1. Introduction
In 1884, Theobald Smith isolated
Hog-cholerabacillus from pigs’ intestines infected with swine fever and named it
Salmonella after his mentor, Daniel Elmer Salmon, an American veterinary pathologist.
Salmonella is one of the common bacterial enteropathogens responsible for sporadic illness or widespread gastrointestinal disease [
1,
2,
3,
4]. Up to 80% of human
Salmonella infections are not associated with a known outbreak; instead, they are considered a sporadic illness, and some are not diagnosed [
5]. The World Health Organization estimated 153 million non-typhoidal
Salmonella (NTS) infections worldwide in 2010, of which 56,969 were lethal and nearly half were foodborne [
6]. A total of 926 salmonellosis foodborne outbreaks were reported across 23 European countries in 2019, resulting in 9169 cases, 1915 hospitalizations, and 7 deaths. Salmonellae were responsible for 17.9% (one in six) of all foodborne outbreaks in 2019 [
7].
Salmonella is considered the most common foodborne organism in imported foods from Africa to the European Union. A large proportion (72.4%) of the foodborne salmonellosis outbreaks were caused by
Salmonella enterica serovar Enteritidis [
8]. The two most common serovars in the United States are
Salmonella enterica serovar Typhimurium (
S. Typhimurium) and serovar Enteritidis (
S. Enteritidis), and these were responsible for 41.5% of the total outbreaks. These two serovars account for nearly 60% of all
Salmonella outbreaks worldwide [
9] and 91% of African cases [
10].
Antimicrobial resistance in
Salmonella is a significant concern for public health safety [
11,
12]. This pathogen must receive more attention, particularly its presence in the food/feed supply chain [
13]. Food animals are often reservoir hosts for
Salmonella; thus, it is difficult to eradicate this organism from them [
14,
15].
Outbreak studies have recognized food commodities, including fresh produce, vegetables, cantaloupes, cereals, alfalfa sprouts, fruit/fruit pulp, pistachios, poultry meat, turkeys, tuna, ground beef, pork, dried/shredded coconut, tomatoes, and mangoes, as sources of foodborne
Salmonella-related outbreaks in the past decade [
16,
17]. The magnitude of infection that has been provoked due to antibiotic-resistant variants is recurrently implied as the causative organisms in these outbreaks, resulting in a higher level of systemic diseases, treatment failures, and a surge in hospitalizations [
18,
19].
This review provides an updated summary of the nomenclature, taxonomy, microbial properties, pathogenesis, transmission, and antimicrobial resistance of human salmonellosis. The recent outbreaks of human salmonellosis reported in different clinical and geographical settings, including the farm-to-table food supply continuum, are also presented.
2. Materials and Methods
Medline (PubMed), Web of Science, Embase, and Google Scholar were searched for relevant articles. The search string that enabled us to locate most studies was Salmonella AND (Africa OR Asia OR Europe OR Central America OR South America OR USA OR Middle East. Additional searches involved using the main medical subject headings (MeSH) terms with Boolean operators and other words that included epidemiology, transmission, virulence, pathogenesis, clinical manifestations, foodborne, and drug resistance. We performed the last search on 21 January 2023. The full texts or abstracts of all articles were screened for eligibility. Duplicates or research articles that did not meet the eligibility criteria were excluded. Studies that were non-English (language), nonbacterial, non-salmonella (organism), or nonhuman (host) were also omitted.
3. Classification and Nomenclature
The nomenclature and naming of
Salmonella are relatively complex and are still evolving. Based on variations in the sequence of the 16S rRNA gene, the
Salmonella genus is classified into two species,
Salmonella bongori and
Salmonella enterica (type species).
S. enterica can be categorized into six subspecies depending on their biochemical properties and genetic relationship [
20,
21,
22]. The subspecies are designated with Roman numerals, as follows: I,
S. enterica subsp.
enterica; II,
S. enterica subsp.
salamae; IIIa,
S. enterica subsp.
arizonae; IIIb,
S. enterica subsp.
diarizonae; IV,
S. enterica subsp.
houtenae; VI,
S. enterica subsp.
indica.
Salmonella enterica subsp.
enterica (I) is the predominant subspecies in mammals, accounting for around 99% of human salmonellosis and
Salmonella infections in warm-blooded animals. On the contrary,
S. bongori and the other five subspecies are predominant in cold-blooded animals and the environment [
23].
Besides phylogenetic classification, Kauffman and White developed a system of identifying
Salmonella serotypes using three chief antigenic determining factors: flagellar (H), capsule (K), and somatic (O) antigens [
23] (
Figure 1). The O antigen is the heat-stable form of lipopolysaccharides found on the outer membrane. Some salmonellae serotype express one or more O antigen types [
24]. H antigens represented by bacterial flagella stimulate the host’s immune response. Most
Salmonella serovars contain two definite genomic regions that encode flagella synthesis. They have the unique capability to express only one of these proteins at a time, which is why they are called diphasic (phase I and II) bacteria. Phase I H antigens account for the immunological character of a serotype, while phase II antigens are common to many serotypes [
25]. K antigens are polysaccharides that are sensitive to heat and attach to the surface of bacterial capsules. It is the least abundant antigen in all serotypes [
26]. A particular subtype of the K-antigen called the virulence (Vi) antigen is detected only in three pathogenic serotypes (not all strains), namely Dublin, Typhi, and Paratyphi C [
27].
The
Salmonella serotypes can be identified by analyzing all the antigenic properties of the bacterium. An agglutination test using antibodies unique to the O antigens can be performed to classify the organism into six serogroups, namely A, B, C1, C2, D, and E. This approach offers crucial evidence for epidemiological study and enables genus and/or species-level identification [
28]. Thus far, more than 2500 serovars have been identified; more than half of these serotypes are part of
S. enterica subsp enterica, which is responsible for most human salmonellosis [
29]. The taxonomy and nomenclature of
Salmonella should follow a standard format (i.e.,
Salmonella enterica subsp
enterica serovar Typhimurium, which is abbreviated as
S. Typhimurium) while describing them in the literature [
30].
More recently, Chattaway et al. [
31] proposed a taxonomic classification based on whole genome sequence data to define a two-tier subtyping approach: multilocus sequence typing (MLST) followed by antigen prediction. Using this approach, the researchers classified 46,000 isolates of
Salmonella enterica subspecies
enterica that showed a 99.96% match to a serovar organized by the Kauffmann and White scheme.
4. Pathogenesis
The severity of human salmonellosis depends on the causative serotypes and the immune health of the patient or the presence of comorbid diseases in the infected individuals. The aging population, children under 5 years, and immunodeficient individuals are more vulnerable to
Salmonella infections.
Salmonella showed an unusual characteristic in the course of its colonization of non-phagocytic cells [
32], whereby it triggers phagocytosis (referred to as trigger mechanism) to be able to enter the host cell (
Figure 2A). These virulence factors are situated in
Salmonella pathogenicity islands (SPIs), the cluster of genes positioned at the large region of the chromosomal DNA and standing for the structural domains that participate in the invasion process [
33]. The bacterium tends to penetrate the intestinal wall epithelial cells after entering the digestive system via contaminated water or food. The transportation of
Salmonella via the barrier of the intestine arises mainly through the specialized microfold (M) cells (
Figure 2B) positioned in the lymphoid tissue, also named Peyer’s patches [
34] or by active penetration of nonphagocytic cells, via the so-called “trigger” process [
35] (
Figure 2C). The type III secretion system (T3SS) is the cell surface multi-protein channel, which allows the bacterium to deliver its effector molecules (payload) into the host cytosol [
36]. The effectors then activate the signal transduction system and induce the rearrangement of the host actin cytoskeleton, leading to the outward projection of the membrane of the epithelial cell to internalize the bacterium. The structure of the projected membrane resembles the process of phagocytosis, which is conducted by normal phagocytic cells [
37] (
Figure 2C).
The intracellular persistence of
Salmonella is vital for pathogenesis, which may vary among hypervirulent to low-virulence strains [
38,
39]. After the engulfment, the bacterium is encased in a host-derived sheath of a membrane known as a vacuole (also named a
Salmonella-containing vacuole, SCV) (
Figure 2A). The host cell then triggers the fusion of the phagosome with lysosomes and the production of enzymes and reactive oxygen species to destroy the captured bacteria [
40]. The bacterium injects effector proteins directly into the vacuole using the T3SS, resulting in the compartment’s structural modification. The modified vacuole structure stops the phagolysosomal fusion, which protects the organism in the intracellular niche to promote safe replication (
Figure 2A). The ability of this organism to evade macrophages enables the bacterium to be transported in the reticuloendothelial system (RES) [
41,
42].
4.1. Salmonella Virulence Factors
4.1.1. Pathogenicity Islands
There have been 24
Salmonella pathogenicity islands (SPIs) identified thus far. These horizontally obtained loci encode genes assisting many virulence factors, including (i) secretion systems, flagella, fimbriae, and capsule synthesis, (ii) colonization of the host cell, (iii) host evasion mechanisms and survival, and (iv) seroconversion [
43]. Among the SPIs, approximately 21 SPIs are typically found in
S. enterica, encoding genes required for pathogenesis, survival inside the host, and maintenance of housekeeping genes [
44,
45]. The genetic and phenotypic characteristics of
Salmonella pathogenicity islands 1 (SPI-1) and 2 (SPI-2) have been well-researched. SPI-1 is commonly found in all species and subspecies [
46]. T3SS encoded by SPI-1 is involved in the export of effector proteins for host cell colonization. Contrary to this, SPI-2 is present only in
S.
enterica, while SPI-22 is found only in
S. bongori [
46]. The SPI-2 gene encodes an additional T3SS critical for survival at the intracellular niche and for preventing SCV acidification.
Several SPIs have been associated with
S. enterica subsp
enterica serovars, and some have been reported to confer fitness advantages on specific serovars. Although SPI-7 encodes the Vi capsule, it was traditionally thought to appear exclusively in strains of
S. Typhi [
47]; however, it has also been found in NTS serovar Dublin [
48,
49]. These two serovars have been associated with invasive diseases in humans. Some serovars carry typhoid toxin genes on SPI-11, even though SPI-11 is common among
S. enterica subsp
enterica [
50]. Researchers have reported that carriers of the typhoid toxin gene
cdtB (cytolethal distending toxin) are more likely to develop invasive diseases [
51]. It has also been proposed that certain SPIs can help colonize a specific host. T6SS located in SPI-19 was shown to be responsible for
S. Gallinarum and
S. Typhimurium colonization and spread to deeper tissues in chickens [
52].
4.1.2. Toxins
Bacterial toxins that are released extracellularly and act on the target host cells are exotoxins. The toxin produced by serotype Typhi, designated the “typhoid toxin”, is responsible for the pathogenesis and typhoid symptoms [
53]. Typhoid toxin is also called
Salmonella cdt, and at least 41 NTS serovars have been reported to harbor this genotoxin [
50,
54]. However, it has not been found in serovars Enteritidis, Typhimurium, or Newport. Scientists have discovered that the
S. Typhi toxin helps to fine-tune the host’s defense against infection by marking specific target cells, including immune and brain cells [
55].
S. Typhimurium DT-104 and other NTS strains also encode an exotoxin called ArtAB toxin (ADP-ribosylating toxin) [
51]. The ArtA and ArtB subunits form the active and pentameric binding subunits, respectively [
56]. These two subunits share homology with pertussis toxin, produced by
Bordetella pertussis [
56].
Several reports indicate that various NTS serovars also secrete trypsin-sensitive, heat-labile cytotoxins. In some cases, the cytotoxic effect is related to the outer membrane. It has been reported that some serovars are capable of producing cytotoxins, including Nchanga, Enteritidis, Saintpaul, Braenderup, Indiana, Choleraesuis, Virchow, and Typhimurium [
57].
4.1.3. Flagella
Flagella are a locomotory cellular appendage in most NTS [
43,
58], except for serovar Gallinarum biovars Pullorum and Gallinarum [
59]. Research findings indicate the involvement of 50 or more genes in synthesizing flagella assembly and maintenance [
60]. Nevertheless, the building block, flagellin, is encoded individually by
fliC, fljB, and
flpA genes for phases 1, 2, and 3, respectively [
61]. Most NTS serovars express 5–10 peritrichous flagella [
61]. Phase variation is one of the crucial characteristics of different
Salmonella serovars, which is a genetic reshuffle (reversible) to switch between the expression of
fliC and
fljB. This method is developed by several bacterial pathogens [
62,
63].
Flagella help
Salmonella to move toward the host epithelial layers and are also a potent inducer of the innate immune response in the host [
64]. Additionally, it has been demonstrated that flagella enable the organism to move toward the tetrathionate and nitrate generated by the host microbiota, which is utilized as alternative terminal electron acceptors for survival inside the host gut [
65,
66].
4.1.4. Fimbriae (Pili)
A thin bacterial cell surface appendage called fimbriae aids the organism in adhering and attaching to the host cell. This structure is produced by several gram-positive and gram-negative bacteria [
67]. Genomic and phenotypic investigations have identified the 39 distinct fimbrial operons in
Salmonella. Of all operons,
agf is found in both
S. bongori and
S. enterica and encodes the nucleator-dependent curli fimbriae, which are aggregative thin structures that help in firm attachment and colonization processes [
68]. The type IV fimbriae are encoded by
pil and
bfp operons; the former operon is present on SPI-7. Hence, it is limited to serovar Paratyphi C, Typhi, and Dublin [
48]. The other 36 fimbrial operons encode chaperone-usher-dependent fimbrial (CUDF) pathways [
68]. Twenty-seven fimbrial operons have been detected in NTS [
68]. The
fim operon is the only CUDF operon found in all
S. enterica isolates [
68]. Many fimbrial genes are not expressed in in vitro environments but are detected during in vivo infection [
69].
4.1.5. Protein Secretion System
Presently, four protein secretion systems have been identified in
Salmonella, including the type I (T1SS), III (T3SS), IV (T4SS), and VI (T6SS) secretion systems (
Figure 3). According to reports, these systems play a pivotal role in host infection [
70]. They are mainly used to transport and translocate the bacterial effector proteins and to transport some (T3SS and T4SS) directly into the target cell cytoplasm, and to modulate signaling cascades of the host cell [
71,
72,
73]. In the host cytosol, the effector proteins can change normal cellular activities, such as membrane transport, cytoskeleton structure, cytokine expression, and signal transduction [
74]. The secretion apparatus contains numerous components comprising over 20 different types of proteins [
75]. Some proteins are homologous to the proteins involved in flagellar assembly, indicating an evolutionary relationship between the two [
76].
Among all the secretion systems, T3SSs are the most advanced and well-studied secretion system in bacteria [
75,
77,
78].
Salmonella outcompetes the gut commensal bacteria with the help of T6SS. T1SS and T3SS-1 trigger this organism’s adherence to the intestine’s epithelial layer. After that, they facilitate invasion of M cells or epithelial cells by releasing effectors via T3SS-1 and T6SS. Once internalized, T3SS-2, T4SS, and T6SS promote the replication and survival of
Salmonella in SCVs [
70].
