
Bioprocess Strategies for Vitamin B12 Production by Microbial Fermentation and Its Market Applications


Abstract
:1. Historical Overview
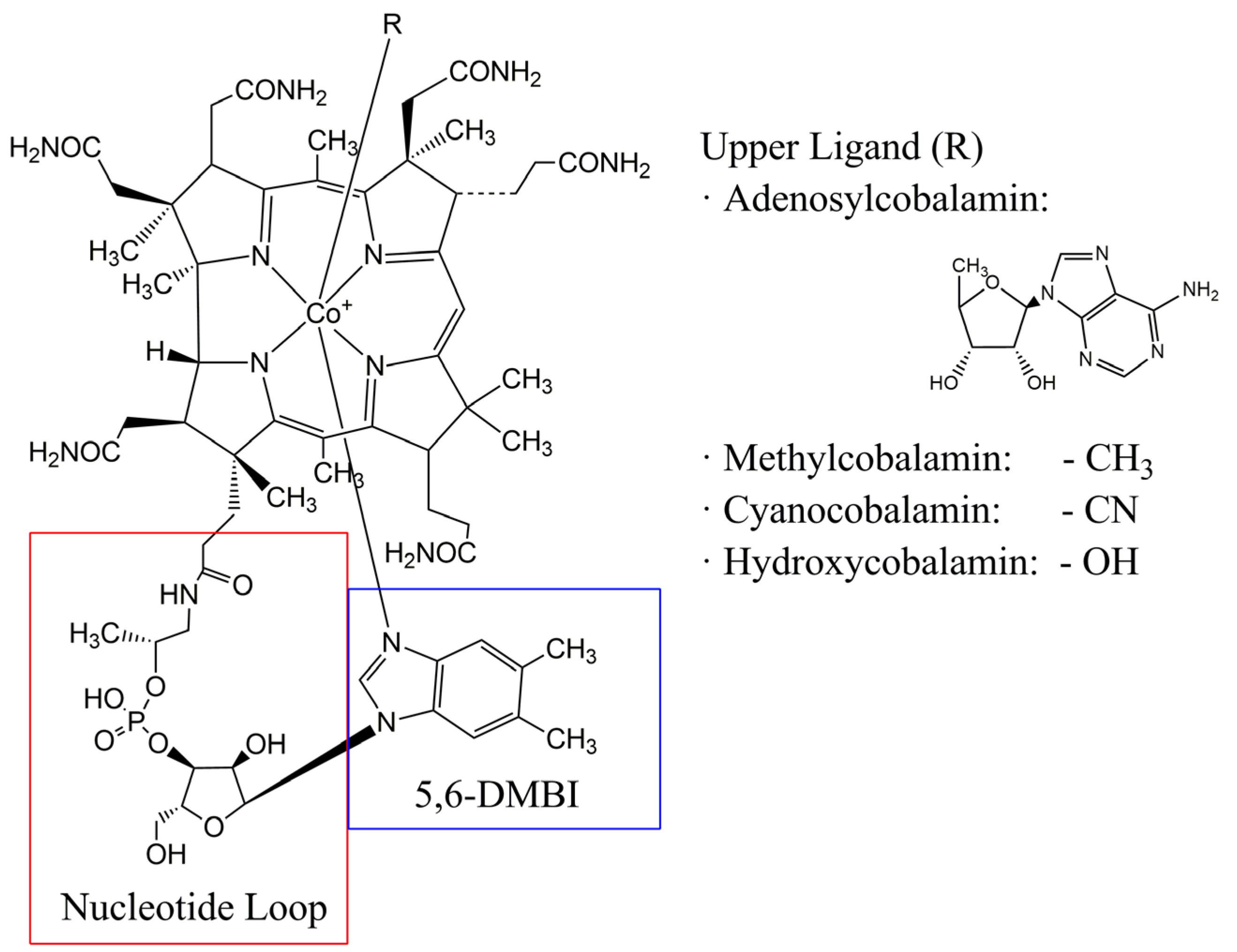
2. Structure of Cobalamin Derivatives and Functions as Enzyme Cofactors
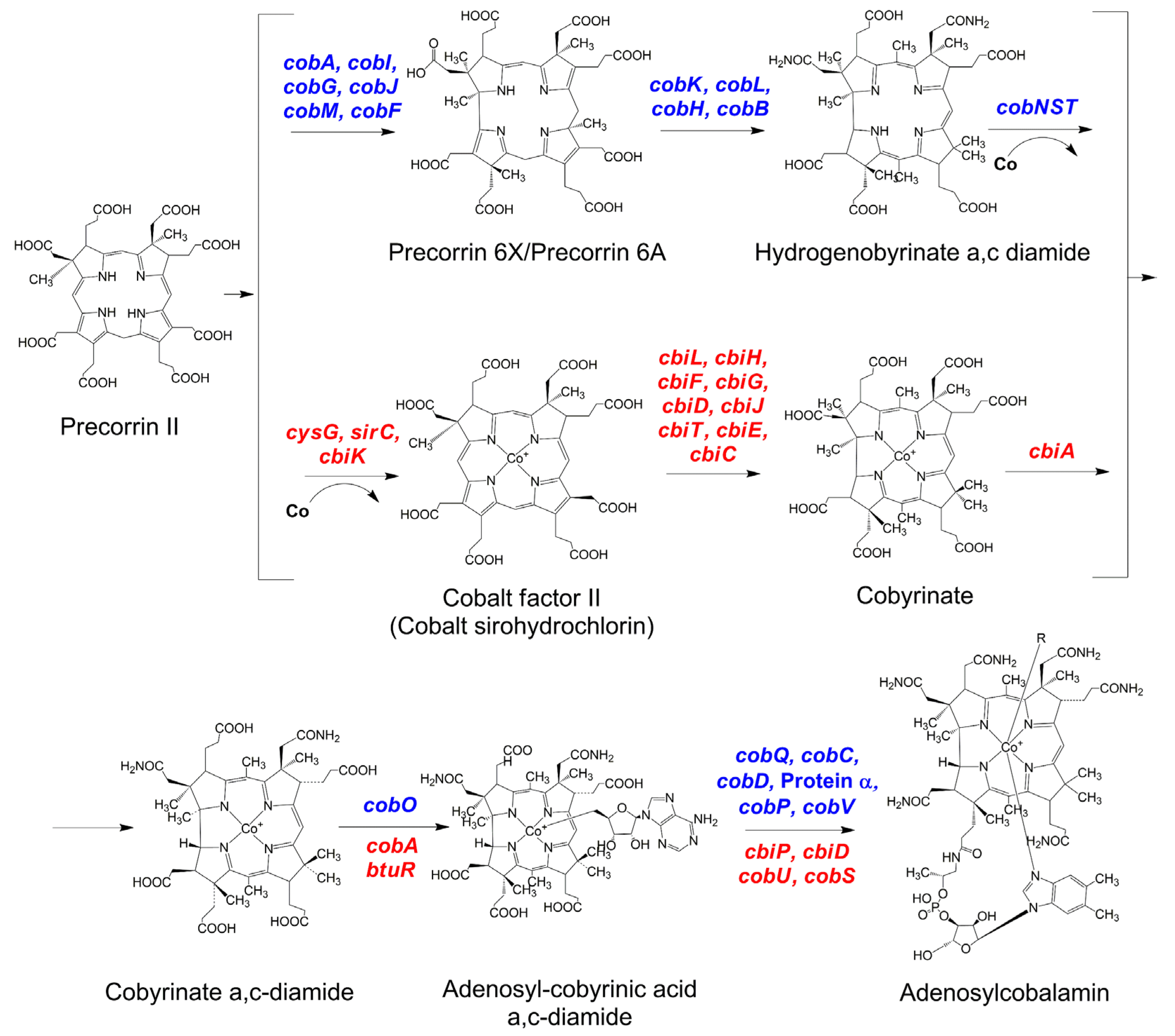
3. Biosynthesis of Vitamin B12: The Aerobic and Anaerobic Pathways
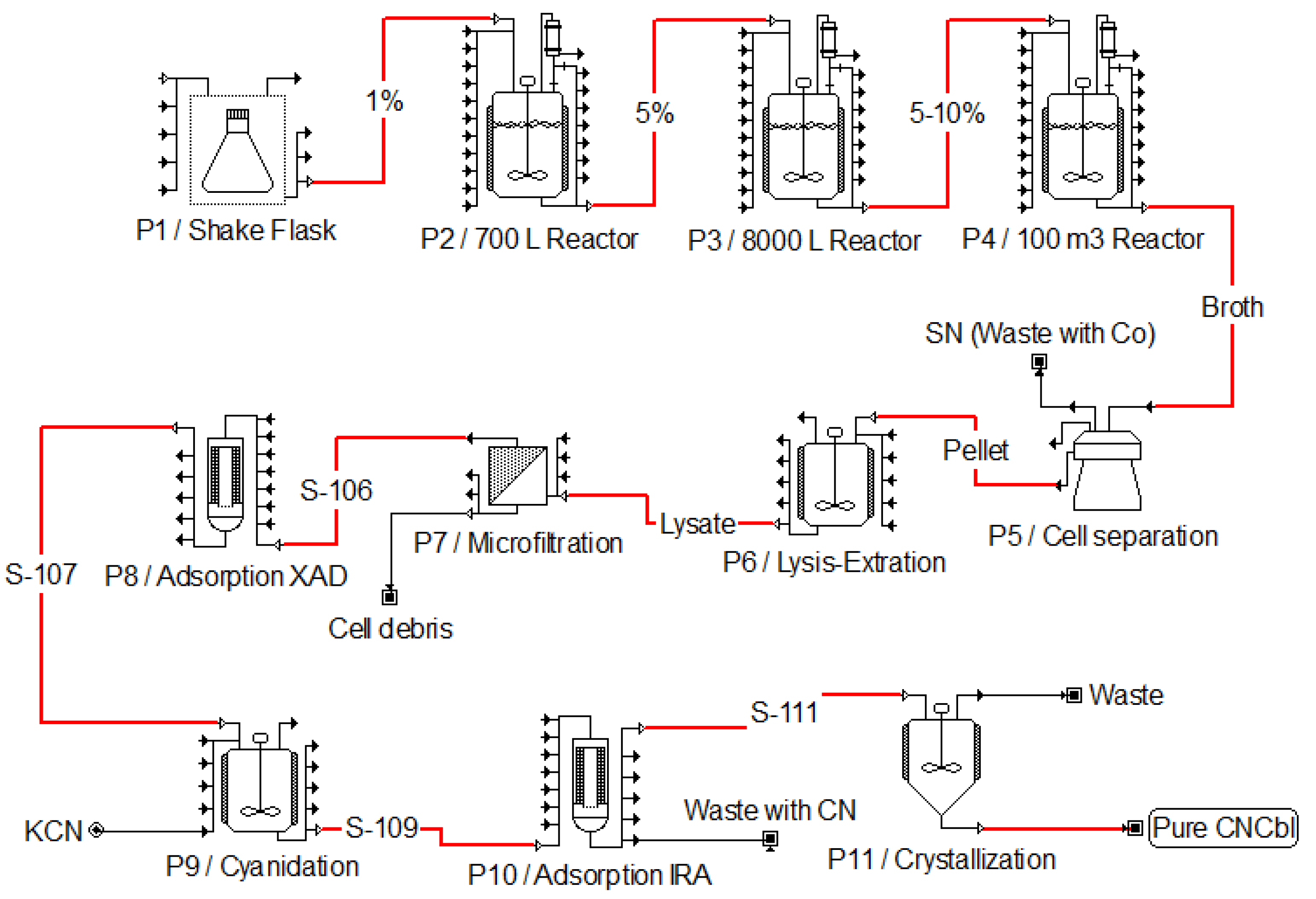
4. Microbial Production of Vitamin B12: Bioprocess Optimization for Cyanocobalamin Production
4.1. Microbial Production in Pseudomonas denitrificans
4.2. Microbial Production in Propionibacterium freudenreichii
5. Vitamin B12 Downstream Processing and Post-Modification Strategies
6. Patents—State of the Art

{kind=link}
{kind=link}
{kind=link}
| Patent Application Number (Reference) | Name | Microorganism/Strain | Innovation | Volumetric Production | Year | |
|---|---|---|---|---|---|---|
| Propionibacterium genus | US4544633A [43] (Expired) | Process for producing vitamin B12 by the fermentation technique, and vitamin B12-producing microorganism | P. freudenreichii (IFO 12424, IFO 12391, IFO 12426) | Creation of propionic-resistant strains (P. freudenreichii FERM-86 and FERM-87) for enhanced CNCbl production | 15 mg/L | 1983 |
| US6492141B1 [107] (Expired) | Process for the production of vitamin B12 | P. freudenreichii CBS 929.97 | O2 effect in production during the anaerobic phase and a "fill and draw" strategy for enhanced production | 19 mg/L | 1999 | |
| US6187761B1 [117] (Expired) | Production and use of compositions comprising high concentrations of vitamin B12 activity | P. freudenreichii subsp. shermanii and P. denitrificans | Method for producing vitamin B12 and making highly concentrated compositions | 10 mg/L | 1999 | |
| US7427397B2 [108] (Expired) | Probiotic Propionibacterium | Propionibacterium jensenii 702 | Propionibacterium jensenii as a probiotic | 0.0012 mg/L | 2004 | |
| EP2376644B1 [118] (Active) | Process for the preparation of a fermentation broth | Lactobacillus plantarum DSM 22,118 and P. freudenreichii DSM 22120 | Fermentation media optimization and co-culture for folate and vitamin B12 production | 1.07 mg/L | 2009 | |
| CN206828509U [110] (Active) | A device for producing propionic acid and co-producing vitamin B12 by semi-continuous fermentation | P. freudenreichii | Simultaneous production of propionic acid and vitamin B12 in a semicontinuous fermentation with propionic acid separation | 20.12 mg/L | 2017 | |
| US9938554 [109] (Active) | Co-cultivation of Propionibacterium and yeast. | P. freudenreichii (ATCC 6207) and yeast cells (DSM 28271) | Co-culture of Propionibacterium and propionic-resistant yeast to decrease the chemical oxygen load (COD) of spent media | 16 mg/L | 2018 | |
| US20200149084A1 [83] (Active) | Sequential co-culturing method for producing a vitamin- and protein-rich food product | Basidiomycota and P. freudenreichii | Co-culture of Basidiomycota genus strains and vitamin B12-producing strains for in situ food fortification | 0.0014 mg/L 1 | 2020 | |
| IN201827044769 A [111] (Active) | Continuous process for co-production of vitamin B12 and organic acids | P. freudenreichii (ATCC 13673) | Co-production of vitamin B12 and organic acids in a continuous fermentation with a single bioreactor | 76.13 mg/L | 2020 | |
| WO21041759 A1 [119] (Active) | Modified Propionibacterium and methods of use | P. freudenreichii (P. UF 1) | Generation of a vitamin B12-overproducing strain by introducing a mutation that decreases the activity of the cbiMcbl riboswitch | n.d. 2 | 2021 | |
| Pseudomonas denitrificans | US3018225A [100] (Expired) | Production of vitamin B12 | P. denitrificans MB-580 | A process for vitamin B12 production with a high-yield strain (P. denitrificans MB-580) | 2.4 mg/L 1 | 1962 |
| US20060019352A1 [101] (Abandoned) | Methods for increasing the production of cobalamins using cob gene expression | P. denitrificans | Overexpression of several genes involved in Cob biosynthesis; generation of several overproducing strains, such as SC-510 | 65 mg/L | 1990 | |
| US6156545A [120] (Expired) | Biosynthesis method enabling the preparation of cobalamins | P. denitrificans G2650 | Enhanced Cob production by the heterologous overexpression of precursors, such as DMBI and O-phospo-L-threonine | 7.9 mg/L | 1996 | |
| CN101538599A [121] (Active) | Method for improving the yield of denitrified pseudomonas vitamin B12 | P. denitrificans J741 | Enhance cob production by betaine addition optimization | 177.49 mg/L | 2008 | |
| CN102399845A [122] (Active) | Vitamin B12 fermentation production control process based on CO2 concentration in tail gas | P. denitrificans MB-580 | Vitamin B12 enhanced production through a carbon dioxide control strategy during fermentation | 164.6 mg/L | 2010 | |
