separately and in combination, to extend shelf-life and control Escherichia coli O157:H7 and Staphylococcus aureus in raw beef patty during refrigerated storage. J. Food Saf. 2016, 36, 227–236. [Google Scholar] [CrossRef]
- Karam, L.; Chehab, R.; Osaili, T.M.; Savvaidis, I.N. Antimicrobial effect of thymol and carvacrol added to a vinegar-based marinade for controlling spoilage of marinated beef (Shawarma) stored in air or vacuum packaging. Int. J. Food Microbiol. 2020, 332, 108769. [Google Scholar] [CrossRef] [PubMed]
- Olatunde, O.O.; Benjakul, S.; Huda, N.; Zhang, B.; Deng, S. Ethanolic Noni (Morinda citrifolia L.) leaf extract dechlorophyllised using sedimentation process: Antioxidant, antibacterial properties and efficacy in extending the shelf-life of striped catfish slices. Int. J. Food Sci. Technol. 2021, 56, 2804–2819. [Google Scholar] [CrossRef]
- Olatunde, O.O.; Della Tan, S.L.; Shiekh, K.A.; Benjakul, S.; Nirmal, N.P. Ethanolic guava leaf extracts with different chlorophyll removal processes: Anti-melanosis, antibacterial properties and the impact on qualities of Pacific white shrimp during refrigerated storage. Food Chem. 2021, 341, 128251. [Google Scholar] [CrossRef]
- Li, Y.; Zhuang, S.; Liu, Y.; Zhang, L.; Liu, X.; Cheng, H.; Liu, J.; Shu, R.; Luo, Y. Effect of grape seed extract on quality and microbiota community of container-cultured snakehead (Channa argus) fillets during chilled storage. Food Microbiol. 2020, 91, 103492. [Google Scholar] [CrossRef]
- Van Haute, S.; Raes, K.; Van Der Meeren, P.; Sampers, I. The effect of cinnamon, oregano and thyme essential oils in marinade on the microbial shelf-life of fish and meat products. Food Control 2016, 68, 30–39. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, D.; Lv, J.; Li, Q.; Kong, C.; Luo, Y. Effect of cinnamon essential oil on bacterial diversity and shelf-life in vacuum-packaged common carp (Cyprinus carpio) during refrigerated storage. Int. J. Food Microbiol. 2017, 249, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Roila, R.; Valiani, A.; Ranucci, D.; Ortenzi, R.; Servili, M.; Veneziani, G.; Branciari, R. Antimicrobial efficacy of a polyphenolic extract from olive oil by-product against “Fior di latte” cheese spoilage bacteria. Int. J. Food Microbiol. 2019, 295, 49–53. [Google Scholar] [CrossRef]
- Derbassi, N.; Pedrosa, M.C.; Heleno, S.; Fernandes, F.; Dias, M.I.; Calhelha, R.C.; Rodrigues, P.; Carocho, M.; Ferreira, I.C.F.R.; Barros, L. Arbutus unedo leaf extracts as potential dairy preservatives: Case study on quark cheese. Food Funct. 2022, 13, 5442–5454. [Google Scholar] [CrossRef] [PubMed]
- Milanović, V.; Sabbatini, R.; Garofalo, C.; Cardinali, F.; Pasquini, M.; Aquilanti, L.; Osimani, A. Evaluation of the inhibitory activity of essential oils against spoilage yeasts and their potential application in yogurt. Int. J. Food Microbiol. 2021, 341, 109048. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Molina, A.K.; Abreu, R.M.; Santo-Buelga, C.; Ferreira, I.C.; Pereira, C.; Barros, L. Natural food colorants and preservatives: A review, a demand, and a challenge. J. Agric. Food Chem. 2022, 70, 2789–2805. [Google Scholar] [CrossRef]
- Schirone, M.; Visciano, P.; Tofalo, R.; Suzzi, G. Foodborne pathogens: Hygiene and safety. Front. Microbiol. 2019, 10, 1974. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef]
- Guan, P.; Wang, X.; Dong, Z.; Song, M.; Zhu, H.; Suo, B. Cinnamaldehyde inactivates Listeria monocytogenes at a low temperature in ground pork by disturbing the expression of stress regulatory genes. Food Biosci. 2023, 51, 102277. [Google Scholar] [CrossRef]
- Bai, J.; Li, J.; Chen, Z.; Bai, X.; Yang, Z.; Wang, Z.; Yang, Y. Antibacterial activity and mechanism of clove essential oil against foodborne pathogens. LWT 2023, 173, 114249. [Google Scholar] [CrossRef]
- Sepahvand, S.; Amiri, S.; Radi, M.; Akhavan, H.R. Antimicrobial activity of thymol and thymol-nanoemulsion against three food-borne pathogens inoculated in a sausage model. Food Bioproc. Tech. 2021, 14, 1936–1945. [Google Scholar] [CrossRef]
- Saraiva, C.; Silva, A.C.; García-Díez, J.; Cenci-Goga, B.; Grispoldi, L.; Silva, A.F.; Almeida, J.M. Antimicrobial activity of Myrtus communis L. and Rosmarinus officinalis L. essential oils against Listeria monocytogenes in cheese. Foods 2021, 10, 1106. [Google Scholar] [CrossRef]
- Ahmed, L.I.; Ibrahim, N.; Abdel-Salam, A.B.; Fahim, K.M. Potential application of ginger, clove and thyme essential oils to improve soft cheese microbial safety and sensory characteristics. Food Biosci. 2021, 42, 101177. [Google Scholar] [CrossRef]
- Rossi, C.; Chaves-López, C.; Možina, S.S.; Di Mattia, C.; Scuota, S.; Luzzi, I.; Jenič, T.; Paparella, A.; Serio, A. Salmonella enterica adhesion: Effect of Cinnamomum zeylanicum essential oil on lettuce. LWT 2019, 111, 16–22. [Google Scholar] [CrossRef]
- Brnawi, W.I.; Hettiarachchy, N.S.; Horax, R.; Kumar-Phillips, G.; Ricke, S. Antimicrobial activity of leaf and bark cinnamon essential oils against Listeria monocytogenes and Salmonella typhimurium in broth system and on celery. J. Food Process. Preserv. 2019, 43, e13888. [Google Scholar] [CrossRef]
- Dai, J.; Li, C.; Cui, H.; Lin, L. Unraveling the anti-bacterial mechanism of Litsea cubeba essential oil against E. coli O157: H7 and its application in vegetable juices. Int. J. Food Microbiol. 2021, 338, 108989. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.; Coimbra, A.T.; Silva, L.; Duarte, A.P.; Ferreira, S. Melissa officinalis essential oil as an antimicrobial agent against Listeria monocytogenes in watermelon juice. Food Microbiol. 2023, 109, 104105. [Google Scholar] [CrossRef]
