Nutrient Properties and Nuclear Magnetic Resonance-Based Metabonomic Analysis of Macrofungi

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivated Edible and Medical Mushrooms
2.2. Proximate Compositions and Trace Minerals
2.3. Coarse Fibers
2.4. Vitamin A and C
2.5. Amino Acids
2.6. Sample Preparation and 1H NMR Spectroscopic Measurement
2.7. Statistical Analysis
3. Results and Discussion
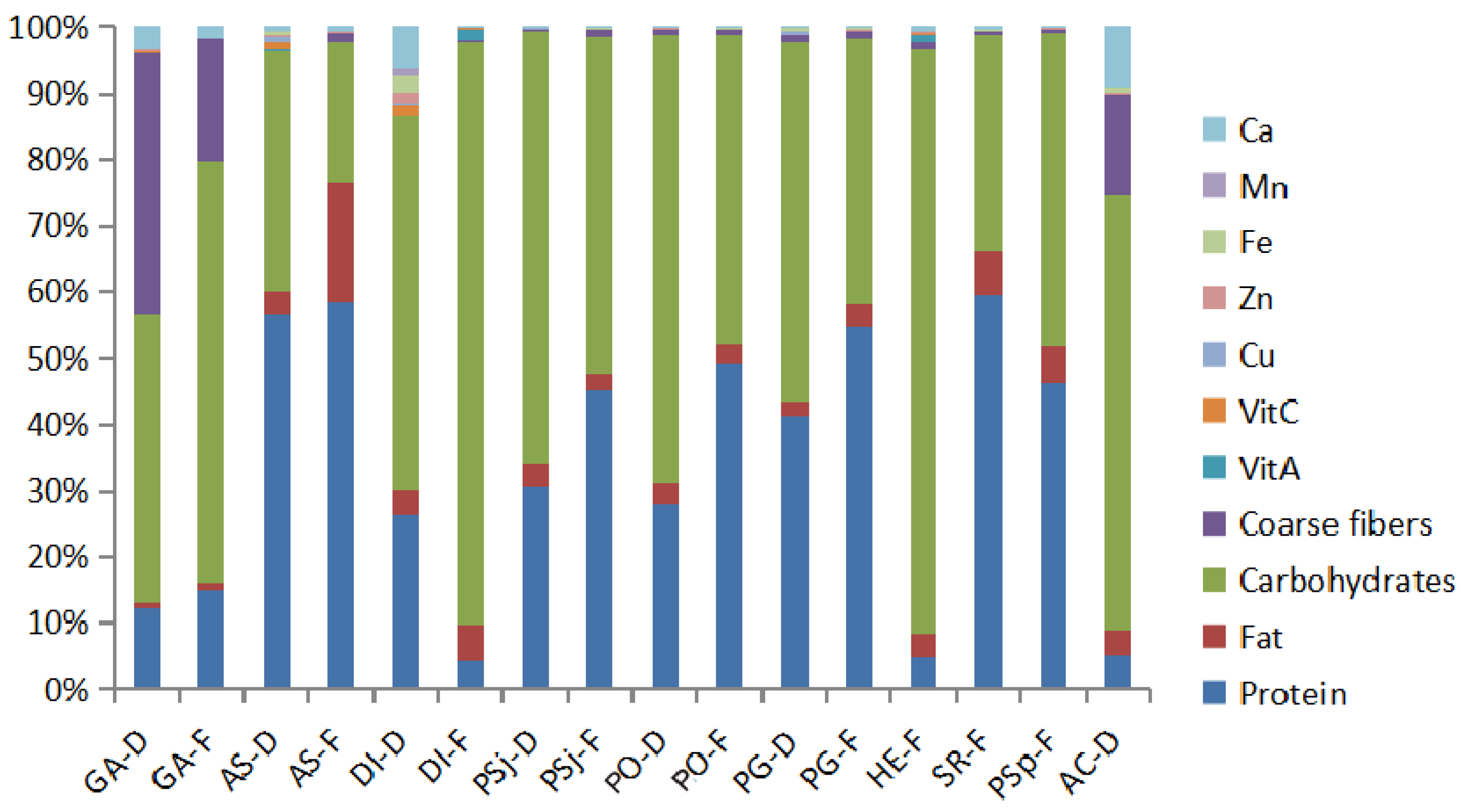
3.1. Proximate Compositions and Trace Elements from Dried and Fresh Mushrooms
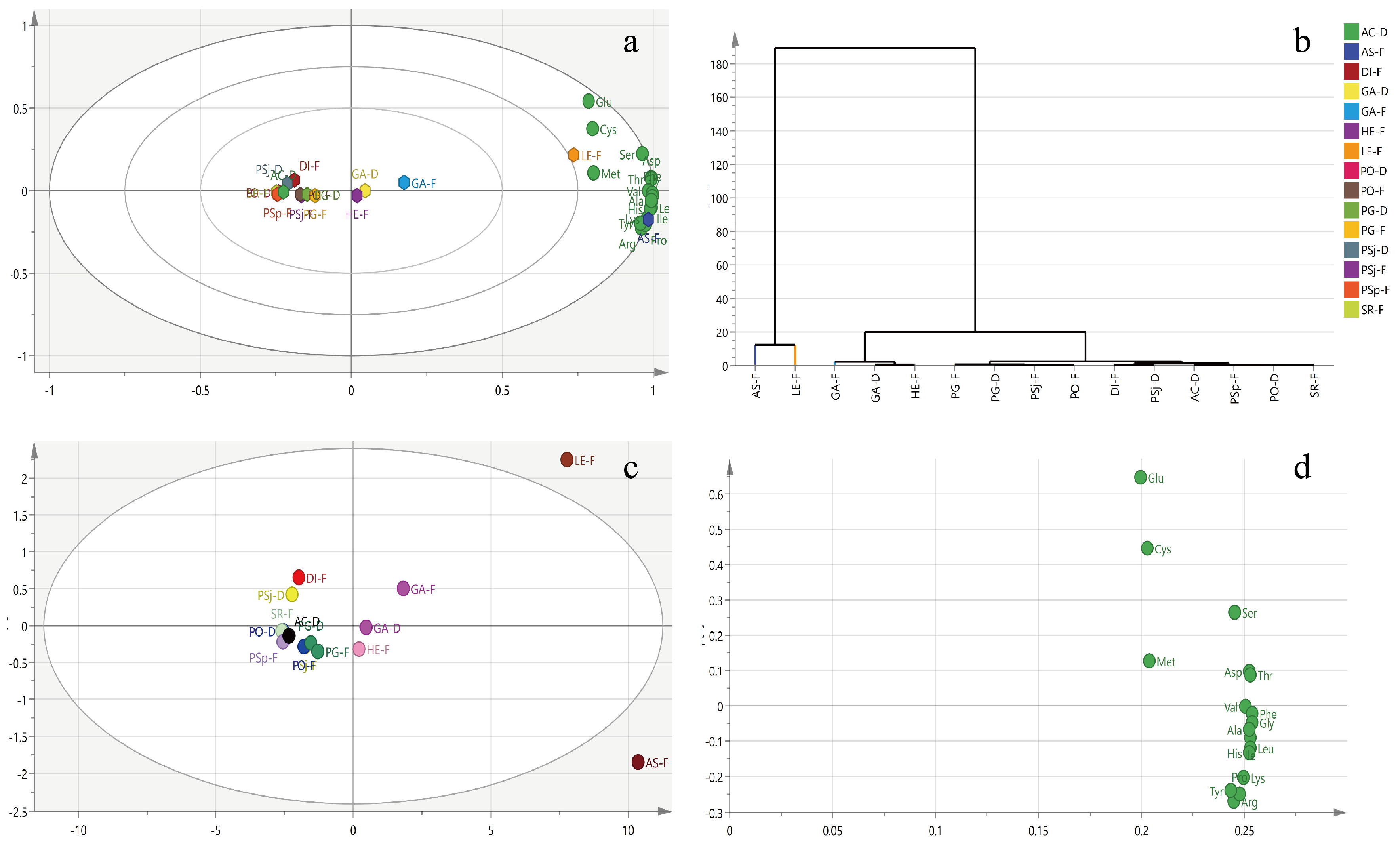
3.2. Amino Acid Derivatives and Peptides
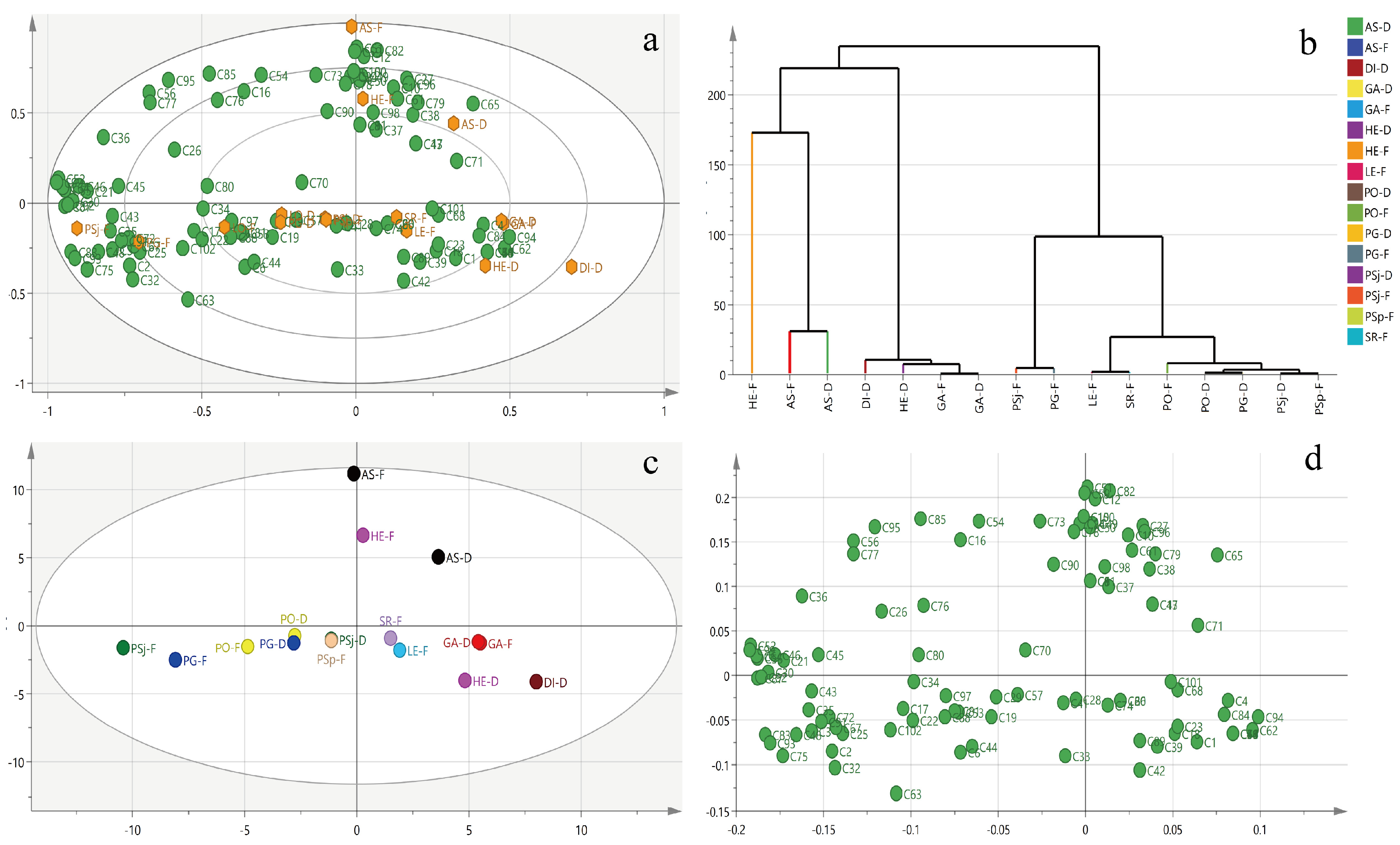
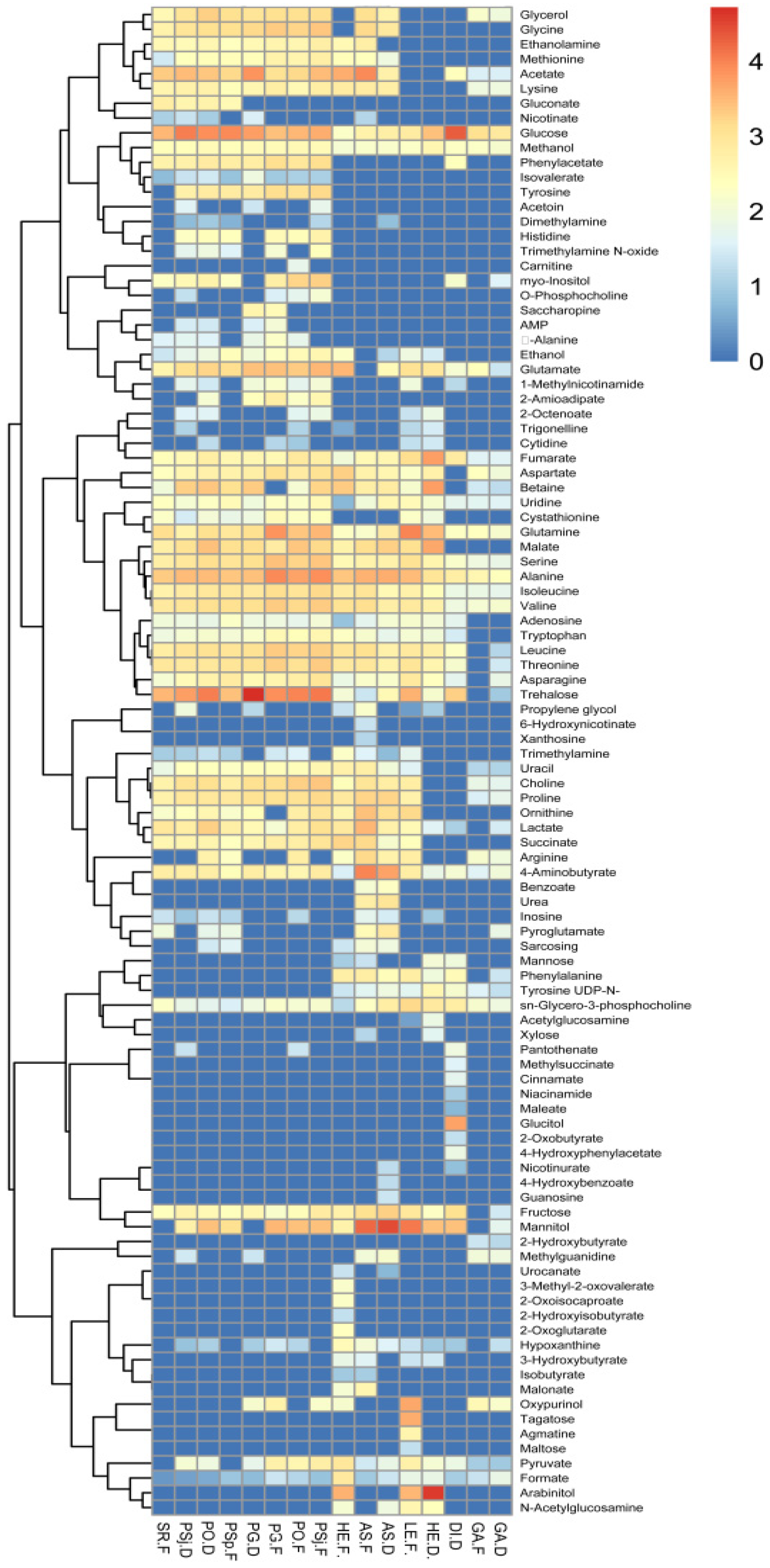
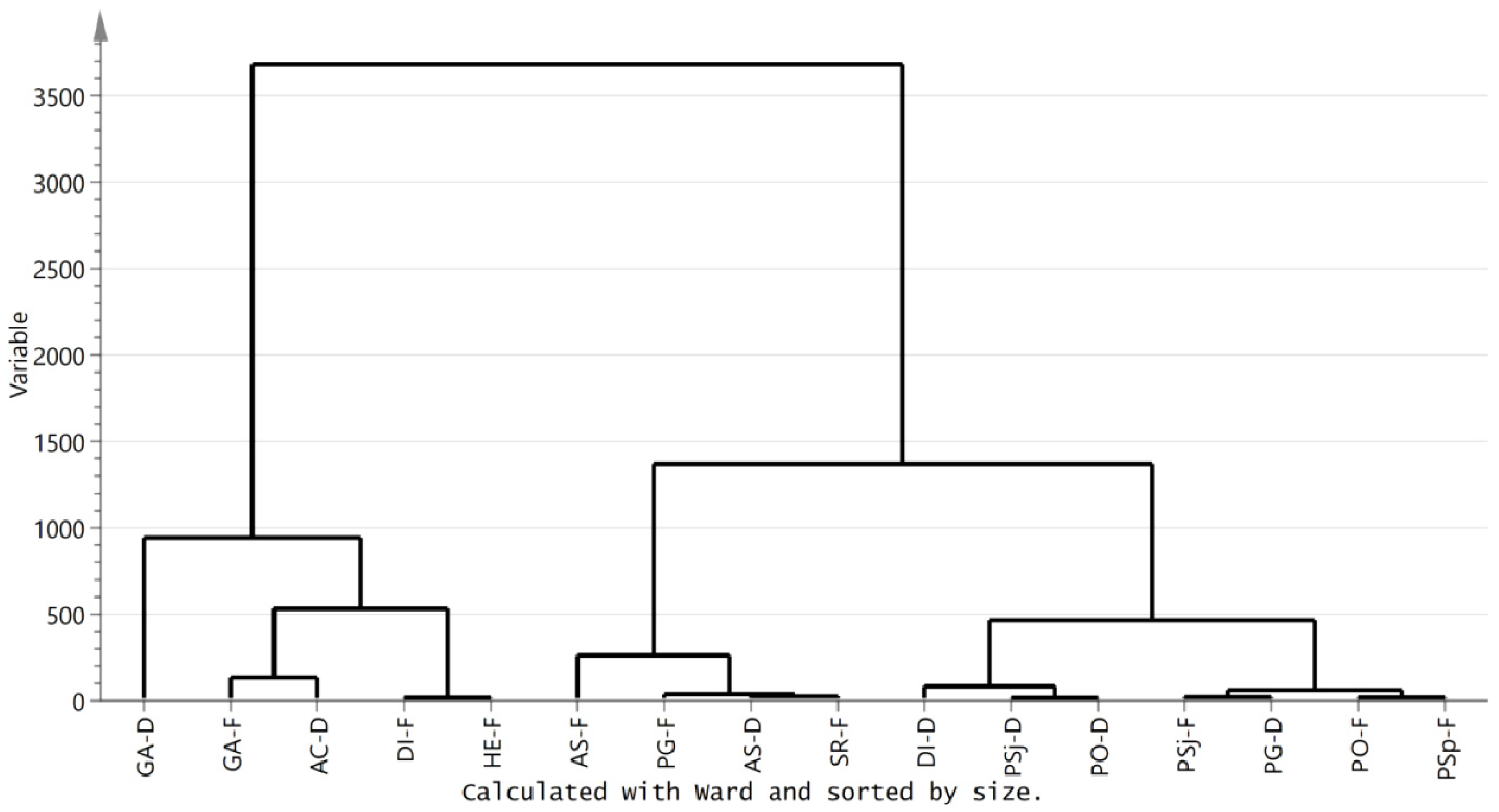
3.3. Metabolic Profiling of Selected Mushrooms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wu, S.R.; Luo, X.L.; Liu, B.; Gui, M.Y. Analyse and advise to research and development of wild edible fungi. Food Sci. Technol. 2010, 35, 100–103. [Google Scholar]
- Zhao, C.; Fan, J.; Liu, Y.; Guo, W.; Cao, H.; Xiao, J.; Wang, Y.; Liu, B. Hepatoprotective activity of Ganoderma lucidum triterpenoids in alcohol-induced liver injury in mice, an iTRAQ-based proteomic analysis. Food Chem. 2019, 271, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Q.; Liu, D.; Wang, D.Y.; Lai, S.S.; Zhong, R.T.; Liu, Y.Y.; Yang, C.F.; Liu, B.; Sarker, M.M.R.; Zhao, C. Hypoglycemic activity and gut microbiota regulation of a novel polysaccharide from Grifola frondosa in type 2 diabetic mice. Food Chem. Toxicol. 2019, 126, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, C.F.; Wai, S.T.C.; Zhang, Y.B.; Portillo, M.Y.; Paoli, P.; Wu, Y.J.; Cheang, W.S.; Liu, B.; Carpéné, C.; et al. Regulation of glucose metabolism by bioactive phytochemicals for the management of type 2 diabetes mellitus. Crit. Rev. Food Sci. Nutr. 2019, 59, 830–847. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Fu, H.; Teng, L.; Hu, Z.; Xu, X.; Chen, J. Anti-tumor activity of the regenerated triple-helical polysaccharide from dictyophora indusiata. Int. J. Biol. Macromol. 2013, 61, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Yang, B.; Tang, J.; Ma, Z.; Gao, Q.; Zhao, M. Structural analysis of water-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their in vivo antioxidant activities. Carbohydr. Polym. 2012, 87, 343–347. [Google Scholar] [CrossRef]
- Kanagasabapathy, G.; Kuppusamy, U.R.; Malek, S.N.A.; Abdulla, M.A.; Chua, K.H.; Sabaratnam, V. Glucan-rich polysaccharides from Pleurotus sajor-caju(fr.) singer prevents glucose intolerance, insulin resistance and inflammation in c57bl/6j mice fed a high-fat diet. BMC Complementary Altern. Med. 2012, 12, 261. [Google Scholar] [CrossRef]
- Cao, X.Y.; Liu, J.L.; Yang, W.; Hou, X.; Li, Q.J. Antitumor activity of polysaccharide extracted from Pleurotus ostreatus mycelia against gastric cancer in vitro and in vivo. Mol. Med. Rep. 2015, 12, 2383. [Google Scholar] [CrossRef]
- Chang, U.M.; Li, C.H.; Lin, L.I.; Huang, C.P.; Kan, L.S.; Lin, S.B. Ganoderiol F, a Ganoderma triterpene, induces senescence in hepatoma HepG2 cells. Life Sci. 2006, 79, 1129–1139. [Google Scholar] [CrossRef]
- Phan, C.W.; David, P.; Naidu, M.; Wong, K.H.; Sabaratnam, V. Therapeutic potential of culinary-medicinal mushrooms for the management of neurodegenerative diseases: Diversity, metabolite, and mechanism. Crit. Rev. Biotechnol. 2015, 35, 355–368. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Liu, Y.Y.; Sarker, M.M.; Yan, X.; Yang, C.F.; Zhao, L.N.; Lv, X.C.; Liu, B.; Zhao, C. Structural characterization and antidiabetic potential of a novel heteropolysaccharide from Grifola frondosa via IRS1/PI3K-JNK signaling pathways. Carbohydr. Polym. 