1. Introduction
Lipases (Enzyme Commission Number: 3.1.1.3) are a class of enzymes that catalyze the hydrolysis of carboxylic ester bonds, producing partial glycerides, fatty acids, and glycerol from triglycerides. These enzymes act at the organic–aqueous solvent interface in the presence of excess water [
1]. There are many reasons (e.g., environmental benefits) for the growing interest in enzyme-mediated reactions compared to chemical processes, and lipases are gaining more importance because of their application in various fields and their properties in terms of enantioselectivity, regioselectivity, and broad substrate specificity [
2]. In addition, lipases have a unique property in that they catalyze the reverse reaction of hydrolysis in reaction media with a low water content or inorganic solvent, meaning switching thermodynamic equilibrium towards synthesis (e.g., esterification) [
3,
4]. Lipases have been employed successfully in synthetic reactions to produce high-value products such as tri- or partial glycerides [
5].
Different sources are available for lipases, including plants, animals, and microorganisms, and, specifically, the versatility of microbial lipases with relatively easy production makes them more attractive for industrial applications [
6]. Among various microbial sources, lipases from thermophilic bacteria can have high application possibilities because of their high stability and activity at higher temperatures than non-thermophilic enzymes. This property is of interest in industrial processes. Also, the high thermal stability of enzymes is considered to be positively correlated with the stability of enzymes in organic solvents [
7] because the combination of enzymes with organic solvents has been commonly used to catalyze a range of reactions. These advantages emphasize lipases from thermophilic bacteria have multiple beneficial properties and the need for screening to develop various useful thermophilic lipases.
As mentioned above, lipases can catalyze synthesis reactions in microaqueous conditions, and this research focuses on one of those reactions, producing glycerides from glycerol, especially partial glycerides (mono- and diglycerides). Partial glycerides are a nonionic type of surfactant and are widely added to different products in the food, pharmaceutical, and cosmetic industries [
8]. Currently, mono- and diglycerides of fatty acids have been authorized as food additives by the European Union in 77 food categories [
9]. and they have been considered safe food additives because most of these compounds can be hydrolyzed in the gastrointestinal tract before absorption, releasing non-toxic glycerol and fatty acids [
9]. Industrially, mono- and diglycerides can be produced in two different ways. They include (1) transesterification using a reaction between natural or hydrogenated fats/oils (fatty acid esters) and glycerol and (2) a direct esterification process using esterification of fatty acids with glycerol [
9]. In this study, both types of synthetic reactions were tested to produce partial glycerides from glycerol and fatty acids or fatty acid esters.
The members of the
Geobacillus and
Anoxybacillus genera have shown considerable potential in biotechnology due to their unique thermostable enzymes, leading to high industrial and economic benefits [
10] and, especially,
G. stearothermophilus and
A. flavithermus are the most frequent thermophilic contaminants in dairy products and manufacturing plants [
11], indicating their easy availability. In addition, a previous study showed simple purification methods (e.g., ethanol precipitation or heat treatment) compatible with these bacteria (Geobacillus stearothermophilus and Anoxybacillus flavithermus) to produce thermophilic lipases [
12]. Thermophilic
Geobacillus and
Anoxybacillus are bacteria known to withstand high temperatures, ranging from 50 to 90 °C, and produce lipases that may have important industrial applications [
13]. Notably, thermophilic lipases of these bacteria prepared from simple heat treatment indicated their potential application in the food industry in a previous study [
14]. With these idea in mind, in this current research, those thermophilic lipases from
G. stearothermophilus and
A. flavithermus by heat treatment purification were selected for the production of partial glycerides to find out their industrial applicability. The partial glyceride synthesis was conducted after additional immobilization to make them additionally fit for industrial purposes. Here, among different types of immobilization, the reaction using covalent bonding between support materials and enzymes was conducted. Immobilization can improve enzyme stability and efficiency through repeated usage [
15]. Also, it allows enzymes to be easily separated from the final products and enhances reusability and the possibility of better control of reactions, resulting in more favorable economic factors [
16].