- Gerardi, C.; Pinto, L.; Baruzzi, F.; Giovinazzo, G. Comparison of antibacterial and antioxidant properties of red (cv. Negramaro) and white (cv. Fiano) skin pomace extracts. Molecules 2021, 26, 5918. [Google Scholar] [CrossRef]
- Tian, L.; Fu, J.; Wu, M.; Liao, S.; Jia, X.; Wang, J.; Yang, S.; Liu, Z.; Liu, Z.; Xue, Z.; et al. Evaluation of gallic acid on membrane damage of Yersinia enterocolitica and its application as a food preservative in pork. Int. J. Food Microbiol. 2022, 374, 109720. [Google Scholar] [CrossRef]
- Phuong, N.N.M.; Le, T.T.; Van Camp, J.; Raes, K. Evaluation of antimicrobial activity of rambutan (Nephelium lappaceum L.) peel extracts. Int. J. Food Microbiol. 2020, 321, 108539. [Google Scholar] [CrossRef]
- Belgacem, I.; Schena, L.; Teixidó, N.; Romeo, F.V.; Ballistreri, G.; Abadias, M. Effectiveness of a pomegranate peel extract (PGE) in reducing Listeria monocytogenes in vitro and on fresh-cut pear, apple and melon. Eur. Food Res. Technol. 2020, 246, 1765–1772. [Google Scholar] [CrossRef]
- Nicolau-Lapeña, I.; Aguiló-Aguayo, I.; Bobo, G.; Viñas, I.; Anguera, M.; Abadias, M. Ferulic acid application to control growth Listeria monocytogenes and Salmonella enterica on fresh-cut apples and melon, and its effect in quality parameters. Postharvest Biol. Technol. 2022, 186, 111831. [Google Scholar] [CrossRef]
- Bombelli, A.; Araya-Cloutier, C.; Vincken, J.P.; Abee, T.; den Besten, H.M. Impact of food-relevant conditions and food matrix on the efficacy of prenylated isoflavonoids glabridin and 6, 8-diprenylgenistein as potential natural preservatives against Listeria monocytogenes. Int. J. Food Microbiol. 2023, 390, 110109. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, S.; Hao, G.; Zhao, L.; Lü, X.; Wang, H.; Wang, L.; Zhang, J.; Ge, W. Antimicrobial activity and mechanism of isothiocyanate from Moringa oleifera seeds against Bacillus cereus and Cronobacter sakazakii and its application in goat milk. Food Control 2022, 139, 109067. [Google Scholar] [CrossRef]
- Dogruyol, H.; Mol, S.; Cosansu, S. Increased thermal sensitivity of Listeria monocytogenes in sous-vide salmon by oregano essential oil and citric acid. Food Microbiol. 2020, 90, 103496. [Google Scholar] [CrossRef]
- Orizano-Ponce, E.; Char, C.; Sepúlveda, F.; Ortiz-Viedma, J. Heat sensitization of Escherichia coli by the natural antimicrobials vanillin and emulsified citral in blended carrot-orange juice. Food Microbiol. 2022, 107, 104058. [Google Scholar] [CrossRef]
- Cheng, S.; Su, R.; Song, L.; Bai, X.; Yang, H.; Li, Z.; Li, Z.; Zhan, X.; Xia, X.; Lü, X.; et al. Citral and trans-cinnamaldehyde, two plant-derived antimicrobial agents can induce Staphylococcus aureus into VBNC state with different characteristics. Food Microbiol. 2023, 112, 104241. [Google Scholar] [CrossRef]
- de Candia, S.; Quintieri, L.; Caputo, L.; Baruzzi, F. Antimicrobial activity of processed spices used in traditional Southern Italian sausage processing. J. Food Process. Preserv. 2017, 41, e13022. [Google Scholar] [CrossRef]
- Palomares-Navarro, J.J.; Bernal-Mercado, A.T.; González-Aguilar, G.A.; Ortega-Ramirez, L.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Antibiofilm action of plant terpenes in Salmonella strains: Potential inhibitors of the synthesis of extracellular polymeric substances. Pathogens 2023, 12, 35. [Google Scholar] [CrossRef]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D.S. Plant natural products targeting bacterial virulence factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Lizardi-Mendoza, J.; Silva-Espinoza, B.A.; Cruz-Valenzuela, M.R.; Gonzalez-Aguilar, G.A.; Nazzaro, F.; Fratianni, F.; Ayala-Zavala, J.F. Phenolic extracts from grape stems inhibit Listeria monocytogenes motility and adhesion to food contact surfaces. J. Adhes. Sci. Technol. 2018, 32, 889–907. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Bernal-Mercado, A.T.; Tapia-Rodriguez, M.R.; Gonzalez-Aguilar, G.A.; Lopez-Zavala, A.A.; Martinez-Tellez, M.A.; Hernandez-Oñate, M.A.; Ayala-Zavala, J.F. Quercetin reduces adhesion and inhibits biofilm development by Listeria monocytogenes by reducing the amount of extracellular proteins. Food Control 2018, 90, 266–273. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Hernandez-Oñate, M.A.; Martinez-Tellez, M.A.; Lopez-Zavala, A.A.; Gonzalez-Aguilar, G.A.; Gutierrez-Pacheco, M.M.; Ayala-Zavala, J.F. Quercetin repressed the stress response factor (sigB) and virulence genes (prfA, actA, inlA, and inlC), lower the adhesion, and biofilm development of L. monocytogenes. Food Microbiol. 2020, 87, 103377. [Google Scholar] [CrossRef]
- Kim, Y.K.; Roy, P.K.; Ashrafudoulla, M.; Nahar, S.; Toushik, S.H.; Hossain, M.I.; Ha, S.D. Antibiofilm effects of quercetin against Salmonella enterica biofilm formation and virulence, stress response, and quorum-sensing gene expression. Food Control 2022, 137, 108964. [Google Scholar] [CrossRef]
- Ortega-Ramirez, L.A.; Gutiérrez-Pacheco, M.M.; Vargas-Arispuro, I.; González-Aguilar, G.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Inhibition of glucosyltransferase activity and glucan production as an antibiofilm mechanism of lemongrass essential oil against Escherichia coli O157: H7. Antibiotics 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Kostoglou, D.; Protopappas, I.; Giaouris, E. Common plant-derived terpenoids present increased anti-biofilm potential against Staphylococcus bacteria compared to a quaternary ammonium biocide. Foods 2020, 9, 697. [Google Scholar] [CrossRef]