4.1.6. Biofilm Production
Besides the planktonic phase,
Salmonella sometimes exists as a sessile or multicellular form (also known as biofilm) that enable the agent to firmly adhere to biotic and/or abiotic surfaces [
79,
80,
81]. Biofilm formation has been linked to
Salmonella persistence on food surfaces, plants, and other fresh produce, and biofilm protects the pathogen during food processing [
82]. Forming biofilm contributes to the dissemination of
Salmonella, since microbes in the biofilm are protected from chemical, mechanical, and physical stressors [
79]. It also contributes to the virulence of
Salmonella, since bacteria in the biofilm are not vulnerable to the host’s immune attack and antimicrobials, leading to the long-term survival of the agent, which ultimately results in chronic infection and carrier state [
83,
84]. The biofilm formed by
Salmonella is mainly made up of O antigen, curli (amyloid fimbriae), biofilm-associated protein (Bap), cellulose, and extracellular DNA [
79,
80,
85]. Curlin subunit gene D (CsgD), regulated by transcriptional factors, bis-3′-5′-cyclic dimeric guanosine monophosphate (c-di-GMP), and sRNAs, regulates biofilm formation by
Salmonella [
86]. The biofilm-forming capacity may vary among serovars [
87] but largely depends on various extrinsic factors, including temperature, source, and contact surface [
88,
89]. In addition, biofilm formation may vary based on their carrier state, i.e., chronic carriage versus sensitive isolates of
Salmonella [
90].
5. Clinical Manifestations
Salmonella causes two forms of the disease in humans: typhoid fever and gastroenteritis. The latter is also known as non-typhoidal salmonellosis [
4]. Human salmonellosis has four clinical manifestations: gastroenteritis, bacteremia, enteric fever, extraintestinal complications, and chronic carrier state [
91].
5.1. Enteric Fever
Salmonella enterica serovar Typhi causes typhoid fever, while
S. Paratyphi A, B, and C cause paratyphoid fever. As the clinical signs of typhoid fever are different from that of paratyphoid fever, the term “enteric fever” (EF) is used to describe both fevers, and both of these agents are called typhoid
Salmonella [
92,
93]. Humans are the only reservoir for both
S. Typhi and
S. Paratyphi, collectively called typhoid salmonellae. Humans acquire the disease via ingesting water or food contaminated with the biowaste of infected individuals. EF onset needs a one or more weeks incubation period, with initial symptoms, such as abdominal pain, headache, and constipation (or diarrhea), accompanied by the onset of fever [
94]. Diarrhea is usually seen in children, whereas constipation is more often observed in immunodeficient patients [
95]. In the course of the illness, enteric fever exhibits two phases, with an initial low-grade (>37.5 °C to 38.2 °C) gradually reaching a high-grade (>38.2 °C to 41.5 °C) fever in the second week. In untreated patients, the fever may linger for a month or more [
96]. Infected individuals may also show bradycardia, myalgia, splenomegaly (enlarged spleen), hepatomegaly (enlarged liver), and rose spots on their abdomen and chest [
97]. Nearly 15% of diseased individuals in an endemic area develop gastrointestinal tract (GIT) complications, including hepatitis, cholecystitis, and pancreatitis. Bleeding is one of the GIT complications that follow the puncture of Peyer’s patches, lymphatic nodules situated at the ileum termini, leading to dysentery. Furthermore, the capability of typhoid salmonellae to persist and survive in the reticuloendothelial system causes a relapse in nearly 10% of the infected individuals [
98].
5.2. Gastroenteritis
Salmonellae serovars other than
S. Paratyphi and
S. Typhi are called NTS and are present in animal reservoirs. Gastroenteritis, an inflammatory condition of the GIT, is a characteristic feature of NTS infection, which is complemented by other clinical signs, such as headache, nausea, vomiting, abdominal pain, non-bloody diarrhea, and myalgia. Splenomegaly and hepatomegaly are less frequently encountered in NTS-infected patients [
99] (
Figure 4). Compared to typhoid infections, NTS has an incubation period of 6–12 h, and the clinical signs are often self-limiting and linger for 10 or a few days [
100]. GI complications of NTS infections include cholecystitis, appendicitis, and pancreatitis without causing terminal ileum perforation [
99]. The severity of the disease and its symptoms would be higher in susceptible individuals, such as immunocompromised patients, aging populations, infants, and young children [
101]. Invasive NTS (iNTS) is prevalent in malnourished and immunocompromised individuals and is widespread in sub-Saharan Africa, resulting in high mortality [
10,
102].
5.3. Bacteremia and Other Extraintestinal Complications
Salmonella bacteremia occurs when
Salmonella invades the intestinal barrier and enters the bloodstream. Almost all serotypes of
Salmonella can cause bacteremia; however,
S. Cholearaesuis and
S. Dublin are the two highly invasive serovars strongly linked with bacteremia [
27,
103]. In contrast to typhoid salmonellae infections, NTS infections are more likely to cause bacteremia. Based on genetic analysis, some serotypes, such as Enteritidis, Typhimurium, and Dublin, but not all serotypes, are suspected of possessing the
Salmonella plasmid virulence gene (
spv), which can cause non-typhoidal bacteremia [
104]. While it is unclear how the presence of the gene increases NTS virulence, expression of the gene prolongs apoptotic cell death, which may permit the pathogen to survive in the host longer [
105]. About 5% of patients who contract NTS develop bacteremia, and the lung is the most impacted organ in some cases. Urinary tract infections (UTIs), cellulitis, pneumonia, meningitis, and endocarditis are the other notable extraintestinal complications [
106,
107].
5.4. Carrier State and Transmission
The
Salmonella chronic carrier state is defined as when bacteria are shed in the stool for at least one year after the onset of the acute infection. Colonization in the gall bladder and secretion through bile is considered a prime source of fecal shedding [
108]. In endemic regions, carriers of
S. Paratyphi and
S. Typhi are responsible for the spread of enteric fever, and the most common means of transmission is the fecal–oral route (drinking of water or ingestion of food contaminated by the feces of chronic carriers) [
109] (
Figure 5). Nearly 4% of enteric fever patients, including infants, the aging population, and women, may serve as chronic carriers [
108]. On the other hand, a carrier state of NTS occurs in only 0.1% of patients, because animals are the primary source of NTS, rather than humans.
Salmonella is a prototype foodborne pathogen whose transmission between animals, plants, and humans has been well-documented [
110].
Salmonella can spread from one farm building and facility to another via pests, including flies, mice, rats, and cockroaches [
111,
112,
113].
Salmonella can be carried in the intestinal tracts of rodents asymptomatically without causing any clinical symptoms and serve as vectors and reservoirs of the disease (
Figure 5). It has also been documented that bacteria are transmitted from cattle to humans through flies, which serve as mechanical vectors [
113]. Wildlife, including wild birds, is a pivotal reservoir for
Salmonella infection [
114]. Studies have shown that human or farm workers’ movement among different farms increases the risk of disease in chickens, hens, and pigs [
115,
116].
6. Epidemiology
Among
Salmonella infections, NTS infections are the most common cause of self-limiting illness. Enteric fever caused by typhoid
Salmonella has a high mortality and morbidity rate and occurs more frequently in developing nations [
117,
118].
6.1. Epidemiology of Enteric Fever
Enteric fever (EF) is endemic in different regions of Asian and African nations and countries in Europe, Central and South America, and the Middle East. EF is rare in the U.S. and some other European countries, with fewer than 10 cases of salmonellosis per 100,000 people each year. Most reported cases in these countries are linked to international travel. Travelers returning from India, Africa, or Pakistan are often the source of this disease [
119,
120,
121]. The rise in cases of
S. Paratyphi infection raises concerns about the efficacy of current vaccines for typhoid fever and suggests the need for a more comprehensive study.
Enteric fever is prevalent in several Asian nations, such as Indonesia, India, Vietnam, China, and Pakistan, with yearly incidence rates surpassing 100 cases per 100,000 people [
122]. Since the data collected by EF are from significant outbreaks, the global incidence of EF reports is more of an estimate. Because of the shortage of diagnostic facilities and effective surveillance technologies in many developing nations, predominantly in sub-Saharan Africa, the prevalence of EF is poorly characterized [
123].
6.2. Epidemiology of Non-Typhoid Salmonella Infections
Despite advances in sanitation and hygiene, the number of NTS infections remains high, posing a problem in developed and developing nations [
2,
124,
125]. Invasive NTS capable of spreading to extraintestinal sites is prevalent in developing countries, particularly in sub-Saharan Africa, with high incidence rates in children under three and HIV-positive individuals [
124,
126]. In Asia, the invasive illness produced by NTS is less common [
127].
Inadequate cooking of foodstuffs, improper storage, and direct contact with raw ingredients are all considered significant causes of
Salmonella outbreaks. Animal commodities, such as milk, poultry, eggs, and other foods, such as peanut butter and chocolate, are frequently linked to epidemics [
128]. Most recently, onion has been implicated in salmonellosis outbreaks in the U.S. [
129].
Animals are considered the primary reservoir of NTS [
130]. Consumption of water or food contaminated with the excrement of infected animals, direct contact with infected animals, or ingestion of infected food animals can cause NTS infection in humans. The global incidence rate of NTS infection is high, as the strains may exist naturally and in wild and domestic animals, such as dogs, cats, amphibians, rodents, and reptiles [
131]. Widespread distribution of food animals, wildlife, and various commodities are primary factors in salmonellae spread in the farm-to-fork food supply continuum.
6.3. Outbreaks of Salmonellosis in Humans
When two or more individuals are afflicted with the same sickness from the same source of contaminated drink or food, such a scenario is known as a foodborne outbreak. Likewise, when two or more individuals suffer from the same disease from animal or animal products and associated environments, the event is classified as a zoonotic outbreak [
132,
133]. A brief overview of outbreaks of salmonellosis in humans on different continents is provided below.
6.3.1. Africa
In Africa, NTS infections appear to be endemic, and are one of the major causes of bacteremia, mostly in children, with 4100 deaths per year [
125]. The prevalence rate is higher in areas where malnutrition, malaria, and HIV are prevalent. Nearly 85.8% of global iNTS deaths have been reported in sub-Saharan Africa [
2]. About 14.3 million typhoid and paratyphoid fever cases in 2017 resulted in 135,900 deaths, 15.8% of which were in sub-Saharan Africa [
134].
Salmonella Typhi is the leading cause of bloodstream infections in eastern and southern Africa, with reports of multiple outbreaks since 2012 [
135]. Malawi has a very high incidence of 444 cases per 100,000 persons per year [
92]. The primary infection source of people’s exposure to
S. Typhi is uncertain [
136]. In Africa, iNTS is mainly associated with HIV patients (both adults and young children), malaria infection, and malnutrition [
137]. Two
Salmonella serotypes, Enteritidis and Typhimurium, have been reported to be the most common causes of iNTS in South Africa, Malawi, Mozambique, Kenya, and Mali, with
S. Typhimurium Sequence Type (ST) 313 (ST313) and S. Enteritidis ST11 being the most frequently reported serovars [
138]. In South Africa (2020 and 2021), although the total number of enteric fever cases across the country was similar to previous years (83 patients in 2020 and 110 patients in 2021), there was a relative upsurge in the number of cases reported from the northwest provinces and Western Cape [
139]. In Nigeria, out of 372 humans screened, 77 (20.7%) were positive for enteric fever, 38 (20.4%) were isolated from non-poultry workers, and 39 (21.0%) were isolated from poultry staff in the three senatorial districts [
140]. A recent study on 16,236 children from Kenya indicated that 1.3% of bloodstream infections was caused by
Salmonella Typhimurium and Enteritidis, while
Salmonella Typhi caused 1.4% of disease.
Salmonella Enteritidis and
S. Typhimurium were not significantly associated with rearing domestic animals. However, rearing chicken was linked to a high prevalence of
S. Typhi (2.1%) infection. The rate of children infected with
Salmonella Typhimurium and Enteritidis was significantly higher in households that used water pots as water storage containers compared to using water directly from the tap (0.6%) [
141].
An extensive drug-resistant (XDR) strain of
Salmonella Typhimurium was reported to cause millions of bloodstream infections per year in sub-Saharan Africa, including in the Democratic Republic of Congo (DRC) [
142]. A recent study conducted in Burkina Faso indicated that among the 106
Salmonella isolates (77 human stools; 14 sandwiches), O antigen-positive
Salmonella was confirmed in 86% (91/106) of the samples, and serogroup O:4,5 was the most common serogroup detected (40%; 36/91).
Salmonella Enteritidis and Typhimurium represented 5.5% (5/91) and 3.3% (3/91), respectively, and were identified only from clinical isolates. Furthermore, 14 serotypes of
Salmonella (12/91 human strains and 2/15 sandwich strains) were evocative of Kentucky and Bargny serotypes [
143]. In Ethiopia (from 2010 to 2020), the pooled prevalence of
Salmonella among human stools and animal-origin foods was 4.8% and 7.7%, respectively [
144].
6.3.2. Middle East and North Africa
Several reports indicate a worrisome rising trend of NTS cases in developing countries, including the Middle East and northern Africa (MENA) [
145,
146]. A systematic review and meta-analysis study conducted on the prevalence of enteric NTS in humans in the MENA countries indicated that there were 6356
Salmonella-positive cases associated with 252,831 human samples. The pooled
Salmonella prevalence in MENA was estimated at 6.6%. The highest pooled prevalence of
Salmonella reports were in Tunisia (10.2%), Morocco (17.9%), and Sudan (9.2%), while the lowest were in Oman (1.2%), Jordan (1.1%), and Palestine (1.2%) [
147].
A recent study in Iran indicated that nearly 94% of
Salmonella isolates were recovered from ≤5-year-old patients, and 99% were NTS. The author found extensive diversity among
Salmonella isolates; serogroup D (46%) was predominant, and
Salmonella Enteritidis (41%) was the most common serotype that showed the highest antimicrobial susceptibility rate (>96%).
S. Newport from human specimens was isolated for the first time in Iran. Most isolates were sensitive to all antimicrobials tested, but 35% of isolates were not-typed (NT), which showed the highest resistance, with 48% being resistant to ≥1 antimicrobial tested [
148].
Malaeb et al. [
149] reviewed published data from Lebanon on
Salmonella susceptibility/resistance patterns and its clinical complications. The estimated incidence was 13.34 cases per 100,000 individuals, and most cases occurred in the 20–39 age group with no significant gender variation. Poor and less developed districts of Lebanon had the highest number of cases, and the peak incidence was in summer [
149].
A case-control study conducted in central Israel indicated that in 18 years (2001–2018), 34 cases of NTS were identified in the bloodstreams of infected patients. The median age was 59 years, with 20% of patients below 20 years of age [
150].
Salmonella infection in Saudi Arabia is highly prevalent during the Hajj and Umrah seasons due to the gathering of many pilgrims [
151]. A retrospective descriptive study conducted in King Khalid University Hospital (KKUH), Riyadh, Saudi Arabia, between May 2017 and December 2018, indicated 22 patients with invasive
Salmonella infection. Fifteen (68%) were females, and seven (32%) were males. The range of ages was from 8 months to 74 years [
152].
6.3.3. Latin America
Typhoid is broadly accepted to be endemic in parts of Latin America; the region has a medium incidence of typhoid fever (53 per 100,000 people), corresponding to >273,000 cases annually [
153]. Using cases reported to the National Public Health Surveillance System in Columbia between 2012 and 2015, typhoid salmonellae was found in 836 patients, with the majority (676/836; 80.1%) of reported cases originating from only 7 departments. They further characterized 402
S. Typhi isolates with available corresponding data recovered from various departments of Colombia through antimicrobial susceptibility testing and molecular subtyping. The majority (235/402; 58.5%) of these typhoid cases occurred in males aged between 10 and 29 years (218/402; 54.2%), with 3 deaths (0.74%). The overwhelming preponderance (339/402; 84.3%) of
S. Typhi were susceptible to all tested antimicrobials. The organism showed the most resistance against ampicillin (30/402;7.5%), followed by nalidixic acid (23/402, 5.7%) [
154].
