
The Identification of a Strain for the Biological Purification of Soy Molasses to Produce Functional Soy Oligosaccharides and Optimize Purification Conditions
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
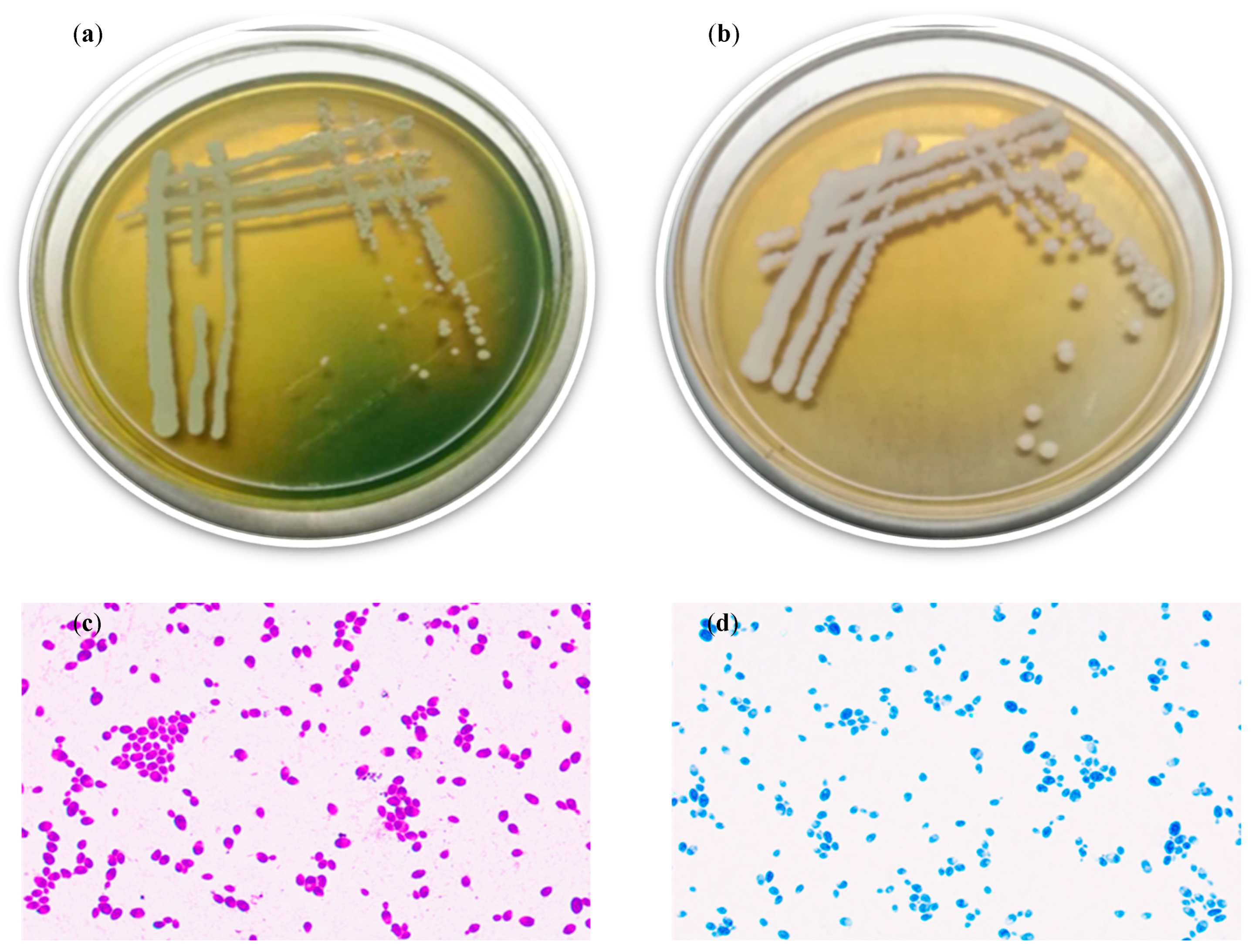
2.2. Identification and Biochemical Characteristics of Strain YT312
2.2.1. Morphological Observation
2.2.2. Physiological and Biochemical Characteristics
2.2.3. Molecular Identification
2.2.4. Strain Performance Determination
2.2.5. Determination of Strain Growth Curve
2.3. Optimization of Fermentation Conditions by Single-Factor Experiments
2.4. Optimization of Fermentation Conditions by Orthogonal Experiment
2.5. Verification of Fermentation Conditions for Soy Molasses Purification
2.6. Determination of Sucrose, Stachyose, Raffinose, and Manninotriose
2.7. Determination of Cell Density
2.8. Determination of Comprehensive Index
2.8.1. Determination of Range Normalization Values
2.8.2. Determination of Weights
2.8.3. Determination of Comprehensive Index
2.9. Data Processing
3. Results and Discussion
3.1. Identification and Biological Characteristics of Strain YT312
3.1.1. Identification of Strain YT312
- Observation of colony morphology and cell structure
- 2.