2018, 198, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Gao, L.; Wang, C.; Liu, B.; Jin, Y.; Xing, Z. Structural characterization and antiviral activity of a novel heteropolysaccharide isolated from Grifola frondosa against enterovirus 71. Carbohydr. Polym. 2016, 144, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Komura, D.L.; Carbonero, E.R.; Gracher, A.H.P.; Baggio, C.H.; Freitas, C.S.; Marcon, R.; Santos, A.R.S.; Gorin, P.A.J.; Iacomini, M. Structure of Agaricus spp. fucogalactans and their anti-inflammatory and antinociceptive properties. Bioresour. Technol. 2010, 101, 6192–6199. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.C.; Jeong, Y.T.; Yang, B.K.; Islam, R.; Koyyalamudi, S.R.; Pang, G.; Cho, K.Y.; Song, C.H. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetic and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.; Bhunia, S.K.; Maity, K.K.; Patra, S.; Mandal, S.; Maiti, S.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. Glucans of Pleurotus florida blue variant: Isolation, purification, characterization and immunological studies. Int. J. Biol. Macromol. 2012, 50, 591–597. [Google Scholar] [CrossRef]
- Barros, L.; Cruz, T.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C. Wild and commercial mushrooms as source of nutrients and nutraceuticals. Food Chem. Toxicol. 2007, 46, 2742–2747. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, Y.; Lai, S.; Cao, H.; Guan, Y.; San Cheang, W.; Liu, B.; Zhao, K.; Miao, S.; Riviere, C.; et al. Effects of domestic cooking process on the chemical and biological properties of dietary phytochemicals. Trends Food Sci. Technol. 2019, 85, 55–66. [Google Scholar] [CrossRef]
- Pereira, E.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Towards chemical and nutritional inventory of Portuguese wild edible mushrooms in different habitats. Food Chem. 2012, 130, 394–403. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhang, J.; Wu, L.H.; Zhao, Y.L.; Li, T.; Li, J.Q.; Wang, Y.Z.; Liu, H.G. A mini-review of chemical composition and nutritional value of edible wild-grown mushroom from China. Food Chem. 2014, 151, 279–285. [Google Scholar] [CrossRef]