- dos Santos Rodrigues, J.B.; de Carvalho, R.J.; de Souza, N.T.; de Sousa Oliveira, K.; Franco, O.L.; Schaffner, D.; de Souza, E.L.; Magnani, M. Effects of oregano essential oil and carvacrol on biofilms of Staphylococcus aureus from food-contact surfaces. Food Control 2017, 73, 1237–1246. [Google Scholar] [CrossRef]
- Ozma, M.A.; Abbasi, A.; Ahangarzadeh Rezaee, M.; Hosseini, H.; Hosseinzadeh, N.; Sabahi, S.; Kafil, H.S. A critical review on the nutritional and medicinal profiles of garlic’s (Allium sativum L.) bioactive compounds. Food Rev. Int. 2022, 1–38. [Google Scholar] [CrossRef]
- Yang, X.; Sha, K.; Xu, G.; Tian, H.; Wang, X.; Chen, S.; Wang, Y.; Li, J.; Chen, J.; Huang, N. Subinhibitory concentrations of allicin decrease uropathogenic Escherichia coli (UPEC) biofilm formation, adhesion ability, and swimming motility. Int. J. Mol. Sci. 2016, 17, 979. [Google Scholar] [CrossRef]
- Cruz-Valenzuela, M.R.; Ayala-Soto, R.E.; Ayala-Zavala, J.F.; Espinoza-Silva, B.A.; González-Aguilar, G.A.; Martín-Belloso, O.; Soliva-Fortuny, R.; Nazzaro, F.; Fratianni, F.; Tapia-Rodriguez, M.T.; et al. Pomegranate (Punica granatum L.) peel extracts as antimicrobial and antioxidant additives used in alfalfa sprouts. Foods 2022, 11, 2588. [Google Scholar] [CrossRef]
- Bouyahya, A.; Chamkhi, I.; Balahbib, A.; Rebezov, M.; Shariati, M.A.; Wilairatana, P.; El Omari, N. Mechanisms, anti-quorum-sensing actions, and clinical trials of medicinal plant bioactive compounds against bacteria: A comprehensive review. Molecules 2022, 27, 1484. [Google Scholar] [CrossRef] [PubMed]
- Fimbres-García, J.O.; Flores-Sauceda, M.; Othon-Díaz, E.D.; García-Galaz, A.; Tapia-Rodríguez, M.R.; Silva-Espinoza, B.A.; Ayala-Zavala, J.F. Facing resistant bacteria with plant essential oils: Reviewing the oregano case. Antibiotics 2022, 11, 1777. [Google Scholar] [CrossRef] [PubMed]
- Guillín, Y.; Cáceres, M.; Torres, R.; Stashenko, E.; Ortiz, C. Effect of essential oils on the inhibition of biofilm and quorum sensing in Salmonella enteritidis 13076 and Salmonella typhimurium 14028. Antibiotics 2021, 10, 1191. [Google Scholar] [CrossRef] [PubMed]
- Tamfu, A.N.; Ceylan, O.; Kucukaydin, S.; Duru, M.E. HPLC-DAD phenolic profiles, antibiofilm, anti-quorum sensing and enzyme inhibitory potentials of Camellia sinensis (L.) O. Kuntze and Curcuma longa L. LWT 2020, 133, 110150. [Google Scholar] [CrossRef]
- Tapia-Rodriguez, M.R.; Bernal-Mercado, A.T.; Gutierrez-Pacheco, M.M.; Vazquez-Armenta, F.J.; Hernandez-Mendoza, A.; Gonzalez-Aguilar, G.A.; Martinez-Tellez, M.A.; Nazzaro, F.; Ayala-Zavala, J.F. Virulence of Pseudomonas aeruginosa exposed to carvacrol: Alterations of the Quorum sensing at enzymatic and gene levels. J. Cell Commun. Signal. 2019, 13, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.D. Clinically important toxins in bacterial infection: Utility of laboratory detection. Clin. Microbiol. Newsl. 2020, 42, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Nwabor, O.F.; Singh, S.; Syukri, D.M.; Voravuthikunchai, S.P. Bioactive fractions of Eucalyptus camaldulensis inhibit important foodborne pathogens, reduce listeriolysin O-induced haemolysis, and ameliorate hydrogen peroxide-induced oxidative stress on human embryonic colon cells. Food Chem. 2021, 344, 128571. [Google Scholar] [CrossRef]
- Rasooly, R.; Molnar, A.; Choi, H.-Y.; Do, P.; Racicot, K.; Apostolidis, E. In-vitro inhibition of staphylococcal pathogenesis by witch-hazel and green tea extracts. Antibiotics 2019, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhao, X.; Yan, H.; Meng, R.; Zhang, Y.; Li, W.; Liu, Z.; Guo, N. Effect of tea tree oil on Staphylococcus aureus growth and enterotoxin production. Food Control 2016, 62, 257–263. [Google Scholar] [CrossRef]
- Zhang, M.; Li, H.; Agyekumwaa, A.K.; Yu, Y.; Xiao, X. Effects of citronellal on growth and enterotoxins production in Staphylococcus aureus ATCC 29213. Toxicon 2022, 213, 92–98. [Google Scholar] [CrossRef]
- Mooyottu, S.; Kollanoor-Johny, A.; Flock, G.; Bouillaut, L.; Upadhyay, A.; Sonenshein, A.L.; Venkitanarayanan, K. Carvacrol and trans-cinnamaldehyde reduce Clostridium difficile toxin production and cytotoxicity in vitro. Int. J. Mol. Sci. 2014, 15, 4415–4430. [Google Scholar] [CrossRef]
- Tian, F.; Woo, S.Y.; Lee, S.Y.; Park, S.B.; Im, J.H.; Chun, H.S. Plant-based natural flavonoids show strong inhibition of aflatoxin production and related gene expressions correlated with chemical structure. Food Microbiol. 2023, 109, 104141. [Google Scholar] [CrossRef]
- Pok, P.S.; Londoño, V.A.G.; Vicente, S.; Romero, S.M.; Pacín, A.; Tolaba, M.; Alzamora, S.M.; Resnik, S.L. Evaluation of citrus flavonoids against Aspergillus parasiticus in maize: Aflatoxins reduction and ultrastructure alterations. Food Chem. 2020, 318, 126414. [Google Scholar] [CrossRef] [PubMed]
- Hamad, G.M.; Mohdaly, A.A.A.; El-Nogoumy, B.A.; Ramadan, M.F.; Hassan, S.A.; Zeitoun, A.M. Detoxification of aflatoxin B1 and ochratoxin A using Salvia farinacea and Azadirachta indica water extract and application in meat products. Appl. Biochem. Biotechnol. 2021, 193, 3098–3120. [Google Scholar] [CrossRef] [PubMed]
- Ponzilacqua, B.; Rottinghaus, G.E.; Landers, B.R.; Oliveira, C.A.F.D. Effects of medicinal herb and Brazilian traditional plant extracts on in vitro mycotoxin decontamination. Food Control 2019, 100, 24–27. [Google Scholar] [CrossRef]