These immobilized thermophilic lipases from
G. stearothermophilus and
A. flavithermus were utilized to produce partial glycerides from glycerol and various types of fatty acids in a solvent-free environment. This system has benefits as it simplifies reaction preparation and removes the drawbacks of solvent usage [
17]. The resulting reaction products by those immobilized thermophilic lipases above were measured using the HPLC method and compared to the pattern by a commercial immobilized lipase from
Thermomyces lanuginosus, which already showed noticeable thermostability and applications in food and different industrial areas [
18]. This comparison was used to evaluate different immobilized thermophilic lipases in terms of glyceryl decanoates synthesis. In addition, the selectivity of different immobilized lipases to distinct substrates was studied to analyze the effect of immobilization methods and microbial sources, which has a limited understanding. This current research will provide better knowledge about substrate selectivity and product patterns depending on different immobilized lipases, which can be helpful in widening manufacturers’ selectivity for proper partial glyceride production.
2. Materials and Methods
2.1. Materials
Silica gel beads (wide pore and 150 Å), molecular sieves (3 Å, 3–5 mm), and glutaraldehyde (25%) were purchased from Alfa Aesar (Haverhill, MA). Methyl decanoate, decanoic acid, and sodium tetraborate were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). 2,4,6-Trinitrobenzene sulfonic acid (5%, w/v) was purchased from Thermo Scientific (Waltham, MA, USA). 3-Aminopyl-triethoxysilane (98%) was purchased from Beantown Chemical (Hudson, NH, USA). Tris-HCl solution 1 M, calcium chloride, isopropanol, 20% (v/v) nitric acid, ethanol (95%), chloroform, HPLC grade acetone, and acetonitrile with 0.1% formic acid, and trihydrochloride were purchased from Fisher Chemicals (Waltham, MA, USA). IMMOZYME TLL-T2-150 (recombinant lipase from Thermomyces lanuginosus) was purchased from ChiralVision (Den Hoorn, The Netherlands). Tryptic soy broth (TSB), starch (from potato), 1.5 mL centrifuge tube, and 1.5 mL two-sided disposable plastic cuvettes polystyrene were from VWR (Darmstadt, Germany). Microfilter (1.0 μm) (Glassfiber Prefilter, Tullagreen, Carrigtwohill Co., Ireland), grade 4 filter paper (110 mm) (Whatman International Ltd, Maidstone, UK), and 0.2 µm HPLC syringe filter (Acrodisc LC PVDF, Gelman Sciences, Ann Arbor, MI, USA) were used. Triton X100 was purchased from Mallinckrodt Specialty Chemicals Co., St. Louis, MO, USA. Glyceryl tridecanoate and 1-decanoyl-rac-glycerol were purchased from Sigma (St. Louis, MO, USA). 1,2-Didecanote-sn-glycerol was purchased from LifeSpan BioSciences, Inc. (Seattle, WA, USA). Glycerol (Fisher Chemicals, Waltham, MA, USA), glycine solution (Mallinckrodt Specialty Chemicals Co., St. Louis, MO, USA), (3-aminopropyl)triethoxysilane (APTES) (Beantown Chemical, BTC, Hudson, NH, USA), and 0.2 M sodium phosphate buffer (pH 7.0) (Fisher Chemicals, Waltham, MA, USA) were used. All other chemicals were of reagent grade and procured from general laboratory suppliers.
