Meta-Analysis of NOS3 G894T Polymorphisms with Air Pollution on the Risk of Ischemic Heart Disease Worldwide

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy and Selection Criteria
2.2. Characteristics of Included Studies
2.3. Quality Assessment
2.4. Data Synthesis and Analysis
3. Results
3.1. Pooled Meta-Analysis
3.2. Subgroup Analysis
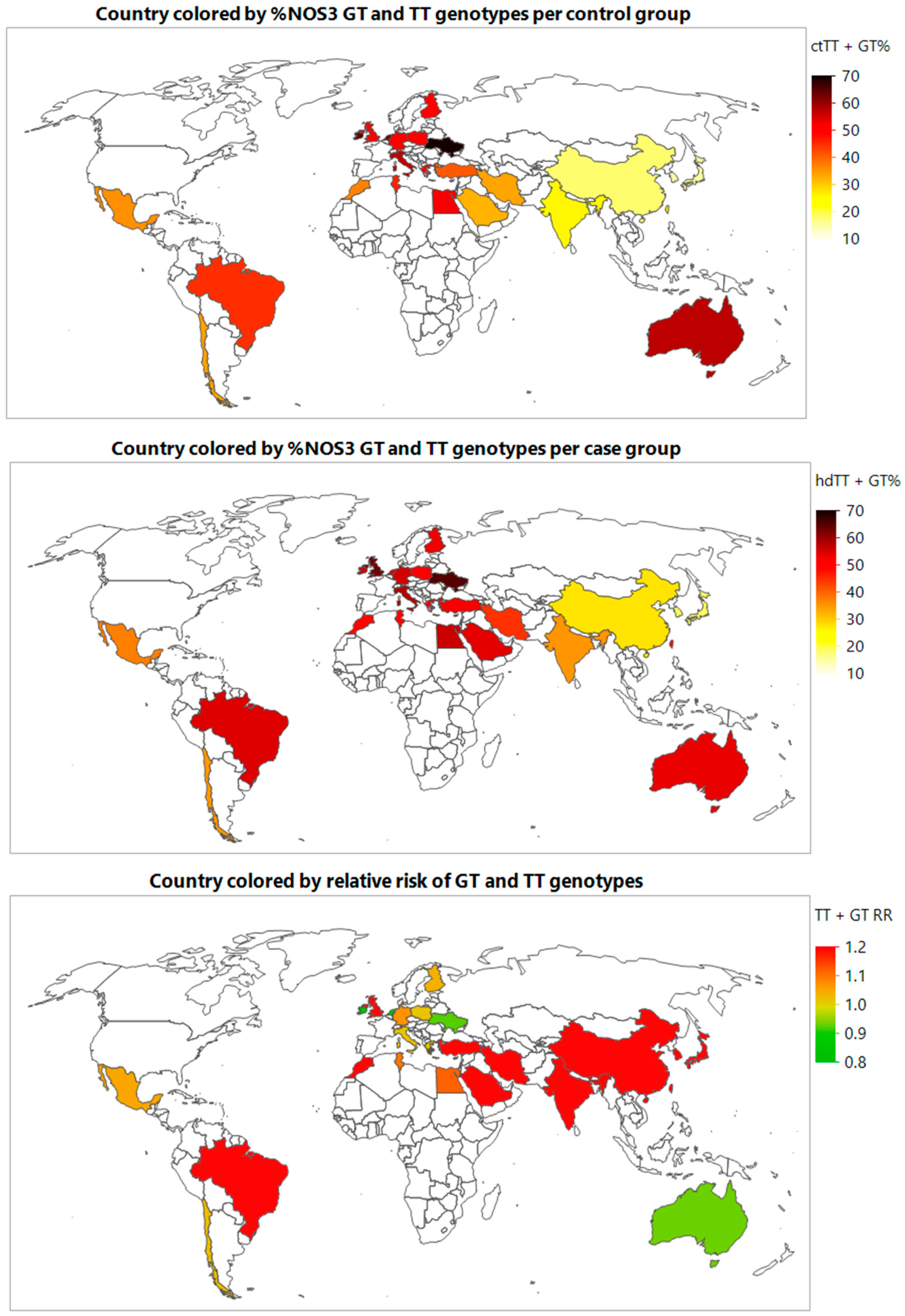
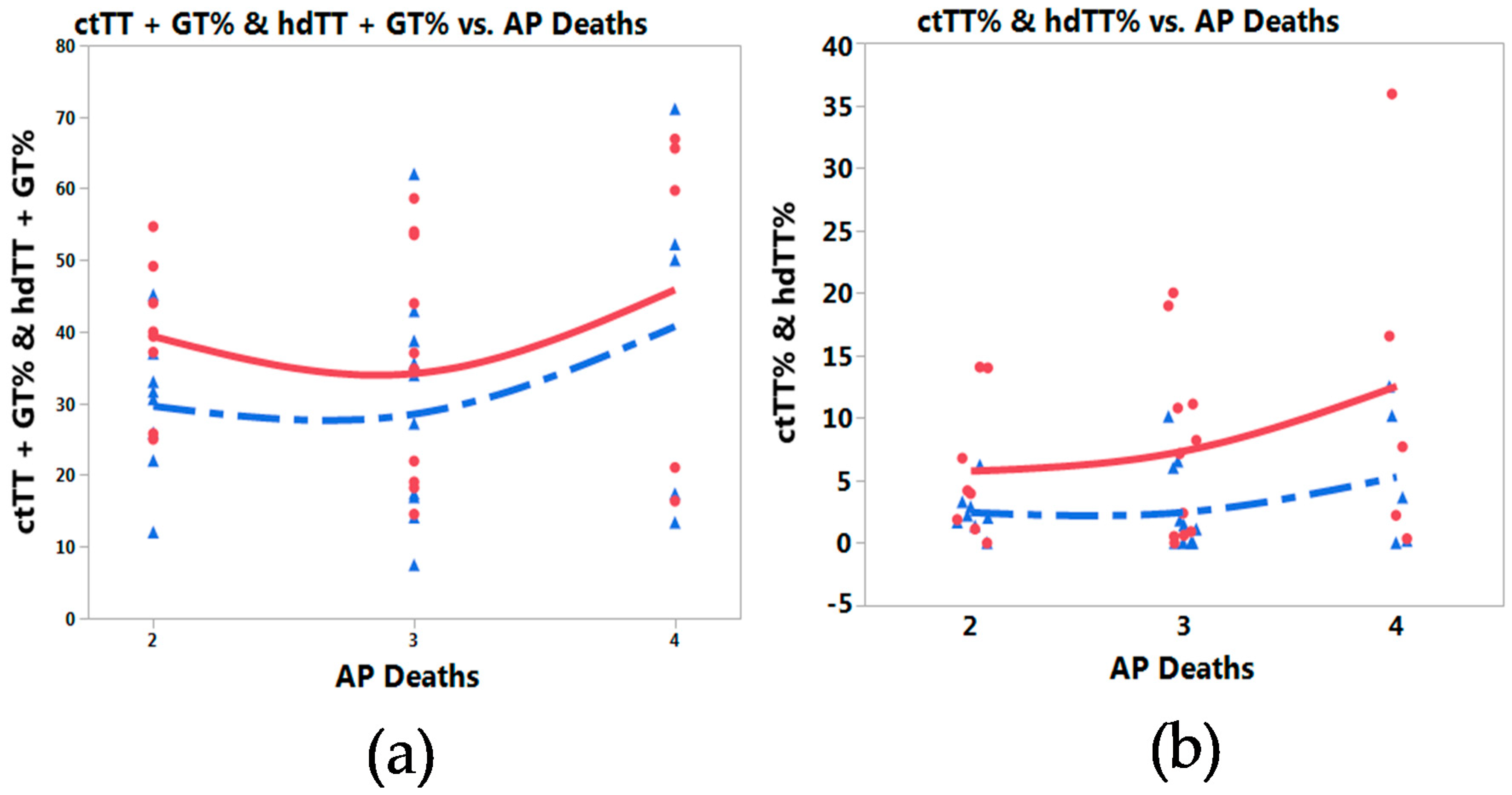
3.3. Meta-Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ferguson, J.F.; Phillips, C.M.; McMonagle, J.; Pérez-Martínez, P.; Shaw, D.I.; Lovegrove, J.A.; Roche, H.M. NOS3 gene polymorphisms are associated with risk markers of cardiovascular disease, and interact with omega-3 polyunsaturated fatty acids. Atherosclerosis 2010, 211, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Jaramillo, P.C.; Munoz, M.A.; Lanas, M.C.; Lanas, Z.F.; Salazar, I.A. Endothelial nitric oxide synthase G894T gene polymorphism in Chilean subjects with coronary artery disease and controls. Clin. Chim. Acta 2006, 371, 102–106. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Cardiovascular Diseases. Available online: http://www.who.int/cardiovascular diseases/en/ (accessed on 1 March 2018).
- Center for Disease Control and Prevention. Heart Disease Facts. Available online: https://www.cdc.gov/heartdisease/facts.htm (accessed on 1 March 2018).
- Heidenreich, P.A.; Trogdon, J.G.; Khavjou, O.A.; Butler, J.; Dracup, K.; Ezekowitz, M.D.; Woo, Y.J. Forecasting the future of cardiovascular disease in the United States: A policy statement from the American Heart Association. Circulation 2011, 123, 933–944. [Google Scholar] [CrossRef] [PubMed]
- American Heart Association. Cardiovascular Disease: A Costly Burden for Americans. Available online: http://www.heart.org/idc/groups/heart-public/@wcm/@adv/documents/downloadable/ucm_491543.pdf (accessed on 1 March 2018).