- Vaz, J.A.; Barros, L.; Martins, A.; Santos-Buelga, C.; Vasconcelos, M.H.; Ferreira, I.C.F.R. Chemical composition of wild edible mushrooms and antioxidant properties of their water soluble polysaccharidic and ethanolic fractions. Food Chem. 2011, 126, 610–616. [Google Scholar] [CrossRef]
- Minyoung, K.; Lllmin, C.; Sunjoo, L.; Joungkuk, A.; Eunhye, K.; Mijung, K.; Sunlim, K.; Hyungin, M.; Heemyong, R.; Kang, E.Y. Comparison of free amino acid, carbohydrates concentrations in Korean edible and medicinal mushrooms. Food Chem. 2009, 113, 386–393. [Google Scholar]
- Pck, C. The nutritional and health benefits of mushrooms. Nutr. Bull. 2010, 35, 292–299. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 16th ed.; Cunniff, P.A., Ed.; AOAC International: Arlington, VA, USA, 1995; Volume 1. [Google Scholar]
- Allen, W.F. A micro-Kjeldahl method for nitrogen determination. Oil Fat Ind. 1931, 10, 391–397. [Google Scholar] [CrossRef]
- Brzostowski, A.; Falandysz, J.; Jarzyńska, G.; Zhang, D. Bioconcentration potential of metallic elements by Poison Pax (Paxillus involutus) mushroom. J. Environ. Sci. Health Part A 2011, 46, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Brzostowski, A.; Jarzyńska, G.; Kojta, A.K.; Wydmańska, D.; Falandysz, J. Variations in metal levels accumulated in Poison Pax (Paxillus involutus) mushroom collected at one site over four years. J. Environ. Sci. Health Part A 2011, 46, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Morán, J.I.; Alvarez, V.A.; Cyras, V.P.; Vázquez, A. Extraction of cellulose and preparation of nanocellulose from sisal fibers. Cellulose 2008, 15, 149–159. [Google Scholar] [CrossRef]
