
Enhanced Gel Properties of Duck Myofibrillar Protein by Plasma-Activated Water: Through Mild Structure Modifications

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of Myofibrillar Proteins
2.3. Preparation of PAW
2.4. Preparation of DMP Gels
2.5. Gel Strength
2.6. Water Holding Capacity
2.7. Dynamic Rheological Behavior
2.8. Molecular Forces
2.9. Microstructure
2.10. Protein Solubility
2.11. Gel Electrophoresis
2.12. Total Sulfhydryl Content
2.13. Carbonyl Content
2.14. Secondary Structure Analysis
2.15. Surface Hydrophobicity
2.16. UV Absorption Spectral Analysis
2.17. Fluorescence Measurement
2.18. Statistical Analysis
3. Results and Discussion
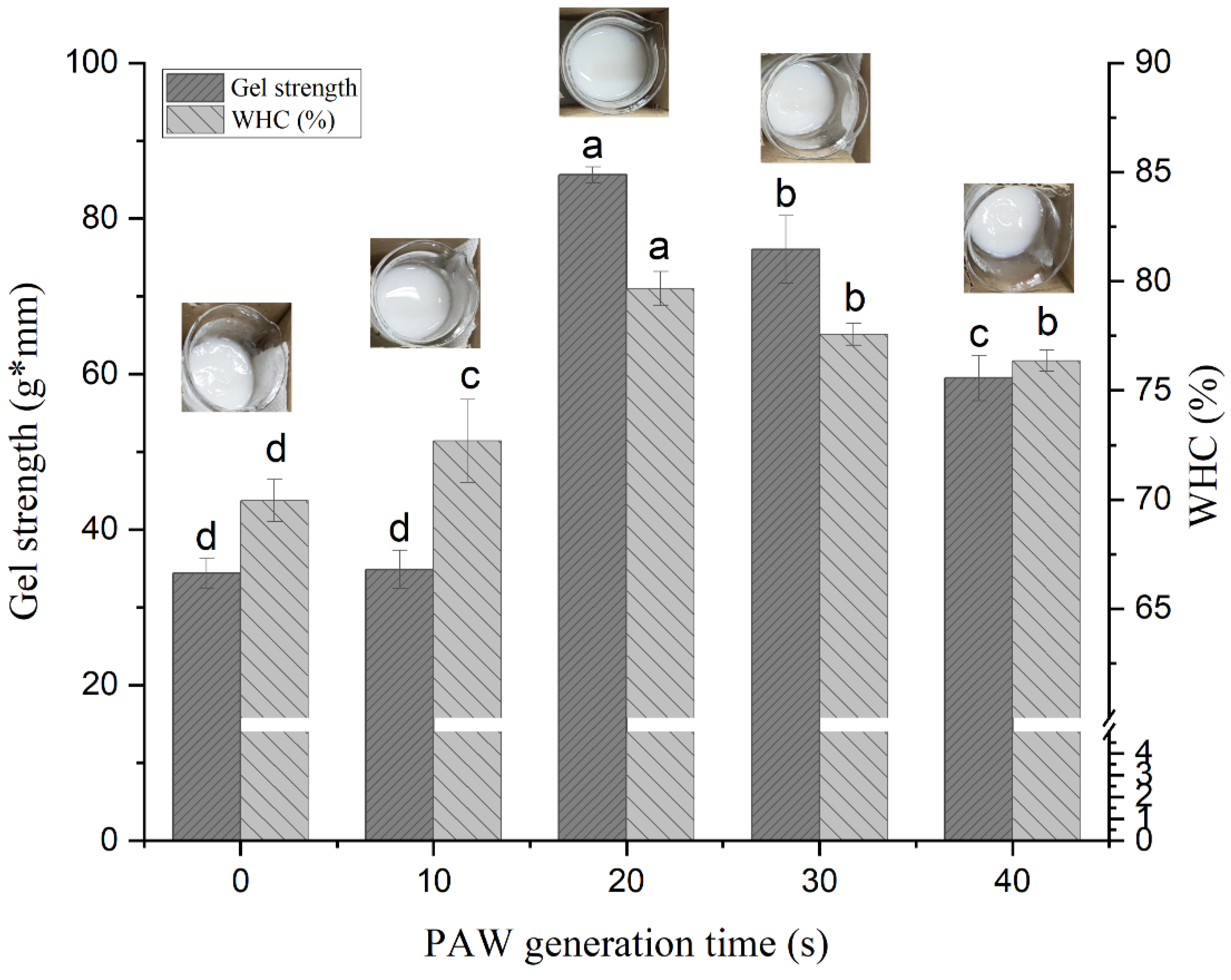
3.1. Gel Strength and WHC
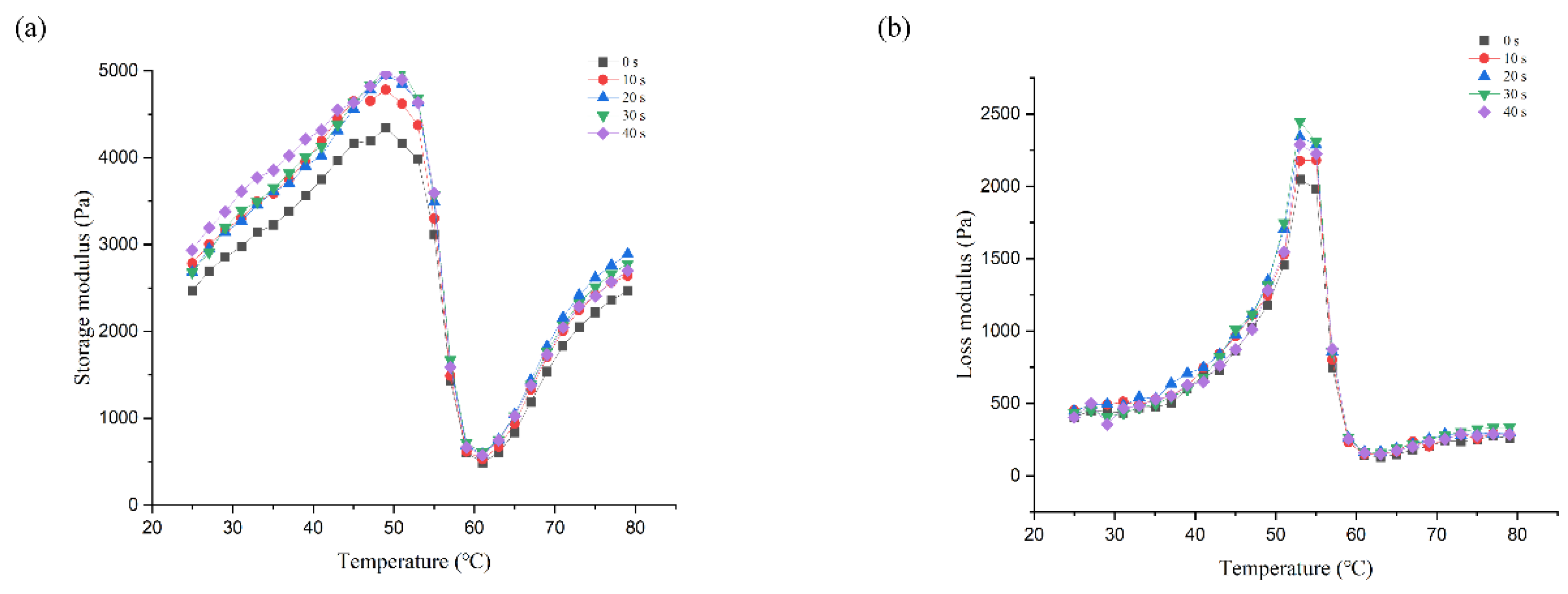
3.2. Dynamic Rheology
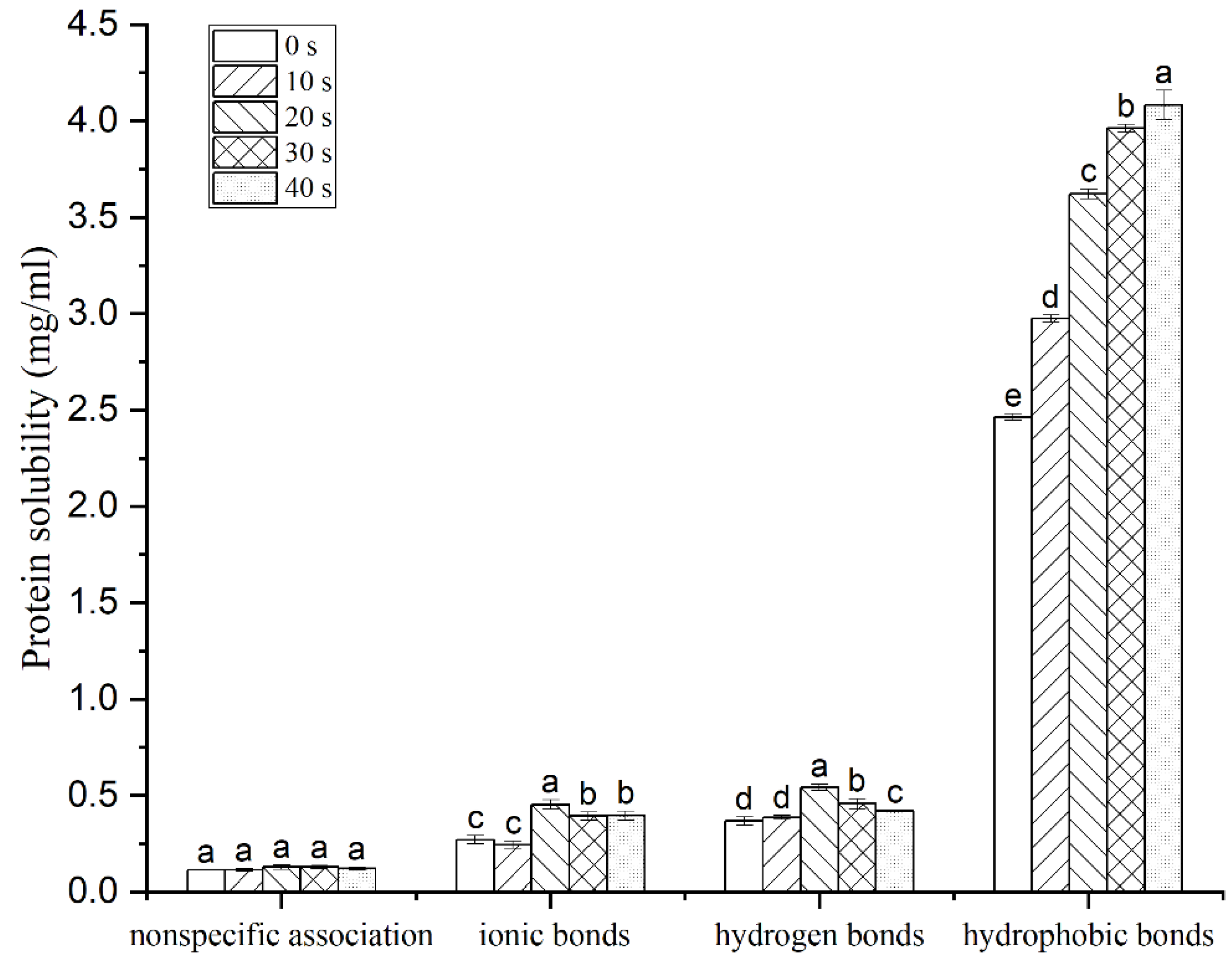
3.3. Molecular Forces
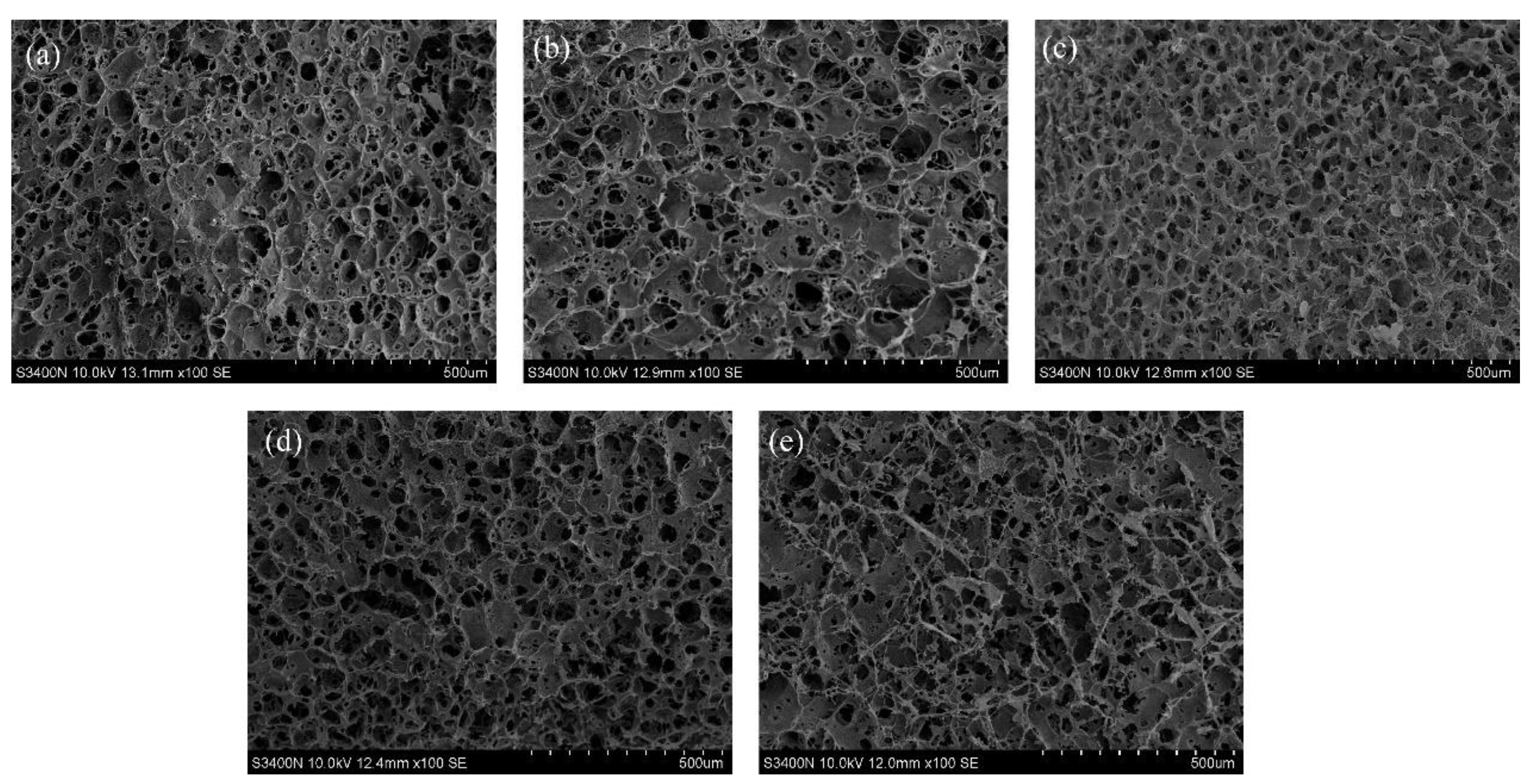
3.4. The Microstructure
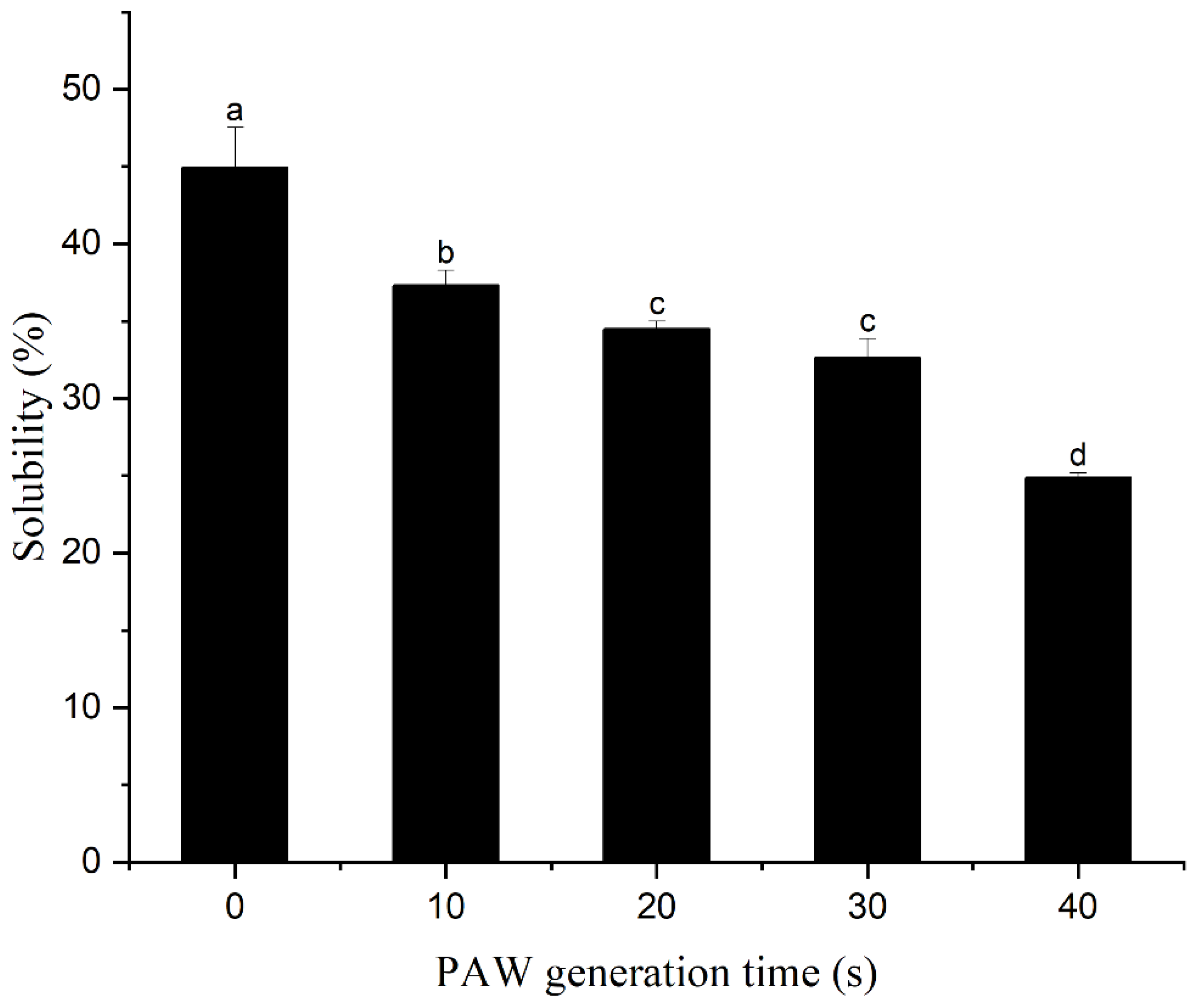
3.5. Solubility
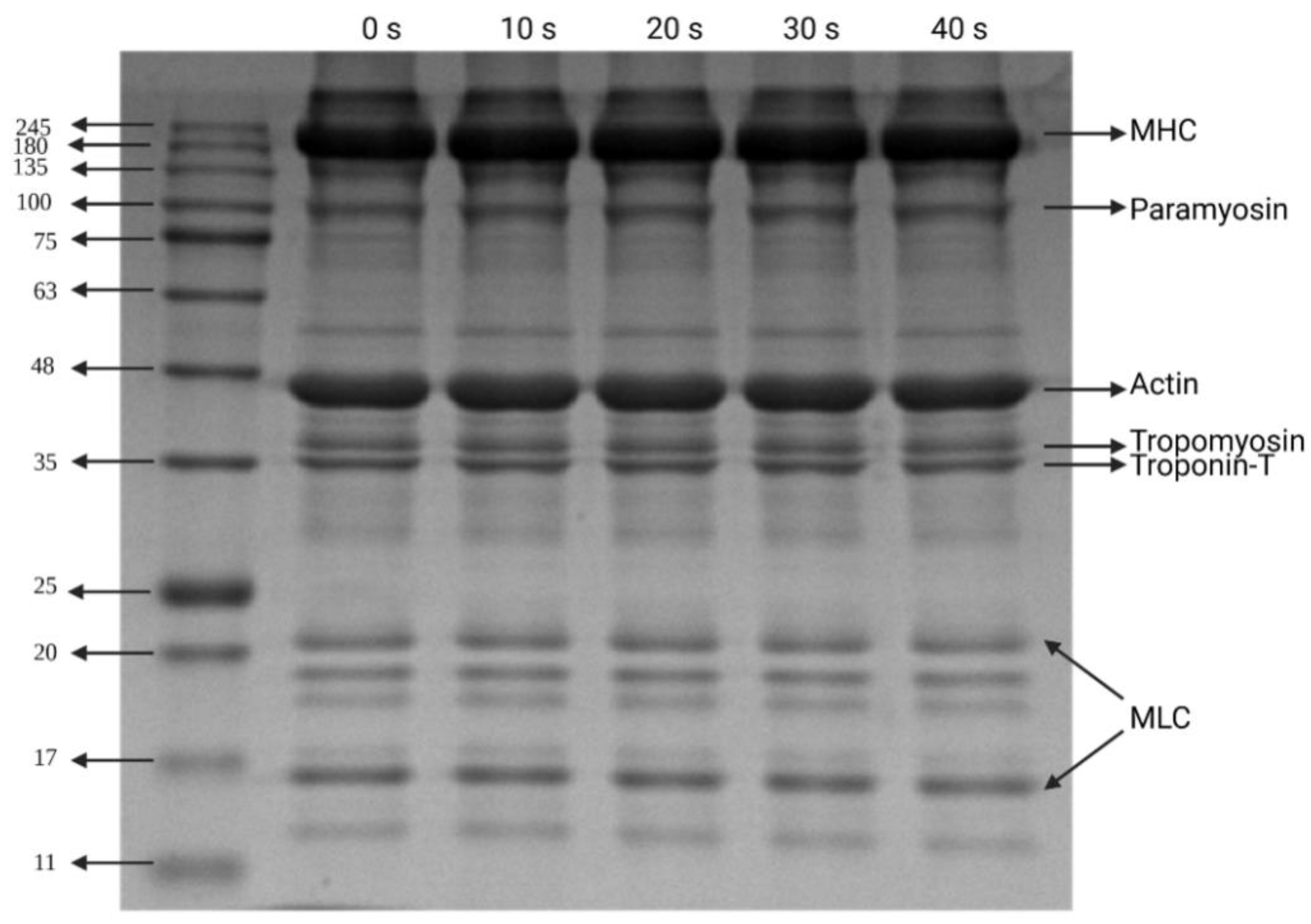
3.6. Electrophoresis Patterns
3.7. Sulfhydryl and Carbonyl Content
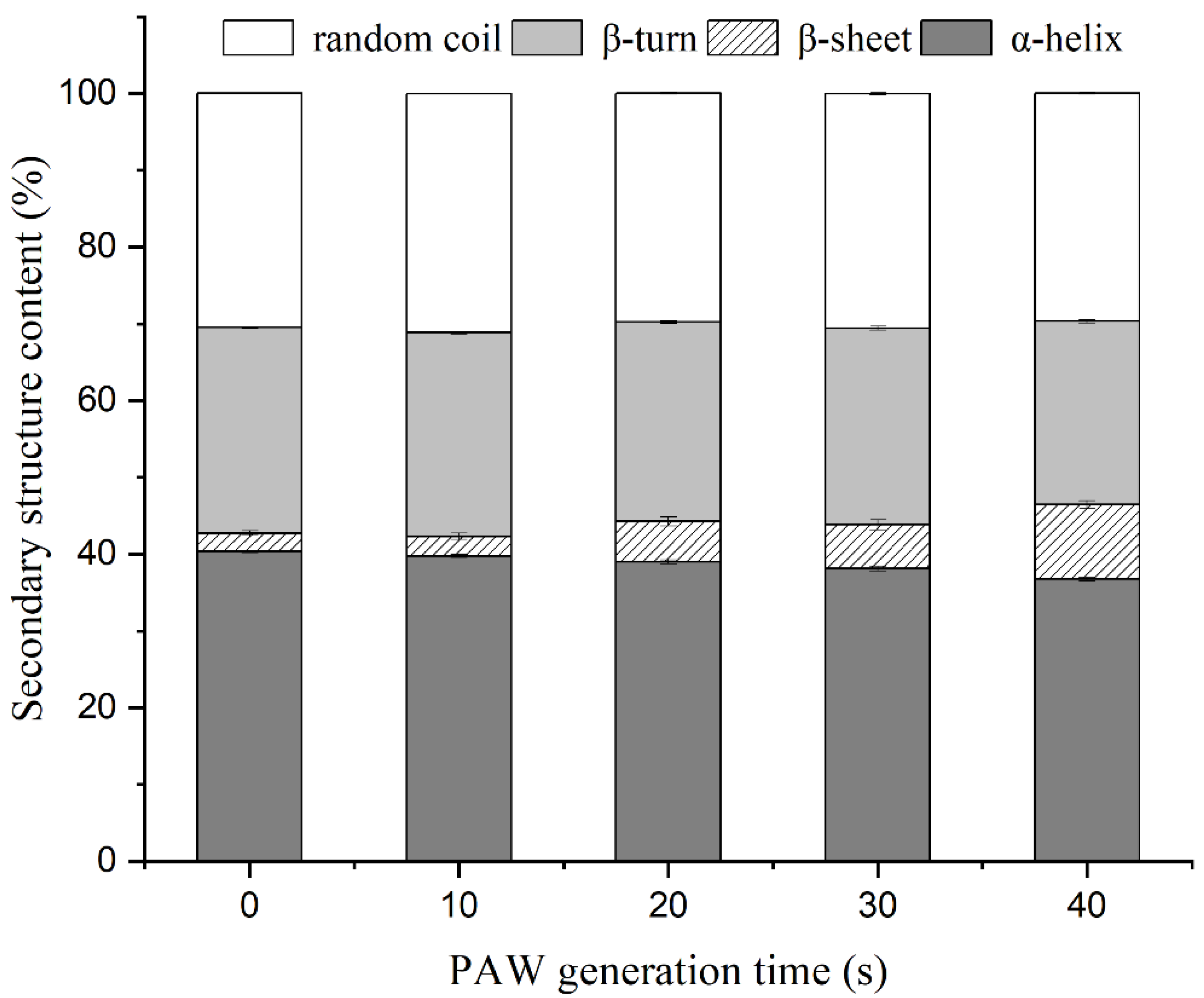
3.8. Secondary Structure Changes of MP
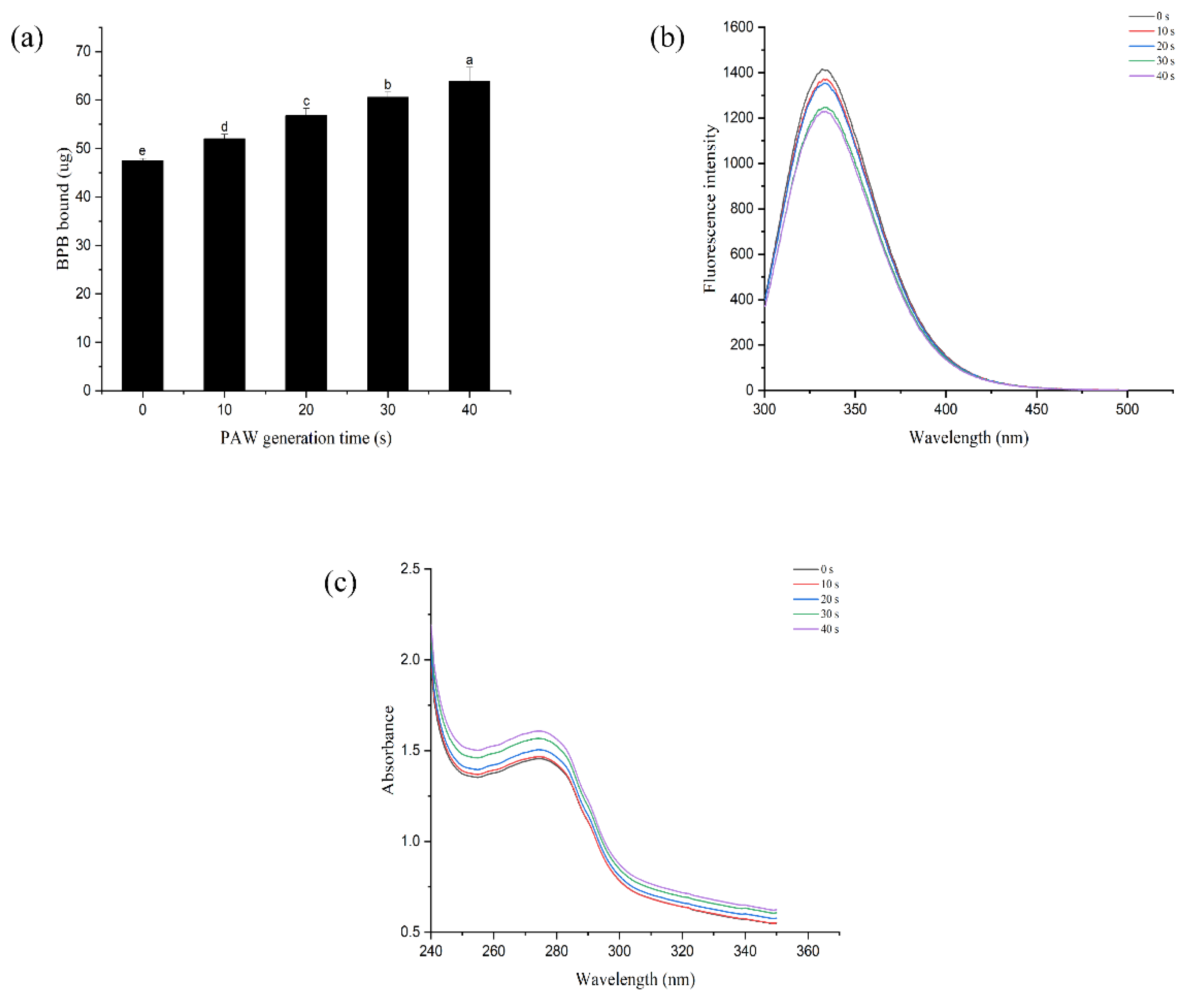
3.9. Tertiary Structure of MP
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Meat, Duck. Available online: http://www.fao.org/faostat/en/#search/duck (accessed on 1 October 2018).
- Biswas, S.; Banerjee, R.; Bhattacharyya, D.; Patra, G.; Das, A.K.; Das, S.K. Technological investigation into duck meat and its products—A potential alternative to chicken. World Poult. Sci. J. 2019, 75, 609–620. [Google Scholar] [CrossRef]
- Cobos, A.; Veiga, A.; Diaz, O. Chemical and fatty acid composition of meat and liver of wild ducks (Anas platyrhynchos). Food Chem. 2000, 68, 77–79. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Y.; Dang, Y.; Cao, J.; Pan, D.; Guo, Y.; He, J. Water-insoluble dietary fibers from oats enhance gel properties of duck myofibrillar proteins. Food Chem. 2021, 344, 128690. [Google Scholar] [CrossRef]
- Xiong, Y.L. Myofibrillar protein from different muscle fiber types: Implications of biochemical and functional properties in meat processing. Crit. Rev. Food Sci. Nutr. 1994, 34, 293–320. [Google Scholar] [CrossRef]
- Wang, P.; Li, Y.; Qu, Y.; Wang, B.; Sun, J.; Miao, C.; Huang, M.; Huang, H.; Zhang, C. Improving gelling properties of myofibrillar proteins incorporating with cellulose micro/nanofibres. Int. J. Food Sci. Technol. 2022, 57, 3428–3439. [Google Scholar] [CrossRef]