2.2. Preparation of Bacterial Cultures
Two different thermophilic bacteria were used in this study, including
G. stearothermophilus (ATCC 7953) (NAMSA, Northwood, OH, USA) and
A. flavithermus TNO-09.006 (an agar slant was kindly provided by Remco Kort at Vrije University). Bacterial frozen stocks (20% glycerol in TSB,
v/
v) and experimental cultures for cell lysates were prepared using TSB for
G. stearothermophilus and TSB supplemented with 0.2% (
w/
v) potato starch for
A. flavithermus according to a previously optimized method [
12]. The resulting overnight bacterial cultures showed optical density at 600 nm (OD
600) of around 1.0 by using a spectrophotometer (BioSpec 1601; Shimadzu, Pleasanton, CA, USA).
2.3. Preparation of Bacterial Cell Lysates
After centrifugation at 7100×
g and 10 °C for 10 min (Sorvall RC 5B centrifuge, Dupont Instruments, Wilmington, DE, USA) of overnight bacterial cultures to obtain intracellular lipases due to their lower preparation cost [
19], the resultant harvested cells in Tris-HCl buffer (50 mM Tris–HCl, pH 7.5) of each bacteria (
G. stearothermophilus and
A. flavithermus) were sonicated according to a previous method [
14] using a 4.5 mm microtip and a Qsonica Q500 (Qsonica, Newtown, CT, USA) at 40% amplitude for 1 min, followed by filtration using a microfilter (1.0 μm) to remove cell debris and obtain crude lysates.
2.4. Partial Lipase Purification by Heat Treatment
Lipases from each bacterial cell lysate (
G. stearothermophilus and
A. flavithermus) were purified according to a previously reported method using heat treatment [
14]. In short, the crude cell lysates of each bacteria were maintained at 70 °C for 10 min. After centrifugation at 10,000×
g and 4 °C for 30 min to remove denatured proteins, the supernatants containing thermophilic lipases were collected and analyzed for total protein concentrations using the Pierce BCA protein assay kit (Thermo Scientific, Rockford, IL, USA) according to the manufacturer’s protocol.
2.5. Preparation of Silica Beads and Thermophilic Lipase Immobilization
Derivatization of silica beads and immobilization of thermophilic lipases to the resulting silica gel beads using glutaraldehyde as a crosslinker were carried out according to a previously established method [
20]. In brief, silica gel beads were cleaned by washing with 20% nitric acid at 100 °C for 60 min and then nitric acid was removed using distilled water. Clean silica gel beads in 3 volumes of a degassed 10% APTES solution (pH 4.0) were incubated at 70 °C for 3 h to add amino groups to silica gel beads, and the mixture was washed using a distilled water before drying in an oven at 100 °C overnight. Washed aminopropyl silica gel beads were incubated with 2 volumes of degassed 0.2 M sodium phosphate buffer (pH 7.0) containing 15% (
v/
v) glutaraldehyde at 22 °C for 16 h, followed by washing using a 25 mM sodium phosphate buffer (pH 7.0). The resulting silica gel beads were mixed with 10 volumes of individual lipase solutions (3 mg of proteins per 1 mL of 50 mM Tris-HCl buffer [pH 7.5]). After incubation for 4 h (
G. stearothermophilus) or 6 h (
A. flavithermus), the mixture was filtered under suction. The dried silica gel beads were incubated in 25 mM sodium phosphate buffer (pH 7.0) containing 10 mM glycine at 4 °C overnight, followed by drying at 100 °C in an oven overnight. The information of final immobilized thermophilic lipases from
G. stearothermophilus (ILGS) and
A. flavithermus (ILAF) are shown in
Table 1, including their hydrolytic activity as reported in a previous study [
14]. This enzyme activity assay was conducted using p-nitrophenyl acetate as the substrate according to a previous method [
14]. Briefly, for the reaction, 900 μL of 1 mM p-nitrophenyl acetate solution was mixed with 100 μL of enzyme solution in 50 mM Tris HCl (pH 7.5) containing 1 mM CaCl
2. After the incubation at 20 °C for 1 min, the absorbance of reaction mixtures was analyzed at 410 nm every 5 s over 1 min. The blank was 900 μL of substrate solution and 100 μL of water. One unit of lipase activity (U) was defined as 1 μmol substrate hydrolyzed per minute under the assay conditions.