- Arnett, D.K.; Baird, A.E.; Barkley, R.A.; Basson, C.T.; Boerwinkle, E.; Ganesh, S.K.; O’Donnell, C.J. Relevance of genetics and genomics for prevention and treatment of cardiovascular disease: A scientific statement from the American Heart Association council on epidemiology and prevention, the stroke council, and the functional genomics and translational biology interdisciplinary working group. Circulation 2007, 115, 2878–2901. [Google Scholar] [CrossRef] [PubMed]
- Gluba, A.; Banach, M.; Rysz, J.; Piotrowski, G.; Fendler, W.; Pietrucha, T. Is polymorphism within eNOS gene associated with the late onset of myocardial infarction? A pilot study. Angiology 2009, 60, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.G.; Andreassi, M.G.; Paradossi, U.; Botto, N.; Manfredi, S.; Masetti, S.; Biagini, A. Evidence for association of a common variant of the endothelial nitric oxide synthase gene (Glu298Asp polymorphism) to the presence, extent, and severity of coronary artery disease. Heart 2002, 82, 525–528. [Google Scholar] [CrossRef]
- Gardemann, A.; Lohre, J.; Cayci, S.; Katz, N.; Tillmanns, H.; Haberbosch, W. The T allele of the missense Glu298Asp endothelial nitric oxide synthase gene polymorphism is associated with coronary heart disease in younger individuals with high atherosclerotic risk profile. Atherosclerosis 2002, 160, 167–175. [Google Scholar] [CrossRef]
- Hingorani, A.D.; Liang, C.F.; Fatibene, J.; Lyon, A.; Monteith, S.; Parson, A.; Brown, M.J. A common variant of the endothelial nitric oxide synthase (Glu298–>Asp) is a major risk factor for coronary artery disease in the UK. Circulation 1999, 100, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Rai, H.; Parveen, F.; Kumar, S.; Kapoor, A.; Sinha, N. Association of endothelial nitric oxide synthase gene polymorphisms with coronary artery disease: An updated meta-analysis and systematic review. PLoS ONE 2009, 9, e113363. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A.; Baghdadi, H.; Allam, A.R. Role of genetic changes in the progression of cardiovascular diseases. Int. J. Biomed. Sci. 2011, 7, 238–248. [Google Scholar] [PubMed]
- Gorchakova, O.; Koch, W.; von Beckerath, N.; Mehilli, J.; Schömig, A.; Kastrati, A. Association of a genetic variant of endothelial nitric oxide synthase with the 1 year clinical outcome after coronary stent placement. Eur. Heart J. 2003, 9, 820–827. [Google Scholar] [CrossRef]
- Pereira, T.V.; Rudnicki, M.; Cheung, B.M.Y.; Baum, L.; Yamada, Y.; Oliveira, P.S.L.; Krieger, J.E. Three endothelial nitric oxide (NOS3) gene polymorphisms in hypertensive and normotensive individuals: Meta-analysis of 53 studies reveals evidence of publication bias. J. Hypertens. 2007, 25, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.S.; Mineo, C.; Shaul, P.W.; Bauer, J.A. Biochemical consequences of the NOS3 Glu298Asp variation in human endothelium: Altered caveolar localization and impaired response to shear. FASEB J. 2007, 21, 2655–2663. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, K.; Liu, Z.; Lu, X. Endothelial nitric oxide synthase (eNOS) 4b/a gene polymorphisms and coronary artery disease: Evidence from a meta-analysis. Int. J. Mol. Sci. 2014, 7, 7987–8003. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.Q.; Wen, J.G.; Zhou, H.H.; Chen, X.P.; Zhang, W. Endothelial nitric oxide synthase gene G894T polymorphism and myocardial infarction: A meta-analysis of 34 studies involving 21068 subjects. PLoS ONE 2014, 9, e87196. [Google Scholar] [CrossRef] [PubMed]
- Casas, J.P.; Bautista, L.E.; Humphries, S.E.; Hingorani, A.D. Endothelial nitric oxide synthase genotype and ischemic heart disease: meta-analysis of 26 studies involving 23028 subjects. Circulation 2004, 109, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Yu, L.; Huang, G.R.; Zhang, L.; Liu, Y.Q.; Wang, T.W.; Xiong, H.Y. Polymorphisms of angiotensin converting enzyme and nitric oxide synthase 3 genes risk factors of high altitude pulmonary edema: A case-control study and meta-analysis. Tohoku J. Exp. Med. 2013, 229, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Bai, P.; Shi, S.; Zhou, B.; Wang, Y.; Song, Y.; Rao, L.; Zhang, L. The G894T polymorphism on endothelial nitric oxide synthase gene is associated with increased coronary heart disease among Asia population: Evidence from a Meta analysis. Thromb. Res. 2012, 130, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Xu, X.; Chu, M.; Guo, Y.; Wang, J. Air particulate matter and cardiovascular disease: The epidemiological, biomedical and clinical evidence. J. Thorac. Dis. 2016, 8, E8–E19. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.; Ezzati, M.; Dockery, D.W. Fine-particulate air pollution and life expectancy in the United States. N. Engl. J. Med. 2009, 360, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Zanobetti, A.; Schwartz, J. Particulate air pollution, progression, and survival after myocardial infarction. Environ. Health Perspect. 2007, 115, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Krewski, D. Evaluating the effects of ambient air pollution on life expectancy. N. Engl. J. Med. 2009, 360, 413–415. [Google Scholar] [CrossRef] [PubMed]