- Xia, M.; Chen, Y.; Guo, J.; Feng, X.; Yin, X.; Wang, L.; Wu, W.; Li, Z.; Sun, W.; Ma, J. Effects of oxidative modification on textural properties and gel structure of pork myofibrillar proteins. Food Res. Int. 2019, 121, 678–683. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Zhou, P.; Zhang, X.; Wang, J. Effects of high pressure modification on conformation and gelation properties of myofibrillar protein. Food Chem. 2017, 217, 678–686. [Google Scholar] [CrossRef]
- Jin, S.; Wang, M.; Yang, H.; Shan, A.; Feng, X. Dietary supplementation of resveratrol improved the oxidative stability and spatial conformation of myofibrillar protein in frozen-thawed duck breast meat. Food Biosci. 2021, 43, 101261. [Google Scholar] [CrossRef]
- Khan, M.A.; Ali, S.; Abid, M.; Cao, J.; Jabbar, S.; Tume, R.K.; Zhou, G. Improved duck meat quality by application of high pressure and heat: A study of water mobility and compartmentalization, protein denaturation and textural properties. Food Res. Int. 2014, 62, 926–933. [Google Scholar] [CrossRef]
- Gavahian, M.; Sarangapani, C.; Misra, N.N. Cold plasma for mitigating agrochemical and pesticide residue in food and water: Similarities with ozone and ultraviolet technologies. Food Res. Int. 2021, 141, 110138. [Google Scholar] [CrossRef]
- Roh, S.H.; Oh, Y.J.; Lee, S.Y.; Kang, J.H.; Min, S.C. Inactivation of Escherichia coli O157:H7, Salmonella, Listeria monocytogenes, and Tulane virus in processed chicken breast via atmospheric in-package cold plasma treatment. LWT 2020, 127, 109429. [Google Scholar] [CrossRef]
- Shi, H.; Ileleji, K.; Stroshine, R.L.; Keener, K.; Jensen, J.L. Reduction of aflatoxin in corn by high voltage atmospheric cold plasma. Food Bioprocess Technol. 2017, 10, 1042–1052. [Google Scholar] [CrossRef]
- Hatab, S.; Koddy, J.K.; Miao, W.; Tang, L.; Xu, H.; Deng, S.; Zheng, B. Atmospheric cold plasma: A new approach to modify protein and lipid properties of myofibrillar protein isolate from hairtail (Trichiurus lepturus) fish. J. Sci. Food Agric. 2022, 102, 2041–2049. [Google Scholar] [CrossRef]
- Sharifian, A.; Soltanizadeh, N.; Abbaszadeh, R. Effects of dielectric barrier discharge plasma on the physicochemical and functional properties of myofibrillar proteins. Innov. Food Sci. Emerg. Technol. 2019, 54, 1–8. [Google Scholar] [CrossRef]
- Ekezie, F.-G.C.; Cheng, J.-H.; Sun, D.-W. Effects of atmospheric pressure plasma jet on the conformation and physicochemical properties of myofibrillar proteins from king prawn (Litopenaeus vannamei). Food Chem. 2019, 276, 147–156. [Google Scholar] [CrossRef]
- Luo, J.; Xu, W.; Liu, Q.; Zou, Y.; Wang, D.; Zhang, J. Dielectric barrier discharge cold plasma treatment of pork loin: Effects on muscle physicochemical properties and emulsifying properties of pork myofibrillar protein. LWT 2022, 162, 113484. [Google Scholar] [CrossRef]