- Friedman, M.; Jürgens, H.S. Effect of pH on the stability of plant phenolic compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef]
- Hatab, S.; Athanasio, R.; Holley, R.; Rodas-Gonzalez, A.; Narvaez-Bravo, C. Survival and reduction of shiga toxin-producing Escherichia coli in a fresh cold-pressed juice treated with antimicrobial plant extracts. J. Food Sci. 2016, 81, M1987–M1995. [Google Scholar] [CrossRef] [PubMed]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Assessment of factors influencing antimicrobial activity of carvacrol and cymene against Vibrio cholerae in food. J. Biosci. Bioeng. 2010, 110, 614–619. [Google Scholar] [CrossRef]
- Salvia-Trujillo, L.; Rojas-Graü, A.; Soliva-Fortuny, R.; Martín-Belloso, O. Physicochemical characterization and antimicrobial activity of food-grade emulsions and nanoemulsions incorporating essential oils. Food Hydrocoll. 2015, 43, 547–556. [Google Scholar] [CrossRef]
- Topuz, O.K.; Özvural, E.B.; Zhao, Q.; Huang, Q.; Chikindas, M.; Gölükçü, M. Physical and antimicrobial properties of anise oil loaded nanoemulsions on the survival of foodborne pathogens. Food Chem. 2016, 203, 117–123. [Google Scholar] [CrossRef]
- Ghazy, O.A.; Fouad, M.T.; Saleh, H.H.; Kholif, A.E.; Morsy, T.A. Ultrasound-assisted preparation of anise extract nanoemulsion and its bioactivity against different pathogenic bacteria. Food Chem. 2021, 341, 128259. [Google Scholar] [CrossRef] [PubMed]
- Ghazy, O.A.; Fouad, M.T.; Morsy, T.A.; Kholif, A.E. Nanoemulsion formulation of Lawsonia inermis extract and its potential antimicrobial and preservative efficacy against foodborne pathogens. Food Control 2023, 145, 109458. [Google Scholar] [CrossRef]
- Bhargava, K.; Conti, D.S.; da Rocha, S.R.; Zhang, Y. Application of an oregano oil nanoemulsion to the control of foodborne bacteria on fresh lettuce. Food Microbiol. 2015, 47, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.K.; Bhargava, K.; Kotturi, H. Antimicrobial activity of cinnamon oil nanoemulsion against Listeria monocytogenes and Salmonella spp. on melons. LWT 2019, 111, 682–687. [Google Scholar] [CrossRef]
- Molet-Rodríguez, A.; Turmo-Ibarz, A.; Salvia-Trujillo, L.; Martín-Belloso, O. Incorporation of antimicrobial nanoemulsions into complex foods: A case study in an apple juice-based beverage. LWT 2021, 141, 110926. [Google Scholar] [CrossRef]
- Luciano, W.A.; Pimentel, T.C.; Bezerril, F.F.; Barão, C.E.; Marcolino, V.A.; Carvalho, R.D.S.F.; dos Santos Lima, M.; Martín-Belloso, O.; Magnani, M. Effect of citral nanoemulsion on the inactivation of Listeria monocytogenes and sensory properties of fresh-cut melon and papaya during storage. Int. J. Food Microbiol. 2023, 384, 109959. [Google Scholar] [CrossRef]
- Ozogul, Y.; Boğa, E.K.; Akyol, I.; Durmus, M.; Ucar, Y.; Regenstein, J.M.; Köşker, A.R. Antimicrobial activity of thyme essential oil nanoemulsions on spoilage bacteria of fish and food-borne pathogens. Food Biosci. 2020, 36, 100635. [Google Scholar] [CrossRef]
- Özogul, Y.; El Abed, N.; Özogul, F. Antimicrobial effect of laurel essential oil nanoemulsion on food-borne pathogens and fish spoilage bacteria. Food Chem. 2022, 368, 130831. [Google Scholar] [CrossRef]
- Özogul, Y.; Özogul, F.; Kulawik, P. The antimicrobial effect of grapefruit peel essential oil and its nanoemulsion on fish spoilage bacteria and food-borne pathogens. LWT 2021, 136, 110362. [Google Scholar] [CrossRef]
- Tang, M.; Liu, F.; Wang, Q.; Wang, D.; Wang, D.; Zhu, Y.; Sun, Z.; Xu, W. Physicochemical characteristics of ginger essential oil nanoemulsion encapsulated by zein/NaCas and antimicrobial control on chilled chicken. Food Chem. 2022, 374, 131624. [Google Scholar] [CrossRef]
- Yang, R.; Miao, J.; Shen, Y.; Cai, N.; Wan, C.; Zou, L.; Chen, C.; Chen, J. Antifungal effect of cinnamaldehyde, eugenol and carvacrol nanoemulsion against Penicillium digitatum and application in postharvest preservation of citrus fruit. LWT 2021, 141, 110924. [Google Scholar] [CrossRef]
- Gundewadi, G.; Sarkar, D.J.; Rudra, S.G.; Singh, D. Preparation of basil oil nanoemulsion using Sapindus mukorossi pericarp extract: Physico-chemical properties and antifungal activity against food spoilage pathogens. Ind. Crops Prod. 2018, 125, 95–104. [Google Scholar] [CrossRef]
- Ribes, S.; Fuentes, A.; Barat, J.M. Effect of oregano (Origanum vulgare L. ssp. hirtum) and clove (Eugenia spp.) nanoemulsions on Zygosaccharomyces bailii survival in salad dressings. Food Chem. 2019, 295, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Zhong, S.; Schwarz, P.; Chen, B.; Rao, J. Physical properties, antifungal and mycotoxin inhibitory activities of five essential oil nanoemulsions: Impact of oil compositions and processing parameters. Food Chem. 2019, 291, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.K.; Singh, V.K.; Das, S.; Prasad, J.; Dwivedy, A.K.; Dubey, N.K. Improvement of in vitro and in situ antifungal, AFB1 inhibitory and antioxidant activity of Origanum majorana L. essential oil through nanoemulsion and recommending as novel food preservative. Food Chem. Toxicol. 2020, 143, 111536. [Google Scholar] [CrossRef]
- Gharsallaoui, A.; Roudaut, G.; Chambin, O.; Voilley, A.; Saurel, R. Applications of spray-drying in microencapsulation of food ingredients: An overview. Food Res. Int. 2007, 40, 1107–1121. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, Y.; Zhong, Q. Physical and antimicrobial properties of spray-dried zein–casein nanocapsules with co-encapsulated eugenol and thymol. J. Food Eng. 2015, 144, 93–102. [Google Scholar] [CrossRef]