2.6. Synthesis Activity of Immobilized Thermophilic Lipases
2.6.1. Reaction
Immobilized lipases (ILGS and ILAF) with commercial immobilized thermophilic lipases from Thermomyces lanuginosus (ILTL) were placed in a water activity chamber containing saturated NaCl solution, adjusting water activity to 0.75 for 24 h. Substrate solutions for synthetic reaction were prepared by blending decanoic acid, methyl decanoate, or vinyl decanoate with glycerol (9:1 (v/v), fatty acids to glycerol) and mixed under magnetic stirring at 65 °C and 100 rpm for 10 min. Each immobilized lipase (100 mg of silica beads containing lipases) was mixed with 1 mL of individual substrate solution and 200 mg of molecular sieves, followed by incubation at 65 °C and 200 rpm for 3 days. Over the reaction, individual samples (1 mL) were collected on days 0, 1, 2, and 3 and diluted with different volumes of chloroform for compositional analysis.
2.6.2. HPLC Analysis
Instead of an analysis method with GC equipment, requiring extra sample preparation (e.g., methylation) [
21], the compositional difference during synthesis reaction by immobilized thermophilic lipases was analyzed using an HPLC system (Beckman System Gold 125 solvent Mobile, Beckman Coulter, Pasadena, CA, USA) equipped with an evaporative light-scattering (ELSD) detector (Agilent Technologies Group, Hesperia, CA, USA). Each sample (20 µL) was injected after heating at 40 °C for 2 min in a water bath. The analysis was conducted using a SynChropak RP-4 C8 (250 × 4.6 mm) column (SynChrom. Inc., Lafayette, IN, USA) and two different mobile phases: (A) 100% acetone and (B) 100% acetonitrile containing 0.1% formic acid. The flow rate was 1 mL per min with a linear gradient as follows: 0–5 min 0 to 60% (A), 5–15 min 60 to 80% (A), 15 to 20 min 80 to 85% (A), and 20 to 45 min 85 to 100% (A). The column temperature was 40 °C using a column heater (Bio-Rad 125-0425 HPLC Column Heater, Redmond, WA, USA). The compounds were detected using an ELSD at 40 °C with 3.55 bar of nitrogen gas. The composition of each reaction mixture was identified by matching retention times of the known standard compounds, and the area of each peak in chromatograms obtained was measured by LP-chrom software (Lipopharm, Poland) (
Figure 1). Each glyceryl decanoate (µmol mL
−1) produced per 1 mg of the immobilized lipases was normalized to the hydrolytic activities (U mg
−1) of each enzyme used to ensure that the same level of the enzyme was used.
2.7. Statistical Analysis
Every experiment was conducted in duplicate. The mean ± standard deviation was used to indicate results. The results were analyzed with repeated measure ANOVA using PROC MIXED (fixed factors: type of substrates, reaction time, and type of products; repeated measure factor: type of products) followed by post hoc analysis using the Simulation method (SAS version 9.4, SAS Institute Inc., Cary, NC, USA). A statistically significant difference was calculated when p < 0.05.
3. Results and Discussion
Thermophilic lipases prepared from
G. stearothermophilus (ILGS) and
A. flavithermus and (ILAF) were immobilized, and their hydrolytic activities, as reported in a previous study [
14], are included in
Table 1. Both ILGS and ILAF showed similar activities (0.011 U mg
−1 for ILGS and 0.010 U mg
−1 for ILAF), and these values were around 10 times lower than that (0.100 U mg
−1) obtained from a commercial immobilized thermophilic lipase from
T. lanuginrsus (ILTL) (
Table 1). These similar hydrolytic activities between ILGS and ILAF and their large difference from ILTL might be due to the different immobilization methods because the final activities of immobilized enzymes are dependent on the immobilization conditions, such as enzyme loading concentrations per support material [
22]. In this context, it is not informative to directly compare the enzyme activities between those prepared in this research and a commercial immobilized lipase without considering influences by different immobilization processes. To overcome this issue, in this current research, these hydrolytic activities of enzymes were utilized later to normalize concentrations of the products (mono-, di-, and triglycerols) during the synthetic reaction by the immobilized thermophilic lipases. This adjustment can be helpful in clarifying the distinct substrate selectivity and product patterns by different immobilized lipases.