- Leal, A.R.; Barros, L.; Barreira, J.C.M.; Sousa, M.J.; Martins, A.; Santos-Buelga, C.; Ferreira, I.C.F.R. Portuguese wild mushrooms at the “pharma–nutrition” interface: Nutritional characterization and antioxidant properties. Food Res. Int. 2013, 50, 1–9. [Google Scholar] [CrossRef]
- Grangeia, C.; Heleno, S.A.; Barros, L.; Martins, A.; Ferreira, I.C.F.R. Effects of trophism on nutritional and nutraceutical potential of wild edible mushrooms. Food Res. Int. 2011, 44, 1029–1035. [Google Scholar] [CrossRef]
- Barros, L.; Venturini, B.A.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C.F.R. Chemical composition and biological properties of portuguese wild mushrooms: A comprehensive study. J. Agric. Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef]
- Liu, P.H.; Yuan, J.; Jiang, Z.H.; Wang, Y.X.; Weng, B.Q.; Li, G.X. A lower cadmium accumulating strain of Agaricus brasiliensis produced by 60Co-γ-irradiation. LWT Food Sci. Technol. 2019, 114, 108370. [Google Scholar] [CrossRef]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
- Ouzouni, P.K.; Petridis, D.; Koller, W.D.; Riganakos, K.A. Nutritional value and metal content of wild edible mushrooms collected from West Macedonia and Epirus, Greece. Food Chem. 2009, 115, 1575–1580. [Google Scholar] [CrossRef]
- Jiang, Z.H. An analysis of amino acids in A. blazei fruitbodies cultured by three different composts. Acta Edulis Fungi 1999, 6, 30–34. (In Chinese) [Google Scholar]
- Li, S.; Wang, A.; Liu, L.; Tian, G.; Wei, S.; Xu, F. Evaluation of nutritional values of shiitake mushroom (Lentinus edodes) stipes. J. Food Meas. Charact. 2018, 12, 2012–2019. [Google Scholar] [CrossRef]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Micrbiol. 2013, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Lou, Y.R.; Tzi, V.; Jander, G. Alteration of plant primary metabolism in response to insect herbivory. Plant Physiol. 2015, 169, 1488–1498. [Google Scholar] [CrossRef]