- Talón, E.; Lampi, A.M.; Vargas, M.; Chiralt, A.; Jouppila, K.; González-Martínez, C. Encapsulation of eugenol by spray-drying using whey protein isolate or lecithin: Release kinetics, antioxidant and antimicrobial properties. Food Chem. 2019, 295, 588–598. [Google Scholar] [CrossRef]
- Sun, X.; Cameron, R.G.; Bai, J. Effect of spray-drying temperature on physicochemical, antioxidant and antimicrobial properties of pectin/sodium alginate microencapsulated carvacrol. Food Hydrocoll. 2020, 100, 105420. [Google Scholar] [CrossRef]
- Plati, F.; Papi, R.; Paraskevopoulou, A. Characterization of oregano essential oil (Origanum vulgare L. subsp. hirtum) particles produced by the novel nano spray drying technique. Foods 2021, 10, 2923. [Google Scholar] [CrossRef]
- Cruz-Molina, A.V.D.L.; Ayala Zavala, J.F.; Bernal Mercado, A.T.; Cruz Valenzuela, M.R.; González-Aguilar, G.A.; Lizardi-Mendoza, J.; Brown-Bojorquez, F.; Silva-Espinoza, B.A. Maltodextrin encapsulation improves thermal and pH stability of green tea extract catechins. J. Food Process. Preserv. 2021, 45, e15729. [Google Scholar] [CrossRef]
- Radünz, M.; dos Santos Hackbart, H.C.; Camargo, T.M.; Nunes, C.F.P.; de Barros, F.A.P.; Dal Magro, J.; Filho, P.J.S.; Gandra, E.A.; Radünz, A.L.; da Rosa Zavareze, E. Antimicrobial potential of spray drying encapsulated thyme (Thymus vulgaris) essential oil on the conservation of hamburger-like meat products. Int. J. Food Microbiol. 2020, 330, 108696. [Google Scholar] [CrossRef] [PubMed]
- do Valle Calomeni, A.; de Souza, V.B.; Tulini, F.L.; Thomazini, M.; Ostroschi, L.C.; de Alencar, S.M.; Massarioli, A.P.; de Carvalho Balieiro, J.C.; de Carvalho, R.A.; Favaro-Trindade, C.S. Characterization of antioxidant and antimicrobial properties of spray-dried extracts from peanut skins. Food Bioprod. Process. 2017, 105, 215–223. [Google Scholar] [CrossRef]
- Vinceković, M.; Viskić, M.; Jurić, S.; Giacometti, J.; Kovačević, D.B.; Putnik, P.; Donsì, F.; Barba, F.J.; Jambrak, A.R. Innovative technologies for encapsulation of Mediterranean plants extracts. Trends Food Sci. Technol. 2017, 69, 1–12. [Google Scholar] [CrossRef]
- de Araújo, J.S.F.; de Souza, E.L.; Oliveira, J.R.; Gomes, A.C.A.; Kotzebue, L.R.V.; da Silva Agostini, D.L.; de Oliveira, D.L.V.; Mazzetto, S.M.; da Silva, A.L.; Cavalcanti, M.T. Microencapsulation of sweet orange essential oil (Citrus aurantium var. dulcis) by liophylization using maltodextrin and maltodextrin/gelatin mixtures: Preparation, characterization, antimicrobial and antioxidant activities. Int. J. Biol. Macromol. 2020, 143, 991–999. [Google Scholar] [CrossRef]
- Viacava, G.E.; Ayala-Zavala, J.F.; González-Aguilar, G.A.; Ansorena, M.R. Effect of free and microencapsulated thyme essential oil on quality attributes of minimally processed lettuce. Postharvest Biol. Technol. 2018, 145, 125–133. [Google Scholar] [CrossRef]
- Garcia-Sotelo, D.; Silva-Espinoza, B.; Perez-Tello, M.; Olivas, I.; Alvarez-Parrilla, E.; González-Aguilar, G.A.; Ayala-Zavala, J.F. Antimicrobial activity and thermal stability of rosemary essential oil: β−cyclodextrin capsules applied in tomato juice. LWT 2019, 111, 837–845. [Google Scholar] [CrossRef]
- Silva, F.; Caldera, F.; Trotta, F.; Nerin, C.; Domingues, F.C. Encapsulation of coriander essential oil in cyclodextrin nanosponges: A new strategy to promote its use in controlled-release active packaging. Innov. Food Sci. Emerg. Technol. 2019, 56, 102177. [Google Scholar] [CrossRef]
- Ozdemir, N.; Pola, C.C.; Teixeira, B.N.; Hill, L.E.; Bayrak, A.; Gomes, C.L. Preparation of black pepper oleoresin inclusion complexes based on beta-cyclodextrin for antioxidant and antimicrobial delivery applications using kneading and freeze drying methods: A comparative study. LWT 2018, 91, 439–445. [Google Scholar] [CrossRef]
- Lin, L.; Gu, Y.; Sun, Y.; Cui, H. Characterization of chrysanthemum essential oil triple-layer liposomes and its application against Campylobacter jejuni on chicken. LWT 2019, 107, 16–24. [Google Scholar] [CrossRef]
- Yang, K.; Liu, A.; Hu, A.; Li, J.; Zen, Z.; Liu, Y.; Tang, S.; Li, C. Preparation and characterization of cinnamon essential oil nanocapsules and comparison of volatile components and antibacterial ability of cinnamon essential oil before and after encapsulation. Food Control 2021, 123, 107783. [Google Scholar] [CrossRef]
- Augustin, M.A.; Sanguansri, P. Challenges and solutions to incorporation of nutraceuticals in foods. Annu. Rev. Food Sci. Technol. 2015, 3, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Bhandari, B. Encapsulation of polyphenols—A review. Trends Food Sci. Technol. 2010, 21, 510–523. [Google Scholar] [CrossRef]
- Arruda, T.R.; Bernardes, P.C.; e Moraes, A.R.F.; Soares, N.D.F.F. Natural bioactives in perspective: The future of active packaging based on essential oils and plant extracts themselves and those complexed by cyclodextrins. Food Res. Int. 2022, 156, 111160. [Google Scholar] [CrossRef]
- Cui, R.; Zhu, B.; Yan, J.; Qin, Y.; Yuan, M.; Cheng, G.; Yuan, M. Development of a sodium alginate-based active package with controlled release of cinnamaldehyde loaded on halloysite nanotubes. Foods 2021, 10, 1150. [Google Scholar] [CrossRef]
- Cometa, S.; Bonifacio, M.A.; Bellissimo, A.; Pinto, L.; Petrella, A.; De Vietro, N.; Iannaccone, G.; Baruzzi, F.; De Giglio, E. A green approach to develop zeolite-thymol antimicrobial composites: Analytical characterization and antimicrobial activity evaluation. Heliyon 2022, 8, e09551. [Google Scholar] [CrossRef]