| CN101748177 A [123] (Active) | Optimized method for producing vitamin B12 through P. denitrificans fermentation and synthetic medium | P. denitrificans | Development and optimization of media and bioprocess conditions for improved vitamin B12 production | 77 mg/L | 2010 | |
| CN102021214 A [124] (Active) | Oxygen consumption rate-based vitamin B12 fermentation production control process | P. denitrificans | Vitamin B12 production optimization through an oxygen control strategy | 171,4 mg/L | 2011 | |
| CN102453740 A [125] (Active) | Culture medium for producing vitamin B12 by fermenting P. denitrificans and fermentation method thereof | P. denitrificans | Use of artificial molasses and bioprocess optimization for a more stable fermentation yield | 198 mg/L | 2012 | |
| CN108949866 A [103] (Active) | Multi-stage rotating speed regulating policy for improving P. denitrificans fermentation for production of vitamin B12 | P. denitrificans | Vitamin B12 production improved by optimization of the culture media and the stirring speed of the bioprocess | 246 mg/L 1 | 2018 | |
| CN108913739 A [126] (Active) | Method for producing vitamin B12 by using P. denitrificans based on pH value control | P. denitrificans | Improved vitamin B12 production by optimization of the bioprocess through pH value control | 248 mg/L | 2018 | |
| CN110205350 A [104] (Active) | Method for improving the yield of vitamin B12 based on the regulation of ammonia nitrogen index | P. denitrificans | A method for improved Cbl production by supplementation with yeast extract controlled by the ammonia nitrogen index | 167 mg/L 1 | 2019 | |
| CN109837320 A [105] (Active) | Method for promoting P. denitrificans to generate vitamin B12 | P. denitrificans | Optimization of media and culture conditions for improved vitamin B12 production | 198 mg/L | 2019 | |
| CN111808158 A [106] (Active) | Preparation method of vitamin B12 crude product | P. denitrificans | Downstream process improvement for AdoCbl extraction | n.d. 2 | 2020 | |
| CN111254173 A [102] (Active) | Screening method and screening culture medium for bacterial strains for high yield of vitamin B12 produced through fermentation production with P. denitrificans | Several high-yield strains of P. denitrificans | Screening for high-vitamin B12 producing P. denitrificans strains and culture medium screening for high vitamin B12 production | 281 mg/L 1 | 2020 | |
| Other producers | US2650896A [127] (Expired) | Cyanide ions in production of vitamin B12 | Streptomyces griseus | Effects of cyanide ions in B12 production | Biological assay | 1953 |
| US2576932A [112] (expired) | Fermentation process to produce vitamin B12 | B. megaterium B-938 | Vitamin B12 production with B. megaterium in a nutrient media with sucrose | 0.45 mg/L | 1983 | |
| US20050227332A1 [128] (Expired) | Method for producing vitamin B12 from hydrogen-metabolizing methane bacterium | A mesophilic methane bacterium obtained from digested sludge | The culture is acclimatized in a H2–CO media and grown in an immobilized bed bioreactor | 25.2 mg/L | 2005 | |
| US20060105432A1 [129] (Abandoned) | Method for the production of vitamin B12 | B. megaterium DSMZ509 | Genetically modified B. megaterium strain | 0.008 mg/L 1 | 2006 | |
| WO2011154820A2 [113] (Application granted) | Vitamin B12-producing probiotic bacterial strains | Lactobacillus reuteri (DSM 17938, DSM 16143, ATCC 55730) | In situ food fortification for increased vitamin B12 production with Lactobacillus reuteri strains | 0.018 mg/L 1 | 2011 | |
| CN104342390 A [114] (Active) | Sinorhizobium meliloti strain and composition and application of Sinorhizobium meliloti strain | S. meliloti (CGMCC 9638) | A S. melitolli strain capable of producing vitamin B12 and optimization of the bioprocess for vitamin B12 production | At least 50 mg/L | 2015 | |
| WO2019109975A1 [116] (Active) | Recombinant strain of Escherichia coli for de novo synthesis of vitamin B12, construction method therefor and application thereof | E. coli | Recombinant E. coli for the de novo synthesis of vitamin B12 | 89 µg/g DCW | 2019 | |
| CN110804598 A [115] (Active) | Procorrin-2C(20)-methyltransferase mutant and mutant gene and application thereof in preparing vitamin B12 | Sinorhizobium (CGMCC 9638) | Generation of a vitamin B12 overproducer strain by overexpressing the precorrin-2C(20)-methyltransferase gene | 115 mg/L | 2020 |
7. Vitamin B12 Market Applications and the State of the Market
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martens, H.; Barg, M.; Warren, D.; Jah, J.-H. Microbial Production of Vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Minot, G.R.; Murphy, W.P. Treatment of Pernicious Anemia by a Special Diet. J. Am. Med. Assoc. 1926, 87, 1666. [Google Scholar] [CrossRef]