- Beluhan, S.; Ranogajec, A. Chemical composition and non-volatile components of Croatian wild edible mushrooms. Food Chem. 2011, 124, 1076–1082. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, Y.J.; Liu, X.Y.; Liu, B.; Cao, H.; Yu, H.; Sarker, S.D.; Nahar, L.; Xiao, J.B. Functional properties, structural studies and chemo-enzymatic synthesis of oligosaccharides. Trends Food Sci. Technol. 2017, 66, 135–145. [Google Scholar] [CrossRef]
- Yang, C.F.; Lai, S.S.; Chen, Y.H.; Liu, D.; Liu, B.; Ai, C.; Wan, X.Z.; Gao, L.Y.; Chen, X.H.; Zhao, C. Anti-diabetic effect of oligosaccharides from seaweed Sargassum confusum via JNK-IRS1/PI3K signalling pathways and regulation of gut microbiota. Food Chem. Toxicol. 2019, 131, 110562. [Google Scholar] [CrossRef]
- Maga, J.A. Mushroom flavor. J. Agric. Food Chem. 1981, 29, 1–4. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Tsai, H.L.; Mau, J.L. Non-volatile taste components of Agaricus blazei, Agrocybe cylindracea and Boletus edulis. Food Chem. 2008, 107, 977–983. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macrofungi | Proximate Composition | Trace Elements (mg/kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein (g/100 g) | Fat (g/100 g) | Carbohydrates (g/100 g) | Coarse Fibers (g/100 g) | Vitamin A (μg/100 g) | Vitamin C (mg/100 g) | Cu | Zn | Fe | Mn | Ca | |
| GA-D | 12.7 | 0.8 | 45 | 40.8 | 15.2 | 10.5 | 4.7 | 19.2 | 13.4 | 11 | 321 |
| GA-F | 14.6 | 1 | 61 | 17.7 | 17.8 | 1.6 | 2.2 | 5.9 | 3.9 | 2.2 | 172 |
| AS-D | 37.2 | 2.2 | 23.9 | 0.075 | <16 | 69.7 | 42.6 | 28 | 28.4 | 1.1 | 50.2 |
| AS-F | 50.4 | 15.7 | 18 | 1.2 | 7.38 | 1.1 | 14.6 | 13.5 | 14.4 | 0.9 | 53.8 |
| DI-D | 19.5 | 2.7 | 41.6 | 0.087 | 21 | 111.4 | 15 | 130 | 184 | 75 | 468 |
| DI-F | 1.9 | 2.6 | 40.2 | 0.085 | 770 | 16.7 | 0.83 | 0.39 | 1.7 | - | 4.1 |
| PSj-D | 23.2 | 2.7 | 49.4 | 0.4 | 1.46 | 0.6 | 0.6 | 4.8 | 5.4 | 0.7 | 14 |
