Cyanobacterial PHA Production—Review of Recent Advances and a Summary of Three Years’ Working Experience Running a Pilot Plant

1
Institute of Environmental Biotechnology, Department of Agrobiotechnology, IFA-Tulln, University of Natural Resources and Life Sciences, Vienna, Tulln 3430, Austria
2
Bioenergy2020+ GmbH, Tulln 3430, Austria
*
Author to whom correspondence should be addressed.
Bioengineering 2017, 4(2), 26; https://doi.org/10.3390/bioengineering4020026
Submission received: 7 March 2017
/
Revised: 15 March 2017
/
Accepted: 16 March 2017
/
Published: 28 March 2017
(This article belongs to the Special Issue Advances in Polyhydroxyalkanoate (PHA) Production)
Abstract
:Cyanobacteria, as photoautotrophic organisms, provide the opportunity to convert CO2 to biomass with light as the sole energy source. Like many other prokaryotes, especially under nutrient deprivation, most cyanobacteria are able to produce polyhydroxyalkanoates (PHAs) as intracellular energy and carbon storage compounds. In contrast to heterotrophic PHA producers, photoautotrophic cyanobacteria do not consume sugars and, therefore, do not depend on agricultural crops, which makes them a green alternative production system. This review summarizes the recent advances in cyanobacterial PHA production. Furthermore, this study reports the working experience with different strains and cultivating conditions in a 200 L pilot plant. The tubular photobioreactor was built at the coal power plant in Dürnrohr, Austria in 2013 for direct utilization of flue gases. The main challenges were the selection of robust production strains, process optimization, and automation, as well as the CO2 availability.
1. Introduction
Polyhydroxyalkanoates (PHAs) are considered as one of the most promising bioplastics. Their mechanical properties are similar to polypropylene and they can be processed in a similar way, including extrusion, injection molding, or fiber spinning [1]. One of the major advantages of PHAs are their biodegradability. They are degraded relatively rapidly by soil organisms, allowing easy composting of PHA waste material [2].
Currently, PHA is produced in large fermenters by heterotrophic bacteria, like Cupriavidus necator or recombinant Escherichia coli [3]. For these fermentation processes large amounts of organic carbon sources like glucose are necessary, accounting for approximately 50% of the total production costs [4]. An alternative way of producing PHA is the use of prokaryotic algae, better known as cyanobacteria. As part of the phytoplankton, they are global primary biomass producers using light as the sole energy source to bind atmospheric CO2 [5]. Burning of fossil fuels has increased the atmospheric CO2 concentration from approximately 300 ppm in 1900 to over 400 ppm today. The latest report of the intergovernmental panel on climate change (IPCC) clearly indicates anthropogenic CO2 emissions as the main driver for climate change [6]. Given these facts, cultivation of cyanobacteria for PHA production could be a more sustainable way of producing bioplastics.
2. Cyanobacteria and Cyanobacterial Energy and Carbon Storage Compounds
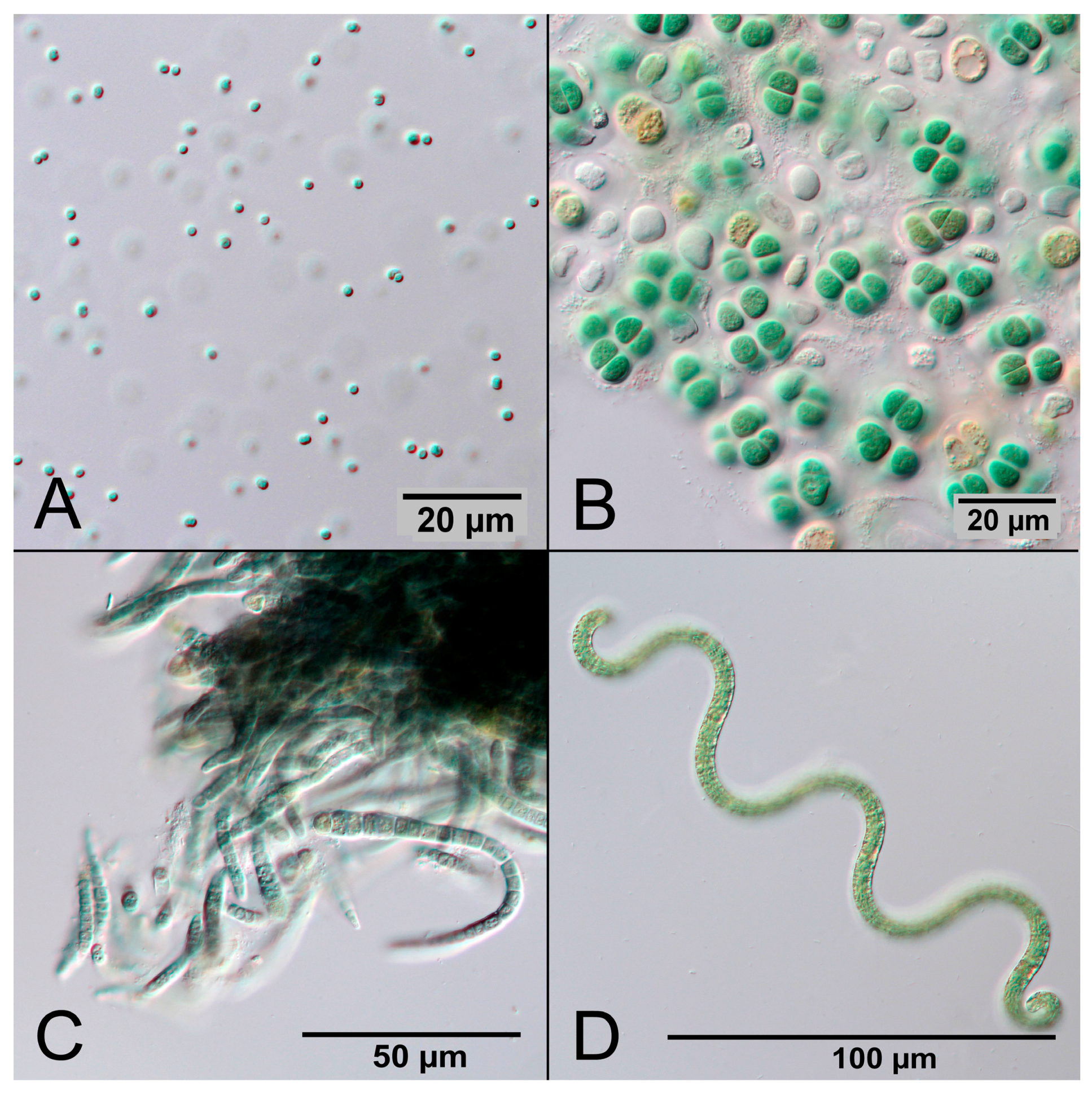
Cyanobacteria are Gram-negative prokaryotes that perform oxygenic photosynthesis. They are abundant in illuminated aquatic ecosystems and contribute significantly to the world carbon and oxygen cycle [7]. According to current evidence, oxygen was nearly absent in the Earth’s early atmosphere until 2.4 billion years ago [8]. Due to oxygenic photosynthesis of early cyanobacteria the CO2-rich atmosphere gradually turned into an oxygen-rich atmosphere, providing the conditions for multicellular life [9,10]. Today there are an estimated 6000 species of cyanobacteria with great diversity, for example ranging in size from the 1 µm small unicellular Synechocystis sp. to the several millimeter-long multicellular filaments of Oscillatoria sp. [11]. The common feature of cyanobacteria is the presence of the pigment phycocyanin, which gives them their typical blue-green color. Figure 1 shows photographs of four different cyanobacterial species.
2.1. Cyanobacteria–Microalgae or Not?
For more than a century, cyanobacteria were considered as an algal group under the general name “blue-green algae”. They were classified under the International Code of Botanical Nomenclature, nowadays called the International Code of Nomenclature for Algae, Fungi, and Plants (ICN). In 1980 the International Code of Nomenclature of Bacteria, nowadays called the International Code of Nomenclature of Prokaryotes (ICNP), was established. Stanier, one of the leading cyanobacteria researchers at that time, proposed the inclusion of cyanobacteria in the ICNP [12]. Nevertheless, the ICNP was not consistently applied for cyanobacteria and cyanobacteria are still covered by the ICN as well. The latest preamble of the ICN clarifies, that this code applies to all organisms traditionally treated as algae, fungi, or plants, including cyanobacteria [13]. Today cyanobacteria continue to be covered by both the Botanical Code (ICN) and Prokaryotic Code (ICNP). An effort to reconcile the status of this group of bacteria has been underway for several decades. Although some progress has been made, a final decision has not yet been reached [14]. From a phylogenetic point of view, there is a clear distinction between prokaryotic cyanobacteria and eukaryotic green algae. However, phycologists regard any organism with chlorophyll a and a thallus not differentiated into roots, stem, and leaves to be an alga. Therefore, in phycology, the term microalgae refers to both eukaryotic green algae and cyanobacteria, microscopic in size [15].
2.2. Cyanobacterial PHA
Polyhydroxyalkanoates (PHAs) can be classified into three groups: short-chain-length-PHA (scl-PHA), medium-chain-length-PHA (mcl-PHA), and long-chain-length-PHA (lcl-PHA). They differ in mechanical and thermal properties [16]. Among the different PHAs, polyhydroxybutyrate (PHB) is by far the most common and the only PHA produced under photoautotrophic conditions reported so far. Other scl-PHAs, like P[3HB-co-3HV], are only produced when adding organic carbon precursors, like valerate, to the medium. No mcl-PHA or lcl-PHA have been reported in cyanobacteria. Therefore, the term PHB is used in this study, if no other specific PHA is described.
PHB is frequently found in cyanobacteria as an energy and carbon storage compound. In the biosphere they often have to cope with unfavourable environmental conditions. One of the most important growth limiting factors is the absence of nutrients. Nitrogen limitation is the most important and best studied trigger for PHB production in cyanobacteria [17,18,19]. Non-diazotrophic strains are not able to bind molecular nitrogen and depend on nitrogen in the form of nitrate or ammonium. Nitrogen-depleted cells cannot synthesize the necessary proteins for reproduction and, therefore, start to accumulate storage compounds like PHB. Another important function of PHB synthesis is to compensate imbalanced metabolic situations, as it acts as an electron sink and delivers new reduction equivalents in the form of NADP+ [18,19,20].
The model organism Synechocystis PCC6803 is the best-studied cyanobacterium, and its genome was fully sequenced in 1997 [21]. Most of the understanding of cyanobacterial PHB formation was gained by research done with Synechocystis PCC6803. Biosynthesis of PHB from the precursor acetyl-CoA takes place in three steps. Acetoacetyl-CoA is produced from two molecules of acetyl-CoA in a Claisen type condensation by β-ketothiolase. Next step is the reduction of acetoacetyl-CoA by the acetoacetyl-CoA reductase to form d-3-hydroxybutyryl-CoA. Ultimately, PHB is formed in a polymerization reaction by the PHA-synthase. The necessary three enzymes are encoded by the four genes phaA (slr1993), phaB (slr1994), phaC (slr1830), and phaE (slr1829). phaA and phaB are organized in one operon encoding for the β-ketothiolase and acetoacetyl-CoA reductase. phaC and phaE are also organized in one operon encoding the two subunits of the type III PHA synthase [22,23].
2.3. Cyanobacterial Glycogen
Regarding PHB synthesis, it should be kept in mind that cyanobacteria also produce glycogen as a second carbon and energy storage compound under nitrogen depletion. In fact, the glycogen content is most often higher than the PHB content and varies between 20% and 60% [24,25,26,27,28]. While PHB is produced in 3–8 larger granules, glycogen is stored in many small granules [18,29,30,31,32,33]. Glycogen is synthesized instantly after nitrogen depletion while PHB synthesis is slower [34]. Glycogen is also produced in non-depleted cells with lower content, aiding the cell to cope with short term energy deficits like the day-night cycle. Glycogen deficient mutants were shown to be highly sensitive to day-night cycles [35]. Glycogen synthesis is a highly-conserved feature abundant in all cyanobacterial genomes reported so far [36]. PHB synthesis on the other hand is common in many, but not all, cyanobacteria [37,38]. Glycogen shows similarities to starch in green algae, while PHB synthesis shows some similarities to triacylglycerol (TAG) synthesis in green algae, where TAG synthesis also serves as an electron sink and consumes excess NADPH [39,40].
