
The Physiological Functions of AbrB on Sporulation, Biofilm Formation and Carbon Source Utilization in Clostridium tyrobutyricum

Abstract
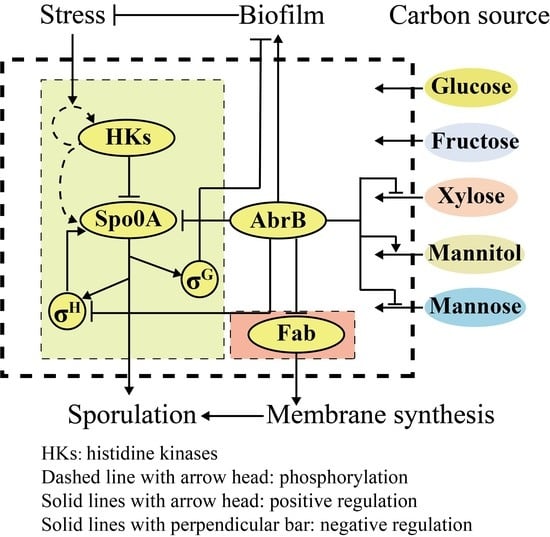
:
1. Introduction
2. Materials and Methods
2.1. AbrB Sequence Alignment and Functional Domain Prediction in C. tyrobutyricum ATCC 25755
2.2. Strains and Culture Conditions
2.3. Plasmids Construction and Transformation
2.4. Biofilm Formation Assay
2.5. Sporulation Assay
2.6. Stress Tolerance Assessment and Plate Survival Assay
2.7. Quantitative Real-Time PCR (qRT-PCR)
2.8. Fermentation Kinetics Studies
2.9. Analytical Methods
3. Results
3.1. Multiple Sequence Alignment and Functional Domain Prediction of AbrB
3.2. Phenotypic Analysis of the AbrB Mutants
3.3. Negative Regulation of AbrB on Sporulation
3.4. Positive Regulation of AbrB on Biofilm Formation
3.5. AbrB could Enhance the Stress Resistance of C. tyrobutyricum
3.6. Effect of AbrB on the Fermentation Performance of C. tyrobutyricum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Strauch, M.A.; Hoch, J.A. Transition-state regulators. sentinels of Bacillus subtilis post-exponential gene expression. Mol. Microbiol. 1993, 7, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Strauch, M.A. Regulation of Bacillus subtilis gene expression during the transition from exponential growth to stationary phase. Prog. Nucleic. Acid Res. Mol. Biol. 1993, 46, 121–153. [Google Scholar] [PubMed]
- Barbieri, G.; Albertini, A.M.; Ferrari, E.; Sonenshein, A.L.; Belitsky, B.R. Interplay of CodY and ScoC in the Regulation of Major Extracellular Protease Genes of Bacillus subtilis. J. Bacteriol. 2016, 198, 907–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, J.B.; Gocht, M.; Marahiel, M.A.; Zuber, P. AbrB, a regulator of gene expression in Bacillus, interacts with the transcription initiation regions of a sporulation gene and an antibiotic biosynthesis gene. Proc. Natl. Acad. Sci. USA 1989, 86, 8457–8461. [Google Scholar] [CrossRef] [Green Version]
- Hamon, M.A.; Stanley, N.R.; Britton, R.A.; Grossman, A.D.; Lazazzera, B.A. Identification of AbrB-regulated genes involved in biofilm formation by Bacillus subtilis. Mol. Microbiol. 2004, 52, 847–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, J.; Wang, Y.; Li, J.; Shen, Q.; Zhang, R. Enhanced root colonization and biocontrol activity of Bacillus amyloliquefaciens SQR9 by abrB gene disruption. Appl. Microbiol. Biotechnol. 2013, 97, 8823–8830. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Yu, Z.; Du, C.; Gong, Y.; Yin, W.; Li, X.; Li, Z.; Römling, U.; Chou, S.-H.; He, J. 2-Methylcitrate cycle. a well-regulated controller of Bacillus sporulation. Environ. Microbiol. 2020, 22, 1125–1140. [Google Scholar] [CrossRef] [PubMed]
- Strauch, M.A.; Perego, M.; Burbulys, D.; AHoch, J. The transition state transcription regulator AbrB of Bacillus subtilis is autoregulated during vegetative growth. Mol. Microbiol. 1989, 3, 1203–1209. [Google Scholar] [CrossRef]
- Strauch, M.A.; Spiegelman, G.B.; Perego, M.; Johnson, W.C.; Burbulys, D.; AHoch, J. The transition state transcription regulator abrB of Bacillus subtilis is a DNA binding protein. EMBO J. 1989, 8, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Strauch, M.A. Delineation of AbrB-binding sites on the Bacillus subtilis spo0H, kinB, ftsAZ, and pbpE promoters and use of a derived homology to identify a previously unsuspected binding site in the bsuB1 methylase promote. J. Bacteriol. 1995, 177, 6999–7002. [Google Scholar] [CrossRef]