| PSj-F | 35.5 | 1.98 | 40.1 | 0.8 | 4.05 | 0.6 | 1.9 | 9 | 8.2 | 1.2 | 18 |
| PO-D | 21.1 | 2.19 | 50.7 | 0.7 | 6.51 | 0.7 | 3.6 | 6.8 | 15 | 0.8 | 7 |
| PO-F | 36.4 | 2.06 | 34.7 | 0.5 | 4.42 | 0.5 | 2.5 | 7.7 | 14 | 1 | 11 |
| PG-D | 32.7 | 1.71 | 42.8 | 0.9 | 21.9 | 0.6 | 28 | 15 | 25 | 1.1 | 24 |
| PG-F | 39.8 | 2.26 | 29 | 0.8 | 2.98 | 0.8 | 6.6 | 13 | 15 | 1.4 | 22 |
| HE-F | 2.3 | 1.8 | 42.9 | 0.42 | 580 | 5.5 | 6.1 | 10.4 | 0.15 | 0.079 | 39 |
| SR-F | 41.1 | 4.5 | 22.4 | 0.4 | 1.5 | 1.6 | 2.1 | 3.4 | 13 | 1.4 | 32 |
| PSp-F | 33.8 | 4.1 | 34.6 | 0.5 | 2.61 | 0.5 | 1.2 | 8.4 | 10 | 1.2 | 8.5 |
| AC-D | 4.1 | 3 | 52.8 | 12.2 | - | 0.6 | 3.2 | 27 | 38 | 12 | 740 |
| Amino Acid | GA-D | GA-F | AS-F | DI-F | PSj-D | PSj-F | PO-D | PO-F | PG-D | PG-F | HE-F | SR-F | PSp-F | AC-D | LE-F |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TEAA | 60.5 | 72 | 133.7 | 9.9 | 7.5 | 13.8 | 7.5 | 13.3 | 15.3 | 17.4 | 36.8 | 7.3 | 6.7 | 6 | 107.7 |
| TNAA | 23.5 | 30 | 91.2 | 6.4 | 6.5 | 10.3 | 6 | 10.6 | 12.4 | 14 | 18.1 | 5.4 | 4.3 | 7.4 | 110.1 |
| CEAA | 1.4 | 2.8 | 5.9 | 1.9 | 1.5 | 1.2 | 0.6 | 1.2 | 1.3 | 1.5 | 2.7 | 0.4 | 0.9 | 0.8 | 4.8 |
| Asp | 5.8 | 8.1 | 17.7 | 1.4 | 0.9 | 2.2 | 1.2 | 2.1 | 2.7 | 3.1 | 5.3 | 0.9 | 0.6 | 1.9 | 16.8 |
| Thr | 3.6 | 4.8 | 11 | 0.9 | 0.8 | 1.5 | 0.8 | 1.4 | 1.7 | 1.9 | 3.2 | 0.8 | 0.4 | 1.2 | 10.3 |
| Met | 33.2 | 36.3 | 33.5 | 0.7 | 0.4 | 0.9 | 0.5 | 0.8 | 0.8 | 0.8 | 11.6 | 0.2 | 0.2 | - | 29.7 |
| Ile | 2.7 | 3.4 | 9.8 | 0.7 | 0.7 | 1.3 | 0.6 | 1.2 | 1.5 | 1.6 | 2 | 0.6 | 0.8 | 1.1 | 7.1 |
| Leu | 4.2 | 5.3 | 16.8 | 1.2 | 1.1 | 2 | 1 | 1.9 | 2.1 | 2.6 | 3.8 | 1.3 | 1.4 | 1.6 | 11.7 |
| Val | 3.5 | 4.7 | 11.9 | 2.7 | 1.6 | 2.3 | 1.6 | 2.4 | 2.4 | 2.7 | 2.9 | 1.6 | 1.8 | - | 9.6 |
| Phe | 2.7 | 3.8 | 10.9 | 0.9 | 0.7 | 1 | 0.6 | 1 | 1.3 | 1.3 | 2.3 | 0.6 | 0.8 | 0.9 | 8.5 |
| Lys | 3.5 | 3.9 | 16.6 | 1 | 0.9 | 1.9 | 0.9 | 1.8 | 2 | 2.5 | 4.1 | 0.9 | 0.3 | 0.8 | 10.2 |