- Rickes, E.L.; Brink, N.G.; Koniuszy, F.R.; Wood, T.R.; Folkers, K. Crystalline Vitamin B12. Science 1948, 107, 396–397. [Google Scholar] [CrossRef] [PubMed]
- Smith, L. Purification of Anti-Pernicious Anemia Factors from Liver. Nature 1948, 161, 638–639. [Google Scholar] [CrossRef]
- Hodgkin, D.C.; Kamper, J.; Lindsey, J.; Mackay, M.; Pickworth, J.; Robertson, J.H.; Shoemaker, C.B.; White, J.G. The Structure of Vitamin B12. I. An Outline of the Crystallographic Investigation of Vitamin B12. Proc. R. Soc. London Ser. A. Math. Phys. Sci. 1957, 242, 228–263. [Google Scholar] [CrossRef]
- Dixon, M.M.; Huang, S.; Matthews, R.G.; Ludwig, M. The Structure of the C-Terminal Domain of Methionine Synthase: Presenting S-Adenosylmethionine for Reductive Methylation of B12. Structure 1996, 4, 1263–1275. [Google Scholar] [CrossRef] [Green Version]
- Mancia, F.; Keep, N.H.; Nakagawa, A.; Leadlay, P.F.; McSweeney, S.; Rasmussen, B.; Bösecke, P.; Diat, O.; Evans, P.R. How Coenzyme B12 Radicals Are Generated: The Crystal Structure of Methylmalonyl-Coenzyme A Mutase at 2 Å Resolution. Structure 1996, 4, 339–350. [Google Scholar] [CrossRef] [Green Version]
- Wuerges, J.; Garau, G.; Geremia, S.; Fedosov, S.N.; Petersen, T.E.; Randaccio, L. Structural Basis for Mammalian Vitamin B12 Transport by Transcobalmin. Proc. Natl. Acad. Sci. USA 2006, 103, 4386–4391. [Google Scholar] [CrossRef] [Green Version]
- Mathews, F.S.; Gordon, M.M.; Chen, Z.; Rajashankar, K.R.; Ealick, S.E.; Alpers, D.H.; Sukumar, N. Crystal Structure of Human Intrinsic Factor: Cobalamin Complex at 2.6-Å Resolution. Proc. Natl. Acad. Sci. USA 2007, 104, 17311–17316. [Google Scholar] [CrossRef] [Green Version]
- Alam, A.; Woo, J.S.; Schmitz, J.; Prinz, B.; Root, K.; Chen, F.; Bloch, J.S.; Zenobi, R.; Locher, K.P. Structural Basis of Transcobalamin Recognition by Human CD320 Receptor. Nat. Commun. 2016, 7, 12100. [Google Scholar] [CrossRef] [Green Version]
- Furger, E.; Frei, D.C.; Schibli, R.; Fischer, E.; Prota, A.E. Structural Basis for Universal Corrinoid Recognition by the Cobalamin Transport Protein Haptocorrin. J. Biol. Chem. 2013, 288, 25466–25476. [Google Scholar] [CrossRef] [Green Version]
- Koutmos, M.; Gherasim, C.; Smith, J.L.; Banerjee, R. Structural Basis of Multifunctionality in a Vitamin B12- Processing Enzyme. J. Biol. Chem. 2011, 286, 29780–29787. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Gherasim, C.; Banerjee, R.; Koutmos, M. Structure of Human B12 Trafficking Protein CblD Reveals Molecular Mimicry and Identifies a New Subfamily of Nitro-FMN Reductases. J. Biol. Chem. 2015, 290, 29155–29166. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Feng, Z.; Hou, W.T.; Jiang, Y.L.; Wang, L.; Sun, L.; Zhou, C.Z.; Chen, Y. Cryo-EM Structure of Human Lysosomal Cobalamin Exporter ABCD4. Cell Res. 2019, 29, 1039–1041. [Google Scholar] [CrossRef]
- Schubert, H.L.; Hill, C.P. Structure of ATP-Bound Human ATP:Cobalamin Adenosyltransferase. Biochemistry 2006, 45, 15188–15196. [Google Scholar] [CrossRef] [Green Version]
- Woodward, R. The Total Synthesis of Vitamin B12. Pure Appl. Chem. 1973, 33, 145–177. [Google Scholar] [CrossRef]
- Eschenmoser, A.; Wintner, C.E. Natural Product Synthesis and Vitamin B12. Science 1977, 196, 1410–1420. [Google Scholar] [CrossRef]
- Fink, R.G. Coenzyme B12-Based Chemical Precedent for Co-C Bond Homolysis and Other Key Elementary Steps; Krautler, B., Arigoni, D., Golding, B.T., Eds.; Wiley-VCH: Weinheim, Germany, 1998. [Google Scholar]
- Hannibal, L.; Axhemi, A.; Glushchenko, A.V.; Moreira, E.S.; Brasch, N.E.; Jacobsen, D.W. Accurate Assessment and Identification of Naturally Occurring Cellular Cobalamins Luciana. Clin. Chem. Lab. Med. 2008, 46, 1739–1746. [Google Scholar] [CrossRef] [Green Version]
- Juzeniene, A.; Nizauskaite, Z. Photodegradation of Cobalamins in Aqueous Solutions and in Human Blood. J. Photochem. Photobiol. B Biol. 2013, 122, 7–14. [Google Scholar] [CrossRef]
- Hogenkamp, H.P.C.; Vergamini, P.J.; Matwiyoff, N.A. The Effect of Temperature and Light on the Carbon43 Nuclear Magnetic Resonance Spectra of Alkylcorrinoids, Selectively Enriched with Carbon-13. J. Chem. SOC. (A) 1975, 23, 2628–2633. [Google Scholar] [CrossRef]
- Obeid, R.; Fedosov, S.N.; Nexo, E. Cobalamin Coenzyme Forms Are Not Likely to Be Superior to Cyano- and Hydroxyl-Cobalamin in Prevention or Treatment of Cobalamin Deficiency. Mol. Nutr. Food Res. 2015, 59, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.R.; Lawrence, J.G.; Bobik, T.A. Cobalamin (Coenzyme B12): Synthesis and Biological Significance. Annu. Rev. Microbiol. 1996, 50, 137–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeper, F.J. The Biosynthesis of Porphyrins, Chlorophylls, and Vitamin B12. Nat. Prod. Rep. 1989, 6, 171–203. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, D.; Blanche, F.; Cameron, B.; Crouzet, J.; Debussche, L.; Remy, E.; Vuilhorgne, M. Vitamin B12 Biosynthesis in Pseudomonas denitrificans. In Vitamin B12, and B12-Proteins; Kräutler, B., Arigoni, D., Golding, B.T., Eds.; Wiley-VCH: Weinheim, Germany, 1998; pp. 63–79. [Google Scholar]