- Physiological and biochemical characteristics
- 3.
- Molecular identification
3.1.2. Biological Characteristics of W. anomalus YT312
3.1.3. Growth Curve of W. anomalus YT312
3.2. Optimization of Fermentation Conditions for Soy Molasses Purification by Single-Factor Experiments
3.2.1. Effect of Inoculum Size of W. anomalus YT312 on Soy Molasses Purification
3.2.2. Effect of Fermentation Temperature on Soy Molasses Purification by W. anomalus YT312
3.2.3. Effect of Shaking Speed on Soy Molasses Purification by W. anomalus YT312
3.2.4. Effect of Dilution Ratio on the Soy Molasses Purification by W. anomalus YT312
3.2.5. Effect of pH Value on the Soy Molasses Purification by W. anomalus YT312
3.2.6. Effect of Fermentation Time on the Soy Molasses Purification by W. anomalus YT312
3.3. Optimization of Fermentation Conditions for Soy Molasses Purification by Orthogonal Experiment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Da Silva, F.B.; Romao, B.B.; Cardoso, V.L.; Filho, U.C.; Ribeiro, E.J. Production of ethanol from enzymatically hydrolyzed soybean molasses. Biochem. Eng. J. 2012, 69, 61–68. [Google Scholar] [CrossRef]
- Rakita, S.; Banjac, V.; Djuragic, O.; Cheli, F.; Pinotti, L. Soybean molasses in animal nutrition. Animals 2021, 11, 514. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Tian, J.J.; Wang, D. Comprehensive utilization of soy molasses. Sci. Technol. Cereals Oils Foods 2010, 18, 22–24. [Google Scholar] [CrossRef]
- Zhang, L.L.; Yang, W.W.; Zhang, Y.Z. Study on soybean oligosaccharide separated from soy molasses. Sci. Technol. Food Ind. 2009, 30, 258–260. [Google Scholar] [CrossRef]
- Long, C.C.; Gibbons, W.R. Conversion of soy molasses, soy solubles, and dried soybean carbohydrates into ethanol. Int. J. Agric. Biol. Eng. 2013, 6, 62–68. [Google Scholar] [CrossRef]
- Rodrigues, M.S.; Moreira, F.S.; Cardoso, V.L.; de Resende, M.M. Soy molasses as a fermentation substrate for the production of biosurfactant using Pseudomonas aeruginosa ATCC 10145. Environ. Sci. Pollut. Res. 2017, 24, 18699–18709. [Google Scholar] [CrossRef] [PubMed]
- Sancheti, A.; Thompson, E.R.; Ju, L.K. Factors influencing the enzymatic hydrolysis of soy molasses into fermentation feedstock. Enzym. Microb. Technol. 2023, 170, 110302. [Google Scholar] [CrossRef]
- Cheng, C.; Zhou, Y.P.; Lin, M.; Wei, P.L.; Yang, S.T. Polymalic acid fermentation by Aureobasidium pullulans for malic acid production from soybean hull and soy molasses: Fermentation kinetics and economic analysis. Bioresour. Technol. 2017, 223, 166–174. [Google Scholar] [CrossRef]
- Duru, K.C.; Slesarev, G.P.; Aboushanab, S.A.; Kovalev, I.S.; Zeidler, D.M.; Kovaleva, E.G.; Bhat, R. An eco-friendly approach to enhance the extraction and recovery efficiency of isoflavones from kudzu roots and soy molasses wastes using ultrasound-assisted extraction with natural deep eutectic solvents (NADES). Ind. Crop. Prod. 2022, 182, 114886. [Google Scholar] [CrossRef]
- Aboushanab, S.A.; Shevyrin, V.A.; Slesarev, G.P.; Melekhin, V.V.; Shcheglova, A.V.; Makeev, O.G.; Kovaleva, E.G.; Kim, K.H. Antioxidant and cytotoxic activities of kudzu roots and soy molasses against pediatric tumors and phytochemical analysis of isoflavones using HPLC-DAD-ESI-HRMS. Plants 2022, 11, 741. [Google Scholar] [CrossRef]