Different glyceride products were analyzed using HPLC equipped with ELSD, and their separation patterns are shown in
Figure 1. The retention times of analytes depend on their interaction with reverse phase (hydrophobic) C8 column and mobile phases. Because of the relatively highly hydrophilic property of glycerol, it was eluted in the void volume without proper interaction with the column (
Figure 1a). Following that, different glycerides were eluted in the order of increasing hydrophobicity (mono-, di-, and triglycerides) (
Figure 1a). Glyceryl monodecanoate was slightly early eluted than decanoic acid, which is thought to be the effect of glycerol (
Figure 1a). The similar separation order by HPLC using reverse phase C18 column and acetonitrile was also reported in a previous study [
21]. This complete separation of substrates and synthesis products was also obtained from the reaction mixtures (
Figure 1b–d), and their composition was analyzed based on the retention time and area of analytes separated.
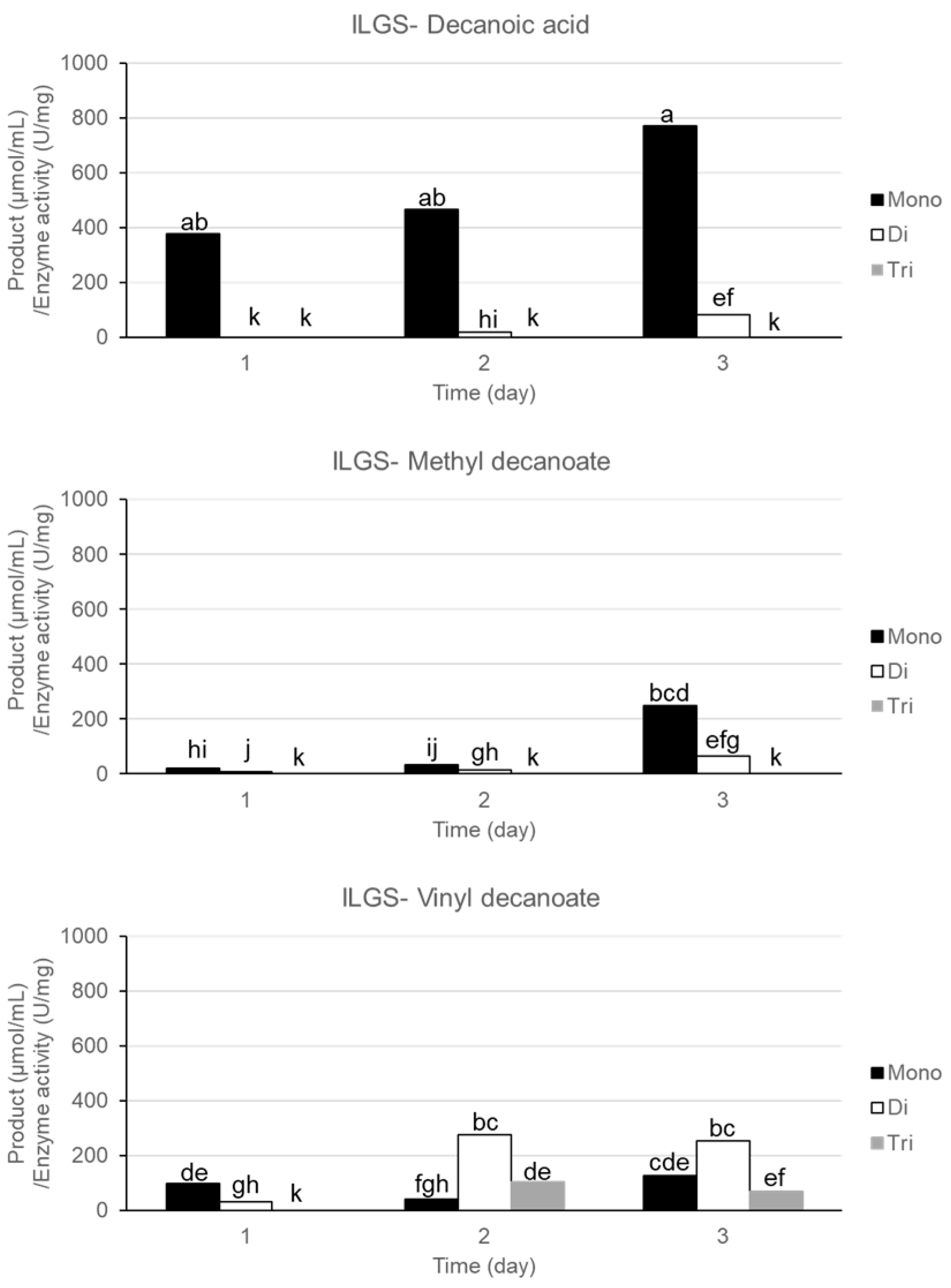
The synthesis of various glyceryl decanoates (mono-, di-, or tri-) by ILGS from glycerol and different fatty acid types (decanoic acid, methyl decanoate, and vinyl decanoate) is shown in
Figure 2. A similar pattern of products was obtained from ILGS between decanoic acid and methyl decanoate as a substrate (mainly glyceryl monodecanoate with some of didecanoate) (
Figure 2). However, the higher reaction products by ILGS were produced using decanoic acids (
Figure 2), indicating the better substrate selectivity of ILGS to decanoic acid. Instead, ILGS synthesized all three types of glyceryl decanoates from vinyl decanoate (primarily glyceryl didecanoate) with a lower amount of total synthetic products compared to those from decanoic acids (