In a recent study Damrow and colleagues compared PHB-deficient mutants to glycogen-deficient mutants of Synechocystis PCC6803. Glycogen-deficient mutants could not switch to a dormant metabolic state and could not recover from nitrogen depletion. Excess carbon was mostly secreted into the medium in the form of 2-oxoglutaric acid and pyruvate, although the PHB content also increased from 8% to 13%. PHB-deficient mutants, on the other hand, behaved very much like the wild-type with the same amount of glycogen accumulation and the same recovery capability. Only double-knockout mutants (glycogen and PHB deficient) were most sensitive and showed a reduced growth rate, signs for a very specific role of PHB in cyanobacteria, which is still not totally clear [41]. The reported studies show that inhibiting glycogen synthesis increases the PHB production, although cells suffer as glycogen plays an important role.
2.4. Nitrogen Chlorosis and Photosynthetic Activity
During nitrogen starvation the cells gradually change from a vegetative state to a dormant state. The most obvious feature of this is the change in colour from blue-green to brownish-yellow. This phenomenon is called “nitrogen chlorosis” and was described already at the begin of the 20th century [42]. It is caused by the degradation of the pigments phycocyanin and chlorophyll. When transferring Synechococcus PCC7942 to a nitrogen depleted medium, 95% phycocyanin was degraded within 24 h, and after 10 days 95% of the chlorophyll was also degraded [43]. Concomitantly, the activities of the photosystems (PS) I and II decrease strongly and are only about 0.1% compared to vegetative cells [44]. A recent and very interesting study examined the awakening of a dormant Synechocystis PCC6803 cell. After the addition of nitrate the yellow culture turned green again within 36 hours. Transmission electron microscopy revealed the rapid degradation of glycogen and PHB. During the first 24 h of this process the cells consumed oxygen. Transcriptome analysis showed the induction of RuBisCO and carboxysom associated RNAs, as well as the photosystem-related RNAs to prepare the cells for vegetative photoautotrophic growth [34]. The results indicate the decrease in photosynthetic activity during nitrogen starvation, which can be considered a significant challenge to photoautotrophic PHB production.
3. Different Cyanobacteria as PHA Producers
3.1. Synechocystis and Synechococcus
Synechocystis and Synechococcus are very small (0.5–2 µm) unicellular cyanobacteria abundant in almost all illuminated saltwater and freshwater ecosystems. One of the first detailed descriptions of PHB accumulation in Synechocystis PCC6803 was provided by Wu and colleagues. Nitrogen starved cells produced 4.1% PHB of cdw while under-balanced culturing conditions PHB content were under the detection limit [45]. The same strain was examined for PHB production some years later, where 9.5% PHB of cdw were produced under nitrogen limitation. Phosphorous-depleted cells showed 11.2% PHA of cdw. Interestingly, balanced cultivated control cultures already contained 4.5% PHB of cdw. Supplementation of acetate and fructose lead to a PHB content of 38% of cdw [46]. Recently, recombinant Synechocystis PCC6803 with overexpression of the native PHA genes were constructed. They showed a PHB content of 26% of cdw under nitrogen-depleted culturing conditions compared to 9.5% of cdw of the wild-type [47]. However, it must be considered that there are legal issues in most countries when cultivating recombinant strains outdoors. In another study the thermophilic strain Synechococcus MA19 showed a PHB content of 55% under phosphate-limited culturing conditions. This study was published in 2001 and still reports the highest PHB content under photoautotrophic conditions [48]. Table 1 shows reported PHA values of Synechocystis and Synechococcus.
3.2. Arthrospira (Spirulina)
Arthrospira (formally Spirulina) is a species of filamentous cyanobacteria that grows naturally in alkaline salt lakes. It has a high protein and vitamin content and is mainly grown as a food supplement. Recent studies have shown its antioxidant, immunomodulatory, and anti-inflammatory activities [49]. From all cyanobacterial species known, only Arthrospira sp. is produced at an industrial scale. The main reason for that is the possibility of cultivation in a highly alkaline environment that prevents contamination and enables the maintenance of a stable culture in open ponds. No exact data are available; however, we estimate the world annual production of around 5000–15,000 tons Arthrospira sp. dry weight per year [50,51,52,53].
The first description of PHB accumulation in Arthrospira was reported by Campbell and colleagues, who described a PHB content of 6% of cdw in a non-optimized mineral medium. Interestingly, the highest PHB content was measured at the end of exponential growth and decreased during stationary phase [54]. In a screening of 23 cyanobacterial strains, Arthrospira platensis had the lowest PHB concentration of only 0.5% in a non-optimized medium [37]. In a screening of several Arthrospira species the PHB amount never exceeded 1% of cdw in photoautotrophic growth. Addition of sodium acetate led to a PHB amount of 2.5% of cdw [55]. In another experiment Arthrospira platensis was grown under phosphate limitation and reached 3.5% PHB of cdw [56]. Arthrospira subsalsa, a strain isolated from the Gujarat coast, India, produced 14.7% PHB of cdw under increased salinity [57]. A detailed ultrastructural analysis of Arthrospira strain PCC8005 was conducted by Deschoenmaker and colleagues. Under nitrogen depleted conditions PHB granules were more abundant and larger. The nitrogen-starved cells showed an estimated four times higher PHB concentration [27]. Nitrogen starvation was performed in Arthrospira maxima and glycogen and PHB content was measured. While the glycogen content increased from around 10% to 60%–70% of cdw, PHB amount remained low at 0.7% of cdw. The addition of sodium acetate increased the PHB amount to 3% of cdw [26].
The performed studies support the idea, that PHB production in Arthrospira is highly strain-dependent. Most Arthrospira species produce PHB only in amounts of lower than 5%, even with the addition of sodium acetate. Arthrospira produces glycogen as storage compound, what has been shown in ultrastructural research, too [27]. Nevertheless, it must be emphasized that Arthrospira, at an industrial scale, is still one of the most promising candidates for PHB production with cyanobacteria. Indeed, PHB nanofibers were produced recently from Arthrospira PHB and showed highly favourable properties [58,59]. The biggest challenge for further research is to increase the relatively low PHB content of Arthrospira. Table 2 shows reported PHA values of Arthrospira.
3.3. Nostoc
Nostoc is a group of filamentous cyanobacteria very common in terrestrial and aquatic habitats. They are capable of fixing atmospheric nitrogen with specialized heterocysts and are suspected to maintain soil fertility in rice fields due to nitrogen fixation [60]. Ge-Xian-Mi, an edible Nostoc species, forms spherical colonies that have been collected in China for centuries [61]. The first reports found for PHB production in Nosctoc muscorum are from 2005, when Sharma and Mallick showed that Nostoc muscroum produced 8.6% PHB of cdw under phosphate and nitrogen limitation during the stationary phase. PHB content could be boosted to 35% of cdw with 0.2% acetate and seven days dark incubation [62]. Limited gas exchange and supply with 0.4% acetate increased the PHB content to 40% [63]. Nostoc muscorum was grown photoautotrophically without combined nitrogen sources and four days of phosphate deficiency increased PHB content from 4% to 22% [56]. The co-polymer P[3HB-co-3HV] could be produced by Nostoc in a propionate- and valerate-supplied medium. The 3HV fraction ranged from 10–40 mol% and showed desirable properties in terms of flexibility and toughness. Nitrogen and phosphate depletion led to a PHA content of 58%–60% of cdw, however, the total cdw did not exceed 1 g/L [64]. Further process optimization led to a PHA productivity of 110 mg/L/d and a P[3HB-co-3HV] content of 78% of cdw, the highest yield in heterotrophic grown cyanobacteria reported so far [65]. Recently, poultry litter was used for cultivation of Nostoc muscorum agardh. The poultry litter contained phosphate, ammonium, nitrate, and nitrite as nutrients for cyanobacterial growth. Optimized conditions, which included the addition of acetate, glucose, valerate, and CO2-enriched air, led to a P[3HB-co-3HV] content of 70% cdw. However, total cdw remained relatively low at 0.68 g/L [66].
The reported studies show that PHB content in Nostoc can be significantly increased with organic carbon sources, especially in the form of acetate. However, those organic carbon sources lead to heterotrophic growth and may suppress CO2 uptake by the cells, which is the most important argument for using cyanobacteria as PHA producers. All of the reported experiments of Nostoc were performed in shaking flasks or small reactors under sterile conditions. In mass cultivation Nostoc would have to be cultivated under non-sterile conditions and organic carbon sources could cause problems maintaining stable cultures. Although optimized conditions of several experiments lead to PHA contents of more than 50% of cdw, the total cdw remained mostly under 1 g/L and the overall productivity and growth rate of Nostoc is relatively low. Table 3 shows reported PHA values of Nostoc.
3.4. Other Cyanobacteria
Recently, the PHB content of 137 cyanobacterial strains representing 88 species in 26 genera was determined under photoautotrophic conditions. High PHB content was highly strain-specific and was not associated with the genera. From the 137 tested strains, 134 produced PHB and the highest content was measured in Calothrix scytonemicola TISTR 8095 (Thailand Institute of Scientific and Technological Research). This strain produced 356.6 mg/L PHB in 44 days and reached a PHB content of 25% of cdw and a total biomass of 1.4 g/L. The PHB content of 25% was reached under nitrogen depletion, while cells with nitrogen supply reached a PHB content of only 0.4%. From the 19 tested Calothrix strains, only six produced more than 5% PHB of cdw. One of the greatest advantages of Calothrix is the relative ease of harvesting the dense flocs of algae, but cultivation of Calothrix is still at a very early stage [38].
The filamentous diazotroph cyanobacterium Aulosira fertilissima produced 10% PHB of cdw under photoautotrophic conditions and phosphate deficiency. The PHB content was boosted to 77% under phosphate deficiency with 0.5% acetate supplementation. This study also shows the positive effect of other carbon sources like citrate, glucose, fructose and maltose on PHB production [67]. Anabaena cylindrica, a filamentous cyanobacterium, was examined for PHB and P[3HB-co-3HV] production. Under nitrogen depletion with acetate supply, Anabeana cylindrica produced 2% PHB of cdw and a total biomass of 0.6 g/L. This organism was also able to produce the co-polymer P[3HB-co-3HV] when supplemented with valerate and propionate [68]. Table 4 shows reported PHA values of different cyanobacterial species.
4. CO2 and Nutrient Supply for Mass Cultivation of Cyanobacteria
4.1. CO2 Supply
Today, commercial microalgae production is still mainly taking place in open ponds. Here, the C source is normally sodium bicarbonate or atmospheric CO2. In order to boost productivities in open systems, or if photobioreactor systems are employed, the use of commercial CO2 from gas cylinders is common [70].
However, current production systems are used for the production of high value products (food, feed additives), where CO2 price is not critical. If PHA is to be produced, which has a lower economic value, cheap CO2 sources are of interest. Although there is considerable literature on various CO2-sources (e.g., flue gases) and microalgae growth, there is very limited literature available on cyanobacteria and alternative CO2-sources. Table 5 summarizes the literature on cyanobacterial growth on flue gases or fermentation gases.
4.2. Nutrient Supply
The cultivation of microalgae and cyanobacteria consume high amounts of nutrients, mainly nitrogen and phosphorous [77]. For research, and even cultivation, mainly synthetic nutrient sources are used [78]. By using alternative nutrient sources, like agro-industrial effluents, waste waters, or anaerobic digestate, questions concerning sustainability of cyanobacteria cultivation, which arise by using fertilizer as a synthetic nutrient source, can be answered [78]. The biomass concentration achieved in open, as well as in closed, cultivation systems are 0.5–1 g/L and 2–9 g/L, respectively [79]. Therefore, large amounts of water are needed. Recycling of process water is another important approach for a more sustainable microalgae cultivation.
In addition to their low costs, the advantages of using alternative nitrogen and phosphorous sources include the production of valuable biomass while removing nutrients from wastewaters, as well as the prevention of competition with food and feed production [78]. On the other hand, new challenges arise, including microbial contaminations, heavy metals and growth inhibitors, suspended solids, or dissolved organic compounds contained in wastewaters, as well as the seasonal composition and fluctuation in amounts [80]. To cope with these challenges recent research focused on cultivating cyanobacteria in anaerobic digestate and agro-industrial effluents or wastewaters for removing nutrients [81,82,83,84,85,86] (see Table 6) and on integrating cultivation processes into biorefinery systems [83].
Additionally, process water and nutrients after harvesting cyanobacterial biomass [79] and product extraction can be directly recycled. Biomass can also be anaerobically digested [87,88] or hydrothermally liquefied via HTL (mineralization of organic nutrients) [89,90] and then recycled. Recycling process water directly can increase the concentration of inhibitory substances and dissolved organic matter from the previous batch produced by cyanobacteria [91], which decrease the productivity of cyanobacteria. Furthermore, nutrient competition may arise by enhanced bacterial growth [79].