In Brazil, serotyping of 3113
Salmonella isolates collected by the National Reference Laboratory for Enteric Diseases between 2011 and 2020 revealed 61 serogroups [
155]. Calarga et al. [
156] studied the prevalence of the antimicrobial-resistant phenotype in 789 NTS strains collected between 2000–2019 in São Paulo, Brazil. Among the non-susceptible isolates, 31.55, 14.06, and 13.18% were resistant to aminoglycosides, tetracycline, and β-lactams, respectively. Moreover, 68 and 11 isolates were MDR and extended-spectrum β-lactamase (ESBL) producers, respectively, whereas one isolate was colistin-resistant [
156].
6.3.4. USA
The Centers for Disease Control and Prevention (CDC), USA, estimates that approximately 1.35 million illnesses, 26,500 hospitalizations, and 420 deaths occur due to NTS infection each year in the U.S., resulting in an estimated
$400 million in direct medical costs [
157]. Between 2009 and 2011, antibiotic-resistant
Salmonella strains that had developed resistance to 5 or more antibiotics caused over 66,000 illnesses in the U.S. [
158]. According to CDC, antibiotic-resistant NTS infections are on the rise, approaching an estimated 10% for ciprofloxacin, 3% for ceftriaxone, and 1% for azithromycin [
157]. Prolonged hospitalization and increased risk of bloodstream infections, treatment failure, and excess mortality have been associated with antimicrobial drug-resistant NTS infections [
102].
In late 2022, a multi-country outbreak of
Salmonella Typhimurium was reported in the USA and UK. The outbreak was associated with chocolate produced in Belgium and was distributed globally to over 113 countries and territories across all WHO regions. While 150 of 151 known cases have been reported in Europe, 1 case has been reported in the U.S. Additional cases are likely reported from other countries, given the broad distribution of the products during the Easter holiday [
159].
6.3.5. Europe
Salmonellosis remains the second most common zoonotic disease in humans in the European Union (EU). The incidence of human salmonellosis has decreased steadily in recent years. Nevertheless, in 2014, 88,175 confirmed human salmonellosis cases, causing 9830 hospitalizations and 65 fatalities, were reported across the EU. Among these, 16,000 cases of human salmonellosis were reported in Germany. As in previous years,
S. Enteritidis was the predominant serovar (44.4% of all isolates), followed by
S. Typhimurium (17.4%) and a monophasic
S. Typhimurium variant (7.8%) [
160].
After a considerable decrease in salmonellosis cases recorded from 2007 to 2014, the incidence was stable between 2015 and 2019. The number of cases in 2020 was significantly lower than in previous years, mainly due to the COVID-19 pandemic. All but two countries reported a decrease in the number of patients due to various factors, including people avoiding hospital and/or clinic visits for mild sickness for fear of the risk of exposure to COVID-19 in healthcare facilities, lower laboratory services because of the reallocation of resources to SARS-CoV-2, limited restaurant visits, frequent hand washing practices, and limited human movement and personal contacts due to travel restrictions [
133].
Notification rates for human salmonellosis also differ between member states in the EU, including area coverage, quality of data, disease severity, surveillance systems, sampling and testing, the prevalence in the food-producing animal population, food and animal trade between member states, and the proportion of travel-associated cases [
161].
In 2020, the majority (58%) of foodborne outbreaks were caused by
S. Enteritidis, similar to previous years. The four most commonly encountered food vehicles in confirmed foodborne outbreaks associated with salmonellosis include ‘eggs and egg products’, ‘pig meat and products thereof’ and ‘bakery products”, as in previous years. Nearly 29 countries reported 53,674 cases, of which 53,169 were classified as confirmed. The number of notifications per 100,000 population was 14.2, considerably fewer than in 2019. Age-standardized notification rates did not differ substantially from crude rates. Of 35,715 cases with known outcomes, 61 were reported to have died, accounting for a case fatality of 0.17%. The highest prevalence was reported by the Czech Republic (98.4 cases per 100,000 population) and Slovakia (62.1), followed by Malta (34.2) and Hungary (30.3) [
133]. Some of the recent outbreaks of human salmonellosis reported from different geographic regions associated with various foodstuffs are summarized in
Table 1.
7. Antimicrobial Resistance
Antimicrobial agents have been extensively used and misused for treating infectious diseases in animals and humans and as promoters of growth in livestock production [
168,
169]. Indiscriminate antibiotic use has led to the evolution of antimicrobial resistance (AMR) against antibiotics available to combat bacterial infections. AMR bacteria have emerged along the food chain, posing critical concerns to public health. Many studies have indicated the colonization, disease, and contamination of food animals and their products by one or more of these bacteria [
169]. These include methicillin-resistant
Staphylococcus aureus (MRSA),
Campylobacter spp., and extended-spectrum beta-lactamase
Enterobacteriaceae, such as
Salmonella,
E. coli, and
Shigella [
170].
Antimicrobial resistance in
Salmonella has become an essential concern to public health worldwide [
171]. The first case of
Salmonella resistance to a single antibiotic, chloramphenicol, was reported in the early 1960s. Since then, nations, including the United Kingdom, United States, and Saudi Arabia, have seen an upsurge of antibiotic-resistant
Salmonella isolates resistant to one or more antimicrobials [
172]. Conventional first-line therapies for
Salmonella infections include chloramphenicol, ampicillin, and trimethoprim-sulfamethoxazole [
173]. Multi-drug resistant (MDR) salmonellae exhibit resistance to various drugs. A recent increase in MDR
Salmonella, and resistance to clinically significant antimicrobials, such as third-generation cephalosporins and fluoroquinolones, pose substantial concerns.
The European Union report on antimicrobial resistance in human indicators and zoonotic bacteria from 2019 to 2020 indicates an increased resistance of salmonellae to sulfonamides, ampicillin, and tetracyclines at high levels (50–80%) [
174]. However, the resistance to third-generation cephalosporins, namely ceftazidime and cefotaxime, was observed at very low levels of 0.5% and 0.8%, respectively. Higher MDR was noted in Italy and Belgium, with a 42.9% and 35% prevalence, respectively, from 2016 to 2020 (
Table 2) [
174].
7.1. Mechanisms of Antibiotic Resistance
A variety of hybrid plasmids (plasmids that contain an inserted piece of foreign DNA) are produced by
Salmonella strains with the MDR phenotype. Plasmids carrying various resistance genes can confer resistance to conventional antibiotics, such as streptomycin, chloramphenicol, tetracycline, and ampicillin [
175,
176,
177]. Several serotypes are also resistant to ciprofloxacin due to the mutations in the chromosomal-linked gene,
gyrA, which also contributes to quinolone resistance [
178]. Resistance to broad-spectrum cephalosporins is attributed to altered genes encoding extended-spectrum β-lactamases, which cleave β-lactam rings of cephamycins and cephalosporins [
179,
180].
Salmonellae use a variety of resistance mechanisms to combat antimicrobials, and drug inactivation is the most common (
Figure 6) [
177,
181,
182,
183]. In this pathway, the functional unit of the antibiotic is inactivated or destroyed via chemical modification with the help of enzymes that hydrolyze phosphorylation, acetylation, and adenylation reactions [
182]. Moreover, enzymes, such as chloramphenicol acetyltransferase and penicillinase, can acetylate the two hydroxyl groups of chloramphenicol, and ß-lactam rings of penicillin and cephalosporin, respectively [
182].
Hindering the target site (cellular structure or enzymes) of antibiotics is the other mechanism of resistance that
Salmonella serotypes can use. For example, quinolone resistance protein (Qnr), a plasmid-encoded protein, confers quinolone resistance by competing with topoisomerase IV and DNA gyrase for binding sites [
182]. As a result, the antibiotic is less likely to bind to DNA gyrase, defending the bacteria from lethal effects [
184]. Moreover,
Salmonella can alter the receptor of the antibiotic in a way that prevents it from binding to it. Rifampicin resistance, for example, is caused by a single-step mutation that results in substitutions of amino acids in the
rpoB gene.
Consequently, the affinity of the antibiotics for DNA polymerase could be lessened, permitting
rpoB transcription to proceed [
182]. It is also known that
Salmonella resists drugs by reducing its membrane permeability, preventing them from entering the cell [
185]. When the membrane protein composition changes, the pores of the membrane transport system are altered, and, hence, antibiotics cannot pass through. Modifying the lipid A moiety of a lipopolysaccharide structure lowers the net negative charge of a membrane, decreasing its attraction for polymyxin and resistance to this antibiotic [
186].
Salmonella can also pump out a drug before it reaches the target site, using efflux pumps or multidrug resistance pumps. Many antibiotics, such as ß-lactams, fluoroquinolones, and carbapenems, can be pumped out using this mechanism [
182]. The genes encoding these efflux pumps (e.g.,
qepA and
oqxAB) are positioned in mobile elements, such as plasmids [
187].
Salmonella can also exert resistance by using chromosomal-linked gene products through mutations that code for the drug’s target or the mechanisms that control the internalization of the drugs [
185]. Single point mutations in chromosomal
parC and
parE, topoisomerase genes responsible for quinolone resistance, and the DNA gyrase genes
gyrA and
gyrB have been reported in some
Salmonella serotypes. The resulting alterations cause the bacterium to be resistant to fluoroquinolones and quinolones [
188].
7.2. Spread of Antimicrobial Resistance
The indiscriminate use of antimicrobial agents other than therapeutic agents, such as the case of growth promoters in food animals and the sharing of human drugs for treating animal infections or other veterinary use, is considered the leading cause of the emergence of resistance of microbes to the first-line as well as the last resort antimicrobial agents [
158,
173]. MDR
Salmonella strains have been transmitted from animals to humans by consuming water or food contaminated with the animals’ feces, eating contaminated food from animal sources, or having direct contact with infected subjects [
189]. Furthermore, MDR
Salmonella strains have been discovered in several exotic pet animals, such as turtles and tortoises, and in their aquatic habitat, potentially exposing individuals to contagious zoonotic infections [
190].
7.3. Clinical Importance
The emergence of MDR
Salmonella serotypes which have developed resistance to multiple antimicrobials substantially influences the treatment of human salmonellosis with antibiotics. Invasive serotype infections are typically fatal and require prompt antibiotic therapy. Third-generation cephalosporins and quinolones have long been the treatment of choice for MDR
Salmonella infections [
191]. The
Salmonella serotypes showing resistance to quinolones and cephalosporins present a new problem in treating infected individuals. The lack of successful antibiotic therapy may result in higher morbidity and fatality [
192]. As a result of the introduction of MDR
Salmonella, the severity of bacterial infections in people and animals has risen. According to epidemiological research, MDR
Salmonella strains generate more severe or fatal infections than non-MDR strains [
193,
194].
8. Prevention of Salmonellosis
Enteric fever spreads chiefly through the fecal–oral route. Historically, enteric fever was prevalent in the U.S. and Western Europe; however, typhoid-causing
Salmonella infection has dramatically reduced with appropriate water and food sanitation, hygienic practices, pasteurization of dairy products, and the prohibition of using human feces as a fertilizer. In Latin America, the occurrence of
Salmonella infections has been reduced with improved sanitary measures. Availability of safe food and drinking water, good sanitation, and the utilization of typhoid vaccines are currently the mainstays of enteric fever prevention [
195].
The primary objective for eliminating the potential transmission pathways of typhoid
Salmonella and NTS is to ensure the safety of drinking water. This critical step has been implemented effectively in developed nations, such as the U.S. and in Europe, but not in underdeveloped and developing continents [
196]. Apart from water,
Salmonella spp. may be found in several foods, including chicken, eggs, meats, nuts, flour, vegetables, and dairy products. Proper food handling and preparation are recommended to eliminate bacterial contamination of food. Food irradiation has been widely advocated in many nations because of its efficiency in lowering the danger of food contamination. Food irradiation technology has been accepted by various public health bodies, including the WHO and the CDC [
197,
198].
Vaccination is one of the crucial strategies to prevent enteric fever infections [
199,
200]. The two vaccinations now licensed to avoid EF are inactive oral and parenteral live attenuated vaccines [
201]. These approved vaccines, however, are only for newborns, and are ineffective in preventing NTS and
S. Paratyphi infections [
202]. One promising and efficient solution to manage NTS is to limit antimicrobials, especially human-use antibiotics, in the feed of food animals and live animals [
173,
194,
203]. In addition, plant-derived antimicrobials, probiotics, or direct-fed-microbes (DFM) are applied to control
Salmonella in meat-producing animals and poultry [
204,
205,
206,
207,
208].
9. Future Perspective and Conclusions
Human salmonellosis is still a significant public health threat across the world and an important cause of foodborne outbreaks all over the globe. This review emphasized providing data on recent episodes of salmonellosis reported in different clinical settings and geographical regions, including Africa, the Middle East and North Africa, Europe, and the USA in the farm-to-fork continuum. Salmonella strains’ genetic makeup allows them to withstand various situations, including animal, human, and non-animal hosts. The organism possesses a wide range of virulence factors involved in the multiple stages of infection. Their cunning survival strategy makes it challenging to eliminate this bacterium from the farm-to-fork food supply chain continuum and makes the infection cycles more difficult to control.
Furthermore, the emergence of MDR Salmonella strains has led to a challenging situation in treating the disease caused by these strains. As we discussed, several Salmonella outbreaks have been reported in different continents associated with various foodstuffs. However, the mortality and morbidity rate of the disease varied, reflecting differences in prevalence in food and animals used for food production, animal trade between countries, the proportion of travel-associated cases, and the quality and coverage of surveillance systems. Several preventative strategies have been recommended to reduce the spread of Salmonella infection, including restricting indiscriminate antibiotic use in food animals, which is by far the most successful. In addition, plant-based antimicrobials, probiotics, and bacteriophages are considered viable alternatives as feed additives. For the prevention of enteric fever, two vaccines have been approved; however, there are no certified vaccinations for S. Paratyphi and NTS infections. More studies into the preparation of vaccines for all Salmonella strains are needed.
The role of ecology in transmitting Salmonella from a One Health (a multidisciplinary and cross-sectoral cooperation) perspective is a crucial approach to managing the disease in the farm-to-fork continuum. Hence, emphasis should be given to the safe and efficient control of insects and rodents, which are the main vectors of contamination and cross-species transmission. In addition, in a free-range environment, safe and efficient barriers are required to prevent the transmission of pathogens among cattle, chickens, and pigs through contaminated feed (e.g., pasture) or the environment.
The mechanisms of Salmonella persistence outside the host are an essential part of their lifecycle and a prerequisite for their evolutionary success as human pathogens. Therefore, detailed investigations are needed to examine the transmission of diseases mainly associated with low-moisture foods, such as peanut butter, spray-dried milk, dry cereal, infant formula, nuts, etc. Furthermore, from a public health point of view, thorough investigations are needed to understand the bacterial factors or other related factors involved in the persistence of the pathogen in chronic infections and the vertical transmission of the agent.
Increasing awareness and intensive implementation of food safety pillars could effectively combat foodborne diseases, including human salmonellosis. All stakeholders, including consumers, farmers, food vendors, and others involved in the food system, must be formally and informally educated on the basic steps of food safety. Food safety training can be conducted in the workplace, school, or community space, via video talks/demonstrations, on any social media platforms (Facebook, Twitter, Instagram, etc.), talk shows, radio interviews, or media briefings. Furthermore, community activities can be initiated by organizing a walk, run, bicycle ride, or fitness classes to publicize food safety as a key to sustaining life and promoting good health.