| His | 1.3 | 1.7 | 5.5 | 0.4 | 0.4 | 0.7 | 0.3 | 0.7 | 0.8 | 0.9 | 1.6 | 0.4 | 0.4 | 0.4 | 3.8 |
| Ser | 3.9 | 4.7 | 8.8 | 0.9 | 0.6 | 1.4 | 0.8 | 1.3 | 1.5 | 1.7 | 3 | 0.7 | 0.1 | 1.3 | 10.8 |
| Glu | 6.6 | 8.6 | 21.6 | 2.4 | 2.5 | 3.1 | 2.1 | 3.5 | 4.9 | 5.1 | 1.8 | 1.8 | 1.5 | 2.1 | 60.8 |
| Gly | 3.4 | 4.3 | 11.6 | 0.7 | 0.8 | 1.3 | 0.7 | 1.2 | 1.6 | 1.8 | 2.6 | 0.6 | 0.9 | 1.2 | 8.9 |
| Ala | 4 | 5.3 | 16 | 1.1 | 1.4 | 2.7 | 1.3 | 2.5 | 2.8 | 3.1 | 3.1 | 1.1 | 1 | 1.3 | 12.2 |
| Arg | 2.7 | 3.6 | 20.9 | 0.7 | 0.1 | 0.5 | 0.2 | 0.9 | 0.3 | 0.6 | 5.2 | 0.3 | 0.2 | 0.4 | 10.6 |
| Pro | 2.9 | 3.5 | 12.3 | 0.6 | 1.1 | 1.3 | 0.9 | 1.2 | 1.3 | 1.7 | 2.4 | 0.9 | 0.6 | 1.1 | 6.8 |
| Cys | 0.1 | 1 | 1.5 | 1.1 | 0.8 | - | - | - | - | - | 0.7 | - | - | - | 1.9 |
| Tyr | 1.3 | 1.8 | 4.4 | 0.8 | 0.7 | 1.2 | 0.6 | 1.2 | 1.3 | 1.5 | 2 | 0.4 | 0.9 | 0.8 | 2.9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Chen, Y.-Q.; Xiao, X.-W.; Zhong, R.-T.; Yang, C.-F.; Liu, B.; Zhao, C. Nutrient Properties and Nuclear Magnetic Resonance-Based Metabonomic Analysis of Macrofungi. Foods 2019, 8, 397. https://doi.org/10.3390/foods8090397
Liu D, Chen Y-Q, Xiao X-W, Zhong R-T, Yang C-F, Liu B, Zhao C. Nutrient Properties and Nuclear Magnetic Resonance-Based Metabonomic Analysis of Macrofungi. Foods. 2019; 8(9):397. https://doi.org/10.3390/foods8090397
Chicago/Turabian StyleLiu, Dan, Yu-Qing Chen, Xiao-Wei Xiao, Ru-Ting Zhong, Cheng-Feng Yang, Bin Liu, and Chao Zhao. 2019. "Nutrient Properties and Nuclear Magnetic Resonance-Based Metabonomic Analysis of Macrofungi" Foods 8, no. 9: 397. https://doi.org/10.3390/foods8090397
APA StyleLiu, D., Chen, Y.-Q., Xiao, X.-W., Zhong, R.-T., Yang, C.-F., Liu, B., & Zhao, C. (2019). Nutrient Properties and Nuclear Magnetic Resonance-Based Metabonomic Analysis of Macrofungi. Foods, 8(9), 397. https://doi.org/10.3390/foods8090397