Although many publications deal with alternative nutrient sources for cultivating cyanobacteria, hardly any of them focus on cyanobacterial PHA production [66,92]. Reasons for that may be that PHA production requires nutrient limitation [93] and the balance between nutrient limitation, decreased growth and production rates is difficult. The colouring of the nutrient source must be respected as well [94].
5. Three Years’ Working Experience Running a Pilot Plant for Photoautotrophic PHB Production
5.1. Location and Reactor Description
The photobioreactor is situated in a glass house at the coal power station in Dürnrohr, Austria. It is a tubular system built from glass elements of Schott AG with an inner diameter of 60 mm, a total length of 80 m and a volume of 200 L (Figure 2). The main design of the photobioreactor is described elsewhere [99,100]. A central degassing unit serves to remove the oxygen as well as to compensate filling level. The medium is circulated with a 400 W centrifugal pump. pH value can be controlled through injection of pure CO2 via a mass flow controller. Additional artificial light is provided by six 250 W gas-discharge lamps. Temperature is controlled with an air conditioner.
5.2. CO2 Supply of the Reactor
The flue gases of the power plant at Dürnrohr usually contain between 11%–13% CO2. Next to the chimney is a CO2 separation plant (acronym SEPPL), providing the possibility to concentrate the CO2 and fill it into gas bottles [101] though, for a more economic approach, the CO2 should be used directly without prior compression. The SEPPL provides this option, as well as the possibility to wash the flue gases after the flue gas cleaning of the plant itself to remove residual NOx and SOx. Unfortunately for our research project, due to the current situation on the energy market, the power station is no longer run in full operation and only runs occasionally for balancing peak demands of the electrical grid. Therefore, a continuous cultivation with direct utilization of flue gas is not possible. Aspects like this must be respected when planning large industrial cultivation plants.
5.3. Automation and pH Control
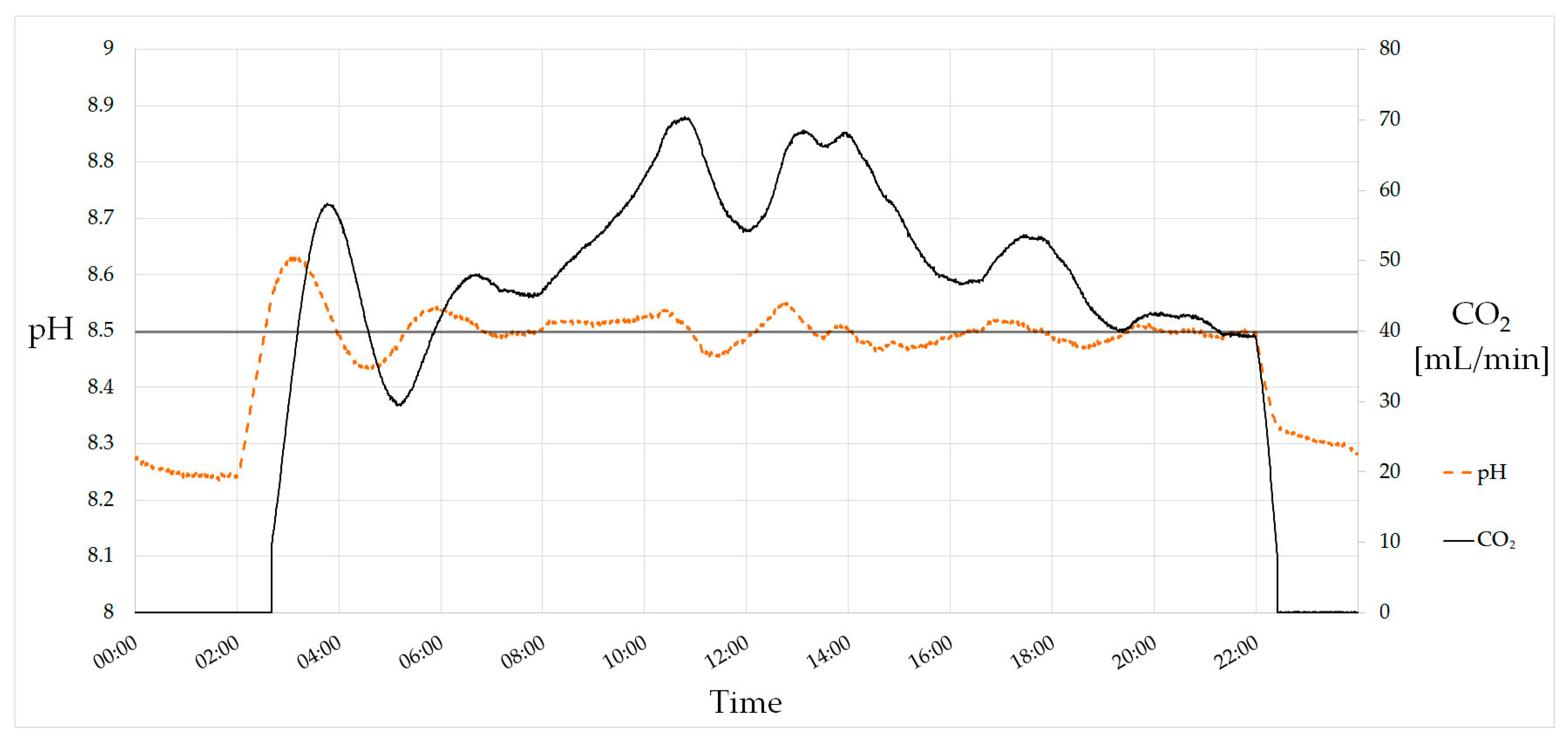
The pH value is one of the most crucial parameters and needs to be controlled carefully. Due to CO2 consumption, the pH value rises during photoautotrophic growth. This can be observed when turning on illumination. The tubular photobioreactor is equipped with a PI (Proportional–Integral) controller for pH setting which adjusts the mass-flow controller for CO2 inlet. This allows an online control of the currently consumed CO2, which is a suitable parameter for growth monitoring. Figure 3 shows a 24-h course of the pH value and the CO2 mass flow. Lamps turned on at 02:00 and off at 22:00, causing a rise and decrease of the pH value, respectively. The setpoint of 8.5 is reached after first overshooting and held during the day. The decrease of CO2 consumption at noon is caused by the shadow of the power plant’s chimney that casts upon the greenhouse at this time.
5.4. Overview of PHB Production Trials
Most of the trials (overview shown in Table 7) were performed using a modified BG 11 medium [102]. Modification in terms of PHB production is necessary, as normal BG 11 medium contains high amounts of nitrogen (1.5 g/L NaNO3) and would not lead to nitrogen limitation. The modified BG 11 contains 0.45 g/L NaNO3 and leads to a self-limitation of the culture. After 8–12 days nitrogen is consumed, PHB production starts and the color of the culture gradually turns yellow. This approach is necessary, as it is not possible to transfer large-scale cultures into a nitrogen-free medium [103].
Synechocystis salina CCALA192 was found to be a very suitable cyanobacterium. It is easy to handle and grows with small inoculation volumes of 1:50. Final biomass and PHB concentrations were in the range of 0.9–2.1 g/L and 4.8% to 9% of cdw, respectively. Synechocystis salina CCALA192 also grew with the addition of acetate, but no significant increase of biomass and PHB concentration was observed compared to photoautotrophic growth. When using acetate, contaminations with fungi were likely to occur and trials had to be stopped. Therefore, this approach was finally abandoned.
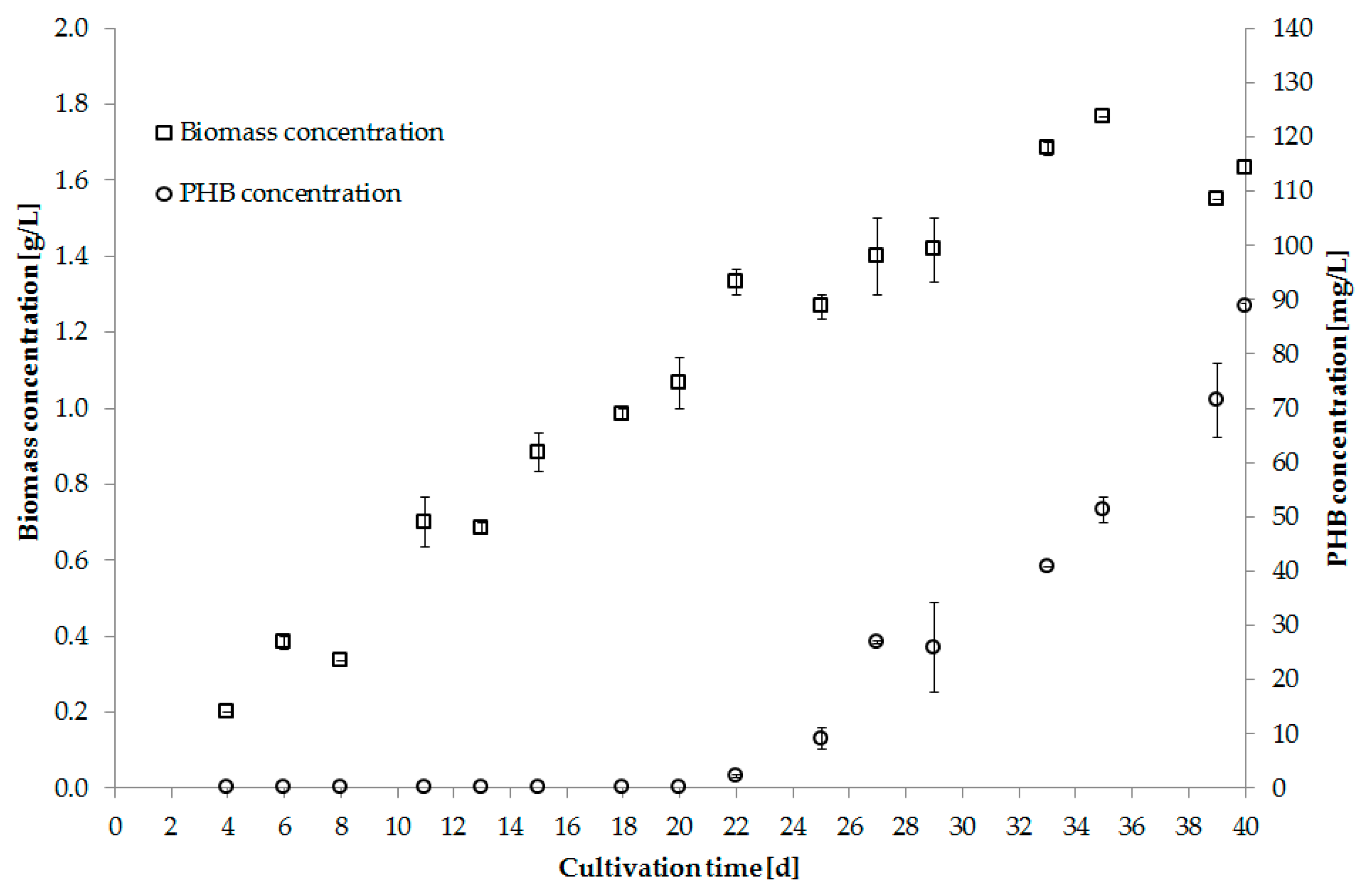
Digestate from a biogas reactor was successfully tested as an alternative nutrient source. The supernatant was produced by centrifugation with prior addition of precipitating agents. Before usage the supernatant was autoclaved and diluted 1:3 with water [92]. Figure 4 shows biomass and PHB production using digestate as nutrient source.
After one and a half years a new degassing system was installed, as the oxygen concentration was mostly above 250% saturation during the day. For an ideal cultivation of cyanobacteria the oxygen saturation should not exceed 200%. The new degasser led to a rise in biomass production with a maximum production rate of 0.25 g/L/d. Efficient degassing affected the cyanobacteria positively. However, during installation of the degasser dirt from the surrounding soil was brought into the reactor and from that time on culture crashes occurred due to ciliate contaminations (see Section 5.6).
The other tested cyanobacteria Chlorogloeopsis fritschii and Arthrospira sp. could not be successfully cultivated in the photobioreactor. It is assumed that these strains were sensitive to shear stress caused by the centrifugal pump [104].