- Xu, L.; Yepez, X.; Applegate, B.; Keener, K.M.; Tao, B.; Garner, A.L. Penetration and microbial inactivation by high voltage atmospheric cold plasma in semi-solid material. Food Bioprocess Technol. 2020, 13, 1688–1702. [Google Scholar] [CrossRef]
- Oliveira, M.; Fernandez-Gomez, P.; Alvarez-Ordonez, A.; Prieto, M.; Lopez, M. Plasma-activated water: A cutting-edge technology driving innovation in the food industry. Food Res. Int. 2022, 156, 111368. [Google Scholar] [CrossRef]
- Han, M.; Zhang, Y.; Fei, Y.; Xu, X.; Zhou, G. Effect of microbial transglutaminase on NMR relaxometry and microstructure of pork myofibrillar protein gel. Eur. Food Res. Technol. 2009, 228, 665–670. [Google Scholar] [CrossRef]
- Shi, T.; Xiong, Z.; Jin, W.; Yuan, L.; Sun, Q.; Zhang, Y.; Li, X.; Gao, R. Suppression mechanism of L-arginine in the heat-induced aggregation of bighead carp (Aristichthys nobilis) myosin: The significance of ionic linkage effects and hydrogen bond effects. Food Hydrocoll. 2020, 102, 105596. [Google Scholar] [CrossRef]
- Chen, H.-H.; Xu, S.-Y.; Wang, Z. Interaction between flaxseed gum and meat protein. J. Food Eng. 2007, 80, 1051–1059. [Google Scholar] [CrossRef]
- Wang, X.-X.; Li, Y.-S.; Zhou, Y.; Ma, F.; Li, P.-J.; Chen, C.-G. Effect of resistant corn starch on the thermal gelling properties of chicken breast myosin. Food Hydrocoll. 2019, 96, 681–687. [Google Scholar]
- Li, M.; Shi, T.; Wang, X.; Bao, Y.; Xiong, Z.; Monto, A.R.; Jin, W.; Yuan, L.; Gao, R. Plasma-activated water promoted the aggregation of Aristichthys nobilis myofibrillar protein and the effects on gelation properties. Curr. Res. Food Sci. 2022, 5, 1616–1624. [Google Scholar] [CrossRef]
- GomezGuillen, M.C.; Borderias, A.J.; Montero, P. Chemical interactions of nonmuscle proteins in the network of sardine (Sardina pilchardus) muscle gels. LWT 1997, 30, 602–608. [Google Scholar] [CrossRef]
- Jiang, D.; Shen, P.; Pu, Y.; Jin, M.; Yu, C.; Qi, H. Enhancement of gel properties of Scomberomorus niphonius myofibrillar protein using phlorotannin extracts under UVA irradiation. J. Food Sci. 2020, 85, 2050–2059. [Google Scholar] [CrossRef]
- Kong, B.; Xiong, Y.L. Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action. J. Agric. Food Chem. 2006, 54, 6059–6068. [Google Scholar] [CrossRef]
- Xu, L.; Zheng, Y.; Zhou, C.; Pan, D.; Geng, F.; Cao, J.; Xia, Q. Kinetic response of conformational variation of duck liver globular protein to ultrasonic stimulation and its impact on the binding behavior of n-alkenals. LWT 2021, 150, 111890. [Google Scholar] [CrossRef]
- Ellman, G.L. A colorimetric method for determining low concentrations of mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Zhang, B.; Fang, C.-D.; Hao, G.-J.; Zhang, Y.-Y. Effect of kappa-carrageenan oligosaccharides on myofibrillar protein oxidation in peeled shrimp (Litopenaeus vannamei) during long-term frozen storage. Food Chem. 2018, 245, 254–261. [Google Scholar] [CrossRef]