- Rich, D.Q.; Schwartz, J.; Mittleman, M.A.; Link, M.; Luttmann-Gibson, H.; Catalano, P.J.; Dockery, D.W. Association of short-term ambient air pollution concentrations and ventricular arrhythmias. Am. J. Epidemiol. 2005, 161, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Dominici, F.; Peng, R.D.; Bell, M.L.; Pham, L.; McDermott, A.; Zeger, S.L.; Samet, J.M. Fine particulate air pollution and hospital admission for cardiovascular and respiratory diseases. JAMA 2006, 295, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Baccarelli, A.; Cassano, P.A.; Litonjua, A.; Park, S.K.; Suh, H.; Sparrow, D.; Vokonas, P.; Schwartz, J. Cardiac autonomic dysfunction: Effects from particulate air pollution and protection by dietary methyl nutrients and metabolic polymorphisms. Circulation 2008, 117, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Raaschou-Nielsen, O.; Andersen, Z.J.; Jensen, S.S.; Ketzel, M.; Sorensen, M.; Hansen, J.; Overvad, K. Traffic air pollution and mortality from cardiovascular disease and all causes: A Danish cohort study. Environ. Health 2012, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.; Yu, C. Meta-prediction of MTHFR gene polymorphism mutations and associated risk for colorectal cancer. Biol. Res. Nurs. 2016, 18, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Chen, Z.; Young, L.; Shiao, P.S. Meta-prediction of the effect of methylenetetrahydrofolate reductase polymorphisms and air pollution on Alzheimer’s disease risk. Int. J. Environ. Res. Public Health 2017, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Nunez, V.; Johns, R.; Shiao, P.S. APOA5 gene polymorphisms and cardiovascular diseases metaprediction in global populations. Nurs. Res. 2017, 66, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, M.C.; Yu, P.; Shiao, S.P.K. MTHFR gene polymorphism-mutations and air pollution as risk factors for breast cancer: A metaprediction study. Nurs. Res. 2017, 66, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Lien, S.-Y.A.; Young, L.; Gau, B.-S.; Shiao, S.P. Meta-prediction of MTHFR gene polymorphism-mutations, air pollution, and risks of leukemia among world populations. Oncotarget 2017, 8, 4387–4398. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Yang, H.L.; Shiao, S.P. Meta-prediction of MTHFR gene polymorphisms and air pollution on the risk of hypertensive disorders in pregnancy worldwide. Int. J. Environ. Res. Public Health 2018, 15, 326. [Google Scholar] [CrossRef] [PubMed]
- Zanobetti, A.; Baccarelli, A.; Schwartz, J. Gene-air pollution interaction and cardiovascular disease: A review. Prog. Cardiovasc. Dis. 2011, 53, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Kim, J.H.; Jung, K.; Hong, Y.C. Associations of air pollution exposure with blood pressure and heart rate variability are modified by oxidative stress genes: A repeated-measures panel among elderly urban residents. Environ. Health 2016, 15, 47. [Google Scholar] [CrossRef] [PubMed]
- Fiordelisi, A.; Piscitelli, P.; Trimarco, B.; Coscion, E.; Iaccarino, G.; Sorriento, D. The mechanisms of air pollution and particulate matter in cardiovascular diseases. Heart Fail. Rev. 2017, 22, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Schneider, A.; Greven, S.; Bellander, T.; Forastiere, F.; Ibald-Mulli, A.; Sunyer, J. Air pollution and inflammatory response in myocardial infarction survivors: Gene-environment interactions in a high-risk group. Inhal. Toxicol. 2007, 19, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Park, S.K.; Vokonas, P.S.; Sparrow, D.; Wilker, E.; Baccarelli, A.; Suh, H.H.; Tucker, K.L.; Wright, R.O.; Schwartz, J. Air pollution and homocysteine: More evidence that oxidative stress-related genes modify effects of particulate air pollution. Epidemiology 2010, 21, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Muto, E.; Hayashi, T.; Yamada, K.; Esaki, T.; Sagai, M.; Iguchi, A. Endothelial-constitutive nitric oxide synthase exists in airways and diesel exhaust particles inhibit the effect of nitric oxide. Life Sci. 1996, 59, 1563–1570. [Google Scholar] [CrossRef]