Author Contributions
Conceptualization, A.D.T., R.R.A.-H. and A.K.B.; contribution to writing A.D.T., R.R.A.-H., A.K.B., R.S.A., M.G.A., M.A.A., A.A.R.F., A.S.A.M., A.M.A.-A., M.S.A., A.M.A., A.H.A. and M.A.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by [Institutional Fund Projects] grant number [IFPRC-109-130-2020].
Data Availability Statement
Not applicable.
Acknowledgments
This research work was funded by Institutional Fund Projects under grant no (IFPRC-109-130-2020). Therefore, the authors gratefully acknowledge technical and financial support from the Ministry of Education, and King Abdulaziz University, Jeddah, Saudi Arabia. AKB acknowledges the funding support of the USDA National Institute of Food and Agriculture (Hatch accession no. 1016249).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jain, P.; Chowdhury, G.; Samajpati, S.; Basak, S.; Ganai, A.; Samanta, S.; Okamoto, K.; Mukhopadhyay, A.K.; Dutta, S. Characterization of non-typhoidal Salmonella isolates from children with acute gastroenteritis, Kolkata, India, during 2000–2016. Brazilian J. Microbiol. 2020, 51, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Stanaway, J.D.; Parisi, A.; Sarkar, K.; Blacker, B.F.; Reiner, R.C.; Hay, S.I.; Nixon, M.R.; Dolecek, C.; James, S.L.; Mokdad, A.H. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef] [PubMed]
- Popa, G.L.; Papa, M.I. Salmonella spp. infection-A continuous threat worldwide. Germs 2021, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.K. Salmonella enterica. In Foodborne Microbial Pathogens: Mechanisms and Pathogenesis; Bhunia, A.K., Ed.; Springer: New York, NY, USA, 2018; pp. 271–287. [Google Scholar] [CrossRef]
- Gut, A.M.; Vasiljevic, T.; Yeager, T.; Donkor, O.N. Salmonella infection–prevention and treatment by antibiotics and probiotic yeasts: A review. Microbiology 2018, 164, 1327–1344. [Google Scholar] [CrossRef]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Doepfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef]
- ECDC. Salmonella the Most Common Cause of Foodborne Outbreaks in the European Union. European Centre for Disease Prevention and Control. 2019. Available online: https://www.ecdc.europa.eu/en/news-events/salmonella-most-common-cause-foodborne-outbreaks-european-union (accessed on 20 October 2022).
- Somorin, Y.M.; Odeyemi, O.A.; Ateba, C.N. Salmonella is the most common foodborne pathogen in African food exports to the European Union: Analysis of the Rapid Alert System for Food and Feed (1999–2019). Food Control 2021, 123, 107849. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Vieira, A.R.; Karlsmose, S.; Lo Fo Wong, D.M.; Jensen, A.B.; Wegener, H.C.; Aarestrup, F.M. Global monitoring of Salmonella serovar distribution from the World Health Organization Global Foodborne Infections Network Country Data Bank: Results of quality assured laboratories from 2001 to 2007. Foodborne Pathog. Dis. 2011, 8, 887–900. [Google Scholar] [CrossRef]
- Uche, I.V.; MacLennan, C.A.; Saul, A. A systematic review of the incidence, risk factors and case fatality rates of invasive nontyphoidal Salmonella (iNTS) disease in Africa (1966 to 2014). PLoS Negl. Trop. Dis. 2017, 11, e0005118. [Google Scholar] [CrossRef]
- Hur, J.; Jawale, C.; Lee, J.H. Antimicrobial resistance of Salmonella isolated from food animals: A review. Food Res. Int. 2012, 45, 819–830. [Google Scholar] [CrossRef]
- Singh, A.K.; Drolia, R.; Bai, X.; Bhunia, A.K. Streptomycin induced stress response in Salmonella enterica serovar Typhimurium shows distinct colony scatter signature. PLoS ONE 2015, 10, e0135035. [Google Scholar] [CrossRef]
- WHO. Stop Using Antibiotics in Healthy Animals to Prevent the Spread of Antibiotic Resistance. 2017. Available online: http://www.who.int/mediacentre/news/releases/2017/antibiotics-animalseffectiveness/en (accessed on 4 November 2022).
- Magwedere, K.; Rauff, D.; De Klerk, G.; Keddy, K.H.; Dziva, F. Incidence of Nontyphoidal Salmonella in Food-Producing Animals, Animal Feed, and the Associated Environment in South Africa, 2012–2014. Clin. Infect. Dis. 2015, 61 (Suppl. S4), S283–S289. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food commodities, United States, 1998–2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, C.A.; Ricke, S.C.; Marcy, J.A. Public health impact of Salmonella spp. on raw poultry: Current concepts and future prospects in the United States. Food Control 2022, 132, 108539. [Google Scholar] [CrossRef]
- Ramatla, T.; Tawana, M.; Onyiche, T.E.; Lekota, K.E.; Thekisoe, O. Prevalence of antibiotic resistance in Salmonella serotypes concurrently isolated from the environment, animals, and humans in South Africa: A systematic review and meta-analysis. Antibiotics 2021, 10, 1435. [Google Scholar] [CrossRef]
- Di Marcantonio, L.; Romantini, R.; Marotta, F.; Chiaverini, A.; Zilli, K.; Abass, A.; Di Giannatale, E.; Garofolo, G.; Janowicz, A. The Current Landscape of Antibiotic Resistance of Salmonella Infantis in Italy: The Expansion of Extended-Spectrum Beta-Lactamase Producers on a Local Scale. Front. Microbiol. 2022, 13, 812481. [Google Scholar] [CrossRef]
- Reeves, M.W.; Evins, G.M.; Heiba, A.A.; Plikaytis, B.D.; Farmer, J.J., 3rd. Clonal nature of Salmonella typhi and its genetic relatedness to other salmonellae as shown by multilocus enzyme electrophoresis, and proposal of Salmonella bongori comb. nov. J. Clin. Microbiol. 1989, 27, 313–320. [Google Scholar] [CrossRef]
- Lan, R.; Reeves, P.R.; Octavia, S. Population structure, origins and evolution of major Salmonella enterica clones. Infect. Gen. Evol. 2009, 9, 996–1005. [Google Scholar] [CrossRef]
- Guard, J. Through the Looking Glass: Genome, Phenome, and Interactome of Salmonella enterica. Pathogens 2022, 11, 581. [Google Scholar] [CrossRef]
- Brenner, F.W.; Villar, R.G.; Angulo, F.J.; Tauxe, R.; Swaminathan, B. Salmonella Nomenclature. J. Clin. Microbiol. 2000, 38, 2465. [Google Scholar] [CrossRef]
- Hu, L.; Kopecko, D.J. Typhoid Salmonella; Marcel Dekker, Inc.: New York, NY, USA, 2003; pp. 151–165. [Google Scholar]
- McQuiston, J.R.; Waters, R.J.; Dinsmore, B.A.; Mikoleit, M.L.; Fields, P.I. Molecular determination of H antigens of Salmonella by use of amicrosphere-based liquid array. J. Clin. Microbiol. 2011, 49, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Parween, F.; Yadav, J.; Qadri, A. The virulence polysaccharide of Salmonella Typhi suppresses activation of Rho family GTPases to limit inflammatory responses from epithelial cells. Front. Cell. Infect. Microbiol. 2019, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Wattiau, P.; Boland, C.; Bertrand, S. Methodologies for Salmonella enterica subsp. enterica subtyping: Gold standards and alternatives. Appl. Environ. Microbiol. 2011, 77, 7877–7885. [Google Scholar] [CrossRef]
- Guibourdenche, M.; Roggentin, P.; Mikoleit, M.; Fields, P.I.; Bockemühl, J.; Grimont, P.A.D.; Weill, F.-X. Supplement 2003–2007 (No. 47) to the white-Kauffmann-Le minor scheme. Res. Microbiol. 2010, 161, 26–29. [Google Scholar] [CrossRef]
- Tindall, B.J.; Grimont, P.A.D.; Garrity, G.M.; Euzeby, J.P. Nomenclature and taxonomy of the genus Salmonella. Int J Syst Evol Microbiol 2005, 55, 521. [Google Scholar] [CrossRef]
- Chattaway, M.A.; Langridge, G.C.; Wain, J. Salmonella nomenclature in the genomic era: A time for change. Sci. Rep. 2021, 11, 7494. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Stecher, B.; Hensel, M. Type III secretion of Salmonella enterica serovar Typhimurium translocated effectors and SseFG. Infect. Immun. 2002, 70, 1403–1409. [Google Scholar] [CrossRef]
- Grassl, G.A.; Finlay, B.B. Pathogenesis of enteric Salmonella infections. Curr. Opin. Gastroenterol. 2008, 24, 22–26. [Google Scholar] [CrossRef]
- Dillon, A.; Lo, D.D. M cells: Intelligent engineering of mucosal immune surveillance. Front. Immunol. 2019, 10, 1499. [Google Scholar] [CrossRef]
- Roche, S.M.; Holbert, S.; Trotereau, J.; Schaeffer, S.; Georgeault, S.; Virlogeux-Payant, I.; Velge, P. Salmonella Typhimurium invalidated for the three currently known invasion factors keeps its ability to invade several cell models. Front. Cell. Infect. Microbiol. 2018, 8, 273. [Google Scholar] [CrossRef]
- Dos Santos, A.M.P.; Ferrari, R.G.; Conte-Junior, C.A. Type three secretion system in Salmonella Typhimurium: The key to infection. Genes Genom. 2020, 42, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Takaya, A.; Tomoyasu, T.; Matsui, H.; Yamamoto, T. The DnaK/DnaJ chaperone machinery of Salmonella enterica serovar Typhimurium is essential for invasion of epithelial cells and survival within macrophages, leading to systemic infection. Infect. Immun. 2004, 72, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Choi, E.; Cho, Y.-J.; Nam, D.; Lee, J.; Lee, E.-J. The Salmonella virulence protein MgtC promotes phosphate uptake inside macrophages. Nat. Commun. 2019, 10, 3326. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, D.; Negi, V.D. Repeated in-vitro and in-vivo exposure leads to genetic alteration, adaptations, and hypervirulence in Salmonella. Microb. Pathog. 2019, 136, 103654. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, J.A.; Crooks, A.L.; Nagy, T.A.; Quintana, J.L.J.; Dalebroux, Z.D.; Detweiler, C.S. Salmonella enterica Infections Are Disrupted by Two Small Molecules That Accumulate within Phagosomes and Differentially Damage Bacterial Inner Membranes. mBio 2022, 13, e01790-22. [Google Scholar] [CrossRef]
- Klumpp, J.; Fuchs, T.M. Identification of novel genes in genomic islands that contribute to Salmonella Typhimurium replication in macrophages. Microbiology 2007, 153, 1207–1220. [Google Scholar] [CrossRef]
- Ibarra, J.A.; Steele-Mortimer, O. Salmonella–the ultimate insider. Salmonella virulence factors that modulate intracellular survival. Cell. Microbiol. 2009, 11, 1579–1586. [Google Scholar]
- Fàbrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef]
- Tomljenovic-Berube, A.M.; Henriksbo, B.; Porwollik, S.; Cooper, C.A.; Tuinema, B.R.; McClelland, M.; Coombes, B.K. Mapping and Regulation of Genes within Salmonella Pathogenicity Island 12 That Contribute to In Vivo Fitness of Salmonella enterica Serovar Typhimurium. Infect. Immun. 2013, 81, 2394–2404. [Google Scholar] [CrossRef]
- Fattinger, S.A.; Sellin, M.E.; Hardt, W.-D. Salmonella effector driven invasion of the gut epithelium: Breaking in and setting the house on fire. Curr. Opin. Microbiol. 2021, 64, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Fookes, M.; Schroeder, G.N.; Langridge, G.C.; Blondel, C.J.; Mammina, C.; Connor, T.R.; Seth-Smith, H.; Vernikos, G.S.; Robinson, K.S.; Sanders, M. Salmonella bongori provides insights into the evolution of the Salmonellae. PLoS Pathog. 2011, 7, e1002191. [Google Scholar] [CrossRef] [PubMed]
- Faucher, S.P.; Curtiss Iii, R.; Daigle, F. Selective capture of Salmonella enterica serovar Typhi genes expressed in macrophages that are absent from the Salmonella enterica serovar Typhimurium genome. Infect. Immun. 2005, 73, 5217–5221. [Google Scholar] [CrossRef]
- Morris, C.; Tam, C.K.P.; Wallis, T.S.; Jones, P.W.; Hackett, J. Salmonella enterica serovar Dublin strains which are Vi antigen-positive use type IVB pili for bacterial self-association and human intestinal cell entry. Microb. Pathog. 2003, 35, 279–284. [Google Scholar] [CrossRef]
- Pickard, D.; Wain, J.; Baker, S.; Line, A.; Chohan, S.; Fookes, M.; Barron, A.; Ó Gaora, P.; Chabalgoity, J.A.; Thanky, N. Composition, acquisition, and distribution of the Vi exopolysaccharide-encoding Salmonella enterica pathogenicity island SPI-7. J. Biotechnol. 2003, 185, 5055–5065. [Google Scholar] [CrossRef] [PubMed]
- Den Bakker, H.C.; Moreno Switt, A.I.; Govoni, G.; Cummings, C.A.; Ranieri, M.L.; Degoricija, L.; Hoelzer, K.; Rodriguez-Rivera, L.D.; Brown, S.; Bolchacova, E. Genome sequencing reveals diversification of virulence factor content and possible host adaptation in distinct subpopulations of Salmonella enterica. BMC Genom. 2011, 12, 425. [Google Scholar] [CrossRef]
- Rodriguez-Rivera, L.D.; Bowen, B.M.; den Bakker, H.C.; Duhamel, G.E.; Wiedmann, M. Characterization of the cytolethal distending toxin (typhoid toxin) in non-typhoidal Salmonella serovars. Gut Pathog. 2015, 7, 19. [Google Scholar] [CrossRef]
- Pezoa, D.; Andrews-Polymenis, H.; Blondel, C.J.; Leiva, L.; Pinto, B.; Alvarez, S.A.; Santiviago, C.A.; Contreras, L.I. The type VI secretion systems encoded in Salmonella pathogenicity islands SPI-6 and SPI-19 contribute to intestinal colonization of chickens and systemic spread by Salmonella enterica serotype Typhimurium. FEBS J. 2012, 279, 227–228. [Google Scholar]
- Galán, J.E. Typhoid toxin provides a window into typhoid fever and the biology of Salmonella Typhi. Proc Nat Acad Sci USA 2016, 113, 6338–6344. [Google Scholar] [CrossRef]
- Thakur, R.; Suri, C.R.; Rishi, P. Contribution of typhoid toxin in the pathogenesis of Salmonella Typhi. Microb. Pathog. 2022, 164, 105444. [Google Scholar] [CrossRef]
- Yang, Y.-A.; Lee, S.; Zhao, J.; Thompson, A.J.; McBride, R.; Tsogtbaatar, B.; Paulson, J.C.; Nussinov, R.; Deng, L.; Song, J. In vivo tropism of Salmonella Typhi toxin to cells expressing a multiantennal glycan receptor. Nat. Microbiol. 2018, 3, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Tamamura, Y.; Tanaka, K.; Uchida, I. Characterization of pertussis-like toxin from Salmonella spp. that catalyzes ADP-ribosylation of G proteins. Sci. Rep. 2017, 7, 2653. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.A.; Eade, C.R.; Wiedmann, M. Embracing diversity: Differences in virulence mechanisms, disease severity, and host adaptations contribute to the success of nontyphoidal Salmonella as a foodborne pathogen. Front. Microbiol. 2019, 10, 1368. [Google Scholar] [CrossRef] [PubMed]
- Horstmann, J.A.; Lunelli, M.; Cazzola, H.; Heidemann, J.; Kühne, C.; Steffen, P.; Szefs, S.; Rossi, C.; Lokareddy, R.K.; Wang, C. Methylation of Salmonella Typhimurium flagella promotes bacterial adhesion and host cell invasion. Nat. Commun. 2020, 11, 2013. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella pathogenicity and host adaptation in chicken-associated serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef]
- De Maayer, P.; Pillay, T.; Coutinho, T.A. Flagella by numbers: Comparative genomic analysis of the supernumerary flagellar systems among the Enterobacterales. BMC Genom. 2020, 21, 670. [Google Scholar] [CrossRef]
- Nedeljković, M.; Sastre, D.E.; Sundberg, E.J. Bacterial flagellar filament: A supramolecular multifunctional nanostructure. Int. J. Mol. Sci. 2021, 22, 7521. [Google Scholar] [CrossRef]