- Chen, J.; Ying, X.; Deng, S.; Li, W.; Peng, L.; Ma, L. Trehalose and alginate oligosaccharides enhance the stability of myofibrillar proteins in shrimp (Litopenaeus vannamei) muscle during frozen storage. J. Food Process. Preserv. 2022, 46, e16469. [Google Scholar] [CrossRef]
- Li, S.; Li, L.; Zhu, X.; Ning, C.; Cai, K.; Zhou, C. Conformational and charge changes induced by L-Arginine and L-lysine increase the solubility of chicken myosin. Food Hydrocoll. 2019, 89, 330–336. [Google Scholar] [CrossRef]
- Xia, W.; Ma, L.; Chen, X.; Li, X.; Zhang, Y. Physicochemical and structural properties of composite gels prepared with myofibrillar protein and lecithin at various ionic strengths. Food Hydrocoll. 2018, 82, 135–143. [Google Scholar] [CrossRef]
- Nyaisaba, B.M.; Miao, W.; Hatab, S.; Siloam, A.; Chen, M.; Deng, S. Effects of cold atmospheric plasma on squid proteases and gel properties of protein concentrate from squid (Argentinus ilex) mantle. Food Chem. 2019, 291, 68–76. [Google Scholar] [CrossRef]
- Zhou, F.; Zhao, M.; Zhao, H.; Sun, W.; Cui, C. Effects of oxidative modification on gel properties of isolated porcine myofibrillar protein by peroxyl radicals. Meat Sci. 2014, 96, 1432–1439. [Google Scholar] [CrossRef]
- Belton, P.S. NMR and the mobility of water in polysaccharide gels. Int. J. Biol. Macromol. 1997, 21, 81–88. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Kong, B.; Shi, S.; Xia, X. Decreased gelling properties of protein in mirror carp (Cyprinus carpio) are due to protein aggregation and structure deterioration when subjected to freeze-thaw cycles. Food Hydrocoll. 2019, 97, 105223. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, D.; Tu, J.; Zhong, Y.; Zhang, D.; Wang, Z.; Tao, X. Mechanisms of change in gel water-holding capacity of myofibrillar proteins affected by lipid oxidation: The role of protein unfolding and cross-linking. Food Chem. 2021, 344, 128587. [Google Scholar] [CrossRef]
- Sun, Y.; Luo, H.; Cao, J.; Pan, D. Structural characteristics of Sheldrake meat and secondary structure of myofibrillar protein: Effects of oxidation. Int. J. Food Prop. 2017, 20, 1553–1566. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; He, Z.; Gan, X.; Li, H. Effect of peroxyl radicals on the structure and gel properties of isolated rabbit meat myofibrillar proteins. Int. J. Food Sci. Technol. 2018, 53, 2687–2696. [Google Scholar] [CrossRef]