5.5. Downstream Processing of Cyanobacterial Biomass
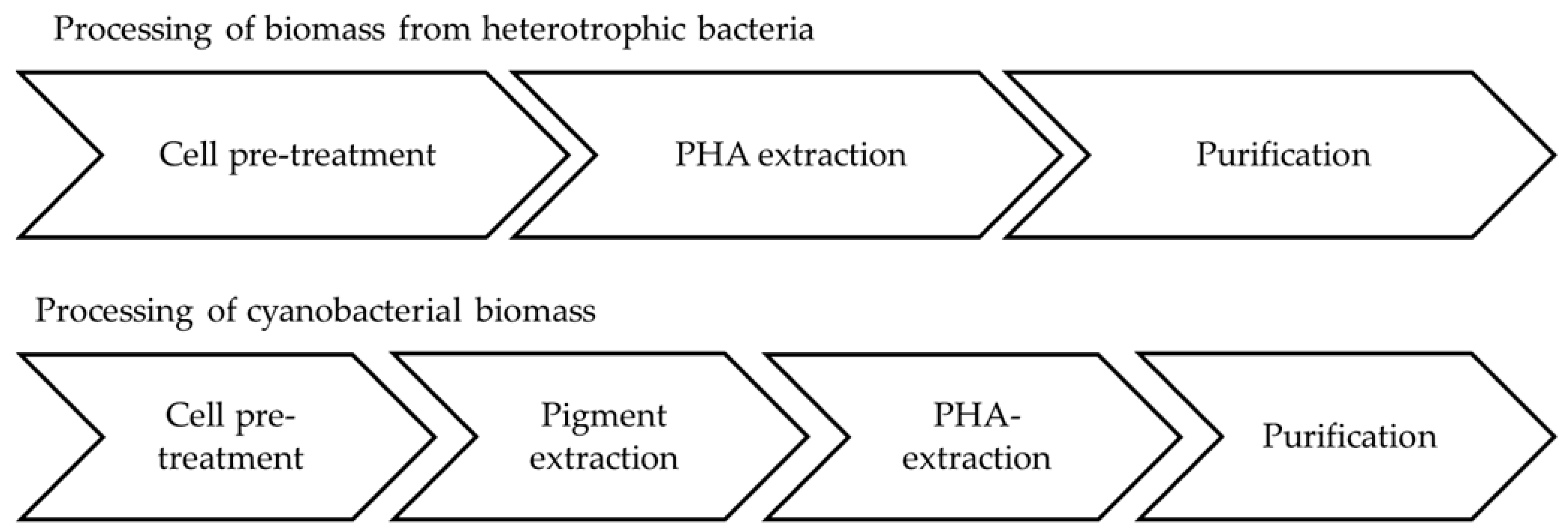
Downstreaming of cyanobacterial cultures is particularly difficult, as cell densities are much lower compared to heterotrophic bacteria. Typical harvesting methods are sedimentation, filtration, or centrifugation [105]. The cyanobacterial biomass was harvested with a nozzle separator and stored at −20 °C. The biomass was then used to evaluate processing steps necessary to gain clean PHB-samples for quality analysis. These downstream trials include (i) different cell disruption methods (milling, ultrasound, French press); (ii) different pigment extraction methods (with acetone and ethanol/methanol before or after extracting PHB); and (iii) different PHB extraction methods (soxhlet extraction with chloroform, biomass digestion with sodium hypochlorite) [106].
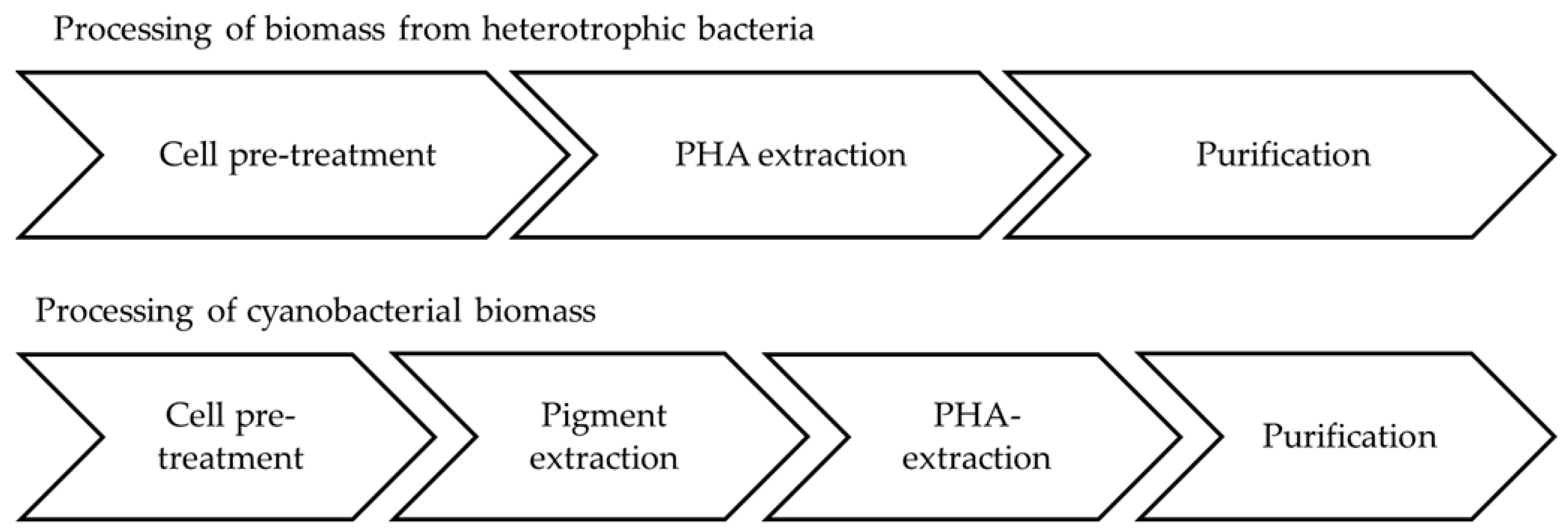
These trials showed that cell disruption with French press worked quite well but is very time consuming. Milling is assumed to decrease the molecular weight (polymer chain length). Pigment removal turned out to be necessary prior to PHB extraction, as pigments influenced the PHB properties negatively. This process step can be of advantage due to the generation of phycocyanin as a valuable side product [107]. A mixture of acetone and ethanol (70:30) was most suitable for this purpose. PHB extraction was performed with hot chloroform via a soxhlet extractor. Figure 5 compares the necessary processing steps of heterotrophs and cyanobacteria.
The PHB analysis showed that the polymers extracted from cyanobacterial biomass are comparable to commercially available PHB. Furthermore, it was shown that not only did the nutrient source, but also biomass pre-treatment and the method of polymer extraction influence the PHB properties. Pigment extraction and sample pre-treatment increased the average molecular weight (Mw) from 0.3 to 1.4 MD, but decreased degradation temperatures and crystallinity from 282 °C to 275 °C (Tonset) and from 296 °C to 287 °C (Tmax), respectively. The Mw ranged from 5.8 to 8.0 MD, by using mineral medium and digestate, respectively. The thermal properties (Tonset: 283–282 °C; Tmax: 293–292 °C), which are important for processing the polymer, are only slightly influenced by the nutrient source and are lower than, but comparable to, commercially available PHB. The crystallinity, responsible for higher final brittleness of the products, is about 17% lower than commercially available PHA.
5.6. Contaminations
Contaminations in non-sterile mass cultivation of microalgae are inevitable. It is only a matter of time before first contaminations appear, whether cultivation is done in open ponds or closed photobioreactors [108]. We observed certain bacterial and fungal contaminations with minor effects on Synechocystis salina CCALA192, when using CO2 as the sole carbon source. Though, when adding acetate to the medium fungal contaminations were prevalent and difficult to control. After one and a half years the pilot plant was revised and a new degassing system was installed. From this moment on rapid culture crashes occurred. The microscopic image revealed a ciliated protozoa ingesting Synechocystis rapidly.
This ciliate forms highly resistant cysts and it is assumed that cysts from the surrounding soil were brought into the system during the revision work. Facts about contaminations in mass cultivation of Synechocystis are scarcely reported. Touloupakis and colleagues reported the grazing of Synechocystis PCC6803 by golden algae Poterioochromonas sp. [109]. High pH values of 10 and above helped to control the contaminant and maintain a stable culture. Unfortunately, the ciliate in our cultures survives those high pH values. Thoroughly cleaning and sanitizing the photobioreactor brought some success, but the ciliate is still occurring and leading to culture crashes. Due to the ciliate’s capability to form cysts, it is very difficult to completely eliminate it from the reactor. Heat sterilization is not possible in tubular photobioreactors. Addressing further research, there is need for special cultivation methods for robustly growing Synechocystis in non-sterile environment.
6. Conclusions
Although not economical today, the idea of a sustainable PHB production with cyanobacteria, CO2, and sunlight is still attractive and, more and more, researchers are working in this field. The main challenges today are similar to biofuel production with green algae: (i) realization of efficient low-cost cultivation systems at large scale; (ii) maintaining stable cultures under non-sterile conditions; (iii) increasing the total productivity and yield; and (iv) economic downstream processing and utilization of the residual biomass.
Looking for suitable production strains it must be considered that PHB production is a very common feature of many, but not all, cyanobacteria. The PHB content of cyanobacteria is highly strain specific, as strains of the same genus were reported with highly varying PHB contents. In addition to the PHB content the growth rate and robustness of a strain is particularly important. The only cyanobacterium cultivated in mass cultivation today is Arthrospira sp. and, therefore, one of the most promising candidates for photoautotrophic PHB production, although most Arthrospira sp. strains still show little PHB content.
Heterotrophic cultivation with acetate boosts the PHB content remarkably, as most reported values over 30% were achieved this way. However, it needs to be considered that using an organic carbon source impairs the most attractive feature of cyanobacteria, converting CO2 to PHB. Using organic sources will also complicate non-sterile mass cultivation and could easily lead to contaminations and culture crashes. PHB production with organic carbon sources should be performed with heterotrophic bacteria, as their PHB productivity, as well as their cell density, are 10–100 times higher.
Nitrogen and phosphorous depletion are the most important factors to increase the PHB content and are often necessary to produce any PHB at all. Therefore, a two-stage cultivation with a self-limiting medium is necessary for large-scale photoautotrophic PHB production. With this strategy PHB was successfully produced in our 200 L photobioreactor. In tubular systems small unicellular organisms, like Synechocystis sp., are preferred over filamentous organisms, mainly because of the shear stress of the pump. Considering all of the difficulties to overcome, establishing a stable cyanobacterial culture is most important and most difficult to achieve.
Acknowledgments
The research project CO2USE was thankfully financed by the Austrian climate and energy fund and FFG (Austrian research promotion agency).