- Sun, Q.H.; Yue, P.B.; Ying, Z.K.; Cardounel, A.J.; Brook, R.D.; Devlin, R.; Rajagopalan, S. Air pollution exposure potentiates hypertension through reactive oxygen species-mediated activation of Rho/ROCK. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1760–1766. [Google Scholar] [CrossRef] [PubMed]
- Panasevich, S.; Leander, K.; Ljungman, P.; Bellander, T.; de Faire, U.; Pershagen, G.; Nyberg, F. Interaction between air pollution exposure and genes in relation to levels of inflammatory markers and risk of myocardial infarction. BMJ Open 2013, 3, e003058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochette, L.; Lorin, J.; Zeller, M.; Guilland, J.C.; Lorgis, L.; Cottin, Y.; Vergely, C. Nitric oxide synthase inhibition and oxidative stress in cardiovascular diseases: Possible therapeutic targets? Pharmacol. Ther. 2013, 140, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Luiking, Y.C.; Ten Have, G.A.; Wolfe, R.R.; Deutz, N.E. Arginine de novo and nitric oxide production in disease states. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E1177–E1189. [Google Scholar] [CrossRef] [PubMed]
- Wolters, M.; Ströhle, A.; Hahn, A. Age-associated changes in the metabolism of vitamin B12 and folic acid: Prevalence, aetiopathogenesis and pathophysiological consequences. Z. Gerontol. Geriatr. 2004, 37, 109–135. [Google Scholar] [CrossRef] [PubMed]
- Bonnot, O.; Klünemann, H.H.; Sede, F.; Tordjman, S.; Cohen, D.; Walterfang, M. Diagnostic and treatment implications of psychosis secondary to treatable metabolic disorders in adults: A systematic review. Orphanet J. Rare Dis. 2014, 28, 65. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Cook, D.J.; Eastwood, S.; Olkin, I.; Rennie, D.; Stroup, D.F. Improving the quality of reports of meta-analyses of randomized controlled trials: The quorom statement. Onkologie 2000, 23, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Downs, S.H.; Black, N. The feasibility of creating a checklist for the assessment of the methodological quality both of randomized and non-randomized studies of health care interventions. J. Epidemiol. Health 1998, 52, 377–384. [Google Scholar] [CrossRef]
- Stroup, D.F.; Berlin, J.A.; Morton, S.C.; Olkin, I.; Williamson, G.D.; Rennie, D.; Moher, D.; Becker, B.J.; Sipe, T.A.; Thacker, S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. Meta-analysis of observational studies in epidemiology (moose) group. JAMA 2000, 283, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- US Environmental Protection Agency. Air Quality Index. Available online: http://www.airnow.gov/index.cfm?action=aqibasics.aqi (accessed on 8 December 2017).
- Kenworthy, J.; Laube, F. Urban transport patterns in a global sample of cities and their linkages to transport infrastructure, land use, economics and environment. World Transp. Policy Pract. 2002, 8, 5–19. [Google Scholar]
- World Health Organization. Deaths Attributable to Urban Air Pollution. 2004. Available online: http://www.who.int/heli/risks/urban/en/uapmap.1.pdf?ua=1 (accessed on 8 December 2017).
- World Health Organization. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks. 2009. Available online: http://www.who.int/healthinfo/global_burden_disease/GlobalHealthRisks_report_full.pdf (accessed on 8 December 2017).
- World Health Organization. Global Health Risks: Deaths from Air Pollution. 2012. Available online: https://commons.wikimedia.org/wiki/ File:Deaths from_air_pollution.png (accessed on 8 December 2017).
- World Health Organization. The Urban Environment: A General Directory of Resources. 2015. Available online: http://www.who.int/heli/risks/urban/urbenvdirectory/en/ (accessed on 8 December 2017).
- Sha, Q.; Zhang, S. A test of Hardy-Weinberg equilibrium in structured populations. Genet. Epidemiol. 2011, 35, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Wittke-Thompson, J.K.; Pluzhnikov, A.; Cox, N.J. Rational inferences about departures from Hardy-Weinberg equilibrium. Am. J. Hum. Genet. 2005, 76, 967–986. [Google Scholar] [CrossRef] [PubMed]
- How AICR Recommendations Cuts Colorectal Cancer Risk for Both Men and Women. Available online: http://www.aicr.org/cancer-research-update/2016/11_02/cru-how-AICR-recommendations-cuts-colorectal-cancer-risk-for-men-and-women.html (accessed on 16 March 2018).
- WCRF-AICR Continuous Update Project. Diet, Nutrition, Physical Activity and Colorectal Cancer. 2017. Available online: http://www.aicr.org/continuous-update-project/reports/colorectal-cancer-2017-report.pdf (accessed on 1 July 2018).