- Xiong, Y.L.; Blanchard, S.P.; Ooizumi, T.; Ma, Y. Hydroxyl radical and ferryl-generating systems promote gel network formation of myofibrillar protein. J. Food Sci. 2010, 75, C215–C221. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Extreme pH treatments enhance the structure-reinforcement role of soy protein isolate and its emulsions in pork myofibrillar protein gels in the presence of microbial transglutaminase. Meat Sci. 2013, 93, 469–476. [Google Scholar] [CrossRef]
- Xia, T.; Xu, Y.; Zhang, Y.; Xu, L.; Kong, Y.; Song, S.; Huang, M.; Bai, Y.; Luan, Y.; Han, M.; et al. Effect of oxidation on the process of thermal gelation of chicken breast myofibrillar protein. Food Chem. 2022, 384, 132368. [Google Scholar] [CrossRef]
- Chanarat, S.; Benjakul, S.; Xiong, Y.L. Physicochemical changes of myosin and gelling properties of washed tilapia mince as influenced by oxidative stress and microbial transglutaminase. J. Food Sci. Technol. Mys. 2015, 52, 3824–3836. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Wang, Y.; Zhuang, H.; Yan, W.; Zhang, J.; Luo, J. Plasma activated water-induced formation of compact chicken myofibrillar protein gel structures with intrinsically antibacterial activity. Food Chem. 2021, 351, 129278. [Google Scholar] [CrossRef]
- Herrero, A.M.; Cambero, M.; Ordonez, J.A.; Castejon, D. Magnetic resonance imaging, rheological properties, and physicochemical characteristics of meat systems with fibrinogen and thrombin. J. Agric. Food Chem. 2007, 55, 9357–9364. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, W.; Huang, J.; Xiong, Y.L. Effects of sodium pyrophosphate coupled with catechin on the oxidative stability and gelling properties of myofibrillar protein. Food Hydrocoll. 2020, 104, 105722. [Google Scholar] [CrossRef]
- Jiang, S.; Zhao, S.; Jia, X.; Wang, H.; Zhang, H.; Liu, Q.; Kong, B. Thermal gelling properties and structural properties of myofibrillar protein including thermo-reversible and thermo-irreversible curdlan gels. Food Chem. 2020, 311, 126018. [Google Scholar] [CrossRef]
- Ni, N.; Wang, Z.; He, F.; Wang, L.; Pan, H.; Li, X.; Wang, Q.; Zhang, D. Gel properties and molecular forces of lamb myofibrillar protein during heat induction at different pH values. Process Biochem. 2014, 49, 631–636. [Google Scholar] [CrossRef]
- Mi, J.; Ni, W.; Huang, P.; Hong, J.; Jia, R.; Deng, S.; Yu, X.; Wei, H.; Yang, W. Effect of acetylated distarch adipate on the physicochemical characteristics and structure of shrimp (Penaeus vannamei) myofibrillar protein. Food Chem. 2022, 373, 131530. [Google Scholar] [CrossRef]
- Liu, R.; Zhao, S.-M.; Xie, B.-J.; Xiong, S.-B. Contribution of protein conformation and intermolecular bonds to fish and pork gelation properties. Food Hydrocoll. 2011, 25, 898–906. [Google Scholar] [CrossRef]