Author Contributions
Clemens Troschl designed and performed experiments, analyzed data, and wrote about 75% of the article. Katharina Meixner designed and performed experiments, analyzed data and wrote about 15% of the article. Bernhard Drosg supervised the work, analyzed data, and wrote about 10% of the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khosravi-Darani, K.; Mokhtari, Z.-B.B.; Amai, T.; Tanaka, K. Microbial production of poly(hydroxybutyrate) from C1 carbon sources. Appl. Microbiol. Biotechnol. 2013, 97, 1407–1424. [Google Scholar] [CrossRef] [PubMed]
- Dawes, E. Polyhydroxybutyrate: An intriguing biopolymer. Biosci. Rep. 1988, 8, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-Q. A microbial polyhydroxyalkanoates (PHA) based bio- and materials industry. Chem. Soc. Rev. 2009, 38, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Halami, P.M. Production of polyhydroxyalkanoate from starch by the native isolate Bacillus cereus CFR06. World J. Microbiol. Biotechnol. 2008, 24, 805–812. [Google Scholar] [CrossRef]
- Ting, C.S.; Rocap, G.; King, J.; Chisholm, S.W. Cyanobacterial photosynthesis in the oceans: The origins and significance of divergent light-harvesting strategies. Trends Microbiol. 2002, 10, 134–142. [Google Scholar] [CrossRef]
- IPCC. 2014: Climate Change 2014: Synthesis Report; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Stanier, R.J.; Cohen-Bazire, G. Phototrophic prokaryotes: The cyanobacteria. Ann. Rev. Microbiol. 1977, 31, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Bekker, A.; Holland, H.D.; Wang, P.-L.; Rumble, D.; Stein, H.J.; Hannah, J.L.; Coetzee, L.L.; Beukes, N.J. Dating the rise of atmospheric oxygen. Nature 2004, 427, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Schirrmeister, B.E.; Sanchez-Baracaldo, P.; Wacey, D. Cyanobacterial evolution during the Precambrian. Int. J. Astrobiol. 2016, 15, 1–18. [Google Scholar] [CrossRef]
- Fischer, W.W.; Hemp, J.; Johnson, J.E. Evolution of Oxygenic Photosynthesis. Annu. Rev. Earth Planet. Sci. 2016, 44, 647–683. [Google Scholar] [CrossRef]
- Nabout, J.C.; da Silva Rocha, B.; Carneiro, F.M.; Sant’Anna, C.L. How many species of Cyanobacteria are there? Using a discovery curve to predict the species number. Biodivers. Conserv. 2013, 22, 2907–2918. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Sistrom, W.R.; Hansen, T.A.; Whitton, B.A.; Castenholz, R.W.; Pfennig, N.; Gorlenko, V.N.; Kondratieva, E.N.; Eimhjellen, K.E.; Whittenbury, R.; et al. Proposal to Place the Nomenclature of the Cyanobacteria (Blue-Green Algae) Under the Rules of the International Code of Nomenclature of Bacteria. Int. J. Syst. Bacteriol. 1978, 28, 335–336. [Google Scholar] [CrossRef]
- McNeill, J.; Barrie, F.R. International Code of Nomenclature for Algae, Fungi, and Plants (Melbourne Code); Koeltz Scientific Books: Oberreifenberg, Germany, 2012. [Google Scholar]
- Parker, C.T.; Tindall, B.J.; Garrity, G.M. International Code of Nomenclature of Prokaryotes. Int. J. Syst. Evol. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Handbook of Microalgal Culture, 1st ed.; Blackwell Science Ltd.: Oxford, UK, 2004. [Google Scholar]
- Tan, G.-Y.; Chen, C.-L.; Li, L.; Ge, L.; Wang, L.; Razaad, I.; Li, Y.; Zhao, L.; Mo, Y.; Wang, J.-Y. Start a Research on Biopolymer Polyhydroxyalkanoate (PHA): A Review. Polymers (Basel.) 2014, 6, 706–754. [Google Scholar] [CrossRef]
- Nakaya, Y.; Iijima, H.; Takanobu, J.; Watanabe, A.; Hirai, M.Y.; Osanai, T. One day of nitrogen starvation reveals the effect of sigE and rre37 overexpression on the expression of genes related to carbon and nitrogen metabolism in Synechocystis sp. PCC 6803. J. Biosci. Bioeng. 2015, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schlebusch, M.; Forchhammer, K. Requirement of the nitrogen starvation-induced protein s110783 for polyhydroxybutyrate accumulation in Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2010, 76, 6101–6107. [Google Scholar] [CrossRef] [PubMed]
- Hauf, W.; Schlebusch, M.; Hüge, J.; Kopka, J.; Hagemann, M.; Forchhammer, K. Metabolic Changes in Synechocystis PCC6803 upon Nitrogen-Starvation: Excess NADPH Sustains Polyhydroxybutyrate Accumulation. Metabolites 2013, 3, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Stal, L.J. Poly(hydroxyalkanoate) in cyanobacteria: An overview. FEMS Microbiol. Lett. 1992, 103, 169–180. [Google Scholar] [CrossRef]
- Kaneko, T.; Sato, S.; Kotani, H.; Tanaka, A.; Asamizu, E.; Nakamura, Y.; Miyajima, N.; Hirosawa, M.; Sugiura, M.; Sasamoto, S.; et al. Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions. DNA Res. 1996, 3, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Hein, S.; Tran, H.; Steinbüchel, A. Synechocystis sp. PCC6803 possesses a two-component polyhydroxyalkanoic acid synthase similar to that of anoxygenic purple sulfur bacteria. Arch. Microbiol. 1998, 170, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Taroncher-Oldenburg, G.; Nishina, K.; Stephanopoulos, G. Identification and analysis of the polyhydroxyalkanoate-specific beta-ketothiolase and acetoacetyl coenzyme A reductase genes in the cyanobacterium Synechocystis sp. strain PCC6803. Appl. Environ. Microbiol. 2000, 66, 4440–4448. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, S.; Izumi, Y.; Matsuda, F.; Hasunuma, T.; Chang, J.S.; Kondo, A. Synergistic enhancement of glycogen production in Arthrospira platensis by optimization of light intensity and nitrate supply. Bioresour. Technol. 2012, 108, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Monshupanee, T.; Incharoensakdi, A. Enhanced accumulation of glycogen, lipids and polyhydroxybutyrate under optimal nutrients and light intensities in the cyanobacterium Synechocystis sp. PCC 6803. J. Appl. Microbiol. 2014, 116, 830–838. [Google Scholar] [CrossRef] [PubMed]
- De Philippis, R.; Sili, C.; Vincenzini, M. Glycogen and poly-β-hydroxybutyrate synthesis in Spirulina maxima. J. Gen. Microbiol. 1992, 138, 1623–1628. [Google Scholar] [CrossRef]
- Deschoenmaeker, F.; Facchini, R.; Carlos, J.; Pino, C.; Bayon-Vicente, G.; Sachdeva, N.; Flammang, P.; Wattiez, R. Nitrogen depletion in Arthrospira sp. PCC 8005, an ultrastructural point of view. J. Struct. Biol. 2016, 196, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, S.; Nishida, A.; Ho, S.-H.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Glycogen production for biofuels by the euryhaline cyanobacteria Synechococcus sp. strain PCC 7002 from an oceanic environment. Biotechnol. Biofuels 2014, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.H.; Keppel, C.; Spalding, M.; Jane, J.L. Effects of growth condition on the structure of glycogen produced in cyanobacterium Synechocystis sp. PCC6803. Int. J. Biol. Macromol. 2007, 40, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Ostle, A.G.; Holt, J.G. Fluorescent Stain for Poly-3-Hydroxybutyrate. Appl. Environ. Microbiol. 1982, 44, 238–241. [Google Scholar] [PubMed]
- Gorenflo, V.; Steinbüchel, A.; Marose, S.; Rieseberg, M.; Scheper, T. Quantification of bacterial polyhydroxyalkanoic acids by Nile red staining. Appl. Microbiol. Biotechnol. 1999, 51, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Tsang, T.K.; Roberson, R.W.; Vermaas, W.F.J. Polyhydroxybutyrate particles in Synechocystis sp. PCC 6803: Facts and fiction. Photosynth. Res. 2013, 118, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Hauf, W.; Watzer, B.; Roos, N.; Klotz, A.; Forchhammer, K. Photoautotrophic Polyhydroxybutyrate Granule Formation Is Regulated by Cyanobacterial Phasin PhaP in Synechocystis sp. Strain PCC 6803. Appl. Environ. Microbiol. 2015, 81, 4411–4422. [Google Scholar] [CrossRef] [PubMed]
- Klotz, A.; Georg, J.; Bučínská, L.; Watanabe, S.; Reimann, V.; Januszewski, W.; Sobotka, R.; Jendrossek, D.; Hess, W.R.; Forchhammer, K. Awakening of a dormant cyanobacterium from nitrogen chlorosis reveals a genetically determined program. Curr. Biol. 2016, 26, 2862–2872. [Google Scholar] [CrossRef] [PubMed]
- Gründel, M.; Scheunemann, R.; Lockau, W.; Zilliges, Y. Impaired glycogen synthesis causes metabolic overflow reactions and affects stress responses in the cyanobacterium Synechocystis sp. PCC 6803. Microbiol. (UK) 2012, 158, 3032–3043. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Knoop, H.; Axmann, I.M.; Steuer, R. The diversity of cyanobacterial metabolism: Genome analysis of multiple phototrophic microorganisms. BMC Genom. 2012, 13, 56. [Google Scholar] [CrossRef] [PubMed]
- Ansari, S.; Fatma, T. Cyanobacterial polyhydroxybutyrate (PHB): Screening, optimization and characterization. PLoS ONE 2016, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kaewbai-ngam, A.; Incharoensakdi, A.; Monshupanee, T. Increased accumulation of polyhydroxybutyrate in divergent cyanobacteria under nutrient-deprived photoautotrophy: An efficient conversion of solar energy and carbon dioxide to polyhydroxybutyrate by Calothrix scytonemicola TISTR 8095. Bioresour. Technol. 2016, 212, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- de Jaeger, L.; Verbeek, R.E.; Draaisma, R.B.; Martens, D.E.; Springer, J.; Eggink, G.; Wijffels, R.H. Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus: (I) Mutant generation and characterization. Biotechnol. Biofuels 2014, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Damrow, R.; Maldener, I.; Zilliges, Y. The multiple functions of common microbial carbon polymers, glycogen and PHB, during stress responses in the non-diazotrophic cyanobacterium Synechocystis sp. PCC 6803. Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.M.; Smith, A.J. Nitrogen chlorosis in blue-green algae. Arch. für Mikrobiol. 1969, 69, 114–120. [Google Scholar] [CrossRef]
- Gorl, M.; Sauer, J.; Baier, T.; Forchhammer, K. Nitrogen-starvation-induced chlorosis in Synechococcus PCC 7942: Adaptation to long-term survival. Microbiology 1998, 144, 2449–2458. [Google Scholar] [CrossRef] [PubMed]
- Sauer, J.; Schreiber, U.; Schmid, R.; Völker, U.; Forchhammer, K. Nitrogen starvation-induced chlorosis in Synechococcus PCC 7942. Low-level photosynthesis as a mechanism of long-term survival. Plant Physiol. 2001, 126, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.F.; Wu, Q.Y.; Shen, Z.Y. Accumulation of poly-beta-hydroxybutyrate in cyanobacterium Synechocystis sp. PCC6803. Bioresour. Technol. 2001, 76, 85–90. [Google Scholar] [CrossRef]
- Panda, B.; Mallick, N. Enhanced poly-β-hydroxybutyrate accumulation in a unicellular cyanobacterium, Synechocystis sp. PCC 6803. Lett. Appl. Microbiol. 2007, 44, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Khetkorn, W.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Enhancement of poly-3-hydroxybutyrate production in Synechocystis sp. PCC 6803 by overexpression of its native biosynthetic genes. Bioresour. Technol. 2016, 214, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, M.; Nakai, K.; Miyake, M.; Asada, Y.; Taya, M. Production of poly-β-hydroxybutyrate by thermophilic cyanobacterium, Synechococcus sp. MA19, under phosphate-limited conditions. Biotechnol. Lett. 2001, 23, 1095–1099. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, L.; Miron, A.; Klimova, B.; Wan, D.; Kuca, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Shimamatsu, H. Mass production of Spirulina, an edible microalga. Hydrobiologia 2004, 512, 39–44. [Google Scholar] [CrossRef]
- Choi, S.-L.; Suh, I.S.; Lee, C.-G. Lumostatic operation of bubble column photobioreactors for Haematococcus pluvialis cultures using a specific light uptake rate as a control parameter. Enzyme Microb. Technol. 2003, 33, 403–409. [Google Scholar] [CrossRef]
- Lu, Y.M.; Xiang, W.Z.; Wen, Y.H. Spirulina (Arthrospira) industry in Inner Mongolia of China: Current status and prospects. J. Appl. Phycol. 2011, 23, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Stevens, S.E.; Balkwill, D.L. Accumulation of poly-beta-hydroxybutyrate in Spirulina platensis. J. Bacteriol. 1982, 149, 361–363. [Google Scholar] [PubMed]
- Vincenzini, M.; Sili, C.; de Philippis, R.; Ena, A.; Materassi, R. Occurrence of poly-beta-hydroxybutyrate in Spirulina species. J. Bacteriol. 1990, 172, 2791–2792. [Google Scholar] [CrossRef] [PubMed]
- Panda, B.; Sharma, L.; Mallick, N. Poly-β-hydroxybutyrate accumulation in Nostoc muscorum and Spirulina platensis under phosphate limitation. J. Plant Physiol. 2005, 162, 1376–1379. [Google Scholar] [CrossRef] [PubMed]
- Shrivastav, A.; Mishra, S.K.; Mishra, S. Polyhydroxyalkanoate (PHA) synthesis by Spirulina subsalsa from Gujarat coast of India. Int. J. Biol. Macromol. 2010, 46, 255–260. [Google Scholar] [CrossRef] [PubMed]