- García-Pastor, L.; Puerta-Fernández, E.; Casadesús, J. Bistability and phase variation in Salmonella enterica. Biochim Biophys Acta (BBA)-Gene Reg. Mech. 2019, 1862, 752–758. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Toma, S.; Terahara, N.; Miyata, T.; Ashihara, M.; Minamino, T.; Namba, K.; Kato, T. Structural and Functional Comparison of Salmonella Flagellar Filaments Composed of FljB and FliC. Biomolecules 2020, 10, 246. [Google Scholar] [CrossRef]
- Dos Santos, A.M.P.; Ferrari, R.G.; Conte-Junior, C.A. Virulence factors in Salmonella Typhimurium: The sagacity of a bacterium. Curr. Microbiol. 2019, 76, 762–773. [Google Scholar] [CrossRef]
- Rivera-Chávez, F.; Bäumler, A.J. The pyromaniac inside you: Salmonella metabolism in the host gut. Annu Rev Microbiol 2015, 69, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.; Thiennimitr, P.; Winter, M.; Butler, B.; Huseby, D. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundarasamy, T.; Govindarajan, D.K.; Kandaswamy, K. A review on pilus assembly mechanisms in Gram-positive and Gram-negative bacteria. Cell Surface 2022, 8, 100077. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Rankin, S.C.; Blanchet, R.T.; Nulton, J.D.; Edwards, R.A.; Schifferli, D.M. Diversification of the Salmonella fimbriae: A model of macro-and microevolution. PLoS ONE 2012, 7, e38596. [Google Scholar] [CrossRef] [PubMed]
- Łaniewski, P.; Baek, C.-H.; Roland, K.L.; Curtiss Iii, R. Analysis of spleen-induced fimbria production in recombinant attenuated Salmonella enterica serovar Typhimurium vaccine strains. MBio 2017, 8, e01189-17. [Google Scholar] [CrossRef]
- Bao, H.; Wang, S.; Zhao, J.-H.; Liu, S.-L. Salmonella secretion systems: Differential roles in pathogen-host interactions. Microbiol. Res. 2020, 241, 126591. [Google Scholar] [CrossRef]
- Halte, M.; Erhardt, M. Protein export via the type III secretion system of the bacterial flagellum. Biomolecules 2021, 11, 186. [Google Scholar] [CrossRef]
- Lombardi, C.; Tolchard, J.; Bouillot, S.; Signor, L.; Gebus, C.; Liebl, D.; Fenel, D.; Teulon, J.-M.; Brock, J.; Habenstein, B. Structural and functional characterization of the type three secretion system (T3SS) needle of Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 573. [Google Scholar] [CrossRef]
- Lian, H.; Jiang, K.; Tong, M.; Chen, Z.; Liu, X.; Galán, J.E.; Gao, X. The Salmonella effector protein SopD targets Rab8 to positively and negatively modulate the inflammatory response. Nat. Microbiol. 2021, 6, 658–671. [Google Scholar] [CrossRef]
- Azimi, T.; Zamirnasta, M.; Sani, M.A.; Dallal, M.M.S.; Nasser, A. Molecular mechanisms of Salmonella effector proteins: A comprehensive review. Infect. Drug. Resist. 2020, 13, 11. [Google Scholar] [CrossRef]
- Dey, S.; Chakravarty, A.; Guha Biswas, P.; De Guzman, R.N. The type III secretion system needle, tip, and translocon. Protein Sci. 2019, 28, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Pallen, M.J.; Gophna, U. Bacterial flagella and Type III secretion: Case studies in the evolution of complexity. Gene Prot. Evol. 2007, 3, 30–47. [Google Scholar]
- Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Jennings, E.; Thurston, T.L.M.; Holden, D.W. Salmonella SPI-2 type III secretion system effectors: Molecular mechanisms and physiological consequences. Cell Host Microbe 2017, 22, 217–231. [Google Scholar] [CrossRef]
- Bai, X.; Nakatsu, C.H.; Bhunia, A.K. Bacterial biofilms and their implications in pathogenesis and food safety. Foods 2021, 10, 2117. [Google Scholar] [CrossRef]
- Bhunia, A.K. Food safety concerns of biofilm-forming zoonotic bacterial pathogens. Indian J. Anim. Health 2021, 60 (Suppl. S2), 44–51. [Google Scholar] [CrossRef]
- Asma, S.T.; Imre, K.; Morar, A.; Herman, V.; Acaroz, U.; Mukhtar, H.; Arslan-Acaroz, D.; Shah, S.R.A.; Gerlach, R. An overview of biofilm formation–combating strategies and mechanisms of action of antibiofilm agents. Life 2022, 12, 1110. [Google Scholar] [CrossRef]
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Harrell, J.E.; Hahn, M.M.; D’Souza, S.J.; Vasicek, E.M.; Sandala, J.L.; Gunn, J.S.; McLachlan, J.B. Salmonella Biofilm Formation, Chronic Infection, and Immunity Within the Intestine and Hepatobiliary Tract. Front. Cell. Infect. Microbiol. 2020, 10, 624622. [Google Scholar] [CrossRef]
- Beshiru, A.; Igbinosa, I.H.; Igbinosa, E.O. Biofilm formation and potential virulence factors of Salmonella strains isolated from ready-to-eat shrimps. PLoS ONE 2018, 13, e0204345. [Google Scholar] [CrossRef]
- González, J.F.; Tucker, L.; Fitch, J.; Wetzel, A.; White, P.; Gunn, J.S. Human bile-mediated regulation of Salmonella curli fimbriae. J. Bacteriol. 2019, 201, e00055-19. [Google Scholar] [CrossRef] [PubMed]
- Sokaribo, A.S.; Hansen, E.G.; McCarthy, M.; Desin, T.S.; Waldner, L.L.; MacKenzie, K.D.; Mutwiri Jr, G.; Herman, N.J.; Herman, D.J.; Wang, Y. Metabolic activation of CsgD in the regulation of Salmonella biofilms. Microorganisms 2020, 8, 964. [Google Scholar] [CrossRef] [PubMed]
- Obe, T.; Richards, A.K.; Shariat, N.W. Differences in biofilm formation of Salmonella serovars on two surfaces under two temperature conditions. J. Appl. Microbiol. 2022, 132, 2410–2420. [Google Scholar] [CrossRef]
- Nair, A.; Rawool, D.B.; Doijad, S.; Poharkar, K.; Mohan, V.; Barbuddhe, S.B.; Kolhe, R.; Kurkure, N.V.; Kumar, A.; Malik, S.V.S. Biofilm formation and genetic diversity of Salmonella isolates recovered from clinical, food, poultry and environmental sources. Infect. Gene. Evol. 2015, 36, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Piras, F.; Fois, F.; Consolati, S.G.; Mazza, R.; Mazzette, R. Influence of temperature, source, and serotype on biofilm formation of Salmonella enterica isolates from pig slaughterhouses. J. Food Prot. 2015, 78, 1875–1878. [Google Scholar] [CrossRef]
- Devaraj, A.; González, J.F.; Eichar, B.; Thilliez, G.; Kingsley, R.A.; Baker, S.; Allard, M.W.; Bakaletz, L.O.; Gunn, J.S.; Goodman, S.D. Enhanced biofilm and extracellular matrix production by chronic carriage versus acute isolates of Salmonella Typhi. PLoS Pathog 2021, 17, e1009209. [Google Scholar] [CrossRef]
- Darby, J.; Sheorey, H. Searching for Salmonella. Aust. J. Gen. Practice 2008, 37, 806. [Google Scholar]
- Meiring, J.E.; Shakya, M.; Khanam, F.; Voysey, M.; Phillips, M.T.; Tonks, S.; Thindwa, D.; Darton, T.C.; Dongol, S.; Karkey, A. Burden of enteric fever at three urban sites in Africa and Asia: A multicentre population-based study. Lancet Global Health 2021, 9, e1688–e1696. [Google Scholar] [CrossRef]
- Connor, B.A.; Schwartz, E. Typhoid and paratyphoid fever in travellers. Lancet Infect. Dis. 2005, 5, 623–628. [Google Scholar] [CrossRef]
- Bhan, M.K.; Bahl, R.; Bhatnagar, S. Typhoid and paratyphoid fever. Lancet 2005, 366, 749–762. [Google Scholar] [CrossRef]
- Thielman, N.M.; Guerrant, R.L. Acute infectious diarrhea. N. Engl. J. Med. 2004, 350, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.A.; Armstrong, M.; Morris-Jones, S.D.; Wright, S.G.; Doherty, T. Imported enteric fever: Case series from the hospital for tropical diseases, London, United Kingdom. Am. J. Trop. Med. 2010, 82, 1121. [Google Scholar] [CrossRef] [PubMed]
- Kuvandik, C.; Karaoglan, I.; Namiduru, M.; Baydar, I. Predictive value of clinical and laboratory findings in the diagnosis of enteric fever. New Microbiol. 2009, 32, 25. [Google Scholar]
- Johnson, R.; Mylona, E.; Frankel, G. Typhoidal Salmonella: Distinctive virulence factors and pathogenesis. Cell. Microbiol. 2018, 20, e12939. [Google Scholar] [CrossRef] [PubMed]
- Acheson, D.; Hohmann, E.L. Nontyphoidal salmonellosis. Clin. Infect. Dis. 2001, 32, 263–269. [Google Scholar] [CrossRef]
- Crump, J.A.; Kretsinger, K.; Gay, K.; Hoekstra, R.M.; Vugia, D.J.; Hurd, S.; Segler, S.D.; Megginson, M.; Luedeman, L.J.; Shiferaw, B. Clinical response and outcome of infection with Salmonella enterica serotype Typhi with decreased susceptibility to fluoroquinolones: A United States foodnet multicenter retrospective cohort study. Antimicrob Agents Chemother 2008, 52, 1278–1284. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Gilchrist, J.J.; MacLennan, C.A. Invasive nontyphoidal Salmonella disease in Africa. EcoSal Plus 2019, 8, 2. [Google Scholar] [CrossRef]
- Woods, D.F.; Reen, F.J.; Gilroy, D.; Buckley, J.; Frye, J.G.; Boyd, E.F. Rapid multiplex PCR and real-time TaqMan PCR assays for detection of Salmonella enterica and the highly virulent serovars Choleraesuis and Paratyphi C. J. Clin. Microbiol. 2008, 46, 4018–4022. [Google Scholar] [CrossRef]
- Guiney, D.G.; Fierer, J. The role of the spv genes in Salmonella pathogenesis. Front. Microbiol. 2011, 2, 129. [Google Scholar] [CrossRef]
- Gulig, P.A.; Doyle, T.J.; Hughes, J.A.; Matsui, H. Analysis of host cells associated with the Spv-mediated increased intracellular growth rate of Salmonella Typhimurium in mice. Infect. Immun. 1998, 66, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Shimoni, Z.; Pitlik, S.; Leibovici, L.; Samra, Z.; Konigsberger, H.; Drucker, M.; Agmon, V.; Ashkenazi, S.; Weinberger, M. Nontyphoid Salmonella bacteremia: Age-related differences in clinical presentation, bacteriology, and outcome. Clin. Infect. Dis. 1999, 28, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Arii, J.; Tanabe, Y.; Miyake, M.; Mukai, T.; Matsuzaki, M.; Niinomi, N.; Watanabe, H.; Yokota, Y.; Kohno, Y.; Noda, M. Clinical and pathologic characteristics of nontyphoidal Salmonella encephalopathy. Neurology 2002, 58, 1641–1645. [Google Scholar] [CrossRef]
- Gonzalez-Escobedo, G.; Gunn, J.S. Gallbladder epithelium as a niche for chronic salmonella carriage. Infect. Immun. 2013, 81, 2920–2930. [Google Scholar] [CrossRef]
- Gopinath, S.; Carden, S.; Monack, D. Shedding light on Salmonella carriers. Trends Microbiol. 2012, 20, 320–327. [Google Scholar] [CrossRef]
- Silva, C.; Calva, E.; Maloy, S. One health and food-borne disease: Salmonella transmission between humans, animals, and plants. Microbiol Spectr 2014, 2, 2-1. [Google Scholar] [CrossRef] [PubMed]
- Marín, C.; Balasch, S.; Vega, S.; Lainez, M. Sources of Salmonella contamination during broiler production in Eastern Spain. Prevent. Vet. Med. 2011, 98, 39–45. [Google Scholar] [CrossRef]
- Gole, V.C.; Woodhouse, R.; Caraguel, C.; Moyle, T.; Rault, J.-L.; Sexton, M.; Chousalkar, K. Dynamics of Salmonella shedding and welfare of hens in free-range egg production systems. Appl. Environ. Microbiol. 2017, 83, e03313-16. [Google Scholar] [CrossRef]
- Jahan, N.A.; Lindsey, L.L.; Larsen, P.A. The role of peridomestic rodents as reservoirs for zoonotic foodborne pathogens. Vector-Borne Zoonotic Dis. 2021, 21, 133–148. [Google Scholar] [CrossRef]
- Aung, K.T.; Khor, W.C.; Octavia, S.; Ye, A.; Leo, J.; Chan, P.P.; Lim, G.; Wong, W.K.; Tan, B.Z.Y.; Schlundt, J. Distribution of Salmonella serovars in humans, foods, farm animals and environment, companion and wildlife animals in Singapore. Int. J. Environ. Res. Public Health 2020, 17, 5774. [Google Scholar] [CrossRef]
- Dos Santos, E.J.E.; Azevedo, R.P.; Lopes, A.T.S.; Rocha, J.M.; Albuquerque, G.R.; Wenceslau, A.A.; Miranda, F.R.; Rodrigues, D.d.P.; Maciel, B.M. Salmonella spp. in wild free-living birds from Atlantic Forest fragments in southern Bahia, Brazil. BioMed Res. Int. 2020, 2020, 7594136. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Sanabria, R.; Alvarado, A.M. Preharvest Salmonella Risk Contamination and the Control Strategies; InTechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Saraf, S.; Jafra, B.S.; Ray, P.; Rawat, A.; Verma, S. Multidrug-Resistant Nontyphoidal Salmonella Associated with Invasive Disease in an Immunocompetent Child. Indian J. Pediatr. 2021, 88, 1266. [Google Scholar] [CrossRef]
- da Silva, K.E.; Tanmoy, A.M.; Pragasam, A.K.; Iqbal, J.; Sajib, M.S.I.; Mutreja, A.; Veeraraghavan, B.; Tamrakar, D.; Qamar, F.N.; Dougan, G. The international and intercontinental spread and expansion of antimicrobial-resistant Salmonella Typhi: A genomic epidemiology study. Lancet Microbe 2022, 3, e567–e577. [Google Scholar] [CrossRef]
- Mølbak, K.; Neimann, J. Risk factors for sporadic infection with Salmonella Enteritidis, Denmark, 1997–1999. Am. J. Epidemiol. 2002, 156, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.A.; Hargrett-Bean, N.T.; Blake, P.A. Salmonella typhi infections in the United States, 1975–1984: Increasing role of foreign travel. Rev. Infect. Dis. 1989, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.K.F.; Winstead, A.; Appiah, G.D.; Friedman, C.R.; Medalla, F.; Hughes, M.J.; Birhane, M.G.; Schneider, Z.D.; Marcenac, P.; Hanna, S.S. Update on extensively drug-resistant Salmonella serotype Typhi infections among travelers to or from Pakistan and report of ceftriaxone-resistant Salmonella serotype Typhi infections among travelers to Iraq—United States, 2018–2019. Morb. Mortal. Wkly. Rep. 2020, 69, 618. [Google Scholar] [CrossRef]
- Pitzer, V.E.; Meiring, J.; Martineau, F.P.; Watson, C.H.; Kang, G.; Basnyat, B.; Baker, S. The Invisible Burden: Diagnosing and Combatting Typhoid Fever in Asia and Africa. Clin. Infect. Dis. 2019, 69 (Suppl. S5), S395–S401. [Google Scholar] [CrossRef]
- Kim, C.L.; Espinoza, L.M.C.; Vannice, K.S.; Tadesse, B.T.; Owusu-Dabo, E.; Rakotozandrindrainy, R.; Jani, I.V.; Teferi, M.; Soura, A.B.; Lunguya, O. The Burden of Typhoid Fever in Sub-Saharan Africa: A Perspective. Res. Rep. Trop. Med. 2022, 13, 1–9. [Google Scholar] [CrossRef]
- Tack, B.; Vanaenrode, J.; Verbakel, J.Y.; Toelen, J.; Jacobs, J. Invasive non-typhoidal Salmonella infections in sub-Saharan Africa: A systematic review on antimicrobial resistance and treatment. BMC Med. 2020, 18, 212. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease “Burden of Illness”, S. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef]
- Gordon, M.A. Invasive nontyphoidal Salmonella disease: Epidemiology, pathogenesis and diagnosis. Curr. Opin. Infect. Dis. 2011, 24, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Ochiai, R.L.; Von Seidlein, L.; Dong, B.; Bhattacharya, S.K.; Agtini, M.D.; Bhutta, Z.A.; Do, G.C.; Ali, M.; Kim, D.R. Non-typhoidal Salmonella rates in febrile children at sites in five Asian countries. Trop. Med. Int. Health 2010, 15, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Ehuwa, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, food safety and food handling practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- CDC. Salmonella Outbreak Linked to Onions. Available online: https://www.cdc.gov/salmonella/oranienburg-09-21/index.html (accessed on 2 December 2021).