- Cong-Gui, C.; Gerelt, B.; Shao-Tong, J.; Nishiumi, T.; Suzuki, A. Effects of high pressure on pH, water-binding capacity and textural properties of pork muscle gels containing various levels of sodium alginate. Asian-Australas J. Anim. Sci. 2006, 19, 1658–1664. [Google Scholar]
- Du, J.; Zhou, C.; Xia, Q.; Wang, Y.; Geng, F.; He, J.; Sun, Y.; Pan, D.; Cao, J. The effect of fibrin on rheological behavior, gelling properties and microstructure of myofibrillar proteins. LWT 2022, 153, 112457. [Google Scholar] [CrossRef]
- Wang, L.; Xia, M.; Zhou, Y.; Wang, X.; Ma, J.; Xiong, G.; Wang, L.; Wang, S.; Sun, W. Gel properties of grass carp myofibrillar protein modified by low-frequency magnetic field during two-stage water bath heating. Food Hydrocoll. 2020, 107, 105920. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Wang, J.-Z.; Zhang, C.-H.; Sun, H.-M.; Wang, C.-Q.; Xie, X.-L. Effects of oxidation on water distribution and physicochemical properties of porcine myofibrillar protein Gel. Food Biophys. 2014, 9, 169–178. [Google Scholar] [CrossRef]
- Cao, H.; Fan, D.; Jiao, X.; Huang, J.; Zhao, J.; Yan, B.; Zhou, W.; Zhang, W.; Zhang, H. Effects of microwave combined with conduction heating on surimi quality and morphology. J. Food Eng. 2018, 228, 1–11. [Google Scholar] [CrossRef]
- Wang, H.; Pei, Z.; Xue, C.; Cao, J.; Shen, X.; Li, C. Comparative study on the characterization of myofibrillar proteins from Tilapia, Golden Pompano and Skipjack Tuna. Foods 2022, 11, 1705. [Google Scholar] [CrossRef]
- Panpipat, W.; Chaijan, M. Effect of atmospheric pressure cold plasma on biophysical properties and aggregation of natural actomyosin from Threadfin Bream (Nemipterus bleekeri). Food Sci. Biotechnol. 2020, 13, 851–859. [Google Scholar] [CrossRef]
- Wang, H.; Song, Y.; Liu, Z.; Li, M.; Zhang, L.; Yu, Q.; Guo, Z.; Wei, J. Effects of iron-catalyzed and metmyoglobin oxidizing systems on biochemical properties of yak muscle myofibrillar protein. Meat Sci. 2020, 166, 108041. [Google Scholar] [CrossRef]
- Hou, H.; He, H.; Wang, Y. Effects of SDS on the activity and conformation of protein tyrosine phosphatase from thermus thermophilus HB27. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Miao, W.; Nyaisaba, B.M.; Koddy, J.K.; Chen, M.; Hatab, S.; Deng, S. Effect of cold atmospheric plasma on the physicochemical and functional properties of myofibrillar protein from Alaska pollock (Theragra chalcogramma). Int. J. Food Sci. Technol. 2020, 55, 517–525. [Google Scholar] [CrossRef]
- Utrera, M.; Estevez, M. Oxidation of myofibrillar proteins and impaired functionality: Underlying mechanisms of the carbonylation pathway. J. Agric. Food Chem. 2012, 60, 8002–8011. [Google Scholar] [CrossRef]