- Crouzet, J.; Cauchois, L.; Blanche, F.; Debussche, L.; Thibaut, D.; Rouyez, M.C.; Rigault, S.; Mayaux, J.F.; Cameron, B. Nucleotide Sequence of a Pseudomonas Dentrificans 5.4-Kilobase DNA Fragment Containing Five Cob Genes and Identification of Structural Genes Encoding S-Adenosyl-L-Methionine: Uroporphyrinogen III Methyltransferase and Cobyrinic Acid a,c-Diamide Synthase. J. Bacteriol. 1990, 172, 5968–5979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamford, N.P.J.; Duggan, S.; Li, Y.; Alanine, A.I.D.; Crouzet, J.; Battersby, A.R. Biosynthesis of Vitamin B12: The Multi-Enzyme Synthesis of Precorrin-4 and Factor IV. Chem. Biol. 1997, 4, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.J.; Warren, M.J. The Anaerobic Biosynthesis of Vitamin B12. Biochem. Soc. Trans. 2012, 40, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.K. The Porphobilinogen Synthase Family of Metalloenzymes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2000, 56, 115–128. [Google Scholar] [CrossRef]
- Jordan, P.M. The Biosynthesis of 5-Aminolaevulinic Acid and Its Transformation into Uroporphyrinogen III. In New Comprehensive Biochemistry; Krebs, J., Michalak, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1991; Volume 19, pp. 1–66. [Google Scholar]
- Raux, E.; Schubert, H.L.; Warren, M.J. Biosynthesis of Cobalamin (Vitamin B12): A Bacterial Conundrum. Cell. Mol. Life Sci. 2000, 57, 1880–1893. [Google Scholar] [CrossRef]
- Warren, M.J.; Raux, E.; Schubert, H.L.; Escalante-Semerena, J.C. The Biosynthesis of Adenosylcobalamin (Vitamin B12). Nat. Prod. Rep. 2002, 19, 390–412. [Google Scholar] [CrossRef]
- Debussche, L.; Couder, M.; Thibaut, D.; Cameron, B.; Crouzet, J.; Blanche, F. Assay, Purification, and Characterization of Cobaltochelatase, a Unique Complex Enzyme Catalyzing Cobalt Insertion in Hydrogenobyrinic Acid a,c- Diamide during Coenzyme B12 Biosynthesis in Pseudomonas denitrificans. J. Bacteriol. 1992, 174, 7445–7451. [Google Scholar] [CrossRef] [Green Version]
- Mathur, Y.; Sreyas, S.; Datar, P.M.; Sathian, M.B.; Hazra, A.B. CobT and BzaC Catalyze the Regiospecific Activation and Methylation of the 5-Hydroxybenzimidazole Lower Ligand in Anaerobic Cobamide Biosynthesis. J. Biol. Chem. 2020, 295, 10522–10534. [Google Scholar] [CrossRef]
- Campbell, G.R.O.; Taga, M.E.; Mistry, K.; Lloret, J.; Anderson, P.J.; Roth, J.R.; Walker, G.C. Sinorhizobium Meliloti BluB Is Necessary for Production of 5,6-Dimethylbenzimidazole, the Lower Ligand of B12. Proc. Natl. Acad. Sci. USA 2006, 103, 4634–4639. [Google Scholar] [CrossRef] [Green Version]
- Balabanova, L.; Averianova, L.; Marchenok, M.; Son, O.; Tekutyeva, L. Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology. Int. J. Mol. Sci. 2021, 22, 4522. [Google Scholar] [CrossRef]
- Deptula, P.; Kylli, P.; Chamlagain, B.; Holm, L.; Kostiainen, R.; Piironen, V.; Savijoki, K.; Varmanen, P. BluB/CobT2 Fusion Enzyme Activity Reveals Mechanisms Responsible for Production of Active Form of Vitamin B12 by Propionibacterium freudenreichii. Microb. Cell Factories 2015, 14, 186. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Vo, T.P.; Ainala, S.K.; Kim, J.R.; Park, S. Analysis, Characterization of Coenzyme B12 Biosynthetic Gene Clusters and Improvement of B12 Biosynthesis in Pseudomonas denitrificans ATCC 13867. FEMS Microbiol. Lett. 2018, 365, fny211. [Google Scholar] [CrossRef]
- Hazra, A.B.; Han, A.W.; Mehta, A.P.; Mok, K.C.; Osadchiy, V.; Begley, T.P.; Taga, M.E. Anaerobic Biosynthesis of the Lower Ligand of Vitamin B12. Proc. Natl. Acad. Sci. USA 2015, 112, 10792–10797. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Kang, J.; Zhang, D. Microbial Production of Vitamin B12: A Review and Future Perspectives. Microb. Cell Fact. 2017, 16, 15. [Google Scholar] [CrossRef] [Green Version]
- Sobczyńska-Malefora, A.; Delvin, E.; McCaddon, A.; Ahmadi, K.R.; Harrington, D.J. Vitamin B12 Status in Health and Disease: A Critical Review. Diagnosis of Deficiency and Insufficiency–Clinical and Laboratory Pitfalls. Crit. Rev. Clin. Lab. Sci. 2021, 58, 399–429. [Google Scholar] [CrossRef]
- Hardlei, T.F.; Obeid, R.; Herrmann, W.; Nexo, E. Cobalamin Analogues in Humans: A Study on Maternal and Cord Blood. PLoS ONE 2013, 8, e61194. [Google Scholar] [CrossRef] [Green Version]
- Kojima, I. Process for Producing Vitamin B12 by the Fermentation Technique, and Vitamin B12-Producing Microorganism. U.S. Patent 4,544,633, 1 October 1985. [Google Scholar]
- Piao, Y.; Yamashita, M.; Kawaraichi, N.; Asegawa, R.; Ono, H.; Murooka, Y. Production of Vitamin B12 in Genetically Engineered Propionibacterium freudenreichii. J. Biosci. Bioeng. 2004, 98, 167–173. [Google Scholar] [CrossRef]