- van den Brom, R.; de Jong, A.; van Engelen, E.; Heuvelink, A.; Vellema, P. Zoonotic risks of pathogens from sheep and their milk borne transmission. Small Rumin Res. 2020, 189, 106123. [Google Scholar] [CrossRef] [PubMed]
- Dróżdż, M.; Małaszczuk, M.; Paluch, E.; Pawlak, A. Zoonotic potential and prevalence of Salmonella serovars isolated from pets. Infect. Ecol. Epidemiol. 2021, 11, 1975530. [Google Scholar] [CrossRef]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef]
- EFSA. The European union one health 2019 zoonoses report. EFSA J. 2021, 19, e06406. [Google Scholar]
- Stanaway, J.D.; Reiner, R.C.; Blacker, B.F.; Goldberg, E.M.; Khalil, I.A.; Troeger, C.E.; Andrews, J.R.; Bhutta, Z.A.; Crump, J.A.; Im, J. The global burden of typhoid and paratyphoid fevers: A systematic analysis for the global burden of disease study 2017. Lancet Infect. Dis. 2019, 19, 369–381. [Google Scholar] [CrossRef]
- N’cho, H.S. Notes from the field: Typhoid fever outbreak—Harare, Zimbabwe, October 2017–February 2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 44–45. [Google Scholar] [CrossRef]
- Gauld, J.S.; Olgemoeller, F.; Nkhata, R.; Li, C.; Chirambo, A.; Morse, T.; Gordon, M.A.; Read, J.M.; Heyderman, R.S.; Kennedy, N. Domestic river water use and risk of typhoid fever: Results from a case-control study in Blantyre, Malawi. Clin. Infect. Dis. 2020, 70, 1278–1284. [Google Scholar] [CrossRef]
- Feasey, N.A.; Masesa, C.; Jassi, C.; Faragher, E.B.; Mallewa, J.; Mallewa, M.; MacLennan, C.A.; Msefula, C.; Heyderman, R.S.; Gordon, M.A. Three epidemics of invasive multidrug-resistant Salmonella bloodstream infection in Blantyre, Malawi, 1998–2014. Clin. Infect. Dis. 2015, 61 (Suppl. S4), S363–S371. [Google Scholar] [CrossRef] [PubMed]
- Pulford, C.V.; Perez-Sepulveda, B.M.; Canals, R.; Bevington, J.A.; Bengtsson, R.J.; Wenner, N.; Rodwell, E.V.; Kumwenda, B.; Zhu, X.; Bennett, R.J. Stepwise evolution of Salmonella Typhimurium ST313 causing bloodstream infection in Africa. Nat. Microbiol. 2021, 6, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Ramatla, T.; Tawana, M.; Onyiche, T.E.; Lekota, K.E.; Thekisoe, O. One health perspective of Salmonella serovars in South Africa using pooled prevalence: Systematic review and meta-analysis. Int. J. Microbiol. 2022, 2022, 8952669. [Google Scholar] [CrossRef] [PubMed]
- Owowo, E.E.; Umoh, V.J.; Okon, I.E. Occurrence of Typhoidal and Non-Typhoidal Salmonellae among Poultry Workers in the Southern, Nigeria. Open. J. Med. Microbiol. 2019, 9, 201. [Google Scholar] [CrossRef]
- Mbae, C.; Mwangi, M.; Gitau, N.; Irungu, T.; Muendo, F.; Wakio, Z.; Wambui, R.; Kavai, S.; Onsare, R.; Wairimu, C. Factors associated with occurrence of salmonellosis among children living in Mukuru slum, an urban informal settlement in Kenya. BMC Infect. Dis. 2020, 20, 422. [Google Scholar] [CrossRef]
- Van Puyvelde, S.; Pickard, D.; Vandelannoote, K.; Heinz, E.; Barbé, B.; de Block, T.; Clare, S.; Coomber, E.L.; Harcourt, K.; Sridhar, S. An African Salmonella Typhimurium ST313 sublineage with extensive drug-resistance and signatures of host adaptation. Nat. Commun. 2019, 10, 4280. [Google Scholar] [CrossRef]
- Nikiema, M.E.M.; Kakou-Ngazoa, S.; Ky/Ba, A.; Sylla, A.; Bako, E.; Addablah, A.Y.A.; Ouoba, J.B.; Sampo, E.; Gnada, K.; Zongo, O. Characterization of virulence factors of Salmonella isolated from human stools and street food in urban areas of Burkina Faso. BMC Microbiol. 2021, 21, 338. [Google Scholar] [CrossRef]
- Abate, D.; Assefa, N. Prevalence and antimicrobial resistance patterns of Salmonella isolates in human stools and animal origin foods in Ethiopia: A systematic review and meta-analysis. Int. J. Health Sci. 2021, 15, 43. [Google Scholar]
- Fardsanei, F.; Dallal, M.M.S.; Douraghi, M.; Memariani, H.; Bakhshi, B.; Salehi, T.Z.; Nikkhahi, F. Antimicrobial resistance, virulence genes and genetic relatedness of Salmonella enterica serotype Enteritidis isolates recovered from human gastroenteritis in Tehran, Iran. J. Global Antimicrob. Resist. 2018, 12, 220–226. [Google Scholar] [CrossRef]
- Andrews-Polymenis, H.L.; Bäumler, A.J.; McCormick, B.A.; Fang, F.C. Taming the elephant: Salmonella biology, pathogenesis, and prevention. Infect. Immun. 2010, 78, 2356–2369. [Google Scholar] [CrossRef]
- Al-Rifai, R.H.; Chaabna, K.; Denagamage, T.; Alali, W.Q. Prevalence of enteric non-typhoidal Salmonella in humans in the Middle East and North Africa: A systematic review and meta-analysis. Zoonoses Public Health 2019, 66, 701–728. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, A.; Hashemi, F.B.; Heshteli, R.R.; Rahmani, M.; Halimi, S. Frequency of Salmonella serotypes among children in Iran: Antimicrobial susceptibility, biofilm formation, and virulence genes. BMC Pediatr. 2022, 22, 557. [Google Scholar] [CrossRef] [PubMed]
- Malaeb, M.; Bizri, A.R.; Ghosn, N.; Berry, A.; Musharrafieh, U. Salmonella burden in Lebanon. Epidemiol. Infect 2016, 144, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Israel, Y.; Muhsen, K.; Rokney, A.; Adler, A. Epidemiological and Clinical Characteristics of Non-Typhoidal Salmonella Bloodstream Infections in Central Israel: A Case-Control Study. Microorganisms 2022, 10, 1942. [Google Scholar] [CrossRef]
- Abd El Ghany, M.; Alsomali, M.; Almasri, M.; Regalado, E.P.; Naeem, R.; Tukestani, A.; Asiri, A.; Hill-Cawthorne, G.A.; Pain, A.; Memish, Z.A. Enteric infections circulating during Hajj seasons, 2011–2013. Emerg. Infect. Dis. 2017, 23, 1640. [Google Scholar] [CrossRef]
- Alharbi, N.A.; Alsaeed, T.S.; Aljohany, A.S.; Alwehaibi, K.K.; Almasaad, M.A.; Alotaibi, R.M.; Alotaibi, B.J.; Alamoudi, E.A. Extra-intestinal Salmonellosis in a Tertiary Care Center in Saudi Arabia. Sudanese J. Paediatr. 2021, 21, 152. [Google Scholar] [CrossRef]
- Crump, J.A.; Mintz, E.D. Global trends in typhoid and paratyphoid fever. Clin. Infect. Dis. 2010, 50, 241–246. [Google Scholar] [CrossRef]
- Diaz-Guevara, P.; Montaño, L.A.; Duarte, C.; Zabaleta, G.; Maes, M.; Martinez Angarita, J.C.; Thanh, D.P.; León-Quevedo, W.; Castañeda-Orjuela, C.; Alvarez Alvarez, C.J. Surveillance of Salmonella enterica serovar Typhi in Colombia, 2012–2015. PLoS Negl. Trop. Dis. 2020, 14, e0008040. [Google Scholar] [CrossRef] [PubMed]
- das Mercês Santos, A.F.; Amparo, L.F.V.; Machado, S.C.A.; Dias, T.S.; Berto, L.H.; da Costa Abreu, D.L.; de Aquino, M.H.C.; dos Prazeres Rodrigues, D.; de Almeida Pereira, V.L. Salmonella serovars associated with human salmonellosis in Brazil (2011–2020). Res. Soc. Dev. 2022, 11, e28011830533. [Google Scholar] [CrossRef]
- Calarga, A.P.; Gontijo, M.T.P.; de Almeida, L.G.P.; de Vasconcelos, A.T.R.; Nascimento, L.C.; de Moraes Barbosa, T.M.C.; de Carvalho Perri, T.M.; Dos Santos, S.R.; Tiba-Casas, M.R.; Marques, E.G.L. Antimicrobial resistance and genetic background of non-typhoidal Salmonella enterica strains isolated from human infections in São Paulo, Brazil (2000–2019). Braz. J. Microbiol. 2022, 53, 1249–1262. [Google Scholar] [CrossRef]
- Kuehn, B. Antibiotic resistance threat grows. JAMA 2019, 322, 2376. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.; Venkitanarayanan, K.; Kollanoor Johny, A. Antibiotic-resistant Salmonella in the food supply and the potential role of antibiotic alternatives for control. Foods 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Disease Outbreak News; Multi-Country Outbreak of Salmonella Typhimurium Linked to Chocolate Products–Europe and the United States of America. 2022. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON369 (accessed on 21 November 2022).
- European Food Safety Authority; European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar]
- Van Goethem, N.; Van Den Bossche, A.; Ceyssens, P.-J.; Lajot, A.; Coucke, W.; Vernelen, K.; Roosens, N.H.C.; De Keersmaecker, S.C.J.; Van Cauteren, D.; Mattheus, W. Coverage of the national surveillance system for human Salmonella infections, Belgium, 2016–2020. PLoS ONE 2021, 16, e0256820. [Google Scholar] [CrossRef] [PubMed]
- Larkin, L.; de la Gandara, M.P.; Hoban, A.; Pulford, C.; Jourdan-Da Silva, N.; de Valk, H.; Browning, L.; Falkenhorst, G.; Simon, S.; Lachmann, R. Investigation of an international outbreak of multidrug-resistant monophasic Salmonella Typhimurium associated with chocolate products, EU/EEA and United Kingdom, February to April 2022. Eurosurveillance 2022, 27, 2200314. [Google Scholar] [CrossRef]
- Plotogea, A.; Taylor, M.; Parayno, A.; Sillje, M.; Stone, J.; Byrnes, R.; Bitzikos, O.; Redford, T.; Waters, S.; Fraser, E. Human Salmonella Enteritidis illness outbreak associated with exposure to live mice in British Columbia, Canada, 2018–2019. Zoonoses Pub. Health 2022, 69, 856–863. [Google Scholar] [CrossRef]
- Whitney, B.M.; McClure, M.; Hassan, R.; Pomeroy, M.; Seelman, S.L.; Singleton, L.N.; Blessington, T.; Hardy, C.; Blankenship, J.; Pereira, E. A Series of Papaya-Associated Salmonella Illness Outbreak Investigations in 2017 and 2019: A Focus on Traceback, Laboratory, and Collaborative Efforts. J. Food Prot. 2021, 84, 2002–2019. [Google Scholar] [CrossRef]
- Motladiile, T.W. Salmonella food-poisoning outbreak linked to the National School Nutrition Programme, North West province, South Africa. South African J. Infect. Dis. 2019, 34, 124. [Google Scholar] [CrossRef]
- Mörk, M.J.; Karamehmedovic, N.; Hansen, A.; Öhd, J.N.; Lindblad, M.; Östlund, E.; Rehn, M.; Jernberg, C. Outbreak of Salmonella Newport linked to imported frozen cooked crayfish in dill brine, Sweden, July to November 2019. Eurosurveillance 2022, 27, 2100918. [Google Scholar]
- CDC. Salmonella; CDC: Atlanta, GA, USA, 2020. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 21 November 2022).