- Xu, K.; Strauch, M.A. In vitro selection of optimal AbrB-binding sites comparison to known in vivo sites indicates flexibility in AbrB binding and recognition of three-dimensional DNA structures. Mol. Microbiol. 1996, 19, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Benson, L.M.; Vaughn, J.L.; Strauch, M.A.; Bobay, B.G.; Thompson, R.; Naylor, S.; Cavanagh, J. Macromolecular assembly of the transition state regulator AbrB in its unbound and complexed states probed by microelectrospray ionization mass spectrometry. Anal. Biochem. 2002, 306, 222–227. [Google Scholar] [CrossRef]
- Vaughn, J.L.; Feher, V.; Naylor, S.; Strauch, M.A.; Cavanagh, J. Novel DNA binding domain and genetic regulation model of Bacillus subtilis transition state regulator abrB. Nat. Struct. Biol. 2000, 7, 1139–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, Z.E.; Strauch, M.A. Role of Cys54 in AbrB multimerization and DNA-binding activity. FEMS Microbiol. Lett. 2001, 203, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Clark, D.; Strauch, M.A. Analysis of abrB mutations, mutant proteins, and why abrB does not utilize a perfect consensus in the -35 region of its sigma A promoter. J. Biol. Chem. 1996, 271, 2621–2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scotcher, M.C.; Rudolph, F.B.; Bennett, G.N. Expression of abrB310 and SinR, and effects of decreased abrB310 expression on the transition from acidogenesis to solventogenesis, in Clostridium acetobutylicum ATCC 824. Appl. Environ. Microbiol. 2005, 71, 1987–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafikhani, S.H.; Leighton, T. AbrB and Spo0E control the proper timing of sporulation in Bacillus subtilis. Curr. Microbiol. 2004, 48, 262–269. [Google Scholar] [CrossRef]
- Strauch, M.A.; Bobay, B.G.; Cavanagh, J.; Yao, F.; Wilson, A.; Breton, Y.L. Abh and AbrB control of Bacillus subtilis antimicrobial gene expression. J. Bacteriol. 2007, 189, 7720–7732. [Google Scholar] [CrossRef] [Green Version]
- Dubnau, D. The regulation of genetic competence in Bacillus subtilis. Mol. Microbiol. 1991, 5, 11–18. [Google Scholar] [CrossRef]
- Shu, C.-C.; Wang, D.; Guo, J.; Song, J.-M.; Chen, S.-W.; Chen, L.-L.; Gao, J.-X. Analyzing AbrB-Knockout Effects through Genome and Transcriptome Sequencing Bacillus licheniformis of DW2. Front Microbiol. 2018, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Klausmann, P.; Lilge, L.; Aschern, M.; Hennemann, K.; Henkel, M.; Hausmann, R.; Heravi, K.M. Influence of B. subtilis 3NA mutations in spo0A and abrB on surfactin production in B. subtilis 168. Microb. Cell Fact. 2021, 20, 188. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, H.; Fang, H.; Sun, Y.; Zhou, H.; Yang, G.; Lu, F. Transcriptome based functional identification and application of regulator AbrB on alkaline protease synthesis in Bacillus licheniformis 2709. Int. J. Biol. Macromol. 2021, 166, 1491–1498. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Yang, Y.; Chen, J.; Chen, L.; Yang, S.; Jiang, W.; Gu, Y. Roles of three AbrBs in regulating two-phase Clostridium acetobutylicum fermentation. Appl. Microbiol. Biotechnol. 2016, 100, 9081–9089. [Google Scholar] [CrossRef] [PubMed]