- Piao, Y.; Kiatpapan, P.; Yamashita, M.; Murooka, Y. Effects of Expression of HemA and HemB Genes on Production of Porphyrin in Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2004, 70, 7561–7566. [Google Scholar] [CrossRef] [Green Version]
- Biedendieck, R.; Malten, M.; Barg, H.; Bunk, B.; Martens, J.H.; Deery, E.; Leech, H.; Warren, M.J.; Jahn, D. Metabolic Engineering of Cobalamin (Vitamin B12) Production in Bacillus Megaterium. Microb. Biotechnol. 2010, 3, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, Y.; Lee, B.; Kang, Z.; Du, G. Development of a Two-Step Cultivation Strategy for the Production of Vitamin B12 by Bacillus Megaterium. Microb. Cell Factories 2014, 13, 102. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, C.; Zhu, X.; Schütz, D.; Fischer, M.; Bisping, B. Cobalamin Is Produced by Acetobacter Pasteurianus DSM 3509. Appl. Microbiol. Biotechnol. 2019, 103, 3875–3885. [Google Scholar] [CrossRef]
- Gu, Q.; Zhang, C.; Song, D.; Li, P.; Zhu, X. Enhancing Vitamin B12 Content in Soy-Yogurt by Lactobacillus Reuteri. Int. J. Food Microbiol. 2015, 206, 56–59. [Google Scholar] [CrossRef]
- Thirupathaiah, Y.; Rani, C.S.; Reddy, M.S.; Venkateswar Rao, L. Effect of Chemical and Microbial Vitamin B12 Analogues on Production of Vitamin B12. World J. Microbiol. Biotechnol. 2012, 28, 2267–2271. [Google Scholar] [CrossRef]
- Wang, P.; Shen, C.; Li, L.; Guo, J.; Cong, Q.; Lu, J. Simultaneous Production of Propionic Acid and Vitamin B12 from Corn Stalk Hydrolysates by Propionibacterium freudenreichii in an Expanded Bed Adsorption Bioreactor. Prep. Biochem. Biotechnol. 2020, 50, 763–767. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Y.; Liu, Y.; Shi, H.; Su, Z. Novel in Situ Product Removal Technique for Simultaneous Production of Propionic Acid and Vitamin B12 by Expanded Bed Adsorption Bioreactor. Bioresour. Technol. 2012, 104, 652–659. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Z.; Jiao, Y.; Liu, S.; Wang, Y. Improved Propionic Acid and 5,6-Dimethylbenzimidazole Control Strategy for Vitamin B12 Fermentation by Propionibacterium freudenreichii. J. Biotechnol. 2015, 193, 123–129. [Google Scholar] [CrossRef]
- Wang, P.; Jiao, Y.; Liu, S. Novel Fermentation Process Strengthening Strategy for Production of Propionic Acid and Vitamin B12 by Propionibacterium freudenreichii. J. Ind. Microbiol. Biotechnol. 2014, 41, 1811–1815. [Google Scholar] [CrossRef]
- Hugenschmidt, S.; Schwenninger, S.M.; Lacroix, C. Concurrent High Production of Natural Folate and Vitamin B12 Using a Co-Culture Process with Lactobacillus Plantarum SM39 and Propionibacterium freudenreichii DF13. Process Biochem. 2011, 46, 1063–1070. [Google Scholar] [CrossRef]
- Xie, C.; Coda, R.; Chamlagain, B.; Varmanen, P.; Piironen, V.; Katina, K. Co-Fermentation of Propionibacterium freudenreichiiand Lactobacillus Brevisin Wheat Bran for in Situproduction of Vitamin B12. Front. Microbiol. 2019, 10, 1541. [Google Scholar] [CrossRef] [PubMed]
- Miyano, K.I.; Ye, K.; Shimizu, K. Improvement of Vitamin B12 Fermentation by Reducing the Inhibitory Metabolites by Cell Recycle System and a Mixed Culture. Biochem. Eng. J. 2000, 6, 207–214. [Google Scholar] [CrossRef]
- Hajfarajollah, H.; Mokhtarani, B.; Mortaheb, H.; Afaghi, A. Vitamin B12 Biosynthesis over Waste Frying Sunflower Oil as a Cost Effective and Renewable Substrate. J. Food Sci. Technol. 2015, 52, 3273–3282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, V.V.; Prakash, G.; Lali, A.M. Growth Engineering of Propionibacterium freudenreichii Shermanii for Organic Acids and Other Value-Added Products Formation. Prep. Biochem. Biotechnol. 2018, 48, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, Y.; Su, Z. Improvement of Adenosylcobalamin Production by Metabolic Control Strategy in Propionibacterium freudenreichii. Appl. Biochem. Biotechnol. 2012, 167, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Kośmider, A.; Białas, W.; Kubiak, P.; Drozdzyńska, A.; Czaczyk, K. Vitamin B12 Production from Crude Glycerol by Propionibacterium freudenreichii ssp. Shermanii: Optimization of Medium Composition through Statistical Experimental Designs. Bioresour. Technol. 2012, 105, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Chamlagain, B.; Deptula, P.; Edelmann, M.; Kariluoto, S.; Grattepanche, F.; Lacroix, C.; Varmanen, P.; Piironen, V. Effect of the Lower Ligand Precursors on Vitamin B12 Production by Food-Grade Propionibacteria. LWT-Food Sci. Technol. 2016, 72, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Chamlagain, B.; Sugito, T.A.; Deptula, P.; Edelmann, M.; Kariluoto, S.; Varmanen, P.; Piironen, V. In Situ Production of Active Vitamin B12 in Cereal Matrices Using Propionibacterium freudenreichii. Food Sci. Nutr. 2018, 6, 67–76. [Google Scholar] [CrossRef]