- Batista, V.S.F.; Nunes, G.L.; Viegas, G.I.; Lucas, B.N.; Bochi, V.C.; Emanuelli, T.; Barin, J.S.; de Menezes, C.R.; da Rosa, C.S. Extraction, characterization and microencapsulation of isoflavones from soybean molasses. Cienc. Rural 2020, 50, e20190341. [Google Scholar] [CrossRef]
- Ni, C.L.; Xu, L.; Zhang, G.P.; Cheng, J.J.; Gao, L. Purification and the antioxidant properties of saponins in soy molasses. Sci. Technol. Food Ind. 2017, 38, 153–157+162. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.Z.; Jiang, L.Z.; Li, C.G.; Qu, Y.Y. Extraction of soybean saponin from soy molasses by ultrasonic wave. J. Chin. Cer. Oils Assoc. 2009, 24, 23–26. [Google Scholar]
- Liu, Y.; Hua, Y.F. Separation and purification of phospholipids from soy molasses. China Oils Fats 2008, 33, 61–64. [Google Scholar] [CrossRef]
- Rakicka-Pustulka, M.; Ziuzia, P.; Pierwola, J.; Szymanski, K.; Wrobel-Kwiatkowska, M.; Lazar, Z. The microbial production of kynurenic acid using Yarrowia lipolytica yeast growing on crude glycerol and soybean molasses. Front. Bioeng. Biotechnol. 2022, 10, 936137. [Google Scholar] [CrossRef] [PubMed]
- Lima, F.A.; Rola, J.C.; de Freitas, M.M.G.; de Afonso, J.M.M.D.; de Resende, M.M. Acid phosphatase immobilization and production study by Trichoderma spp. in soybean molasses. Chem. Eng. Technol. 2022, 45, 979–984. [Google Scholar] [CrossRef]
- Gomes, R.J.; Faria-Tischer, P.C.D.; Tischer, C.A.; Constantino, L.V.; Rosa, M.D.; Chideroli, R.T.; Pereira, U.D.; Spinosa, W.A. Komagataeibacter intermedius V-05: An acetic acid bacterium isolated from vinegar industry, with high capacity for bacterial cellulose production in soybean molasses medium. Food Technol. Biotechnol. 2021, 59, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.C.G.; de Mello, B.S.; Araujo, M.L.G.D.; da Silva, G.H.R.; Sarti, A. Soybean molasses as feedstock for sustainable generation of biomethane using high-rate anaerobic reactor. J. Environ. Chem. Eng. 2021, 9, 105226. [Google Scholar] [CrossRef]
- Roncevic, Z.; Bajic, B.; Dodic, S.; Grahovac, J.; Pajovic-Scepanovic, R.; Dodic, J. Optimization of bioethanol production from soybean molasses using different strains of Saccharomyces cerevisiae. Hem. Ind. 2019, 73, 1–12. [Google Scholar] [CrossRef]
- Hou, A.; Chen, P.; Shi, A.; Zhang, B.; Wang, Y.J. Sugar variation in soybean seed assessed with a rapid extraction and quantification method. Int. J. Agron. 2009, 2009, 484571. [Google Scholar] [CrossRef]
- Viana, P.A.; de Rezende, S.T.; Falkoski, D.L.; Leite, T.D.; Jose, I.C.; Moreira, M.A.; Guimaraes, V.M. Hydrolysis of oligosaccharides in soybean products by Debaryomyces hansenii UFV-1 α-galactosidases. Food Chem. 2007, 103, 331–337. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Hwang, H. Reduction of soybean oligosaccharides and properties of α-D-galactosidase from Lactobacillus curvatus R08 and Leuconostoc mesenteriodes JK55. Food Microbiol. 2008, 25, 815–823. [Google Scholar] [CrossRef]