- Huber, L.; Agunos, A.; Gow, S.P.; Carson, C.A.; Van Boeckel, T.P. Reduction in Antimicrobial Use and Resistance to Salmonella, Campylobacter, and Escherichia coli in Broiler Chickens, Canada, 2013–2019. Emerg. Infect. Dis. 2021, 27, 2434. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low-and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.W.; Moore, J.E.; Rao, J.R. Antimicrobial resistance (AMR): Significance to food quality and safety. Food Qual. Safety 2019, 3, 15–22. [Google Scholar] [CrossRef]
- Al-Ansari, M.M.; Aljubali, M.M.; Somily, A.M.; Albarrag, A.M.; Masood, A. Isolation and molecular characterization of multidrug-resistant Salmonella enterica serovars. J. Infect. Public Health 2021, 14, 1767–1776. [Google Scholar] [CrossRef]
- Yoke-Kqueen, C.; Learn-Han, L.; Noorzaleha, A.S.; Son, R.; Sabrina, S.; Jiun-Horng, S.; Chai-Hoon, K. Characterization of multiple-antimicrobial-resistant Salmonella enterica subsp. enterica isolated from indigenous vegetables and poultry in Malaysia. Lett. Appl. Microbiol. 2008, 46, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Bhunia, A.K. Animal-use antibiotics induce cross-resistance in bacterial pathogens to human therapeutic antibiotics. Curr. Microbiol. 2019, 76, 1112–1117. [Google Scholar] [CrossRef]
- EFSA. The European Union Summary Report on Antimicrobial Resistance in zoonotic and indicator bacteria from humans, animals and food in 2019–2020. EFSA J. 2022, 20, e07209. [Google Scholar]
- Beutlich, J.; Jahn, S.; Malorny, B.; Hauser, E.; Hühn, S.; Schroeter, A.; Rodicio, M.R.; Appel, B.; Threlfall, J.; Mevius, D. Antimicrobial resistance and virulence determinants in European Salmonella genomic island 1-positive Salmonella enterica isolates from different origins. Appl. Environ. Microbiol. 2011, 77, 5655–5664. [Google Scholar] [CrossRef] [PubMed]
- Argüello, H.; Guerra, B.; Rodríguez, I.; Rubio, P.; Carvajal, A. Characterization of antimicrobial resistance determinants and class 1 and class 2 integrons in Salmonella enterica spp. multidrug-resistant isolates from pigs. Genes 2018, 9, 256. [Google Scholar] [CrossRef]
- Wójcicki, M.; Świder, O.; Daniluk, K.J.; Średnicka, P.; Akimowicz, M.; Roszko, M.Ł.; Sokołowska, B.; Juszczuk-Kubiak, E. Transcriptional regulation of the multiple resistance mechanisms in Salmonella—A review. Pathogens 2021, 10, 801. [Google Scholar] [CrossRef]
- Chang, M.-X.; Zhang, J.-F.; Sun, Y.-H.; Li, R.-S.; Lin, X.-L.; Yang, L.; Webber, M.A.; Jiang, H.-X. Contribution of different mechanisms to Ciprofloxacin resistance in Salmonella spp. Front. Microbiol. 2021, 12, 663731. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Lee, S.-K.; Park, M.-S.; Na, H.-T. Analysis of the fluoroquinolone antibiotic resistance mechanism of Salmonella enterica isolates. J. Microbiol. Biotechnol. 2016, 26, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A. Plasmid-mediated antimicrobial resistance in Salmonella enterica. Curr. Issue Mol. Biol. 2003, 5, 113–122. [Google Scholar]
- Chaudhari, R.; Singh, K.; Kodgire, P. Biochemical and Molecular Mechanisms of Antibiotic Resistance in Salmonella spp. Res. Microbiol. 2022, 174, 103985. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 481–511. [Google Scholar] [CrossRef]
- Hooper, D.C.; Jacoby, G.A. Mechanisms of drug resistance: Quinolone resistance. Annals New York Acad. Sci. 2015, 1354, 12–31. [Google Scholar] [CrossRef]
- Hooper, D.C.; Jacoby, G.A. Topoisomerase inhibitors: Fluoroquinolone mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025320. [Google Scholar] [CrossRef]
- Ugboko, H.; De, N. Mechanisms of Antibiotic resistance in Salmonella Typhi. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 461–476. [Google Scholar]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Correia, S.; Poeta, P.; Hébraud, M.; Capelo, J.L.; Igrejas, G. Mechanisms of quinolone action and resistance: Where do we stand? J. Med. Microbiol. 2017, 66, 551–559. [Google Scholar] [CrossRef]
- Zaki, S.A.; Karande, S. Multidrug-resistant typhoid fever: A review. J. Infect. Dev. Ctries. 2011, 5, 324–337. [Google Scholar] [CrossRef]
- Holmberg, S.D.; Osterholm, M.T.; Senger, K.A.; Cohen, M.L. Drug-resistant Salmonella from animals fed antimicrobials. N. Eng. J. Med. 1984, 311, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Sodagari, H.R.; Habib, I.; Shahabi, M.P.; Dybing, N.A.; Wang, P.; Bruce, M. A review of the public health challenges of Salmonella and turtles. Vet. Sci. 2020, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Karon, A.E.; Archer, J.R.; Sotir, M.J.; Monson, T.A.; Kazmierczak, J.J. Human multidrug-resistant Salmonella Newport infections, Wisconsin, 2003–2005. Emerg. Infect. Dis. 2007, 13, 1777. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Dia, N.M.; Gautret, P.; Benkouiten, S.; Belhouchat, K.; Drali, T.; Parola, P.; Brouqui, P.; Memish, Z.; Raoult, D. Acquisition of extended-spectrum cephalosporin-and colistin-resistant Salmonella enterica subsp. enterica serotype Newport by pilgrims during Hajj. Int. J. Antimicrob. Agents 2015, 45, 600–604. [Google Scholar] [CrossRef] [PubMed]
- CDC. Antibiotic Resistance Threats in the United States. 2019. The Full 2019 AR Threats Report, including Methods and Appendices. Available online: www.cdc.gov/DrugResistance/Biggest-Threats.html (accessed on 19 November 2022).
- Pérez-Rodríguez, F.; Mercanoglu Taban, B. A state-of-art review on multi-drug resistant pathogens in foods of animal origin: Risk factors and mitigation strategies. Front. Microbiol. 2019, 10, 2091. [Google Scholar] [CrossRef]
- Crump, J.A. Progress in typhoid fever epidemiology. Clin. Infect. Dis. 2019, 68 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef]
- Clasen, T.; Roberts, I.; Rabie, T.; Schmidt, W.; Cairncross, S. Cochrane review: Interventions to improve water quality for preventing diarrhoea. Evid. Based Child Health A Cochrane Rev. J. 2007, 2, 226–329. [Google Scholar] [CrossRef]
- Stefanova, R.; Vasilev, N.; Spassov, S. Irradiation of food, current legislation framework, and detection of irradiated foods. Food Anal. Methods 2010, 3, 225–252. [Google Scholar] [CrossRef]
- Roberts, P.B. Food irradiation: Standards, regulations and world-wide trade. Radiation Phys. Chem. 2016, 129, 30–34. [Google Scholar] [CrossRef]
- Gayet, R.; Bioley, G.; Rochereau, N.; Paul, S.; Corthésy, B. Vaccination against Salmonella infection: The mucosal way. Microbiol. Mol. Biol. Rev. 2017, 81, e00007-17. [Google Scholar] [CrossRef]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Human Vac. Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed]
- Tennant, S.M.; MacLennan, C.A.; Simon, R.; Martin, L.B.; Khan, M.I. Nontyphoidal Salmonella disease: Current status of vaccine research and development. Vaccine 2016, 34, 2907–2910. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.C.; Ho, V.A.; Khiem, H.B.; Trach, D.D.; Bay, P.V.; Thanh, T.C.; Kossaczka, Z.; Bryla, D.A.; Shiloach, J.; Robbins, J.B. The efficacy of a Salmonella Typhi Vi conjugate vaccine in two-to-five-year-old children. N. Engl. J. Med. 2001, 344, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Angulo, F.J.; Talbot, E.A.; Gagnon, E.R.; Greenblatt, J. Common Ground for the Control of Multidrug-Resistant Salmonella in Ground Beef. Clin. Infect. Dis. 2006, 42, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Bhunia, A.K. Microbes as a tool to defend against antibiotic resistance in food animal production. Indian J. Anim. Hlth 2019, 58, 1–18. [Google Scholar] [CrossRef]
- Tellez, G.; Pixley, C.; Wolfenden, R.E.; Layton, S.L.; Hargis, B.M. Probiotics/direct fed microbials for Salmonella control in poultry. Food Res. Int. 2012, 45, 628–633. [Google Scholar] [CrossRef]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (direct-fed microbials) in poultry nutrition and their effects on nutrient utilization, growth and laying performance, and gut health: A systematic review. Animals 2020, 10, 1863. [Google Scholar] [CrossRef]
- Salim, H.M.; Kang, H.K.; Akter, N.; Kim, D.W.; Kim, J.H.; Kim, M.J.; Na, J.C.; Jong, H.B.; Choi, H.C.; Suh, O.S. Supplementation of direct-fed microbials as an alternative to antibiotic on growth performance, immune response, cecal microbial population, and ileal morphology of broiler chickens. Poult. Sci. 2013, 92, 2084–2090. [Google Scholar] [CrossRef]
- Burkholder, K.; Bhunia, A. Salmonella enterica serovar Typhimurium adhesion and cytotoxicity during epithelial cell stress is reduced by Lactobacillus rhamnosus GG. Gut Pathog. 2009, 1, 14. [Google Scholar] [CrossRef]
Figure 2.
Schematic demonstration of (A) resistance mechanism of Salmonella to phagocytic destruction: 1. prevention of phagolysosome fusion, 2. resistance to oxidative and nitrosative stress, 3. resistance to antimicrobial peptides, 4. inhibition of macrophage-mediated inflammation). (B) Mechanisms of internalization (course of infection) of Salmonella. Once Salmonella reaches the lumen of the gut, they can pass the epithelial layer either by (1) a passive uptake expedited by dendritic cells; (2) invasion via the M cells; (3 and 4) bacterial replication and survival; (5) secretion of cytokines, inflammation, and/or apoptosis; (6 and 7) basolateral invasion to reseed back into the lumen of the gut, which leads to shedding of the agent via feces. (C) Colonization of non-phagocytic host cells (fibroblast, endothelial, and epithelial cells) by a trigger mechanism. SCV, Salmonella-containing vacuole.
Figure 2.
Schematic demonstration of (A) resistance mechanism of Salmonella to phagocytic destruction: 1. prevention of phagolysosome fusion, 2. resistance to oxidative and nitrosative stress, 3. resistance to antimicrobial peptides, 4. inhibition of macrophage-mediated inflammation). (B) Mechanisms of internalization (course of infection) of Salmonella. Once Salmonella reaches the lumen of the gut, they can pass the epithelial layer either by (1) a passive uptake expedited by dendritic cells; (2) invasion via the M cells; (3 and 4) bacterial replication and survival; (5) secretion of cytokines, inflammation, and/or apoptosis; (6 and 7) basolateral invasion to reseed back into the lumen of the gut, which leads to shedding of the agent via feces. (C) Colonization of non-phagocytic host cells (fibroblast, endothelial, and epithelial cells) by a trigger mechanism. SCV, Salmonella-containing vacuole.
Figure 3.
Schematic elucidation of the different types of secretion systems that participate in Salmonella infection, namely I (T1SS), III (T3SS), IV (T4SS), and VI (T6SS). HM, host cell membrane; IM, bacterial inner membrane; OM, outer membrane.
Figure 3.
Schematic elucidation of the different types of secretion systems that participate in Salmonella infection, namely I (T1SS), III (T3SS), IV (T4SS), and VI (T6SS). HM, host cell membrane; IM, bacterial inner membrane; OM, outer membrane.
Figure 4.
Clinical manifestation of human salmonellosis. The first clinical presentation is gastroenteritis complemented by headache, nausea, vomiting, abdominal pain, non-bloody diarrhea, and myalgia. The second clinical manifestation is bacteremia, other extraintestinal complications, and splenomegaly, hepatomegaly, and urinary tract infections.
Figure 4.
Clinical manifestation of human salmonellosis. The first clinical presentation is gastroenteritis complemented by headache, nausea, vomiting, abdominal pain, non-bloody diarrhea, and myalgia. The second clinical manifestation is bacteremia, other extraintestinal complications, and splenomegaly, hepatomegaly, and urinary tract infections.
Figure 5.
Diagram showing the primary reservoir, source of infection, and the transmission mechanism of human salmonellosis. Insects, reptiles, humans, and animals can serve as a reservoir for human salmonellosis. Uncooked, undercooked, and/or contaminated foods (poultry, beef, pork, egg, milk, milk products, vegetables, flour, nuts, etc.) are familiar sources of infection.
Figure 5.
Diagram showing the primary reservoir, source of infection, and the transmission mechanism of human salmonellosis. Insects, reptiles, humans, and animals can serve as a reservoir for human salmonellosis. Uncooked, undercooked, and/or contaminated foods (poultry, beef, pork, egg, milk, milk products, vegetables, flour, nuts, etc.) are familiar sources of infection.
Figure 6.
Schematic showing antibiotic resistance mechanisms in Salmonella spp.
Figure 6.
Schematic showing antibiotic resistance mechanisms in Salmonella spp.
Table 1.
Summary of worldwide Salmonella causing diarrheal diseases (from 2019–2022).
Table 1.
Summary of worldwide Salmonella causing diarrheal diseases (from 2019–2022).
| Year | Salmonella enterica Serovar | No of Cases | Source of Country | Food Source (s) | References |
|---|
| 2018 | Concord | NA | Israel | Tahini products | [3] |
| 2018 | Unidentified serovar | NA | Australia | Chicken sandwich | [3] |
| 2019 | Unidentified serovar | NA | USA | Backyard poultry | [3] |
| 2021 | Oranienburg | 1040 | USA | Onion | [129] |
| 2022 | Typhimurium | 324 | Europe and USA | Chocolate products | [162] |
| 2022 | Enteritidis | NA | Canada | Exposure to live mice | [163] |
| 2017 and 2019 | Multiple serovars | 325 | United States | Whole, fresh Maradol papayas | [164] |
| 2019 | Heidelberg | 164 (48.5%) | North West Province, South Africa | School lunch at public primary day school | [165] |
| 2019 | Newport | 25 | Sweden | Imported frozen cooked crayfish in dill brine | [166] |
| 2019 | Oranienburg | 26 | USA (14 states) | Contact with pet turtles | [167] |
| 2019 | Six different serovars: Amsterdam, Havana, Kintambo, Mbandaka, Orion, and Senftenberg) | 121 | Five EU/EEA countries | Imported sesame-based products (originating from Syria) | [133] |
Table 2.
Complete susceptibility and multi-drug resistance (MDR) in Salmonella spp. isolated from humans in 2020 in Europe.
Table 2.
Complete susceptibility and multi-drug resistance (MDR) in Salmonella spp. isolated from humans in 2020 in Europe.
| Country * | Susceptibility to All Tested Antibiotics (%) | Multi-Resistant (%) |
|---|
| Austria (N = 894) | 61.5 | 12.9 |
| Belgium (N = 706) | 39.8 | 35.0 |
| Denmark (N = 252) | 71.8 | 19.8 |
| Estonia (N = 95) | 68.4 | 22.1 |
| France (N = 713) | 64.7 | 19.9 |
| Ireland (N = 713) | 65.4 | 12.6 |
| Italy (N = 872) | 42.7 | 42.9 |
| Luxembourg (N = 90) | 41.1 | 31.1 |
| Netherlands (N = 494) | 55.3 | 28.9 |
| Portugal (N = 238) | 50.8 | 29.8 |
| Romania (N = 36) | 38.9 | 11.1 |
| Slovenia (N = 172) | 62.8 | 19.8 |
| Spain (N = 766) | 55.6 | 24.2 |
| Sweden (N = 344) | 77.6 | 15.1 |
| Total | 55.9 | 25.4 |
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).