- Obana, N.; Nakamura, K.; Nomura, N. A Sporulation Factor Is Involved in the Morphological Change of Clostridium perfringens Biofilms in Response to Temperature. J. Bacteriol. 2014, 196, 1540–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagerlund, A.; Dubois, T.; Økstad, O.-A.; Verplaetse, E.; Gilois2, N.; Bennaceur, I.; Perchat, S.; Gominet, M.; Aymerich, S.; Kolstø, A.-B.; et al. SinR Controls Enterotoxin Expression in Bacillus thuringiensis Biofilms. PLoS ONE 2014, 9, e87532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lin, M.; Yu, L.; Xu, M.; Tang, I.-C.; Yang, S.-T.; Yan, Q. Metabolic engineering of Clostridium tyrobutyricum for n-butanol production from sugarcane juice. Appl. Microbiol. Biotechnol. 2017, 101, 4327–4337. [Google Scholar] [CrossRef]
- Fu, H.; Yu, L.; Lin, M.; Wang, J.; Xiu, Z.; Yang, S.-T. Metabolic engineering of Clostridium tyrobutyricum for enhanced butyric acid production from glucose and xylose. Metab. Eng. 2017, 40, 50–58. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Qin, S.; Yang, Z.; Bai, X.; Suo, Y.; Wang, J. Butyric acid production from spent coffee grounds by engineered Clostridium tyrobutyricum overexpressing galactose catabolism genes. Bioresour. Technol. 2020, 304, 122977. [Google Scholar] [CrossRef]
- Lee, J.; Jang, Y.-S.; Han, M.-J.; Kim, J.Y.; Lee, S.Y. Deciphering Clostridium tyrobutyricum Metabolism Based on the Whole-Genome Sequence and Proteome Analyses. MBio 2016, 7, e00743-16. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Cheng, C.; Cao, G.; Yang, S.-T.; Ren, N. Comparative transcriptome analysis of Clostridium tyrobutyricum expressing a heterologous uptake hydrogenase. Sci. Total Environ. 2020, 749, 142022. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain resource annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.R.; Young, D.I.; Young, M. Conjugative plasmid transfer from Escherichia coli to Clostridium acetobutylicum. J. Gen. Microbiol. 1990, 136, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Fu, H.; Feng, J.; Hu, J.; Wang, J. Direct conversion of untreated cane molasses into butyric acid by engineered Clostridium tyrobutyricum. Bioresour. Technol. 2020, 301, 122764. [Google Scholar] [CrossRef]
- Zhang, J.; Zong, W.; Hong, W.; Zhanga, Z.-T.; Wanga, Y. Exploiting endogenous CRISPR-Cas system for multiplex genome editing in Clostridium tyrobutyricum and engineer the strain for high-level butanol production. Metab. Eng. 2018, 47, 49–59. [Google Scholar] [CrossRef]
- Chen, K.-Y.; Rathod, J.; Chiu, Y.-C.; Chen, J.-W.; Tsai, P.-J.; Huang, I.-H. The Transcriptional Regulator Lrp Contributes to Toxin Expression, Sporulation, and Swimming Motility in Clostridium difficile. Front Cell Infect. Microbiol. 2019, 9, 356. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.; Dai, K.; Fu, H.; Wang, J. Enhanced ethanol production from lignocellulosic hydrolysates by Thermoanaerobacterium aotearoense SCUT27/Delta argR(1864) with improved lignocellulose-derived inhibitors tolerance. Renew. Energy 2021, 173, 652–661. [Google Scholar] [CrossRef]
- Perego, M.; Spiegelman, G.B.; Hoch, J.A. Structure of the gene for the transition state regulator, abrB. regulator synthesis is controlled by the spo0A sporulation gene in Bacillus subtilis. Mol. Microbiol. 1988, 2, 689–699. [Google Scholar] [CrossRef]