- Requena, J.R.; Chao, C.C.; Levine, R.L.; Stadtman, E.R. Glutamic and aminoadipic semialdehydes are the main carbonyl products of metal-catalyzed oxidation of proteins. Proc. Natl. Acad. Sci. USA. 2001, 98, 69–74. [Google Scholar] [CrossRef]
- Lund, M.N.; Heinonen, M.; Baron, C.P.; Estevez, M. Protein oxidation in muscle foods: A review. Mol. Nutr. Food Res. 2011, 55, 83–95. [Google Scholar] [CrossRef]
- Estevez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Choi, S.-M.; Ma, C.-Y. Structural characterization of globulin from common buckwheat (Fagopyrum esculentum Moench) using circular dichroism and Raman spectroscopy. Food Chem. 2007, 102, 150–160. [Google Scholar]
- Guo, M.; Liu, S.; Ismail, M.; Farid, M.M.; Ji, H.; Mao, W.; Gao, J.; Li, C. Changes in the myosin secondary structure and shrimp surimi gel strength induced by dense phase carbon dioxide. Food Chem. 2017, 227, 219–226. [Google Scholar] [CrossRef]
- Amiri, A.; Sharifian, P.; Morakabati, N.; Mousakhani-Ganjeh, A.; Mirtaheri, M.; Nilghaz, A.; Guo, Y.-G.; Pratap-Singh, A. Modification of functional, rheological and structural characteristics of myofibrillar proteins by high-intensity ultrasonic and papain treatment. Innov. Food Sci. Emerg. Technol. 2021, 72, 102748. [Google Scholar] [CrossRef]
- Han, Z.; Cai, M.-j.; Cheng, J.-H.; Sun, D.-W. Effects of microwave and water bath heating on the interactions between myofibrillar protein from beef and ketone flavour compounds. Int. J. Food Sci. Technol. 2019, 54, 1787–1793. [Google Scholar] [CrossRef]
- Tan, C.; Xu, Q.-D.; Chen, N.; He, Q.; Sun, Q.; Zeng, W.-C. Cross-linking effects of EGCG on myofibrillar protein from common carp (Cyprinus carpio) and the action mechanism. J. Food Biochem. 2022, 46, e14416. [Google Scholar] [CrossRef]
- Gao, M.-R.; Xu, Q.-D.; Zeng, W.-C. Effect of tea polyphenols on the tenderness of yak meat. J. Food Process. Preserv. 2020, 44, e14433. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, W.; Feizollahi, E.; Roopesh, M.S.; Chen, L. Improvement of pea protein gelation at reduced temperature by atmospheric cold plasma and the gelling mechanism study. Innov. Food Sci. Emerg. Technol. 2021, 67, 102567. [Google Scholar] [CrossRef]
- Fu, Q.; Liu, R.; Wang, H.; Hua, C.; Song, S.; Zhou, G.; Zhang, W. Effects of oxidation in vitro on structures and functions of myofibrillar protein from beef muscles. J. Agric. Food Chem. 2019, 67, 5866–5873. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Liu, Q.; Chen, Q.; Kong, B. Solubilization and stable dispersion of myofibrillar proteins in water through the destruction and inhibition of the assembly of filaments using high-intensity ultrasound. Ultrason. Sonochem. 2020, 67, 105160. [Google Scholar] [CrossRef]