- De Morais, M.G.; Stillings, C.; Roland, D.; Rudisile, M.; Pranke, P.; Costa, J.A.V.; Wendorff, J. Extraction of poly(3-hydroxybutyrate) from Spirulina LEB 18 for developing nanofibers. Polímeros 2015, 25, 161–167. [Google Scholar] [CrossRef]
- De Morais, M.G.; Stillings, C.; Dersch, R.; Rudisile, M.; Pranke, P.; Costa, J.A.V.; Wendorff, J. Biofunctionalized nanofibers using Arthrospira (spirulina) biomass and biopolymer. Biomed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K.; Gudder, D.A.; Mollenhauer, D. Review the Ecology of Nostoc. J. Phycol. 1995, 18, 2–18. [Google Scholar] [CrossRef]
- Qiu, B.; Liu, J.; Liu, Z.; Liu, S. Distribution and ecology of the edible cyanobacterium Ge-Xian-Mi (Nostoc) in rice fields of Hefeng County in China. J. Appl. Phycol. 2002, 14, 423–429. [Google Scholar] [CrossRef]
- Sharma, L.; Mallick, N. Accumulation of poly-β-hydroxybutyrate in Nostoc muscorum: Regulation by pH, light-dark cycles, N and P status and carbon sources. Bioresour. Technol. 2005, 96, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Mallick, N. Enhancement of poly-β-hydroxybutyrate accumulation in Nostoc muscorum under mixotrophy, chemoheterotrophy and limitations of gas-exchange. Biotechnol. Lett. 2005, 27, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Bhati, R.; Mallick, N. Production and characterization of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) co-polymer by a N 2-fixing cyanobacterium, Nostoc muscorum Agardh. J. Chem. Technol. Biotechnol. 2012, 87, 505–512. [Google Scholar] [CrossRef]
- Bhati, R.; Mallick, N. Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymer production by the diazotrophic cyanobacterium Nostoc muscorum Agardh: Process optimization and polymer characterization. Algal Res. 2015, 7, 78–85. [Google Scholar] [CrossRef]
- Bhati, R.; Mallick, N. Carbon dioxide and poultry waste utilization for production of polyhydroxyalkanoate biopolymers by Nostoc muscorum Agardh: A sustainable approach. J. Appl. Phycol. 2016, 28, 161–168. [Google Scholar] [CrossRef]
- Samantaray, S.; Mallick, N. Production and characterization of poly-β-hydroxybutyrate (PHB) polymer from Aulosira fertilissima. J. Appl. Phycol. 2012, 24, 803–814. [Google Scholar] [CrossRef]
- Lama, L.; Nicolaus, B.; Calandrelli, V.; Manca, M.C.; Romana, I.; Gambacorta, A. Effect of growth conditions on endo- and exopolymer biosynthesis in Anabaena cylindrical 10 C. Phytochemistry 1996, 42, 655–659. [Google Scholar] [CrossRef]
- Gopi, K.; Balaji, S.; Muthuvelan, B. Isolation Purification and Screening of Biodegradable Polymer PHB Producing Cyanobacteria from Marine and Fresh Water Resources. Iran. J. Energy Environ. 2014, 5, 94–100. [Google Scholar] [CrossRef]
- Benemann, J. Microalgae for biofuels and animal feeds. Energies 2013, 6, 5869–5886. [Google Scholar] [CrossRef]
- Dineshbabu, G.; Uma, V.S.; Mathimani, T.; Deviram, G.; Arul Ananth, D.; Prabaharan, D.; Uma, L. On-site concurrent carbon dioxide sequestration from flue gas and calcite formation in ossein effluent by a marine cyanobacterium Phormidium valderianum BDU 20041. Energy Convers. Manag. 2016, in press. [Google Scholar] [CrossRef]
- Chen, H.W.; Yang, T.S.; Chen, M.J.; Chang, Y.C.; Lin, C.Y.; Wang, E.I.C.; Ho, C.L.; Huang, K.M.; Yu, C.C.; Yang, F.L.; et al. Application of power plant flue gas in a photobioreactor to grow Spirulina algae, and a bioactivity analysis of the algal water-soluble polysaccharides. Bioresour. Technol. 2012, 120, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Kumar, A.; Pathak, A.K.; Guria, C. Carbon dioxide assisted Spirulina platensis cultivation using NPK-10:26:26 complex fertilizer in sintered disk chromatographic glass bubble column. J. CO2 Util. 2014, 8, 49–59. [Google Scholar] [CrossRef]
- He, L.; Subramanian, V.R.; Tang, Y.J. Experimental analysis and model-based optimization of microalgae growth in photo-bioreactors using flue gas. Biomass Bioenergy 2012, 41, 131–138. [Google Scholar] [CrossRef]
- Ferreira, L.S.; Rodrigues, M.S.; Converti, A.; Sato, S.; Carvalho, J.C.M. Arthrospira (spirulina) platensis cultivation in tubular photobioreactor: Use of no-cost CO2 from ethanol fermentation. Appl. Energy 2012, 92, 379–385. [Google Scholar] [CrossRef]
- Sumardiono, S.; Syaichurrozi, I.; Budi Sasongko, S. Utilization of Biogas as Carbon Dioxide Provider for Spirulina platensis Culture. Curr. Res. J. Biol. Sci. 2014, 6, 53–59. [Google Scholar]
- Cuellar-Bermudez, S.P.; Aleman-Nava, G.S.; Chandra, R.; Garcia-Perez, J.S.; Contreras-Angulo, J.R.; Markou, G.; Muylaert, K.; Rittmann, B.E.; Parra-Saldivar, R. Nutrients utilization and contaminants removal. A review of two approaches of algae and cyanobacteria in wastewater. Algal Res. 2016. [Google Scholar] [CrossRef]
- Markou, G.; Vandamme, D.; Muylaert, K. Microalgal and cyanobacterial cultivation: The supply of nutrients. Water Res. 2014, 65, 186–202. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Mishra, S.K.; Shrivastav, A.; Park, M.S.; Yang, J.W. Recent trends in the mass cultivation of algae in raceway ponds. Renew. Sustain. Energy Rev. 2015, 51, 875–885. [Google Scholar] [CrossRef]
- Markou, G.; Georgakakis, D. Cultivation of filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: A review. Appl. Energy 2011, 88, 3389–3401. [Google Scholar] [CrossRef]
- Chaiklahan, R.; Chirasuwan, N.; Siangdung, W.; Paithoonrangsarid, K.; Bunnag, B. Cultivation of spirulina platensis using pig wastewater in a semi-continuous process. J. Microbiol. Biotechnol. 2010, 20, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Cicci, A.; Bravi, M. Production of the freshwater microalgae scenedesmus dimorphus and arthrospira platensis by using cattle digestate. Chem. Eng. Trans. 2014, 38, 85–90. [Google Scholar] [CrossRef]
- Fouilland, E.; Vasseur, C.; Leboulanger, C.; Le Floc’h, E.; Carré, C.; Marty, B.; Steyer, J.-P.P.; Sialve, B. Coupling algal biomass production and anaerobic digestion: Production assessment of some native temperate and tropical microalgae. Biomass Bioenergy 2014, 70, 564–569. [Google Scholar] [CrossRef]
- Markou, G.; Chatzipavlidis, I.; Georgakakis, D. Cultivation of Arthrospira (Spirulina) platensis in olive-oil mill wastewater treated with sodium hypochlorite. Bioresour. Technol. 2012, 112, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Olguín, E.J.; Galicia, S.; Mercado, G.; Pérez, T. Annual productivity of Spirulina (Arthrospira) and nutrient removal in a pig wastewater recycling process under tropical conditions. J. Appl. Phycol. 2003, 15, 249–257. [Google Scholar] [CrossRef]
- Prajapati, S.K.; Kumar, P.; Malik, A.; Vijay, V.K. Bioconversion of algae to methane and subsequent utilization of digestate for algae cultivation: A closed loop bioenergy generation process. Bioresour. Technol. 2014, 158, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Daelman, M.R.J.; Sorokin, D.; Kruse, O.; van Loosdrecht, M.C.M.; Strous, M. Haloalkaline Bioconversions for Methane Production from Microalgae Grown on Sunlight. Trends Biotechnol. 2016, 34, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Nolla-Ardevol, V.; Strous, M.; Tegetmeyer, H.E.E. Anaerobic digestion of the microalga Spirulina at extreme alkaline conditions: Biogas production, metagenome and metatranscriptome. Front. Microbiol. 2015, 6, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Schideman, L.; Yu, G.; Zhang, Y. A synergistic combination of algal wastewater treatment and hydrothermal biofuel production maximized by nutrient and carbon recycling. Energy Environ. Sci. 2013, 6, 3765. [Google Scholar] [CrossRef]
- Zheng, M.; Schideman, L.C.; Tommaso, G.; Chen, W.-T.; Zhou, Y.; Nair, K.; Qian, W.; Zhang, Y.; Wang, K. Anaerobic digestion of wastewater generated from the hydrothermal liquefaction of Spirulina: Toxicity assessment and minimization. Energy Convers. Manag. 2016, in press. [Google Scholar] [CrossRef]
- Depraetere, O.; Pierre, G.; Noppe, W.; Vandamme, D.; Foubert, I.; Michaud, P.; Muylaert, K. Influence of culture medium recycling on the performance of Arthrospira platensis cultures. Algal Res. 2015, 10, 48–54. [Google Scholar] [CrossRef]
- Meixner, K.; Fritz, I.; Daffert, C.; Markl, K.; Fuchs, W.; Drosg, B. Processing recommendations for using low-solids digestate as nutrient solution for production with Synechocystis salina. J. Biotechnol. 2016, 240, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Gopi, K.; Muthuvelan, B. A review on production of poly-b-hydroxybutyrates from cyanobacteria for the production of bio plastics. Algal Res. 2013, 2, 278–285. [Google Scholar] [CrossRef]
- Marcilhac, C.; Sialve, B.; Pourcher, A.M.; Ziebal, C.; Bernet, N.; Béline, F. Digestate color and light intensity affect nutrient removal and competition phenomena in a microalgal-bacterial ecosystem. Water Res. 2014, 64, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.R.; Costa, J.A.V. Mixotrophic cultivation of microalga Spirulina platensis using molasses as organic substrate. Aquaculture 2007, 264, 130–134. [Google Scholar] [CrossRef]
- Phang, S.M.; Miah, M.S.; Yeoh, B.G.; Hashim, M.A. Spirulina cultivation in digested sago starch factory wastewater. J. Appl. Phycol. 2000, 12, 395–400. [Google Scholar] [CrossRef]
- Massa, M.; Buono, S.; Langellotti, A.L.; Castaldo, L.; Martello, A.; Paduano, A.; Sacchi, R.; Fogliano, V. Evaluation of anaerobic digestates from different feedstocks as growth media for Tetradesmus obliquus, Botryococcus braunii, Phaeodactylum tricornutum and Arthrospira maxima. New Biotechnol. 2017, 36, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Ge, X.; Park, S.Y.; Li, Y. Comparison of Synechocystis sp. PCC6803 and Nannochloropsis salina for lipid production using artificial seawater and nutrients from anaerobic digestion effluent. Bioresour. Technol. 2013, 144, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Molina, E.; Fernández, J.; Acién, F.G.; Chisti, Y. Tubular photobioreactor design for algal cultures. J. Biotechnol. 2001, 92, 113–131. [Google Scholar] [CrossRef]
- Acién Fernández, F.G.; Fernández Sevilla, J.M.; Sánchez Pérez, J.A.; Molina Grima, E.; Chisti, Y. Airlift-driven external-loop tubular photobioreactors for outdoor production of microalgae: Assessment of design and performance. Chem. Eng. Sci. 2001, 56, 2721–2732. [Google Scholar] [CrossRef]
- Rabensteiner, M.; Kinger, G.; Koller, M.; Gronald, G.; Hochenauer, C. Pilot plant study of ethylenediamine as a solvent for post combustion carbon dioxide capture and comparison to monoethanolamine. Int. J. Greenh. Gas Control 2014, 27, 1–14. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Drosg, B.; Fritz, I.; Gattermayr, F.; Silvestrini, L. Photo-autotrophic Production of Poly(hydroxyalkanoates) in Cyanobacteria. Chem. Biochem. Eng. Q. 2015, 29, 145–156. [Google Scholar] [CrossRef]
- Michels, M.H.A.; van der Goot, A.J.; Vermuë, M.H.; Wijffels, R.H. Cultivation of shear stress sensitive and tolerant microalgal species in a tubular photobioreactor equipped with a centrifugal pump. J. Appl. Phycol. 2016, 28, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Milledge, J.J.; Heaven, S. A review of the harvesting of micro-algae for biofuel production. Rev. Environ. Sci. Biotechnol. 2013, 12, 165–178. [Google Scholar] [CrossRef]
- Heinrich, D.; Madkour, M.H.; Al-Ghamdi, M.; Shabbaj, I.I.; Steinbüchel, A. Large scale extraction of poly(3-hydroxybutyrate) from Ralstonia eutropha H16 using sodium hypochlorite. AMB Express 2012, 2, 59. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.; Acién, F.G.; Fernández-Sevilla, J.M.; González, C.V.; Bermejo, R. Development of a process for large-scale purification of C-phycocyanin from Synechocystis aquatilis using expanded bed adsorption chromatography. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, W.; Chen, L.; Wang, J.; Liu, T. The contamination and control of biological pollutants in mass cultivation of microalgae. Bioresour. Technol. 2013, 128, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Touloupakis, E.; Cicchi, B.; Benavides, A.M.S.; Torzillo, G. Effect of high pH on growth of Synechocystis sp. PCC 6803 cultures and their contamination by golden algae (Poterioochromonas sp.). Appl. Microbiol. Biotechnol. 2015, 100, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Microscopic photographs of different cyanobacterial species made in DIC (differential interference contrast). (A) Synechocystis sp.; (B) Cyanosarcina sp.; (C) Calothrix sp.; and (D) Arthrospira sp.