- De Fatima Viana, S.; Monteze Guimaraes, V.; Chamel Jose, I.; Goreti de Almeida e Oliveira, M.; Brunoro Costa, N.M.; Goncalves de Barros, E.; Alves Moreira, M.; Tavares de Rezende, S. Hydrolysis of oligosaccharides in soybean flour by soybean α-galactosidase. Food Chem. 2005, 93, 665–670. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, J.Z.; Liu, H.Y.; Yang, G.Q. Soybean oligosaccharide, stachyose, and raffinose in broilers diets: Effects on odor compound concentration and microbiota in cecal digesta. Poult. Sci. 2020, 99, 3532–3539. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Li, C.R.; Shan, Z.X.; Yin, S.J.; Wang, Y.; Wang, C.C.; Liu, T.H.; Wang, N.F.; Guo, Q.B. In vitro fermentability of soybean oligosaccharides from wastewater of Tofu production. Polymers 2022, 14, 1704. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Bai, J.Y.; Wang, B.T.; Yu, L.L.; Tian, F.W.; Zhao, J.X.; Zhang, H.; Suo, H.Y.; Chen, W.; Zhai, Q.X. Stachyose modulates gut microbiota and alleviates DSS-induced ulcerative colitis in mice. Food Sci. Hum. Wellness 2023, 12, 2211–2220. [Google Scholar] [CrossRef]
- Guo, Y.A.; Song, L.Q.; Huang, Y.M.; Li, X.P.; Xiao, Y.C.; Wang, Z.H.; Ren, Z.H. Latilactobacillus sakei Furu2019 and stachyose as probiotics, prebiotics, and synbiotics alleviate constipation in mice. Front. Nutr. 2023, 9, 1039403. [Google Scholar] [CrossRef] [PubMed]
- Teague, K.D.; Tellez-Isaias, G.; Chai, J.M.; Petrone-Garcia, V.; Vuong, C.N.; Blanch, A.; Rasmussen, S.H.; Brown, K.; Zhao, J.C.; Rochell, S.J. Dietary soy galactooligosaccharides affect the performance, intestinal function, and gut microbiota composition of growing chicks. Poult. Sci. 2023, 102, 102440. [Google Scholar] [CrossRef]
- Fan, G.S.; Liu, P.X.; Chang, X.; Yin, H.; Cheng, L.J.; Teng, C.; Gong, Y.; Li, X.T. Isolation and identification of a high-yield ethyl caproate-producing yeast from Daqu and optimization of its fermentation. Front. Microbiol. 2021, 12, 663744. [Google Scholar] [CrossRef]
- Nagata, Y.; Osono, T.; Hasegawa, M.; Hobara, S.; Hirose, D.; Tanabe, Y.; Kudoh, S.; Uchida, M. Application of the Biolog EcoPlateTM technique for assessing the microbial metabolic diversity in moss profiles of continental Antarctica. Polar Sci. 2023, 35, 100924. [Google Scholar] [CrossRef]
- Gizaw, B.; Alemu, T.; Ebsa, G.; Wako, D. Isolation of Hexavalent chromium tolerant fungal species from urban vegetable farm soil and effluent waste in Addis Ababa and Rift valley, Ethiopia. World J. Microbiol. Biotechnol. 2023, 39, 81. [Google Scholar] [CrossRef] [PubMed]
- Krüger-Haker, H.; Ji, X.; Bartel, A.; Fessler, A.T.; Hanke, D.; Jiang, N.S.; Tedin, K.; Maurischat, S.; Wang, Y.; Wu, C.M.; et al. Metabolic characteristics of porcine LA-MRSA CC398 and CC9 isolates from Germany and China via Biolog Phenotype MicroArrayTM. Microorganisms 2022, 10, 2116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Duan, S.L.; He, C.F.; Wang, L.; Song, J.; Wang, X.; Chen, Z.R. Purification of oligosaccharides through fermentation technology with screening of strains. Sci. Technol. Rev. 2011, 29, 24–28. [Google Scholar] [CrossRef]