- Deptula, P.; Chamlagain, B.; Edelmann, M.; Sangsuwan, P.; Nyman, T.A.; Savijoki, K.; Piironen, V.; Varmanen, P. Food-like Growth Conditions Support Production of Active Vitamin B12 by Propionibacterium freudenreichii 2067 without DMBI, the Lower Ligand Base, or Cobalt Supplementation. Front. Microbiol. 2017, 8, 368. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, Y.; Wu, J.; Fang, H.; Jin, Z.; Zhang, D. Metabolic Profiling Analysis of the Vitamin B12 Producer Propionibacterium freudenreichii. Microbiologyopen 2021, 10, e1199. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Wang, Z.; Wang, Y.; Ma, Y.; Su, Z. Metabolic Flux Analysis of Simultaneous Production of Vitamin B12 and Propionic Acid in a Coupled Fermentation Process by Propionibacterium freudenreichii. Appl. Biochem. Biotechnol. 2021, 193, 3045–3061. [Google Scholar] [CrossRef]
- Cheng, X.; Chen, W.; Peng, W.; Li, K. Improved Vitamin B12 Fermentation Process by Adding Rotenone to Regulate the Metabolism of Pseudomonas denitrificans. Appl. Biochem. Biotechnol. 2014, 173, 673–681. [Google Scholar] [CrossRef]
- Li, K.T.; Liu, D.H.; Li, Y.L.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. An Effective and Simplified PH-Stat Control Strategy for the Industrial Fermentation of Vitamin B12 by Pseudomonas denitrificans. Bioprocess Biosyst. Eng. 2008, 31, 605–610. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, H.; Li, Y.; Chu, J.; Huang, M.; Zhuang, Y.; Zhang, S. Improved Vitamin B12 Production by Step-Wise Reduction of Oxygen Uptake Rate under Dissolved Oxygen Limiting Level during Fermentation Process. Bioresour. Technol. 2010, 101, 2845–2852. [Google Scholar] [CrossRef]
- Wang, Z.; Shi, H.; Wang, P. The Online Morphology Control and Dynamic Studies on Improving Vitamin B12 Production by Pseudomonas denitrificans with Online Capacitance and Specific Oxygen Consumption Rate. Appl. Biochem. Biotechnol. 2016, 179, 1115–1127. [Google Scholar] [CrossRef]
- Xia, W.; Wei, P.; Kun, C. Interactive Performances of Betaine on the Metabolic Processes of Pseudomonas denitrificans. J. Ind. Microbiol. Biotechnol. 2014, 42, 273–278. [Google Scholar] [CrossRef]
- Xia, W.; Chen, W. Industrial Vitamin B12 Production by Pseudomonas denitrificans Using Maltose Syrup and Corn Steep Liquor as the Cost-Effective Fermentation Substrates. Bioprocess Biosyst. Eng. 2015, 38, 1065–1073. [Google Scholar] [CrossRef]
- Li, K.T.; Zhou, J.; Cheng, X.; Wei, S.J. Study on the Dissolved Oxygen Control Strategy in Large-Scale Vitamin B12 Fermentation by Pseudomonas denitrificans. J. Chem. Technol. Biotechnol. 2012, 87, 1648–1653. [Google Scholar] [CrossRef]
- Vandamme, E.J.; Revuelta, J.L. (Eds.) Industrial Biotechnology of Vitamins, Biopigments, and Antioxidants; Wiley-VCH: Weinheim, Germany, 2016; ISBN 9783527681754. [Google Scholar]
- CEIC. China Production: Year to Date: Vitamin E. Available online: https://www.ceicdata.com/en/china/pharmaceuticalproduction-ytd-antiparasitics-vitamins-and-minerals/cn-production-ytd-vitamin-e (accessed on 21 June 2022).
- Li, K.T.; Liu, D.H.; Li, Y.L.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. Improved Large-Scale Production of Vitamin B12 by Pseudomonas denitrificans with Betaine Feeding. Bioresour. Technol. 2008, 99, 8516–8520. [Google Scholar] [CrossRef]
- Li, K.T.; Liu, D.H.; Li, Y.L.; Chu, J.; Wang, Y.H.; Zhuang, Y.P.; Zhang, S.L. Influence of Zn2+, Co2+ and Dimethylbenzimidazole on Vitamin B12 Biosynthesis by Pseudomonas denitrificans. World J. Microbiol. Biotechnol. 2008, 24, 2525–2530. [Google Scholar] [CrossRef]
- Kusel, J.; Fa, Y.; Demain, A. Betaine Stimulation of Vitamin B12 Biosynthesis in Pseudomonas denitrificans May Be Mediated by an Increase in Activity of δ-Aminolaevulinic Acid Synthase. J. Gen. Microbiol. 1984, 130, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, J.Z.; Huang, J.S.; Mao, Z.W. Genome Shuffling of Propionibacterium shermanii for Improving Vitamin B12 Production and Comparative Proteome Analysis. J. Biotechnol. 2010, 148, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Escalante-Semerena, J.C. Single-Enzyme Conversion of FMNH2 to 5,6-Dimethylbenzimidazole, the Lower Ligand of B12. Proc. Natl. Acad. Sci. USA 2007, 104, 2921–2926. [Google Scholar] [CrossRef] [Green Version]
- Signorini, C.; Carpen, A.; Coletto, L.; Borgonovo, G.; Galanti, E.; Capraro, J.; Magni, C.; Abate, A.; Johnson, S.K.; Duranti, M.; et al. Enhanced Vitamin B12 Production in an Innovative Lupin Tempeh Is Due to Synergic Effects of Rhizopus and Propionibacterium in Cofermentation. Int. J. Food Sci. Nutr. 2018, 69, 451–457. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Frettloeh Martin Sequential Co-Culturing Method for Producing a Vitamin- and Protein-Rich Food Product. U.S. Patent 2020-0149084 A1, 14 May 2020.