Figure 1.
Microscopic photographs of different cyanobacterial species made in DIC (differential interference contrast). (A) Synechocystis sp.; (B) Cyanosarcina sp.; (C) Calothrix sp.; and (D) Arthrospira sp.
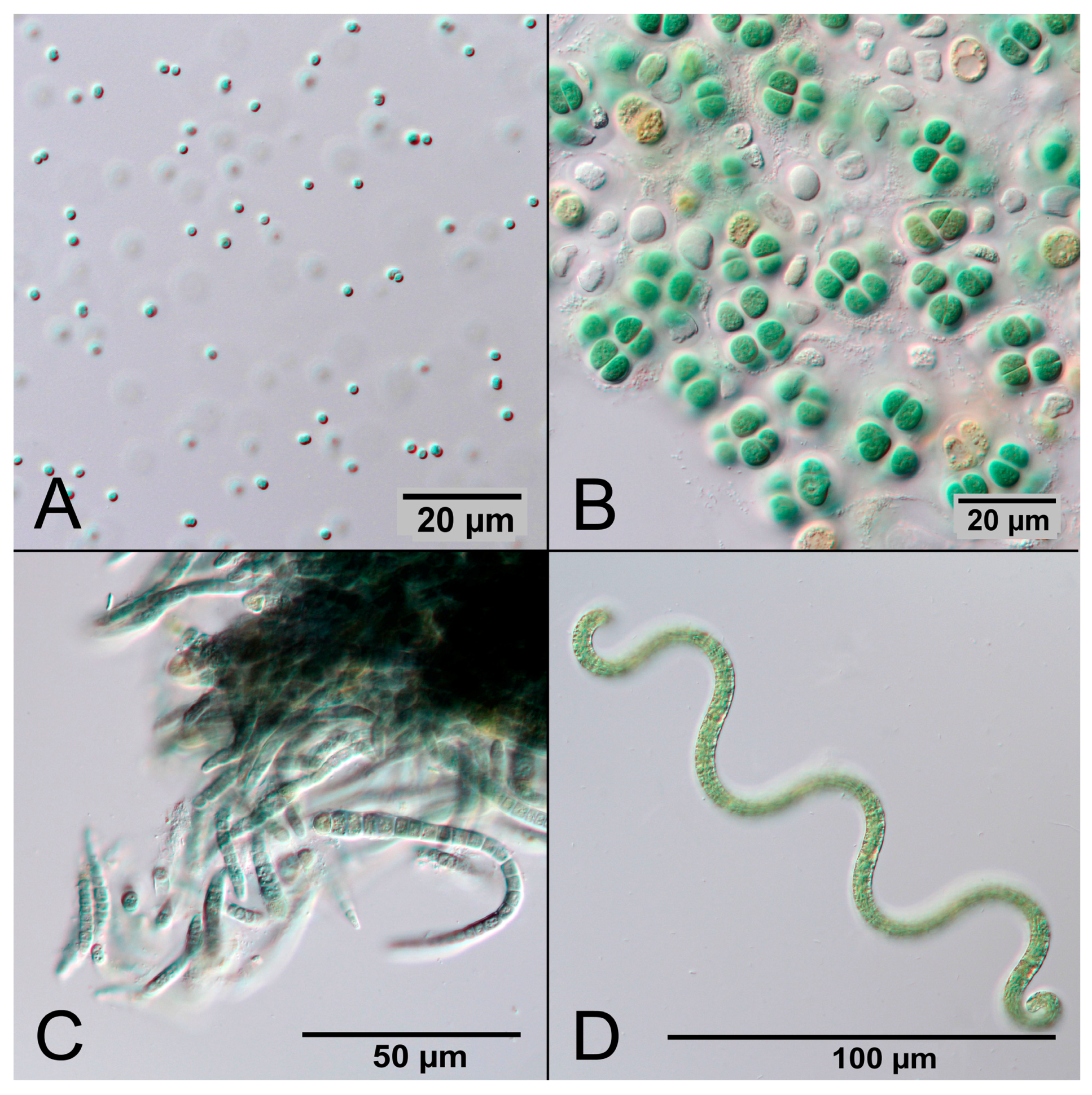
Figure 2.
Two-hundred litre tubular photobioreactor with Synechocystis salina CCALA192. The central tower serves as a degasser. The centrifugal pump is situated at the lowest point of the reactor on the left side.
Figure 2.
Two-hundred litre tubular photobioreactor with Synechocystis salina CCALA192. The central tower serves as a degasser. The centrifugal pump is situated at the lowest point of the reactor on the left side.

Figure 3.
PI-controlled pH value. The setpoint of the pH is 8.5. Lamps turn on at 02:00 and turn off at 22:00, causing a rise and decrease of the pH value due to CO2 consumption. In total, 59 L (118 g) of CO2 were consumed on this day.
Figure 3.
PI-controlled pH value. The setpoint of the pH is 8.5. Lamps turn on at 02:00 and turn off at 22:00, causing a rise and decrease of the pH value due to CO2 consumption. In total, 59 L (118 g) of CO2 were consumed on this day.
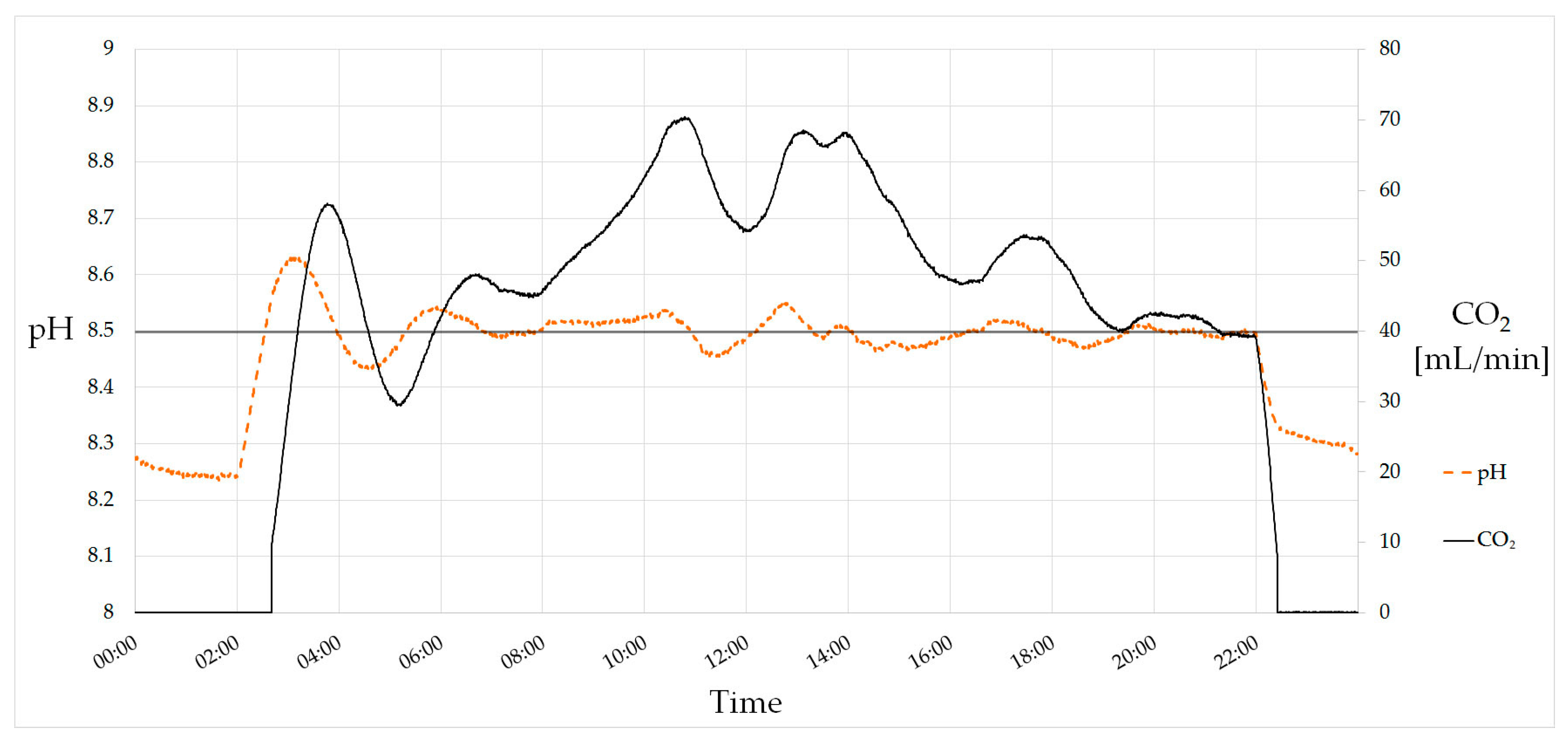
Figure 4.
Biomass [g/L] and PHB [g/L] concentration of Synechocystis salina using digestate supernatant as nutrient source (Trial 5).
Figure 4.
Biomass [g/L] and PHB [g/L] concentration of Synechocystis salina using digestate supernatant as nutrient source (Trial 5).
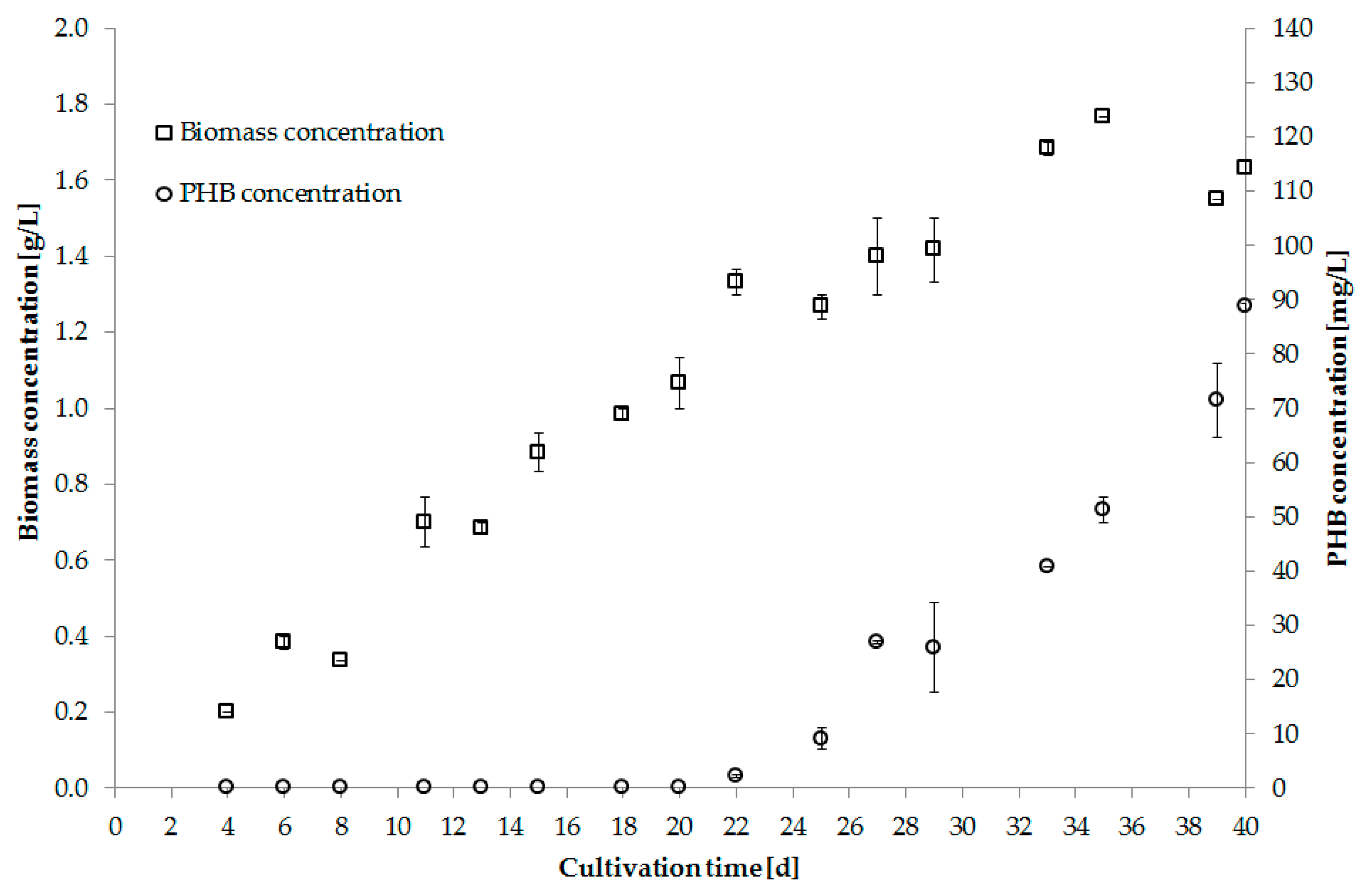
Figure 5.