- Cui, X.Q.; Liu, C.; Dong, Y.M.; He, C.F.; Wang, C.T. Isolation and purification of oligosaccharides from soybean molasses through fermentation. Food Sci. 2009, 30, 343–346. [Google Scholar]
- Liu, Y.L.; Tian, Y.; Bao, D.Q. Preparation of functional soybean oligosaccharide from soybean molasses by fermentation. J. Henan Univ. Technol. 2010, 31, 1–5. [Google Scholar] [CrossRef]
- Liu, F.; Li, X.X.; Ge, H.R.; Qi, X.F.; Yu, D.Y. The preparation of functional oligosaccharide in the process of fermentation from soybean molasses. Food Ind. 2016, 37, 93–98. [Google Scholar]
- Yang, R.; Zhong, X.F.; Tang, Y.B.; Ma, J.H.; Tian, S.B.; Du, Y.H.; Fan, G.S. Screening of microbial strains for the production of functional soybean oligosaccharides from soybean molasses. China Brew. 2024. accepted. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Chapter 7—Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 87–110. [Google Scholar] [CrossRef]
- Fan, G.S.; Sun, B.G.; Xu, D.; Teng, C.; Fu, Z.L.; Du, Y.H.; Li, X.T. Isolation and identification of high-yield ethyl acetate-producing yeast from Gujinggony Daqu and its fermentation characteristic. J. Am. Soc. Brew. Chem. 2018, 76, 117–124. [Google Scholar] [CrossRef]
- Buchana, R.E.; Gibbons, N.E. Bergey’s Manual of Determinative Bacteriology, 8th ed.; Science Press: Beijing, China, 1984; pp. 382–533. [Google Scholar]
- Dong, X.Z.; Cai, M.Y. Ommon Bacteria Manual System Identification; Science Press: Beijing, China, 2001; pp. 348–392. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5’ end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef]
- Zhou, R.L.; Ren, Z.Q.; Ye, J.; Fan, Y.W.; Liu, X.R.; Yang, J.Y.; Deng, Z.Y.; Li, J. Fermented soybean dregs by Neurospora crassa: A traditional prebiotic food. Appl. Biochem. Biotechnol. 2019, 189, 608–625. [Google Scholar] [CrossRef]
- Fu, Z.L.; Sun, B.G.; Li, X.T.; Fan, G.S.; Teng, C.; Alaa, A.; Jia, Y.M. Isolation and characterization of a high ethyl acetate-producing yeast from Laobaigan Daqu and its fermentation conditions for producing high-quality Baijiu. Biotechnol. Biotechnol. Equip. 2018, 32, 1218–1227. [Google Scholar] [CrossRef]
- Francesca, N.; Carvalho, C.; Almeida, P.M.; Sannino, C.; Settanni, L.; Sampaio, J.P.; Moschetti, G. Wickerhamomyces sylviae f.a., sp. nov., an ascomycetous yeast species isolated from migratory birds. Int. J. Syst. Evol. Microbiol. 2013, 63, 4824–4830. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.S.; Cheng, L.J.; Fu, Z.L.; Sun, B.G.; Teng, C.; Jiang, X.Y.; Li, X.T. Screening of yeasts isolated from Baijiu environments for 2-phenylethanol production and optimization of production conditions. 3 Biotech 2020, 10, 275. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.S.; Liu, P.X.; Wu, Q.H.; Fu, Z.L.; Cheng, L.J.; Zhu, Y.T.; Zhu, Y.P.; Yang, R.; Li, X.T. Optimization of cultural conditions for ethyl alcohol production by Saccharomyces cerevisiae YF1914 in aerobic conditions and its aroma-producing characteristics. Sci. Technol. Food Ind. 2019, 40, 52–58. [Google Scholar] [CrossRef]
- Zhang, J.G. Biological Purification and Transformation of Functional Components in Soybean Molasses. Master’ Thesis, Tianjin University of Science and Technology, Tianjin, China, 2019. [Google Scholar] [CrossRef]