Figure 2). This result suggests the highest substrate selectivity of ILGS to decanoic acid showing a major product, glyceryl monodecanoate and potential to control synthetic product patterns from a same enzyme usage according to the source of substrates. In addition, the patterns of reaction products were irrespective of the type of esterification reactions because the product pattern from methyl decanoate (transesterification) corresponded to that from decanoic acid (direct esterification) instead of that from vinyl decanoate (transesterification). This might indicate the higher importance of fatty acid forms than the esterification types in terms of reaction products and also suggest distinct esterification mechanisms in lipase active sites according to dissimilar functional groups of fatty acid esters.
The synthesis of different glyceryl decanoates by ILAF is indicated in
Figure 3. Highly similar synthetic product patterns from decanoic acid vinyl decanoate by ILAF were obtained compared to those by ILGS. In contrast, ILAF (glycerol didecanoate as a primary product) showed different synthetic product patterns from methyl decanoate in comparison to ILGS (glycerol monodecanoate as a major product) (
Figure 2 and
Figure 3). Notably, similar substrate selectivity between ILAF and ILGS was obtained (
Figure 2 and
Figure 3), indicating the highest concentration of glyceryl monodecanoate as a main product among different glyceryl decanoates when decanoic acid was used (
Figure 2 and
Figure 3). Based on the fact that lipase substrate specificity can be determined by the modification of their active site structures in the presence of specific substrates [
23], the immobilization method utilized in the current study might limit this structural flexibility of different lipases, leading to the similar substrate selectivity between ILAF and ILGS. A similar effect of lipase lid modification on substrate specificity was previously reported [
24]. However, ILAF mainly produced glyceryl didecanoate after 3 days of the reaction from methyl decanoate, which was different from the product pattern of ILGS. This synthetic product deviation is thought to have occurred due to different amino acid sequences in active sites of lipases from microbial sources, even though the same immobilization procedure was conducted. This hypothesis has to be further supported by analysis of the amino acid sequences of lipases utilized.