Comparison of processing steps needed to extract PHA from heterotrophic bacteria and cyanobacteria.
Figure 5.
Comparison of processing steps needed to extract PHA from heterotrophic bacteria and cyanobacteria.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Synechocystis and Synechococcus as PHA producers. (cdw = cell dry weight, n.r. = not reported).
Table 1.
Synechocystis and Synechococcus as PHA producers. (cdw = cell dry weight, n.r. = not reported).
| Carbon Source | Cyanobacterium | Culture Condition | %PHA of cdw | PHA Composition | Total cdw | Reference |
|---|---|---|---|---|---|---|
| Photoautotrophic | Synechocystis PCC6803 | Photoautotrophic, nitrogen lim. | 4.1% | PHB | 0.65 g/L | [45] |
| Synechocystis PCC6803 | Photoautotrophic, nitrogen lim. | 9.5% | PHB | n.r. | [46] | |
| Synechocystis PCC6803 | Photoautotrophic, phosphate lim. | 11.2% | PHB | n.r. | [46] | |
| Synechocystis PCC6803 (recombinant) | Photoautotrophic, nitrogen lim. | 26% | PHB | n.r. | [47] | |
| Synechococcus MA19 | Photoautotrophic, phosphate lim., 50 °C | 55% | PHB | 4.4 g/L | [48] | |
| Heterotrophic | Synechocystis PCC6803 | Acetate + Fructose supplementation | 38% | PHB | n.r. | [46] |
| Synechocystis PCC6803 (recombinant) | Acetate supplementation | 35% | PHB | n.r. | [47] |
Table 2.
Arthrospira as a PHA producer. (cdw = cell dry weight, n.r. = not reported).
| Carbon Source | Cyanobacterium | Culture Condition | %PHA of cdw | PHA Composition | Total cdw | Reference |
|---|---|---|---|---|---|---|
| Photoautotrophic | Arthrospira platensis | Photoautotrophic | 6% | PHB | n.r. | [54] |
| Arthrospira sp. | Photoautotrophic | <1% | PHB | n.r. | [55] | |
| Arthrospira platensis | Photoautotrophic, phosphate lim. | 3.5% | PHB | 0.3 g/L | [56] | |
| Arthrospira subsalsa | Photoautotrophic, nitrogen lim. | 14.7% | PHB | 1.97 g/L | [57] | |
| Arthrospira platensis | n.r. | 22% | PHB | n.r. | [59] | |
| Heterotrophic | Arthrospira maxima | Acetate + CO2 | 5% | PHB | 1.4 g/L | [26] |
| Arthrospira sp. | Acetate + CO2 | 2.5% | PHB | n.r. | [55] |
Table 3.
Nostoc as a PHA producer. (cdw = cell dry weight, n.r. = not reported).
| Carbon Source | Cyanobacterium | Culture Condition | %PHA of cdw | PHA Composition | Total cdw | Reference |
|---|---|---|---|---|---|---|
| Photoautotrophic | Nostoc muscorum | Photoautotrophic, nitrogen and phosphorous lim. | 8.7% | PHB | n.r. | [62] |
| Nostoc muscorum agardh | Photoautotrophic, 10% CO2 | 22% | PHB | 1.1 g/L | [66] | |
| Nostoc muscorum | Photoautotrophic, nitrogen and phosphorous lim. | 22% | PHB | 0.13 g/L | [56] | |
| Heterotrophic | Nostoc muscorum agardh | Acetate, valerate, nitrogen lim. | 58% | P[3HB-co-3HV] | 0.29 g/L | [64] |
| Nostoc muscorum | Acetate, limited gas exchange | 40% | PHB | n.r. | [63] | |
| Nostoc muscorum agardh | Acetate, glucose, valerate, 10% CO2 | 70% | P[3HB-co-3HV] | 0.98 g/L | [66] | |
| Nostoc muscorum agardh | Acetate, glucose, valerate, nitrogen lim. | 78% | P[3HB-co-3HV] | 0.56 g/L | [65] | |
| Nostoc muscorum | Acetate, dark incubation, nitrogen and phosphorous lim. | 35% | PHB | n.r. | [62] |
Table 4.
Different cyanobacterial species as PHA producers. (cdw = cell dry weight, n.r. = not reported).
Table 4.
Different cyanobacterial species as PHA producers. (cdw = cell dry weight, n.r. = not reported).
| Carbon Source | Cyanobacterium | Culture Condition | %PHA of cdw | PHA Composition | Total cdw | Reference |
|---|---|---|---|---|---|---|
| Photoautotrophic | Phormidium sp. TISTR 8462 | Photoautotrophic, nitrogen lim. | 14.8% | PHB | n.r. | [38] |
| Oscillatoria jasorvensis TISTR 8980 | Photoautotrophic, nitrogen lim. | 15.7% | PHB | n.r. | [38] | |
| Calothrix scytonemicola TISTR 8095 | Photoautotrophic, nitrogen lim. | 25.2% | PHB | n.r. | [38] | |
| Anabaena sp. | Photoautotrophic | 2.3% | PHB | n.r. | [69] | |
| Aulosira fertilissima | Photoautotrophic, phosphorous lim. | 10% | PHB | n.r. | [67] | |
| Heterotrophic | Aulosira fertilissima | Acetate, phosphorous lim. | 77% | PHB | n.r. | [67] |
| Aulosira fertilissima | Maltose, balanced | 15.9% | PHB | 2.3 g/L | [67] |
Table 5.
Growing cyanobacteria with alternative CO2-sources.
| Type of Gas | Cyanobacterium | CO2 Source | Reference |
|---|---|---|---|
| Flue gases | Phormidium valderianum | Coal combustion flue gas | [71] |
| Atrhrospira platensis | Coal combustion flue gas | [72] | |
| Arthrospira sp. | Synthetic flue gas | [73] | |
| Synechocystis sp. | Flue gas from natural gas combustion | [74] | |
| CO2 rich fermentation gases | Arthrospira platensis | CO2-offgas from ethanol fermentation | [75] |
| Arthrospira platensis | Biogas | [76] |
Table 6.
Overview of agro-industrial effluents and wastewaters and anaerobic digestates used as nutrient sources for cultivating cyanobacteria.
Table 6.
Overview of agro-industrial effluents and wastewaters and anaerobic digestates used as nutrient sources for cultivating cyanobacteria.
| Nutrient Source | Cyanobacterium | Total cdw/Growth Rate | Product/Purpose | Reference | |
|---|---|---|---|---|---|
| Agro-industrial effluents and waste waters | Raw cow manure | Arthrospira maxima | 3.15 g/L | Biomass production | [80] |
| Molasses | Arthrospira platensis | 2.9 g/L | Biomass production | [95] | |
| Olive-oil mill wastewater | Arthrospira platensis | 1.69 g/L | Nutrient removal | [84] | |
| Poultry litter | Nostoc muscorum agardh | 0.62 g/L | PHA production | [66] | |
| Anaerobic digestate | Waste from pig farm | Arthrospira platensis | 20 g/m2/d | Nutrient removal | [81] |
| Digested sago effluent | Arthrospira platensis | 0.52–0.61 g/L | Nutrient removal | [96] | |
| Digestate from municipal solid waste | Arthrospira platensis | Growth rate 0.04 d−1 | Nutrient removal | [97] | |
| Digestate from vegetable waste | Arthrospira platensis | Growth rate 0.20 d−1 | Nutrient removal | [97] | |
| Waste from pig farm | Arthrospira sp. | 15 g/m2/d | Nutrient removal | [85] | |
| Algal digestate | Chroococcus sp. | 0.79 g/L | Nutrient removal | [86] | |
| Digestate sludges | Lyngbya aestuarii | 0.28 g/L | Biomass production | [83] | |
| Digestates of Scenedesmus spp. | Lyngbya aestuarii | 0.11 g/L | Biomass production | [83] | |
| Thin stillage digestate | Synechocystis cf. salina Wislouch | 1.6 g/L | PHB production | [92] | |
| Anaerobic digester effluent | Synechocystis sp. | 0.15 g/L | Lipid production | [98] | |
Table 7.
Overview of selected trials conducted in a tubular photobioreactor at pilot scale.
| Trial | Strain | Nutrient Solution | Cultivation Time | Final Biomass Concentration | Final PHB-Concentration of cdw |
|---|---|---|---|---|---|
| 1. Mineral medium | Synechocystis salina CCALA192 | Optimized BG11 | June 21 days | 2.0 ± 0.12 g/L | 6.6% ± 0.5% |
| 2. Acetate addition | Synechocystis salina CCALA192 | Optimized BG11, 20 mM acetate | July 26 days | 1.9 ± 0.02 g/L | 6.0% ± 0.1% |
| 3. Acetate addition | Synechocystis salina CCALA192 | Optimized BG11, 60 mM acetate | September 24 days | Trial cancelled, due to contaminations with fungi | |
| 4. 24 h illumination | Synechocystis salina CCALA192 | Optimized BG11 | October 27 days | 1.8 ± 0.02 g/L | 4.8% ± 0.0% |
| 5. Alternative nutrient source | Synechocystis salina CCALA192 | Digestate supernatant | November–December 40 days | 1.6 ± 0.02 g/L | 5.5% ± 0.3% |
| 6. Mineral medium | Synechocystis salina CCALA192 | Optimized BG11 | December–January 30 days | 2.1 ± 0.03 g/L | 6.0% ± 0.02% |
| 7. Optimal degassing | Synechocystis salina CCALA192 | Optimized BG11 | May 7 days | 0.9 ± 0.03 g/L (Trial prematurely cancelled due to ciliates) | 9% ± 0.1% (Trial prematurely cancelled due to ciliates) |
| 8. Chlorogloeopsis fritschii CCALA39 | Chlorogloeopsis fritschii CCALA39 | Optimized BG11 | February 11 days | Trial cancelled, due to lack of growth | |
| 9. Arthrospira | Arthrospira sp. | Spirulina Medium | October 7 days | Trial cancelled, due to lack of growth | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Troschl, C.; Meixner, K.; Drosg, B. Cyanobacterial PHA Production—Review of Recent Advances and a Summary of Three Years’ Working Experience Running a Pilot Plant. Bioengineering 2017, 4, 26. https://doi.org/10.3390/bioengineering4020026
AMA Style
Troschl C, Meixner K, Drosg B. Cyanobacterial PHA Production—Review of Recent Advances and a Summary of Three Years’ Working Experience Running a Pilot Plant. Bioengineering. 2017; 4(2):26. https://doi.org/10.3390/bioengineering4020026
Chicago/Turabian StyleTroschl, Clemens, Katharina Meixner, and Bernhard Drosg. 2017. "Cyanobacterial PHA Production—Review of Recent Advances and a Summary of Three Years’ Working Experience Running a Pilot Plant" Bioengineering 4, no. 2: 26. https://doi.org/10.3390/bioengineering4020026
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.