- Cheng, R.W.; Wang, F.Q.; Xu, Y.R.; Wei, L.; Ma, J.H.; Gao, P.; Yang, R.; Liu, X.Y.; Fan, G.S. Optimization of submerged fermentation conditions for glucanase production by Burkholderia pyrrocinia B1213 using Jiuzao. Emir. J. Food Agric. 2023, 35, 468–480. [Google Scholar] [CrossRef]
- Fang, W.H. Isolation of the Soybean Molasses and the Microbiological Purification of the Oligosaccharide. Master’s Thesis, Jiangnan University, Wuxi, China, 2004. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Level |
|---|---|
| Inoculum size (%, v/v) | 0.05, 0.125, 0.25, 0.375, 0.5, 1.0, 2.0, 4.0, 6.0, 8.0, and 10.0 |
| Temperature (°C) | 15, 18, 21, 24, 27, 30, 33, 36, and 39 |
| Shaking speed (rpm) | 0, 50, 100, 150, 200, 250, and 300 |
| Dilution ratio (folds) | 2, 4, 6, 8, 10, 12, 14, and 16 |
| pH | 3, 4, 5, 6, 7, 8, 9, and 10 |
| Time (h) | 2, 4, 6, 8, 10, 12, 14, and 16 |
| Experiment Group | Inoculum Size (A) (%, v/v) | Temperature (B) (°C) | Shaking Speed (C) (rpm) | Dilution Ratio (D) (Folds) | pH (E) | Time (F) (h) | Blank |
|---|---|---|---|---|---|---|---|
| 1 | 0.125 (1) | 27 (1) | 50 (1) | 6 (1) | 5 (1) | 10 (1) | 1 |
| 2 | 0.125 | 30 (2) | 100 (2) | 8 (2) | 6 (2) | 12 (2) | 2 |
| 3 | 0.125 | 33 (3) | 150 (3) | 10 (3) | 7 (3) | 14 (3) | 3 |
| 4 | 0.25 (2) | 27 | 50 | 8 | 6 | 14 | 3 |
| 5 | 0.25 | 30 | 100 | 10 | 7 | 10 | 1 |
| 6 | 0.25 | 33 | 150 | 6 | 5 | 12 | 2 |
| 7 | 0.375 (3) | 27 | 100 | 6 | 7 | 12 | 3 |
| 8 | 0.375 | 30 | 150 | 8 | 5 | 14 | 1 |
| 9 | 0.375 | 33 | 50 | 10 | 6 | 10 | 2 |
| 10 | 0.125 | 27 | 150 | 10 | 6 | 12 | 1 |
| 11 | 0.125 | 30 | 50 | 6 | 7 | 14 | 2 |
| 12 | 0.125 | 33 | 100 | 8 | 5 | 10 | 3 |
| 13 | 0.25 | 27 | 100 | 10 | 5 | 14 | 2 |
| 14 | 0.25 | 30 | 150 | 6 | 6 | 10 | 3 |
| 15 | 0.25 | 33 | 50 | 8 | 7 | 12 | 1 |
| 16 | 0.375 | 27 | 150 | 8 | 7 | 10 | 2 |
| 17 | 0.375 | 30 | 50 | 10 | 5 | 12 | 3 |
| 18 | 0.375 | 33 | 100 | 6 | 6 | 14 | 1 |
| Sugar Fermentation Test | Carbon Source Assimilation Test | Nitrogen Source Assimilation Test | Others Test | ||||
|---|---|---|---|---|---|---|---|
| Sugar | Characteristics | Carbon Source | Characteristics | Nitrogen Source | Characteristics | Test | Result |
| Saccharose | Acid and gas production; growth | Soluble starch | + | Urea | + | Hydrogen sulfide test | − |
| Maltose | Not all produce acid; gas production; growth | Ethyl alcohol | + | Potassium nitrate | + | Indole test | + |
| Xylose | No acid and gas; growth | Mannitol | + | Potassium nitrite | + | Urea test | − |
| Lactose | No acid and gas; growth | Citric acid | + | L-Phenylalanine | + | Methyl red test | + |
| Galactose | Not all produce acid; gas production; growth | Rhamnose | + | Ammonium sulfate | + | Voges–Proskauer test | − |
| Arabinose | No acid and gas; growth | Trehalose | + | Gelatin liquefication test | − | ||
| Sorbinose | No acid and gas; growth | Formic acid | − | Citrate test | + | ||
| Glucose | Not all produce acid; gas production; growth | Glycerol | + | Starch hydrolysis test | − | ||
| Fructose | + | Litmus milk test | + | ||||
| Experiment Group | Sucrose | Raffinose | Stachyose | Manninotriose | K | ||||