- Al-Hinai, M.A.; Jones, S.W.; Papoutsakis, E.T. The Clostridium sporulation programs-diversity and preservation of endospore differentiation. Mol. Biol. Rev. 2015, 79, 19–37. [Google Scholar] [CrossRef]
- Weir, J.; Predich, M.; Dubnau, E.; Nair, G.; Smith, I. Regulation of spo0H, a gene coding for the Bacillus subtilis sigma H factor. J. Bacteriol. 1991, 173, 521–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonenshein, A.L. Control of sporulation initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Al-Hinai, M.A.; Jones, S.W.; Papoutsakis, E.T. σK of Clostridium acetobutylicum is the first known sporulation-specific sigma factor with two developmentally separated roles, one early and one late in sporulation. J. Bacteriol. 2014, 196, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Kirk, D.G.; Dahlsten, E.; Zhang, Z.; Korkeala, H.; Lindström, M. Involvement of Clostridium botulinum ATCC 3502 sigma factor K in early-stage sporulation. Appl. Environ. Microbiol. 2012, 78, 4590–4596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore. assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef]
- Li, J.; Freedman, J.C.; Evans, D.R.; McClane, B.A. CodY Promotes Sporulation and Enterotoxin Production by Clostridium perfringens Type A Strain SM101. Infect. Immun. 2017, 85, e00855-16. [Google Scholar] [CrossRef] [Green Version]
- Pedrido, M.E.; Oña Pd Ramirez, W.; Leñini, C.; Goñi, A.; Grau, R. Spo0A links de novo fatty acid synthesis to sporulation and biofilm development in Bacillus subtilis. Mol. Microbiol. 2013, 87, 348–367. [Google Scholar] [CrossRef]
- Minoru, K.; Yoko, S.; Masayuki, K.; Miho, F.; Mao, T. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Lazazzera, B.A. The sporulation transcription factor Spo0A is required for biofilm development in Bacillus subtilis. Mol. Microbiol. 2001, 42, 1199–1209. [Google Scholar] [CrossRef]
- Kim, M.; Kim, K.-Y.; Lee, K.M.; Youn, S.H.; Lee, S.-M.; Woo, H.M.; Oh, M.-K.; Um, Y. Butyric acid production from softwood hydrolysate by acetate-consuming Clostridium sp S1 with high butyric acid yield and selectivity. Bioresour. Technol. 2016, 218, 1208–1214. [Google Scholar] [CrossRef]
- Fu, H.; Hu, J.; Guo, X.; Feng, J.; Yang, S.-T.; Wang, J. Butanol production from Saccharina japonica hydrolysate by engineered Clostridium tyrobutyricum- The effects of pretreatment method and heat shock protein overexpression. Bioresour. Technol. 2021, 335, 122977. [Google Scholar] [CrossRef]
- Bao, T.; Feng, J.; Jiang, W.; Fu, H.; Wang, J.; Yang, S.T. Recent advances in n-butanol and butyrate production using engineered Clostridium tyrobutyricum. World J. Microbiol. Biotechnol. 2020, 36, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hong, W.; Guo, L.; Wang, Y.; Wang, Y. Enhancing plasmid transformation efficiency and enabling CRISPR-Cas9Cpf1-based genome editing in Clostridium tyrobutyricum. Biotechnol. Bioengin. 2020, 117, 2911–2917. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Li, J.; Wang, X.; Zhan, Y.; Leu, S.-Y.; Wang, Y. Bioaugmentation with Clostridium tyrobutyricum to improve butyric acid production through direct rice straw bioconversion. Bioresour. Technol. 2018, 263, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Suo, Y.; Liao, Z.; Qu, C.; Fu, H.; Wang, J. Metabolic engineering of Clostridium tyrobutyricum for enhanced butyric acid production from undetoxified corncob acid hydrolysate. Bioresour. Technol. 2018, 271, 266–273. [Google Scholar] [CrossRef]