- Wang, Y.; Yi, S.; Lu, R.; Sameen, D.E.; Ahmed, S.; Dai, J.; Qin, W.; Li, S.; Liu, Y. Preparation, characterization, and 3D printing verification of chitosan/halloysite nanotubes/tea polyphenol nanocomposite films. Int. J. Biol. Macromol. 2021, 166, 32–44. [Google Scholar] [CrossRef]
- Jiang, Y.; Yin, H.; Zhou, X.; Wang, D.; Zhong, Y.; Xia, Q.; Deng, Y.; Zhao, Y. Antimicrobial, antioxidant and physical properties of chitosan film containing Akebia trifoliata (Thunb.) Koidz. peel extract/montmorillonite and its application. Food Chem. 2021, 361, 130111. [Google Scholar] [CrossRef]
- Riaz, A.; Lei, S.; Akhtar, H.M.S.; Wan, P.; Chen, D.; Jabbar, S.; Abid, M.; Hashim, M.M.; Zeng, X. Preparation and characterization of chitosan-based antimicrobial active food packaging film incorporated with apple peel polyphenols. Int. J. Biol. Macromol. 2018, 114, 547–555. [Google Scholar] [CrossRef]
- Wang, K.; Lim, P.N.; Tong, S.Y.; San Thian, E. Development of grapefruit seed extract-loaded poly (ε-caprolactone)/chitosan films for antimicrobial food packaging. Food Packag. Shelf Life 2019, 22, 100396. [Google Scholar] [CrossRef]
- Surendhiran, D.; Li, C.; Cui, H.; Lin, L. Fabrication of high stability active nanofibers encapsulated with pomegranate peel extract using chitosan/PEO for meat preservation. Food Packag. Shelf Life 2020, 23, 100439. [Google Scholar] [CrossRef]
- Gutiérrez-Pacheco, M.M.; Ortega-Ramírez, L.A.; Silva-Espinoza, B.A.; Cruz-Valenzuela, M.R.; González-Aguilar, G.A.; Lizardi-Mendoza, J.; Miranda, R.; Ayala-Zavala, J.F. Individual and combined coatings of chitosan and carnauba wax with oregano essential oil to avoid water loss and microbial decay of fresh cucumber. Coatings 2020, 10, 614. [Google Scholar] [CrossRef]
- Saberi, B.; Chockchaisawasdee, S.; Golding, J.B.; Scarlett, C.J.; Stathopoulos, C.E. Characterization of pea starch-guar gum biocomposite edible films enriched by natural antimicrobial agents for active food packaging. Food Bioprod. Process. 2017, 105, 51–63. [Google Scholar] [CrossRef]
- Chollakup, R.; Pongburoos, S.; Boonsong, W.; Khanoonkon, N.; Kongsin, K.; Sothornvit, R.; Sukyai, P.; Sukatta, U.; Harnkarnsujarit, N. Antioxidant and antibacterial activities of cassava starch and whey protein blend films containing rambutan peel extract and cinnamon oil for active packaging. LWT 2020, 130, 109573. [Google Scholar] [CrossRef]
- Issa, A.; Ibrahim, S.A.; Tahergorabi, R. Impact of sweet potato starch-based nanocomposite films activated with thyme essential oil on the shelf-life of baby spinach leaves. Foods 2017, 6, 43. [Google Scholar] [CrossRef]
- Aloui, H.; Deshmukh, A.R.; Khomlaem, C.; Kim, B.S. Novel composite films based on sodium alginate and gallnut extract with enhanced antioxidant, antimicrobial, barrier and mechanical properties. Food Hydrocoll. 2021, 113, 106508. [Google Scholar] [CrossRef]
- Kapetanakou, A.E.; Nestora, S.; Evageliou, V.; Skandamis, P.N. Sodium alginate–cinnamon essential oil coated apples and pears: Variability of Aspergillus carbonarius growth and ochratoxin A production. Food Res. Int. 2019, 119, 876–885. [Google Scholar] [CrossRef]
- Nouri, A.; Yaraki, M.T.; Ghorbanpour, M.; Wang, S. Biodegradable κ-carrageenan/nanoclay nanocomposite films containing Rosmarinus officinalis L. extract for improved strength and antibacterial performance. Int. J. Biol. Macromol. 2018, 115, 227–235. [Google Scholar] [CrossRef]
- He, S.; Wang, Y. Antimicrobial and antioxidant effects of kappa-carrageenan coatings enriched with cinnamon essential oil in pork meat. Foods 2022, 11, 2885. [Google Scholar] [CrossRef]
- Göksen, G.; Fabra, M.J.; Ekiz, H.I.; López-Rubio, A. Phytochemical-loaded electrospun nanofibers as novel active edible films: Characterization and antibacterial efficiency in cheese slices. Food Control 2020, 112, 107133. [Google Scholar] [CrossRef]
- Shankar, S.; Rhim, J.W. Preparation of antibacterial poly (lactide)/poly (butylene adipate-co-terephthalate) composite films incorporated with grapefruit seed extract. Int. J. Biol. Macromol. 2018, 120, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Suwanamornlert, P.; Kerddonfag, N.; Sane, A.; Chinsirikul, W.; Zhou, W.; Chonhenchob, V. Poly (lactic acid)/poly (butylene-succinate-co-adipate) (PLA/PBSA) blend films containing thymol as alternative to synthetic preservatives for active packaging of bread. Food Packag. Shelf Life 2020, 25, 100515. [Google Scholar] [CrossRef]
- Requena, R.; Vargas, M.; Chiralt, A. Eugenol and carvacrol migration from PHBV films and antibacterial action in different food matrices. Food Chem. 2019, 277, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Pabast, M.; Shariatifar, N.; Beikzadeh, S.; Jahed, G. Effects of chitosan coatings incorporating with free or nano-encapsulated Satureja plant essential oil on quality characteristics of lamb meat. Food Control 2018, 91, 185–192. [Google Scholar] [CrossRef]
- Amor, G.; Sabbah, M.; Caputo, L.; Idbella, M.; De Feo, V.; Porta, R.; Fechtali, T.; Mauriello, G. Basil essential oil: Composition, antimicrobial properties, and microencapsulation to produce active chitosan films for food packaging. Foods 2021, 10, 121. [Google Scholar] [CrossRef]
- Pan, J.; Ai, F.; Shao, P.; Chen, H.; Gao, H. Development of polyvinyl alcohol/β-cyclodextrin antimicrobial nanofibers for fresh mushroom packaging. Food Chem. 2019, 300, 125249. [Google Scholar] [CrossRef]