- Quesada-Chanto, A.; Afschar, A.S.; Wagner, F. Microbial Production of Propionic Acid and Vitamin B12 Using Molasses or Sugar. Appl. Microbiol. Biotechnol. 1994, 41, 378–383. [Google Scholar] [CrossRef]
- Haddadin, M.S.Y.; Abu-Reesh, I.M.; Haddadin, F.A.S.; Robinson, R.K. Utilisation of Tomato Pomace as a Substrate for the Production of Vitamin B12—A Preliminary Appraisal. Bioresour. Technol. 2001, 78, 225–230. [Google Scholar] [CrossRef]
- Assis, D.A.D.; Matte, C.; Aschidamini, B.; Rodrigues, E.; Záchia Ayub, M.A. Biosynthesis of Vitamin B12 by Propionibacterium freudenreichii subsp. Shermanii ATCC 13673 Using Liquid Acid Protein Residue of Soybean as Culture Medium. Biotechnol. Prog. 2020, 36, e3011. [Google Scholar] [CrossRef]
- Gardner, N.; Champagne, C.P. Production of Propionibacterium Shermanii Biomass and Vitamin B12 on Spent Media. J. Appl. Microbiol. 2005, 99, 1236–1245. [Google Scholar] [CrossRef]
- Moine, G.; Hohmann, H.-P.; Kurth, R.; Paust, J.; Hähnlein, W.; Pauling, H.; Weimann, B.-J.; Kaesler, B. Vitamins, 6. B Vitamins. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH GmbH & Co. KGaA: Weinheim, Germany, 2011. [Google Scholar]
- Survase, S.A.; Bajaj, I.B.; Singhal, R.S. Biotechnological Production of Vitamins. Food Technol. Biotechnol. 2006, 44, 381–396. [Google Scholar]
- Nielsen, M.J.; Rasmussen, M.R.; Andersen, C.B.F.; Nexø, E.; Moestrup, S.K. Vitamin B12 Transport from Food to the Body’s Cells—A Sophisticated, Multistep Pathway. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 345–354. [Google Scholar] [CrossRef]
- Berlin, H.; Berlin, R.; Brante, G. Oral Treatment of Pernicious Anemia with High Doses of Vitamin B12 without Intrinsic Factor. Acta Med. Scand. 1968, 184, 247–258. [Google Scholar] [CrossRef]
- Gamboa, J.M.; Leong, K.W. In vitro and in vivo Models for the Study of Oral Delivery of Nanoparticles. Adv. Drug Deliv. Rev. 2013, 65, 800–810. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, S.P.; He, J. Microencapsulation of Vitamins in Food Applications to Prevent Losses in Processing and Storage: A Review. Food Res. Int. 2020, 137, 109326. [Google Scholar] [CrossRef]
- Matos, M.; Gutiérrez, G.; Iglesias, O.; Coca, J.; Pazos, C. Enhancing Encapsulation Efficiency of Food-Grade Double Emulsions Containing Resveratrol or Vitamin B12 by Membrane Emulsification. J. Food Eng. 2015, 166, 212–220. [Google Scholar] [CrossRef]
- Ramalho, M.J.; Loureiro, J.A.; Pereira, M.C. Poly (Lactic-Co-Glycolic Acid) Nanoparticles for the Encapsulation and Gastrointestinal Release of Vitamin B9 and Vitamin B12. ACS Appl. Nano Mater. 2021, 4, 6881–6892. [Google Scholar] [CrossRef]
- Bochicchio, S.; Barba, A.A.; Grassi, G.; Lamberti, G. Vitamin Delivery: Carriers Based on Nanoliposomes Produced via Ultrasonic Irradiation. LWT-Food Sci. Technol. 2016, 69, 9–16. [Google Scholar] [CrossRef]
- Estevinho, B.N.; Mota, R.; Leite, J.P.; Tamagnini, P.; Gales, L.; Rocha, F. Application of a Cyanobacterial Extracellular Polymeric Substance in the Microencapsulation of Vitamin B12. Powder Technol. 2019, 343, 644–651. [Google Scholar] [CrossRef]
- Carlan, I.C.; Estevinho, B.N.; Rocha, F. Study of Microencapsulation and Controlled Release of Modified Chitosan Microparticles Containing Vitamin B12. Powder Technol. 2017, 318, 162–169. [Google Scholar] [CrossRef]
- Fidaleo, M.; Tacconi, S.; Sbarigia, C.; Passeri, D.; Rossi, M.; Tata, A.M.; Dini, L. Current Nanocarrier Strategies Improve Vitamin B12 Pharmacokinetics, Ameliorate Patients’ Lives, and Reduce Costs. Nanomaterials 2021, 11, 743. [Google Scholar] [CrossRef]
- Long, R.A. Production of Vitamin B12. U.S. Patent 3,018,225, 23 January 1962. [Google Scholar]
- Crouzet, J.; Debussche, L.; Schil, S.L.; Thibaut, D. Methods of Increasing the Production of Cobalamins Using Cob Gene Expression. U.S. Patent 2006/0019352 A1, 26 January 2006. [Google Scholar]
- Zhendong, L.; Qing, C.; Weikai, X.; Hua, L.; Lulu, Y. Screening Method and Screening Culture Medium for Bacterial Strain for High Yield of Vitamin B12 Produced through Fermentation Production with Pseudomonas denitrificans. CN Patent 111254173 A, 9 June 2020. [Google Scholar]
- Yonggan, H.; Yanmao, L.; Hui, L.; Xin, W. Multi-Stage Rotating Speed Regulating Policy for Improving Pseudomonas denitrificans Fermentation for Production of Vitamin B12. CN Patent 108949866 A, 7 December 2018. [Google Scholar]
- Wang, J.; Wang, J.; Hanzhong, L.; Yanxia, C.; Mu, Q.; Yingran, L.; Xueran, L.; Jingkun, Z.; Hong, W.; Yuan, Y. Method for Improving Yield of Vitamin B12 Based of Regulation of Ammonia Nitrogen Index. CN Patent 110205350 A, 6 September 2019. [Google Scholar]
- Ruiyan, L.; Jiexi, M.; Wei, L.; Caixia, Z.; Fengyu, K. Method for Promoting Pseudomonas denitrificans to Generate Vitamin B12. CN Patent 109837320 A, 4 June 2019. [Google Scholar]
- Pengdong, H.; Lizhong, P.; Hongwei, G.; Yi, S.; Qiong, M. Preparation Method of Vitamin B12 Crude Product. CN Patent 111808158 A, 23 October 2020. [Google Scholar]
- Hunik, J.H. Process for the Production of Vitamin B12. U.S. Patent 6,492,141 B1, 10 December 2002. [Google Scholar]
- Michelle, I.; Adams, C. Probiotic Propionibacterium. U.S. Patent 7.427,397 B2, 23 September 2008. [Google Scholar]
- Gregor, K.; Stefan, F.; Mirjan, S.; Hrvoje, P. Co-Cultivation of Propionibacterium and Yeast. U.S. Patent 9,938,554, 10 April 2018. [Google Scholar]
- Zhiguo, W.Z.; Guoxia, X.; Liquan, W.; Yunshan, W.S. Device for Producing Propionic Acid and Co-Producing Vitamin B12 by Semi-Continuous Fermentation 2017. CN Patent 206828509 U, 2 January 2018. [Google Scholar]
- Mallinath, L.A.; Pillai, V. Continuous Process for Co-Production of Vitamin B12 and Organic Acids. IN Patent 201827044769, 2 October 2020. [Google Scholar]
- Garibaldi, J.A.; Kosuke, I.; Lewis, J.C.; Mcginnis, J. Fermentation Process for Production of Vitamin B12. U.S. Patent 2576,932, 4 December 1994. [Google Scholar]
- Mogna, G.; Paolo Strozzi, G.; Mogna, L. Vitamin B12 Producing Lactobacillus Reuteri Strains 201. International Patent 2011/154820 A2, 15 December 2011. [Google Scholar]
- Dawei, Z.; Miaomiao, X.; Sha, L.; Wenjuan, Z.; Ping, Z.; Huan, F. Sinorhizobium Meliloti Strain and Composition and Application of Sinorhizobium Meliloti Strain. CN Patent 104342390 A, 11 February 2015. [Google Scholar]
- Dawei, Z.; Huina, D. Procorrin-2C(20)-Methyltransferase Mutant and Mutant Gene and Application Thereof in Preparing Vitamin B12 2020. CN Patent 110804598 A, 18 February 2020. [Google Scholar]