- Cao, X.; Chen, Z.; Liang, L.; Guo, L.; Jiang, Z.; Tang, F.; Yun, Y.; Wang, Y. Co-valorization of paper mill sludge and corn steep liquor for enhanced n-butanol production with Clostridium tyrobutyricum Δcat1, adhE2. Bioresour. Technol. 2019, 296, 122347. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Kim, K.-Y.; Lee, K.M.; Lee, S.-M.; Gong1, G.; Oh, M.K.; Um, Y. Enhanced butyric acid production using mixed biomass of brown algae and rice straw by Clostridium tyrobutyricum ATCC25755. Bioresour. Technol. 2019, 273, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Malkmes, M.J.; Zhu, L.; Huang, H.; Jiang, L. Metal-organic frameworks coupling simultaneous saccharication and fermentation for enhanced butyric acid production from rice straw under visible light by Clostridium tyrobutyricum CtΔack.cat1. Bioresour. Technol. 2021, 332, 125117. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Z.; Lei, M.; Zhu, J.; Yang, Y.; Wu, S.; Yu, B.; Niu, H.; Ying, H.; Liu, D.; et al. Effects of Spo0A on Clostridium acetobutylicum with an emphasis on biofilm formation. World J. Microbiol. Biotechnol. 2020, 36, 80. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Darfeuille-Michaud, A. The σE Pathway Is Involved in Biofilm Formation by Crohn’s Disease Associated Adherent-Invasive Escherichia coli. J. Bacteriol. 2013, 195, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Miazek, K.; Remacle, C.; Richel, A.; Goffin, D. Effect of Lignocellulose Related Compounds on Microalgae Growth and Product Biosynthesis. A Review. Energies 2014, 7, 4446–4481. [Google Scholar] [CrossRef] [Green Version]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Minton, N.P. A modular system for Clostridium shuttle plasmids. J. Microbiol. Methods 2009, 78, 79–85. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Strain | Sugar Consumption Rate (g/L/h) | Butyrate | Acetate | ||
|---|---|---|---|---|---|---|
| g/L | g/L/h | g/L | g/L/h | |||
| Glucose | Ct | 0.71 ± 0.05 | 18.78 ± 0.17 | 0.26 ± 0.00 | 2.49 ± 0.03 | 0.03 ± 0.00 |
| Ct/ΔabrB | 0.67 ± 0.04 | 17.42 ± 0.14 * | 0.24 ± 0.00 * | 1.94 ± 0.06 | 0.03 ± 0.00 | |
| Xylose | Ct | 0.58 ± 0.02 | 18.40 ± 0.28 | 0.26 ± 0.00 | 0.44 ± 0.00 | 0.00 ± 0.00 |
| Ct/ΔabrB | 0.45 ± 0.01 ** | 13.71 ± 0.57 ** | 0.19 ± 0.01 ** | - | - | |
| Glucose/Xylose | Ct | 0.97 ± 0.02(G)/ 0.02 ± 0.01(X) | 7.01 ± 0.07 | 0.10 ± 0.00 | 2.02 ± 0.04 | 0.17 ± 0.00 |
| Ct/ΔabrB | 0.65 ± 0.02(G)/0.19 ± 0.02(X) ** | 13.11 ± 0.92 * | 0.18 ± 0.01 * | 1.06 ± 0.28 * | 0.09 ± 0.02 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, K.; Guo, X.; Zhang, H.; Fu, H.; Wang, J. The Physiological Functions of AbrB on Sporulation, Biofilm Formation and Carbon Source Utilization in Clostridium tyrobutyricum. Bioengineering 2022, 9, 575. https://doi.org/10.3390/bioengineering9100575
Luo K, Guo X, Zhang H, Fu H, Wang J. The Physiological Functions of AbrB on Sporulation, Biofilm Formation and Carbon Source Utilization in Clostridium tyrobutyricum. Bioengineering. 2022; 9(10):575. https://doi.org/10.3390/bioengineering9100575
Chicago/Turabian StyleLuo, Kui, Xiaolong Guo, Huihui Zhang, Hongxin Fu, and Jufang Wang. 2022. "The Physiological Functions of AbrB on Sporulation, Biofilm Formation and Carbon Source Utilization in Clostridium tyrobutyricum" Bioengineering 9, no. 10: 575. https://doi.org/10.3390/bioengineering9100575