- Viera, A.J. Odds ratios and risk ratios: What’s the difference and why does it matter? South. Med. J. 2008, 101, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Scott, I. Interpreting risks and ratios in therapy trials. Aust. Prescr. 2008, 31, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Deeks, J.J.; Higgins, J.; Altman, D.G. Analysing Data and Undertaking Meta-Analyses. In Cochrane Handbook for Systematic Reviews of Interventions; Higgins, J., Green, S., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2008; pp. 243–296. [Google Scholar]
- Albrecht, J. Key Concepts and Techniques in GIS; Sage: Thousand Oaks, CA, USA, 2007. [Google Scholar]
- Pereira, C.; Denise, A.; Lespinet, O. A meta-approach for improving the prediction and the functional annotation of ortholog groups. BMC Genom. 2014, 15 (Suppl. 6), S16. [Google Scholar] [CrossRef] [PubMed]
- Strobl, C.; Malley, J.; Tutz, G. An introduction to recursive partitioning: Rationale, application, and characteristics of classification and regression trees, bagging, and random forests. Psychol. Methods 2009, 14, 323–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, W.W.; Wong, T.W.; Wong, A.H.; Hui, D.S. Effect of dust storm events on daily emergency admissions for respiratory diseases. Respirology 2012, 17, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Lai, L. Public health risks of prolonged fine particle events associated with stagnation and air quality index based on fine particle matter with a diameter <2.5 μm in the Kaoping region of Taiwan. Int. J. Biometeorol. 2016, 60, 1907–1917. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Tukey’s honesty significant difference (HSD) test. In Encyclopedia of Research Design; Salkind, N., Ed.; Sage: Thousand Oaks, CA, USA, 2010; pp. 1565–1570. [Google Scholar]
- Mao, W. Leave-one-out cross-validation-based model selection for multi-input multi-output support vector machine. Neural Comput. Appl. 2014, 24, 441–451. [Google Scholar] [CrossRef]
- Yu, C.H. Exploratory data analysis in the context of data mining and resampling. Int. J. Psychol. Res. 2010, 3, 9–22. [Google Scholar] [CrossRef]
- Yu, C.H. Dancing with the Data: The Art and Science of Data Visualization; LAP: Saarbrucken, Germany, 2014. [Google Scholar]
- Cook, E.F.; Goldman, L. Empiric comparison of multivariate analytic techniques: Advantages and disadvantages of recursive partitioning analysis. J. Chronic Dis. 1984, 37, 721–731. [Google Scholar] [CrossRef]
- Kattan, M.W.; Hess, K.R.; Beck, J.R. Experiments to determine whether recursive partitioning (CART) or an artificial neural network overcomes theoretical limitations of Cox proportional hazards regression. Comput. Biomed. Res. 1998, 31, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. Prediction and entropy. In A Celebration of Statistics; Atkinson, A.C., Fienberg, S.E., Eds.; Springer: New York, NY, USA, 1985; pp. 1–24. [Google Scholar]
- Faraway, J.J. Extending the Linear Model with r (Texts in Statistical Science); Chapman & Hall/CRC: New York, NY, USA, 2005. [Google Scholar]
- Lu, Y.; Fang, T.B. Examining personal air pollution exposure, intake, and health danger zone using time geography and 3D geovisualization. ISPRS Int. J. Geo-Inf. 2014, 4, 32–46. [Google Scholar] [CrossRef]
- Packard, G. On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Biol. J. Linn. Soc. 2014, 113, 1167–1178. [Google Scholar] [CrossRef]
- Xiao, X.; White, E.P.; Hooten, M.B.; Durham, S.L. On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Ecology 2011, 92, 1887–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.H.; Lee, H.S.; Gan, S.; Brown, E. Nonlinear modeling with big data in SAS and JMP. In Proceedings of the Western Users of SAS Software Conference (paper presented), Long Beach, CA, USA, 22 September 2017. [Google Scholar]
- Health Effects Institute. State of Global Air 2018 Special Report. Health Effects Institute: Boston, MA. Available online: https://www.stateofglobalair.org/sites/default/files/soga-2018-report.pdf (accessed on 21 April 2018).
- Miller, M.; Shaw, C.; Langrish, J. From particles to patients: Oxidative stress and the cardiovascular effects of air pollution. Future Cardiol. 2012, 8, 577–602. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Guo, Y.; Zhang, Y.; Westerdahl, D.; Mo, Y.; Liang, F.; Pan, X. Spatiotemporal analysis of particulate air pollution and ischemic heart disease mortality in Beijing, China. Environ. Health 2014, 13, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.S.; Zhou, J.; Wong, P.W.; Kowalisyn, J.; Strokosch, G. Intermediate homocysteinemia: A thermolabile variant of methylenetetrahydrofolate reductase. Am. J. Hum. Genet. 1998, 43, 414–421. [Google Scholar]
- Leclerc, D.; Sibani, S.; Rozen, R. Molecular biology of methylenetetrahydrofolate reductase (MTHFR) and overview of mutations/polymorphisms. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2000. Available online: http://www.ncbi.nlm.nih.gov/books/NBK6561/ (accessed on 29 May 2018).