 ,
, 






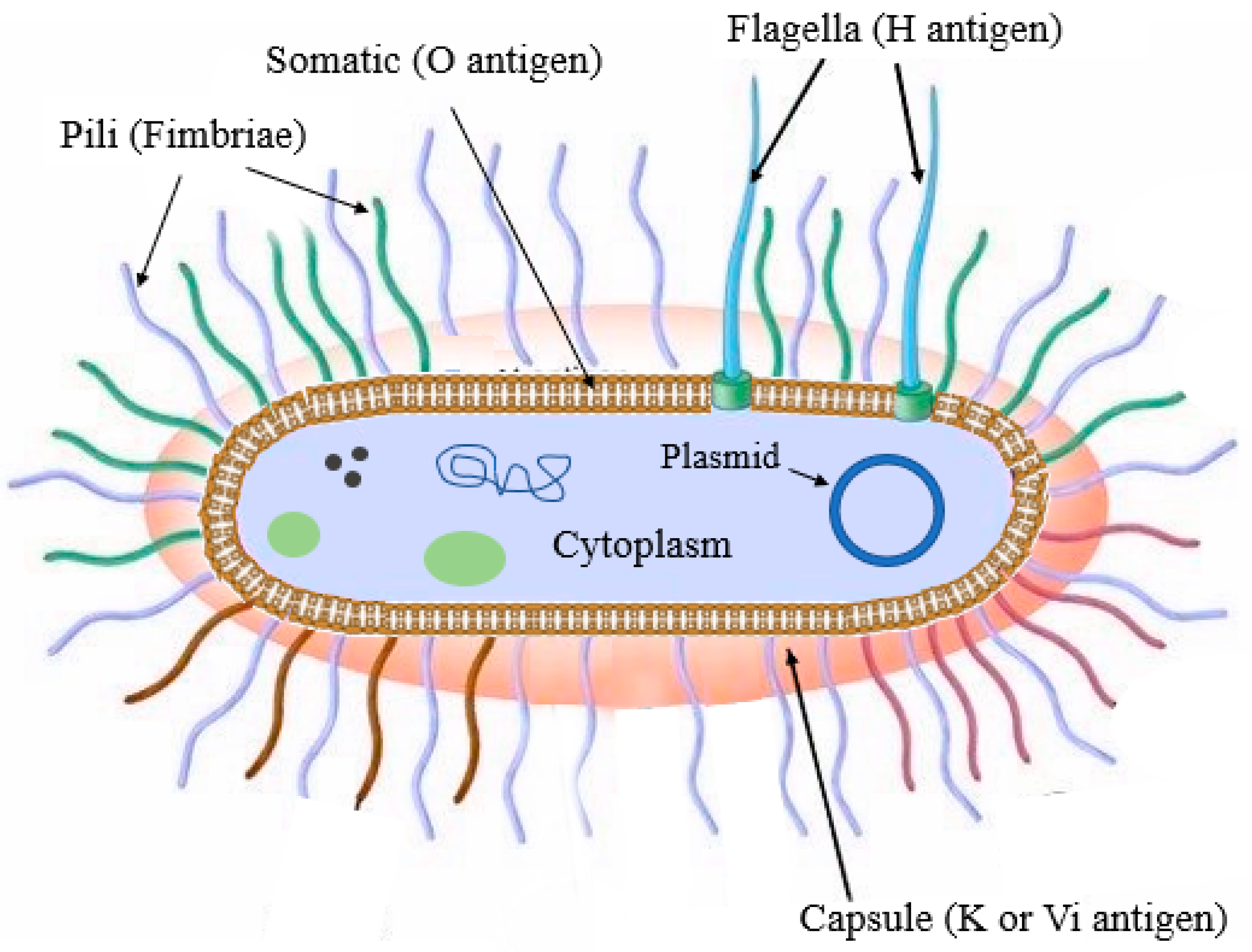{kind=link}
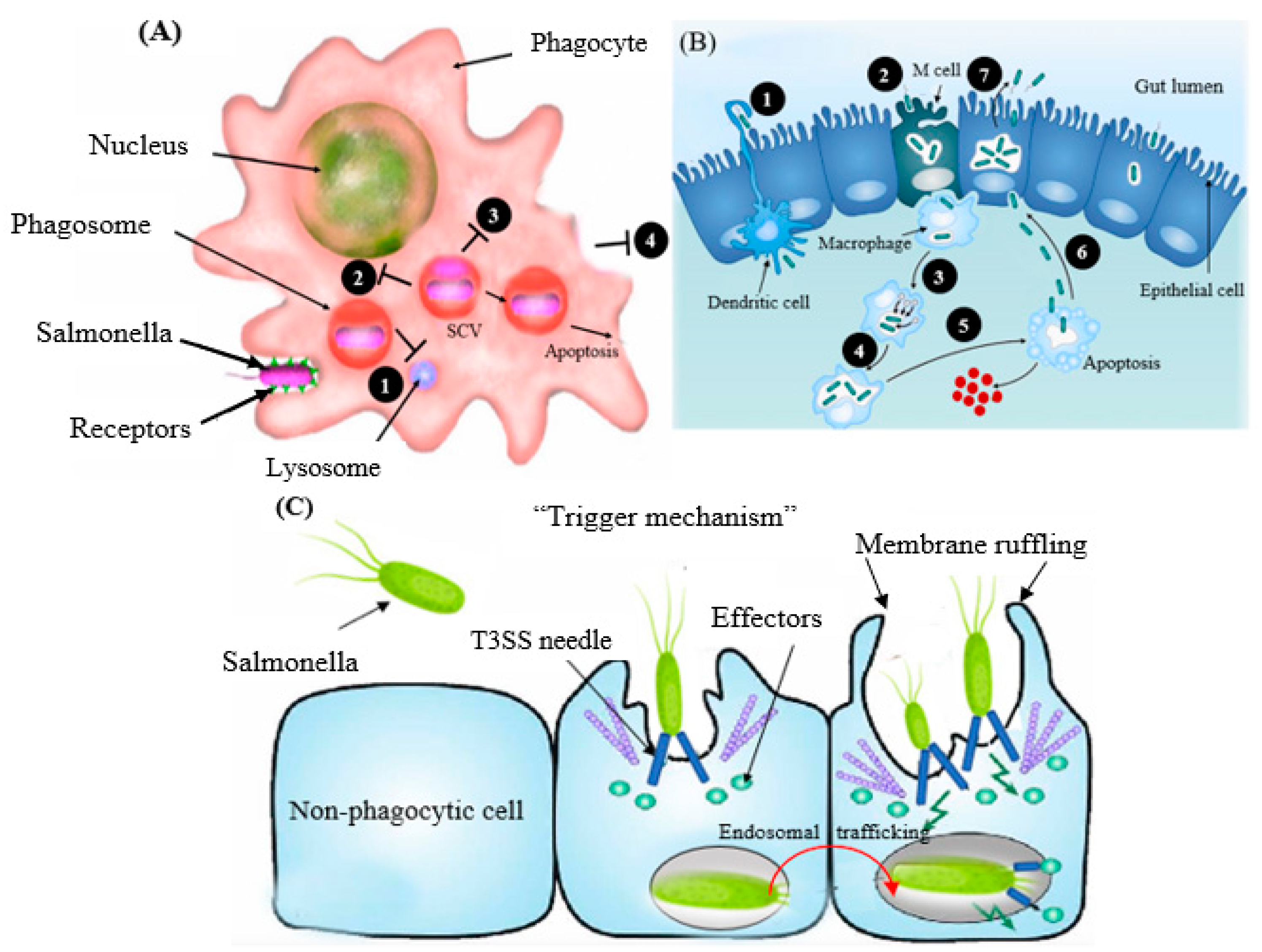{kind=link}
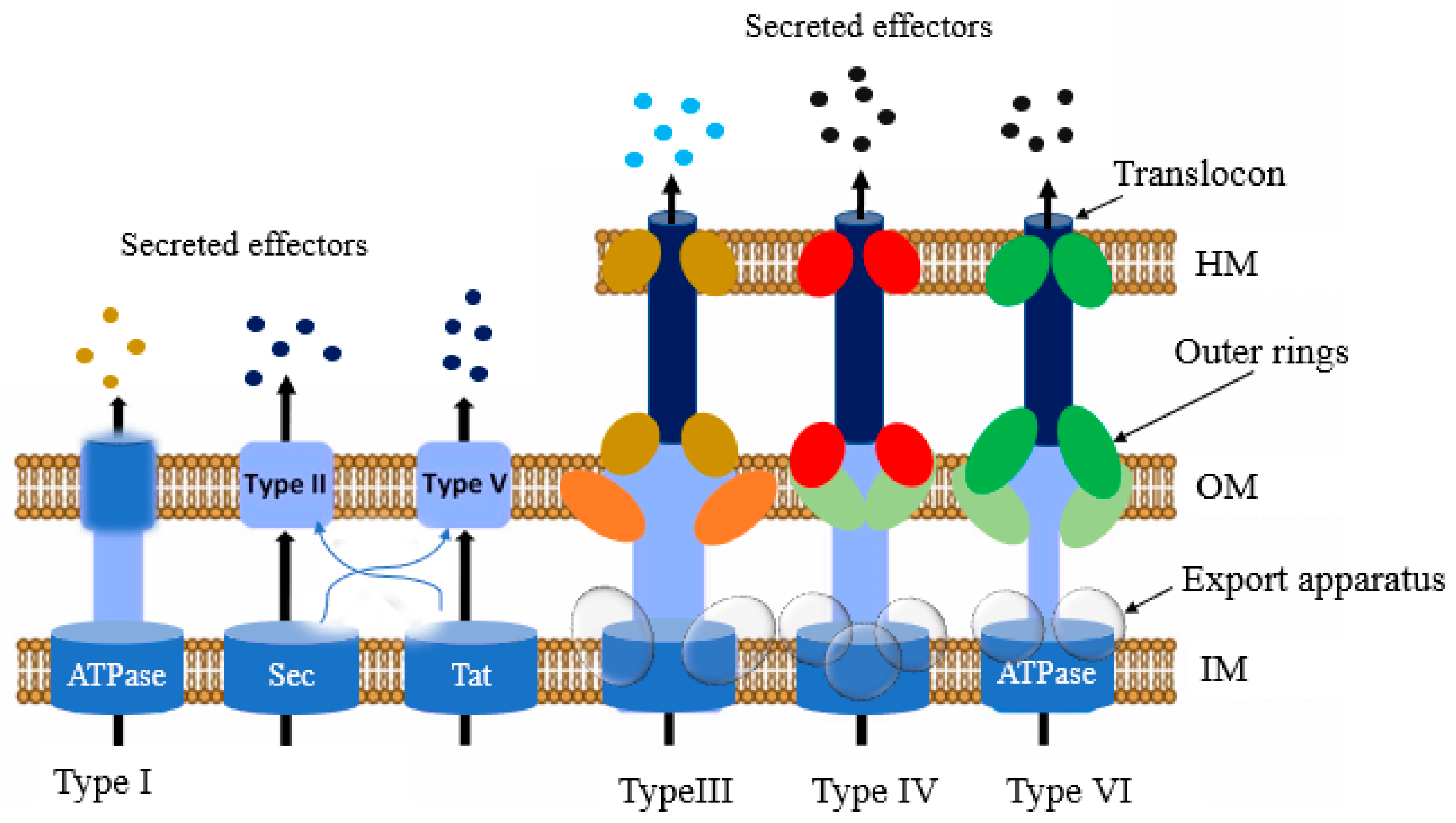{kind=link}
{kind=link}
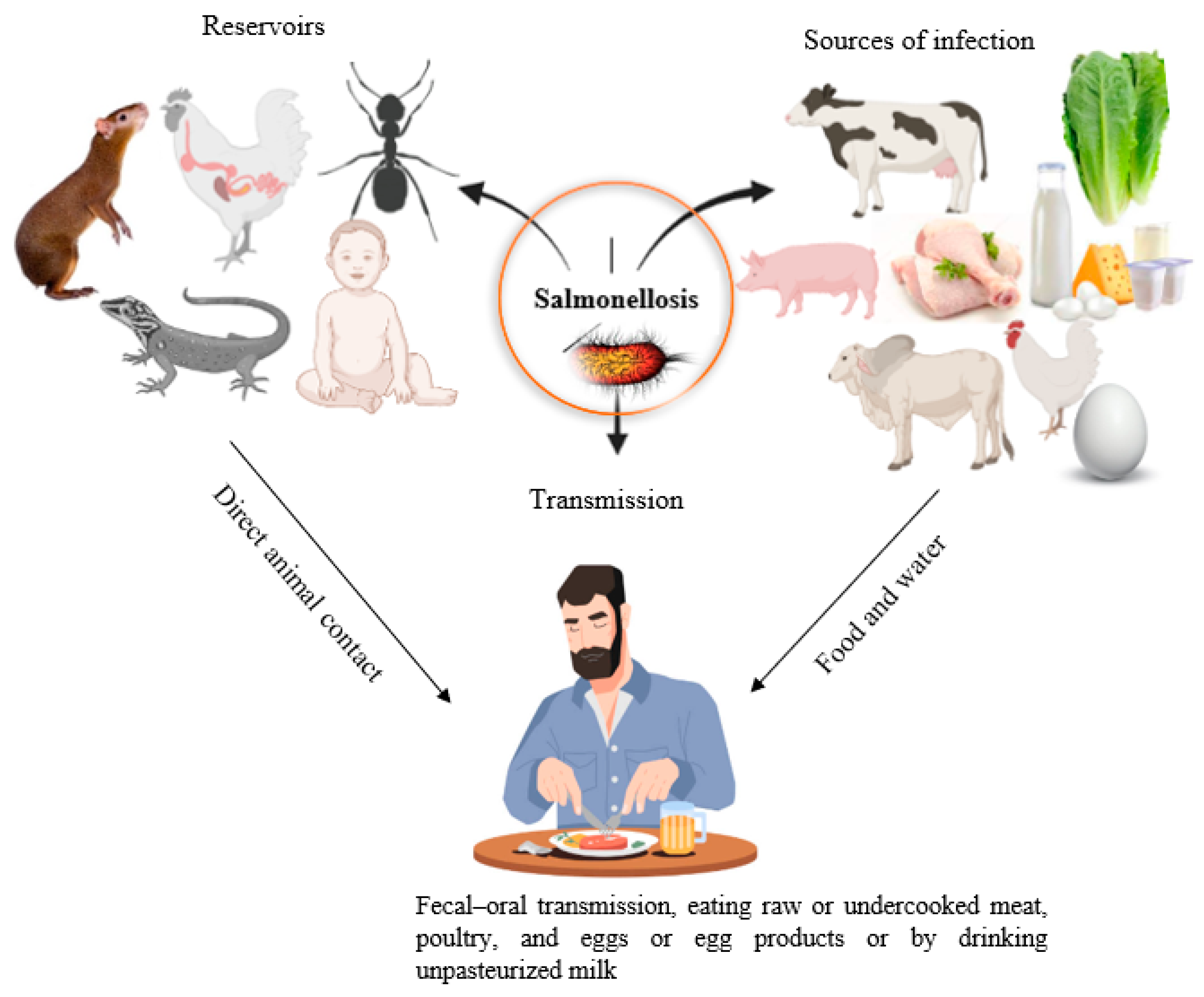{kind=link}
{kind=link}