|---|---|---|---|---|---|---|---|---|---|
| Retention Rate (%) | Range ω | Retention Rate (%) | Range τ | Retention Rate (%) | Range φ | Content (mg/mL) | Range σ | ||
| 1 | 85.3 | 0.07 | 85.0 | 0.86 | 91.0 | 0.91 | 2.7 | 0.99 | 0.49 |
| 2 | 63.7 | 0.30 | 83.4 | 0.83 | 95.0 | 0.97 | 2.2 | 0.99 | 0.62 |
| 3 | 0.2 | 1.00 | 38.8 | 0.34 | 80.1 | 0.75 | 54.6 | 0.32 | 0.83 |
| 4 | 45.8 | 0.50 | 80.9 | 0.80 | 71.0 | 0.61 | 37.1 | 0.55 | 0.57 |
| 5 | 45.6 | 0.50 | 66.8 | 0.65 | 96.2 | 0.99 | 2.7 | 0.99 | 0.71 |
| 6 | 18.7 | 0.80 | 33.8 | 0.26 | 84.9 | 0.82 | 3.2 | 0.98 | 0.76 |
| 7 | 23.5 | 0.74 | 32.6 | 0.26 | 89.2 | 0.89 | 1.6 | 1.00 | 0.75 |
| 8 | 0.0 | 1.00 | 11.9 | 0.02 | 52.2 | 0.33 | 29.9 | 0.64 | 0.64 |
| 9 | 46.5 | 0.49 | 33.4 | 0.26 | 95.1 | 0.97 | 2.7 | 0.99 | 0.66 |
| 10 | 30.8 | 0.66 | 75.8 | 0.73 | 96.8 | 1.00 | 2.7 | 0.99 | 0.80 |
| 11 | 23.3 | 0.75 | 86.2 | 0.87 | 92.0 | 0.93 | 1.6 | 1.00 | 0.84 |
| 12 | 91.5 | 0.00 | 40.1 | 0.34 | 90.6 | 0.91 | 2.2 | 0.99 | 0.40 |
| 13 | 0.0 | 1.00 | 35.0 | 0.28 | 29.8 | 0.00 | 79.8 | 0.00 | 0.53 |
| 14 | 71.4 | 0.22 | 60.8 | 0.57 | 81.8 | 0.78 | 8.3 | 0.91 | 0.48 |
| 15 | 41.2 | 0.55 | 96.6 | 0.99 | 81.4 | 0.77 | 11.6 | 0.87 | 0.68 |
| 16 | 29.7 | 0.68 | 54.3 | 0.51 | 75.9 | 0.69 | 12.1 | 0.87 | 0.67 |
| 17 | 4.4 | 0.95 | 49.5 | 0.45 | 73.8 | 0.66 | 28.4 | 0.66 | 0.78 |
| 18 | 10.3 | 0.89 | 73.0 | 0.72 | 65.9 | 0.54 | 11.5 | 0.87 | 0.74 |
| Dependent Variable VAR00001 | Tests of Between-Subject Effects | ||||
|---|---|---|---|---|---|
| III Sum of Squares | Degree of Freedom | Mean Square | F | Significance | |
| Corrected model | 0.248 | 12 | 0.021 | 3.255 | 0.101 |
| Intercept | 7.924 | 1 | 7.924 | 1249.081 | 0.000 |
| Inoculum size (A) | 0.021 | 2 | 0.011 | 1.691 | 0.275 |
| Temperature (B) | 0.007 | 2 | 0.004 | 0.563 | 0.602 |
| Shaking speed (C) | 0.016 | 2 | 0.008 | 1.222 | 0.370 |
| Dilution ratio (D) | 0.046 | 2 | 0.023 | 3.607 | 0.107 |
| pH (E) | 0.069 | 2 | 0.035 | 5.443 | 0.056 |
| Time (F) | 0.089 | 2 | 0.044 | 7.008 | 0.035 * |
| Error | 0.032 | 5 | 0.006 | ||
| Total | 8.204 | 18 | |||
| Corrected total | 0.280 | 17 | |||
| R2 = 0.887 (Adjusted R2 = 0.614) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Ma, J.; Wang, Z.; Du, Y.; Tian, S.; Fan, G.; Liu, X.; Teng, C. The Identification of a Strain for the Biological Purification of Soy Molasses to Produce Functional Soy Oligosaccharides and Optimize Purification Conditions. Foods 2024, 13, 296. https://doi.org/10.3390/foods13020296
Yang R, Ma J, Wang Z, Du Y, Tian S, Fan G, Liu X, Teng C. The Identification of a Strain for the Biological Purification of Soy Molasses to Produce Functional Soy Oligosaccharides and Optimize Purification Conditions. Foods. 2024; 13(2):296. https://doi.org/10.3390/foods13020296
Chicago/Turabian StyleYang, Ran, Jinghao Ma, Zechen Wang, Yihua Du, Shubin Tian, Guangsen Fan, Xiaoyan Liu, and Chao Teng. 2024. "The Identification of a Strain for the Biological Purification of Soy Molasses to Produce Functional Soy Oligosaccharides and Optimize Purification Conditions" Foods 13, no. 2: 296. https://doi.org/10.3390/foods13020296