- Mahmoodi, K.; Nasehi, L.; Karami, E.; Soltanpour, M.S. Association of nitric oxide levels and endothelial nitric oxide synthase G894T polymorphism with coronary artery disease in the Iranian population. Vasc. Specialist Int. 2016, 32, 105. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.C.; Hsiao, J.Y.; Tien, K.J.; Chang, S.J.; Lin, P.C.; Hsu, S.C.; Lin, S.R. The association of endothelial nitric oxide synthase G894T polymorphism with C-reactive protein level and metabolic syndrome in a Chinese study group. Metabolism 2008, 57, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Mackawy, A.M.; Khan, A.A.; Badawy, M.E. Association of the endothelial nitric oxide synthase gene G894T polymorphism with the risk of diabetic nephropathy in Qassim region, Saudi Arabia—A pilot study. Meta Gene 2014, 2, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y. Endothelial nitric oxide synthase G894T gene polymorphism and essential hypertension in the Chinese population: A meta-analysis involving 11,248 subjects. Intern. Med. 2011, 50, 2099–2106. [Google Scholar] [PubMed]
- Angeline, T.; Krithiga, H.R.; Isabel, W.; Asirvatham, A.J.; Poornima, A. Endothelial nitric oxide synthase gene polymorphism (G894T) and diabetes mellitus (type II) among South Indians. Oxid. Med. Cell. Longev. 2011, 2011, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Tso, A.W.; Tan, K.C.; Wat, N.M.; Janus, E.D.; Lam, T.H.; Lam, K.S. Endothelial nitric oxide synthase G894T (Glu298Asp) polymorphism was predictive of glycemic status in a 5-year prospective study of Chinese subjects with impaired glucose tolerance. Metabolism 2006, 55, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Li, B.; Li, J.; Sun, Y. The Effects of Age, Period, and Cohort on Mortality from Ischemic Heart Disease in China. Int. J. Environ. Res. Public Health 2017, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Mu, Z.; Jiang, B.; Wang, W.; Yu, H.; Zhang, L.; Li, J. Acute effects of particulate air pollution on ischemic heart disease hospitalizations in Shanghai, China. Int. J. Environ. Res. Public Health 2017, 14, 168. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Cases | Controls | Tests of Association | ||
|---|---|---|---|---|---|
| (Number of Studies) | n = 16,219 (%) | n = 12,222 (%) | Model | RR (95% CI) | p |
| TT (61) | 1551 (9.56) | 759 (6.21) | Random | 1.44 (1.23, 1.67) | 0.0001 |
| Caucasian (25) | 1230 (12.30) | 593 (10.21) | Random | 1.30 (1.08, 1.56) | 0.0051 |
| Hispanic (2) | 33 (6.00) | 17 (4.43) | Fixed | 1.34 (0.76, 2.34) | 0.3044 |
| East Asian (18) | 79 (2.60) | 26 (0.74) | Fixed | 2.17 (1.46, 3.25) | 0.0001 |
| South Asian (6) | 40 (4.16) | 18 (2.08) | Fixed | 2.11 (1.21, 3.65) | 0.0076 |
| Middle East (4) | 65 (9.31) | 25 (4.26) | Fixed | 2.35 (1.47, 3.74) | 0.0003 |
| African (6) | 104 (10.54) | 80 (8.06) | Random | 1.45 (0.87, 2.41) | 0.1457 |
| GT (61) | 6066 (37.40) | 3971 (32.49) | Random | 1.37 (1.18, 1.57) | 0.0001 |
| Caucasian (25) | 4271 (42.71) | 2538 (43.69) | Random | 0.96 (0.91, 1.02) | 0.2571 |
| Hispanic (2) | 175 (31.81) | 126 (32.81) | Fixed | 0.96 (0.79, 1.16) | 0.6801 |
| East Asian (18) | 672 (22.12) | 518 (14.66) | Random | 1.38 (1.19, 1.59) | 0.0001 |
| South Asian (6) | 286 (29.79) | 229 (26.44) | Fixed | 1.17 (1.01, 1.36) | 0.0304 |
| Middle East (4) | 272 (38.96) | 180 (30.61) | Random | 1.25 (0.92, 1.70) | 0.1388 |
| African (6) | 390 (39.55) | 380 (38.27) | Random | 1.02 (0.83, 1.26) | 0.8032 |