- Chen, Z.; Zong, L.; Chen, C.; Xie, J. Development and characterization of PVA-Starch active films incorporated with β-cyclodextrin inclusion complex embedding lemongrass (Cymbopogon citratus) oil. Food Packag. Shelf Life 2020, 26, 100565. [Google Scholar] [CrossRef]
- Lee, J.Y.; Garcia, C.V.; Shin, G.H.; Kim, J.T. Antibacterial and antioxidant properties of hydroxypropyl methylcellulose-based active composite films incorporating oregano essential oil nanoemulsions. LWT 2019, 106, 164–171. [Google Scholar] [CrossRef]
- Shokri, S.; Parastouei, K.; Taghdir, M.; Abbaszadeh, S. Application an edible active coating based on chitosan-Ferulago angulata essential oil nanoemulsion to shelf life extension of Rainbow trout fillets stored at 4 °C. Int. J. Biol. Macromol. 2020, 153, 846–854. [Google Scholar] [CrossRef]
- Abbasi, Z.; Aminzare, M.; Hassanzad Azar, H.; Rostamizadeh, K. Effect of corn starch coating incorporated with nanoemulsion of Zataria multiflora essential oil fortified with cinnamaldehyde on microbial quality of fresh chicken meat and fate of inoculated Listeria monocytogenes. J. Food Sci. Technol. 2021, 58, 2677–2687. [Google Scholar] [CrossRef]
- Corrêa, J.A.F.; dos Santos, J.V.G.; Evangelista, A.G.; Pinto, A.C.S.M.; de Macedo, R.E.F.; Luciano, F.B. Combined application of phenolic acids and essential oil components against Salmonella Enteritidis and Listeria monocytogenes in vitro and in ready-to-eat cooked ham. LWT 2021, 149, 111881. [Google Scholar] [CrossRef]
- Nikkhah, M.; Hashemi, M. Boosting antifungal effect of essential oils using combination approach as an efficient strategy to control postharvest spoilage and preserving the jujube fruit quality. Postharvest Biol. Technol. 2020, 164, 111159. [Google Scholar] [CrossRef]
- Purkait, S.; Bhattacharya, A.; Bag, A.; Chattopadhyay, R.R. Synergistic antibacterial, antifungal and antioxidant efficacy of cinnamon and clove essential oils in combination. Arch. Microbiol. 2020, 202, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, D.; Cao, Y.; Zhang, Y.; Xiao, X.; Liu, F.; Yu, Y. Synergistic inactivation of Escherichia coli O157: H7 and Staphylococcus aureus by gallic acid and thymol and its potential application on fresh-cut tomatoes. Food Microbiol. 2022, 102, 103925. [Google Scholar] [CrossRef]
- Meira, N.V.; Holley, R.A.; Bordin, K.; de Macedo, R.E.; Luciano, F.B. Combination of essential oil compounds and phenolic acids against Escherichia coli O157: H7 in vitro and in dry-fermented sausage production. Int. J. Food Microbiol. 2017, 260, 59–64. [Google Scholar] [CrossRef]
- Nikkhah, M.; Hashemi, M.; Najafi, M.B.H.; Farhoosh, R. Synergistic effects of some essential oils against fungal spoilage on pear fruit. Int. J. Food Microbiol. 2017, 257, 285–294. [Google Scholar] [CrossRef]
- Aaliya, B.; Sunooj, K.V.; Navaf, M.; Akhila, P.P.; Sudheesh, C.; Mir, S.A.; Sabu, S.; Sasidharan, A.; Theingi Hlaing, M.; George, J. Recent trends in bacterial decontamination of food products by hurdle technology: A synergistic approach using thermal and non-thermal processing techniques. Food Res. Int. 2021, 147, 110514. [Google Scholar] [CrossRef]
- Pinto, L.; Baruzzi, F.; Cocolin, L.; Malfeito-Ferreira, M. Emerging technologies to control Brettanomyces spp. in wine: Recent advances and future trends. Trends Food Sci. Technol. 2020, 99, 88–100. [Google Scholar] [CrossRef]
- Patrignani, F.; Siroli, L.; Braschi, G.; Lanciotti, R. Combined use of natural antimicrobial based nanoemulsions and ultra high pressure homogenization to increase safety and shelf-life of apple juice. Food Control 2020, 111, 107051. [Google Scholar] [CrossRef]
- Chien, S.Y.; Sheen, S.; Sommers, C.; Sheen, L.Y. Combination effect of high-pressure processing and essential oil (Melissa officinalis extracts) or their constituents for the inactivation of Escherichia coli in ground beef. Food Bioproc. Tech. 2019, 12, 359–370. [Google Scholar] [CrossRef]
- Sheen, S.; Huang, C.Y.; Chuang, S. Synergistic effect of high hydrostatic pressure, allyl isothiocyanate, and acetic acid on the inactivation and survival of pathogenic Escherichia coli in ground chicken. J. Food Sci. 2022, 87, 5042–5053. [Google Scholar] [CrossRef] [PubMed]
- González-González, C.R.; Labo-Popoola, O.; Delgado-Pando, G.; Theodoridou, K.; Doran, O.; Stratakos, A.C. The effect of cold atmospheric plasma and linalool nanoemulsions against Escherichia coli O157: H7 and Salmonella on ready-to-eat chicken meat. LWT 2021, 149, 111898. [Google Scholar] [CrossRef]
- Olatunde, O.O.; Benjakul, S.; Vongkamjan, K. Cold plasma combined with liposomal ethanolic coconut husk extract: A potential hurdle technology for shelf-life extension of Asian sea bass slices packaged under modified atmosphere. Innov. Food Sci. Emerg. Technol. 2020, 65, 102448. [Google Scholar] [CrossRef]
- Van Haute, S.; Raes, K.; Devlieghere, F.; Sampers, I. Combined use of cinnamon essential oil and MAP/vacuum packaging to increase the microbial and sensorial shelf life of lean pork and salmon. Food Packag. Shelf Life 2017, 12, 51–58. [Google Scholar] [CrossRef]