- Li, F.; Wang, B.; Liu, Q.; Chen, Q.; Zhang, H.; Xia, X.; Kong, B. Changes in myofibrillar protein gel quality of porcine longissimus muscle induced by its stuctural modification under different thawing methods. Meat Sci. 2019, 147, 108–115. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, Y.; Chen, L.; Wu, H.; Zhang, Y.; Suo, Z.; Wang, S.; Liang, Y.; Xu, X.; Zhou, G.; et al. Inhibition of epigallocatechin-3-gallate/protein interaction by methyl-beta-cyclodextrin in myofibrillar protein emulsion gels under oxidative stress. J. Agric. Food Chem. 2018, 66, 8094–8103. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, W.; Roopesh, M.S.; Chen, L. Pre-treatment by combining atmospheric cold plasma and pH-shifting to prepare pea protein concentrate powders with improved gelling properties. Food Res. Int. 2022, 154, 111028. [Google Scholar] [CrossRef]
- Zhu, W.; Huan, H.; Bu, Y.; Li, X.; Shiuan, D.; Li, J.; Sun, X. Effects of hydroxyl radical induced oxidation on water holding capacity and protein structure of jumbo squid (Dosidicus gigas) mantle. Int. J. Food Sci. Technol. 2019, 54, 2159–2168. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Li, S.; Oladejo, A.O.; Wang, Y.; Huang, S.; Zhou, C.; Wang, Y.; Mao, L.; Zhang, Y.; et al. Effects of low power density multi-frequency ultrasound pretreatment on the enzymolysis and the structure characterization of defatted wheat germ protein. Ultrason. Sonochem. 2017, 38, 410–420. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAW Generation Time (s) | 0 | 10 | 20 | 30 | 40 |
|---|---|---|---|---|---|
| Sulfhydryl groups (nmol/mg protein) | 44.58 ± 0.92 bc | 45.73 ± 0.18 ab | 47.12 ± 0.71 a | 45.17 ± 0.91 b | 43.08 ± 1.54 c |
| Carbonyl content (nmol/mg protein) | 1.14 ± 0.05 e | 1.44 ± 0.06 d | 1.78 ± 0.02 c | 2.07 ± 0.06 b | 2.44 ± 0.14 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, W.; Roopesh, M.S.; Pan, D.; Du, L. Enhanced Gel Properties of Duck Myofibrillar Protein by Plasma-Activated Water: Through Mild Structure Modifications. Foods 2023, 12, 877. https://doi.org/10.3390/foods12040877
Rao W, Roopesh MS, Pan D, Du L. Enhanced Gel Properties of Duck Myofibrillar Protein by Plasma-Activated Water: Through Mild Structure Modifications. Foods. 2023; 12(4):877. https://doi.org/10.3390/foods12040877
Chicago/Turabian StyleRao, Wei, M. S. Roopesh, Daodong Pan, and Lihui Du. 2023. "Enhanced Gel Properties of Duck Myofibrillar Protein by Plasma-Activated Water: Through Mild Structure Modifications" Foods 12, no. 4: 877. https://doi.org/10.3390/foods12040877
APA StyleRao, W., Roopesh, M. S., Pan, D., & Du, L. (2023). Enhanced Gel Properties of Duck Myofibrillar Protein by Plasma-Activated Water: Through Mild Structure Modifications. Foods, 12(4), 877. https://doi.org/10.3390/foods12040877