Commercial immobilized lipases (ILTLs) were added in this research to compare with those prepared in the lab (
Figure 4). The lipases from
T. lanuginosus were reviewed in terms of food and other industrial applications in a previous study [
18]. Commercial ILTL showed the highest substrate selectivity to methyl decanoate (primary production of glyceryl monodecanoate and no synthesis of glyceryl tridecanoate) (
Figure 4). This different substrate selectivity of commercial ILTL for synthetic reaction compared to the lab-made ILGS and ILAF might be explained by several factors, including the different sources of microbes and the different immobilization conditions. The immobilization method, in particular, was considered as a primary element because of similar substrate selectivity between ILGS and ILAF regardless of microbial sources.
Based on the manufacturer’s product description, precise immobilization conditions for commercial ILTL are unclear. However, this detail still contains important information, indicating a difference in covalently attached supporting materials for immobilization (acrylic bead [size: 150–300 µm] for ILTL and silica gel bead [<74 µm] for ILGS and ILAF). Even though similar immobilization methods using covalent bonding were assumed to be utilized for ILTL, ILGS, and ILAF, the different supporting materials were hypothesized to influence the interaction between substrate and lipases attached, leading to distinct substrate selectivity. In a previous study, the effect of different supporting materials for lipase immobilization on substrate specificity and enzyme stability was reported, although the reaction was hydrolytic [
25]. The results of the current and previous studies indicate the importance of proper supporting materials selection as well as physicochemical reaction condition optimization in terms of final products.
In the case of the production of glyceryl tridecanoates, all immobilized lipases (ILGS, ILAF, and ILTL) did not show good applicability because of their low amount over 3 days compared to partial glycerides. Instead, for the production of partial glycerides (glyceryl mono-and didecanoates), all three immobilized lipases indicated potential usefulness. Notably, as mentioned above, because these partial glycerides are non-toxic additives and widely used in different industries, both ILGS and ILAG prepared in the current study can indicate new microbial sources applicable in different fields for the production of partial glycerides. In particular, both ILGS and ILAF showed the best compatibility with decanoic acids, while methyl decanoate is a better substrate for commercial ILTL. These distinct substrate selectivities can lead to different final product mixtures due to unique byproduct production and also influence downstream processing. Water can be produced as a byproduct by the direct esterification of glycerol and decanoic acids, while the transesterification of glycerol and methyl decanoate can produce methanol, thereby requiring additional processing to manage toxic byproducts. In this context, both ILGS and ILAF might be more environmentally compatible with producing partial glycerides from glycerol due to their highest substrate selectivity to decanoic acids. In addition, the synthetic reactions tested in the current research for immobilized lipases selected a solvent-free environment, utilizing only substrates and lipase catalysts. This reaction system enhances additional environmental and economic advantages by removing drawbacks of the use of organic solvents, which are generally used. In further research, the stability and recovery rate of these immobilized lipases can be analyzed depending on different pH and temperature ranges. Also, different immobilization materials and conditions can be studied in terms of their efficiency and substrate selectivity. This further information can be necessary to develop readily available immobilized lipases.
In the current study, microbial source was initially hypothesized as a primary factor for differentiating substrate selectivity due to the potential difference of amino acid sequences in enzyme active sites. However, based on the synthesis results, this active site sequence might be less important than the overall binding pocket structure of the enzyme. Different immobilization methods might largely determine this shape and decide compatible substrate types. This fact might highlight the importance of immobilization optimization in terms of desired substrates and the potential of lipases, which can be simple and low-cost regardless of the microbial sources.







{kind=link}
{kind=link}
{kind=link}
{kind=link}