| GG (61) | 8613 (53.10) | 7442 (60.89) | Random | 0.92 (0.89, 0.95) | 0.0001 |
| Caucasian (25) | 4498 (44.98) | 2677 (46.09) | Random | 0.95 (0.89, 1.01) | 0.139 |
| Hispanic (2) | 342 (62.18) | 241 (62.76) | Fixed | 0.99 (0.89, 1.10) | 0.9341 |
| East Asian (18) | 2286 (75.27) | 2989 (84.60) | Random | 0.91 (0.87, 0.95) | 0.0001 |
| South Asian (6) | 634 (66.04) | 619 (71.48) | Random | 0.97 (0.84, 1.13) | 0.7656 |
| Middle East (4) | 361 (51.72) | 383 (65.14) | Fixed | 0.77 (0.70, 0.85) | 0.0001 |
| African (6) | 492 (49.90) | 533 (53.68) | Random | 0.91 (0.76, 1.07) | 0.2809 |
| TT + GT (61) | 7617 (46.96) | 4730 (38.70) | Random | 1.15 (1.09, 1.22) | 0.0001 |
| Caucasian (25) | 5501 (55.01) | 3131 (53.90) | Random | 1.03 (0.98, 1.09) | 0.176 |
| Hispanic (2) | 208 (37.81) | 143 (37.24) | Fixed | 1.00 (0.85, 1.19) | 0.9339 |
| East Asian (18) | 751 (24.73) | 544 (15.40) | Random | 1.44 (1.26, 1.64) | 0.0001 |
| South Asian (6) | 326 (33.96) | 247 (28.52) | Fixed | 1.24 (1.08, 1.43) | 0.0018 |
| Middle East (4) | 337 (48.28) | 205 (34.86) | Fixed | 1.42 (1.24, 1.63) | 0.0001 |
| African (6) | 494 (50.10) | 460 (46.32) | Random | 1.10 (0.92, 1.33) | 0.2638 |
| T allele (61) | 4585 (28.20) | 2744 (22.5) | Random | 1.18 (1.11, 1.25) | 0.0001 |
| Caucasian (25) | 3366 (33.66) | 1862 (32.05) | Random | 1.07 (1.00, 1.16) | 0.0466 |
| Hispanic (2) | 120 (21.82) | 80 (20.83) | Fixed | 1.04 (0.81, 1.34) | 0.7399 |
| East Asian (18) | 415 (13.66) | 285 (8.06) | Fixed | 1.48 (1.28, 1.71) | 0.0001 |
| South Asian (6) | 183 (19.06) | 132 (15.24) | Fixed | 1.30 (1.06, 1.60) | 0.0106 |
| Middle East (4) | 201 (28.80) | 115 (19.56) | Fixed | 1.52 (1.24, 1.87) | 0.0001 |
| African (6) | 299 (30.32) | 270 (27.19) | Fixed | 1.10 (0.95, 1.26) | 0.1743 |
| G allele (61) | 11646 (71.80) | 9428 (77.2) | Random | 0.95 (0.93, 0.97) | 0.0001 |
| Caucasian (25) | 6634 (66.34) | 3946 (67.94) | Random | 0.96 (0.92,0.99) | 0.0304 |
| Hispanic (2) | 429 (78.00) | 304 (79.17) | Fixed | 0.98 (0.92, 1.05) | 0.7404 |
| East Asian (18) | 2622 (86.34) | 3248 (91.93) | Fixed | 0.95 (0.93, 0.97) | 0.0001 |
| South Asian (6) | 777 (80.94) | 734 (84.76) | Random | 1.02 (0.89, 1.16) | 0.7493 |
| Middle East (4) | 497 (71.20) | 473 (80.44) | Fixed | 0.87 (0.82, 0.93) | 0.0001 |
| African (6) | 687 (69.68) | 723 (72.81) | Fixed | 0.96 (0.90, 1.01) | 0.1761 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johns, R.; Chen, Z.-F.; Young, L.; Delacruz, F.; Chang, N.-T.; Yu, C.H.; Shiao, S.P.K. Meta-Analysis of NOS3 G894T Polymorphisms with Air Pollution on the Risk of Ischemic Heart Disease Worldwide. Toxics 2018, 6, 44. https://doi.org/10.3390/toxics6030044
Johns R, Chen Z-F, Young L, Delacruz F, Chang N-T, Yu CH, Shiao SPK. Meta-Analysis of NOS3 G894T Polymorphisms with Air Pollution on the Risk of Ischemic Heart Disease Worldwide. Toxics. 2018; 6(3):44. https://doi.org/10.3390/toxics6030044
Chicago/Turabian StyleJohns, Robin, Zhao-Feng Chen, Lufei Young, Flordelis Delacruz, Nien-Tzu Chang, Chong Ho Yu, and S. Pamela K. Shiao. 2018. "Meta-Analysis of NOS3 G894T Polymorphisms with Air Pollution on the Risk of Ischemic Heart Disease Worldwide" Toxics 6, no. 3: 44. https://doi.org/10.3390/toxics6030044
APA StyleJohns, R., Chen, Z.-F., Young, L., Delacruz, F., Chang, N.-T., Yu, C. H., & Shiao, S. P. K. (2018). Meta-Analysis of NOS3 G894T Polymorphisms with Air Pollution on the Risk of Ischemic Heart Disease Worldwide. Toxics, 6(3), 44. https://doi.org/10.3390/toxics6030044