- Shiekh, K.A.; Benjakul, S. Melanosis and quality changes during refrigerated storage of Pacific white shrimp treated with Chamuang (Garcinia cowa Roxb.) leaf extract with the aid of pulsed electric field. Food Chem. 2020, 309, 125516. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, L.; He, Q.; Arabi, S.A.; Zhao, H.; Chen, W.; Ye, X.; Liu, D. Synergistic antibacterial effects of ultrasound and thyme essential oils nanoemulsion against Escherichia coli O157: H7. Ultrason. Sonochem. 2020, 66, 104988. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Khang, D. Synergistic effects of 915 MHz microwave heating and essential oils on inactivation of foodborne pathogen in hot-chili sauce. Int. J. Food Microbiol. 2023, 398, 110210. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, S.; Guo, K.; Guo, D.; Li, J.; Wang, M.; Wang, Y.; Zhang, C.; Xia, X.; Shi, C. Efficacy of 405-nm LED illumination and citral used alone and in combination for the inactivation of Cronobacter sakazakii in reconstituted powdered infant formula. Food Res. Int. 2022, 154, 111027. [Google Scholar] [CrossRef]
- Silva-Espinoza, B.A.; Palomares-Navarro, J.J.; Tapia-Rodriguez, M.R.; Cruz-Valenzuela, M.R.; González-Aguilar, G.A.; Silva-Campa, E.; Pedroza-Montero, M.; Almeida-Lopes, M.; Miranda, R.; Ayala-Zavala, J.F. Combination of ultraviolet light-C and clove essential oil to inactivate Salmonella typhimurium biofilms on stainless steel. J. Food Saf. 2020, 40, e12788. [Google Scholar] [CrossRef]
- Chelliah, R.; Jo, K.H.; Yan, P.; Chen, X.; Jo, H.Y.; Madar, I.H.; Sultan, G.; Oh, D.H. Unravelling the sanitization potential of slightly acidic electrolyzed water combined Thymus vulgaris based nanoemulsion against foodborne pathogens and its safety assessment. Food Control 2023, 146, 109527. [Google Scholar] [CrossRef]
- Shankar, S.; Danneels, F.; Lacroix, M. Coating with alginate containing a mixture of essential oils and citrus extract in combination with ozonation or gamma irradiation increased the shelf life of Merluccius sp. fillets. Food Packag. Shelf Life 2019, 22, 100434. [Google Scholar] [CrossRef]
- Reinholds, I.; Pugajeva, I.; Bavrins, K.; Kuckovska, G.; Bartkevics, V. Mycotoxins, pesticides and toxic metals in commercial spices and herbs. Food Addit. Contam. Part B 2017, 10, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, N.; Höferl, M.; Buchbauer, G. Pesticides in essential oils and selected fragrance extracts. Some examples. A review. Flavour Frag. J. 2018, 33, 373–384. [Google Scholar] [CrossRef]
- Corrales, M.; Fernandez, A.; Pinto, M.G.V.; Butz, P.; Franz, C.M.; Schuele, E.; Tauscher, B. Characterization of phenolic content, in vitro biological activity, and pesticide loads of extracts from white grape skins from organic and conventional cultivars. Food Chem. Toxicol. 2010, 48, 3471–3476. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, H.; Ying, G.; Yang, M.; Nian, Y.; Liu, J.; Kong, W. Relationship of mycotoxins accumulation and bioactive components variation in ginger after fungal inoculation. Front. Pharmacol. 2017, 8, 331. [Google Scholar] [CrossRef]
- Hassan, H.F.; Koaik, L.; Khoury, A.E.; Atoui, A.; El Obeid, T.; Karam, L. Dietary exposure and risk assessment of mycotoxins in thyme and thyme-based products marketed in Lebanon. Toxins 2022, 14, 331. [Google Scholar] [CrossRef]
Figure 1.
Mechanisms of action of plant antimicrobials against foodborne bacteria.

Figure 2.
Anti-quorum sensing potential of carvacrol in foodborne bacteria.

Figure 3.
Hurdle effect of plant bioactive compounds against E. coli O157:H7 contaminating chicken meat when combined with high pressure or cold plasma treatments. The picture was made by drawing the main results from references [277,278].

Table 2.
Advantages and disadvantages of the spray-drying process for producing stable and functional plant antimicrobial powders.
Table 2.
Advantages and disadvantages of the spray-drying process for producing stable and functional plant antimicrobial powders.
| Plant Antimicrobial | Spray Drying Inlet Temperature (°C) | Protective Matrix | Microbial Targets | Advantages | Disadvantages | Data from Ref. |
|---|---|---|---|---|---|---|
| Eugenol and thymol | 105 | Zein/casein | E. coli O157:H7, L. monocytogenes Scott A | Good dispersion in water and good stability during storage | not reported | [223] |
| Eugenol | 180 | Whey protein/maltodextrin/chitosan | E. coli, L. innocua | High encapsulation efficiency and thermal stability | Chitosan inclusion negatively affects thermal stability, releasing and antimicrobial properties of the powder | [224] |
| Carvacrol | 100–190 | Pectin/sodium alginate | E. coli K12 | Better thermal stability | High inlet temperature affects dissolution time and hygroscopicity | [225] |
| Oregano EO | 100 | Whey protein/maltodextrin | E. coli, S. aureus | Low residence time, high yield, low inlet temperature | Low throughput, extended processing hours, high production cost | [226] |
| Green tea extract | 150 | Maltodextrin | - | High thermal stability and reduced weight loss | not reported | [227] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pinto, L.; Tapia-Rodríguez, M.R.; Baruzzi, F.; Ayala-Zavala, J.F. Plant Antimicrobials for Food Quality and Safety: Recent Views and Future Challenges. Foods 2023, 12, 2315. https://doi.org/10.3390/foods12122315
AMA Style
Pinto L, Tapia-Rodríguez MR, Baruzzi F, Ayala-Zavala JF. Plant Antimicrobials for Food Quality and Safety: Recent Views and Future Challenges. Foods. 2023; 12(12):2315. https://doi.org/10.3390/foods12122315
Chicago/Turabian StylePinto, Loris, Melvin R. Tapia-Rodríguez, Federico Baruzzi, and Jesús Fernando Ayala-Zavala. 2023. "Plant Antimicrobials for Food Quality and Safety: Recent Views and Future Challenges" Foods 12, no. 12: 2315. https://doi.org/10.3390/foods12122315
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.