- Zhang, D.; Fang, H. Recombinant Strain of Escherichia Coli for de Novo Synthesis of Vitamin B12, Construction Method Therefor and Application Thereof. International Patent 2019/109975A1, 13 June 2019. [Google Scholar]
- Bijl, H.L. Production and Use of Compositions Comprising High Concentrations of Vitamin B12 Activity. U.S. Patent 6,187,761 B1, 13 February 2001. [Google Scholar]
- Hugenschmidt, S.; Miescher Schwenninger, S.; Lacroix, C. Process for the Preparation of a Fermentation Broth. EU Patent 2 376 644 B1, 23 April 2014. [Google Scholar]
- Mansour, M. Modified Propionibacterium and Methods of Use. International Patent 2021/041759, 4 March 2021. [Google Scholar]
- Remy, E.; Examiner, P.; Achutamurthy, P. Biosynthesis Method Enabling the Preparation of Cobalamins. U.S. Patent 6,156,545, 5 December 2000. [Google Scholar]
- Zhang, S.; Zhao, Q.; Li, K.; Liu, D.; Hongzhuang, Y.; Wang, Z.; Li, Y. Method for Improving Yield of Denitrified Pseudomonas Vitamin B12. CN Patent 101538599 A, 23 September 2009. [Google Scholar]
- Zhang, S.; Chen, X.; Li, Y.; Zhenguo, W.; Xie, L.; Cao, Y.; Tang, L.; Wang, Z.; Zhuang, Y. Vitamin B12 Fermentation Production Control Process Based on CO2 Concentration in Tail Gas. CN Patent 102399845 A, 13 September 2010. [Google Scholar]
- Zhang, S.; Zhang, Y.; Wang, Z.; Wang, H.; Zhuang, Y.; Chu, J.; Siliang, Z.; Yiming, Z.; Yinping, Z.; Huiyuan, W.; et al. Optimized Method for Producing Vitamin B12 through Pseudomonas denitrificans Fermentation and Synthetic Medium. CN Patent 101748177 A, 9 December 2008. [Google Scholar]
- Wang, H.; Chu, J.; Wang, Z.; Zhuang, Y.; Zhang, S.; Zejian, W.; Huiyan, W.; Yingping, Z.; Siliang, Z.; Ju, C. Oxygen Consumption Rate-Based Vitamin B12 Fermentation Production Control Process. CN Patent 102021214 A, 22 September 2009. [Google Scholar]
- Ren, Y.; Leng, X.; Wang, Y.; Qi, N.; Dong, Y.; Xiaohong, L.; Nai, Q.; Yong, R.; Youshan, W.; Yuan, D. Culture Medium for Producing Vitamin B12 by Fermenting Pseudomonas denitrificans and Fermentation Method Thereof. CN Patent 102453740 A, 16 May 2012. [Google Scholar]
- Ju, Y.; Yang, H.; Yanmao, L.; Guangyong, Z. Method for Producing Vitamin B12 by Using Pseudomonas denitrificans Based on PH Value Control. CN Patent 108913739 A, 30 November 2018. [Google Scholar]
- Mcdaniel, L.E.; Harold, B. Cyanide Ion in Production of Vitamin B12. U.S. Patent 2,650,896, 1 September 1953. [Google Scholar]
- Takaaki, M.; Zhenya, Z. Method for Producing Vitamin B12 from Hydrogen-Metabolizing Methane Bacte-Rium. U.S. Patent 2005/0227332 A1, 13 October 2005. [Google Scholar]
- Barg, H.; Jahn, D. Method for the Production of Vitamin B12. U.S. Patent 2006/0105432 A1, 18 May 2006. [Google Scholar]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.D.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Safety and Efficacy of Vitamin B12 (in the Form of Cyanocobalamin) Produced by Ensifer spp. as a Feed Additive for All Animal Species Based on a Dossier Submitted by VITAC EEIG. EFSA J. 2018, 16, e05336. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, G.; Laganà, A.S.; Rapisarda, A.M.C.; la Ferrera, G.M.G.; Buscema, M.; Rossetti, P.; Nigro, A.; Muscia, V.; Valenti, G.; Sapia, F.; et al. Vitamin B12 among Vegetarians: Status, Assessment and Supplementation. Nutrients 2016, 8, 767. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, F.; Yabuta, Y.; Tanioka, Y.; Bito, T. Biologically Active Vitamin B12 Compounds in Foods for Preventing Deficiency among Vegetarians and Elderly Subjects. J. Agric. Food Chem. 2013, 61, 6769–6775. [Google Scholar] [CrossRef]
- Titcomb, T.J.; Tanumihardjo, S.A. Global Concerns with B Vitamin Statuses: Biofortification, Fortification, Hidden Hunger, Interactions, and Toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1968–1984. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Cave, G.; Lu, C. Vitamin B12 (Cobalamin) and Micronutrient Fortification in Food Crops Using Nanoparticle Technology. Front. Plant Sci. 2021, 12, 1451. [Google Scholar] [CrossRef]
- Pawlak, R.; Rusher, D.R. A Review of 89 Published Case Studies of Vitamin B12 Deficiency. J. Hum. Nutr. Food Sci. 2013, 1, 1008. [Google Scholar]
- Watkins, D.; Rosenblatt, D.S. Inborn Errors of Cobalamin Absorption and Metabolism. Am. J. Med. Genet. Part C Semin. Med. Genet. 2011, 157, 33–44. [Google Scholar] [CrossRef]
- Shepherd, G.; Velez, L.I. Role of Hydroxocobalamin in Acute Cyanide Poisoning. Ann. Pharmacother. 2008, 42, 661–669. [Google Scholar] [CrossRef]
- Linnell, J.C.; Smith, A.D.; Smith, C.L.; Wilson, M.J.; Matthews, D.M. Effects of Smoking on Metabolism and Excretion of Vitamin B12. BMJ 1968, 2, 215–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.; Rosenberg, I.; Oakley, G.; Omenn, S. Considering the Case for Vitamin B12 Fortification of Flour. Food Nutr. Bull. 2010, 31 (Suppl. S1), S36–S46. [Google Scholar] [CrossRef] [PubMed]
- Hamel, J. A Review of Acute Cyanide Poisoning with a Treatment Update. Crit. Care Nurse 2011, 31, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Reports and Data Vitamin B12 (Cobalamin) Market to Reach USD 409.7 Million By 2027. Available online: https://www.globenewswire.com/news-release/2020/08/26/2084313/0/en/Vitamin-B12-Cobalamin-Market-To-Reach-USD-409-7-Million-By-2027-Reports-and-Data.html (accessed on 22 April 2022).
- Rabah, H.; Rosa do Carmo, F.L.; Jan, G. Dairy Propionibacteria: Versatile Probiotics. Microorganisms 2017, 5, 24. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Li, Y. (Eds.) Atlas of Oral Microbiology: From Healthy Microflora to Disease; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Acevedo-Rocha, C.G.; Gronenberg, L.S.; Mack, M.; Commichau, F.M.; Genee, H.J. Microbial Cell Factories for the Sustainable Manufacturing of B Vitamins. Curr. Opin. Biotechnol. 2019, 56, 18–29. [Google Scholar] [CrossRef]



| Microorganism/Strain | Main Media Components | Scale | Summary/Innovation | Volumetric Production | Volumetric Productivity (mg/L/h) | Reference |
|---|---|---|---|---|---|---|
| B. megaterium DSM 319 | Terrific broth media | 250 mL shake flask | Precursor supplementation and pO2 control | 0.21 mg/L c | 0.006 mg/L/h c | [47] |
| Lactobacillus reuteri ZJ03 | Soymilk | 250 mL shake flask | Different carbon source supplementation | 0.204 mg/L | 0.003 mg/L/h | [49] |
| P. freudenreichii subsp. shermanii NRRL-B-4327, 3523 and NRRL-B-3524 | Sodium lactate broth | 250 mL shake flask | Vitamin B12 analogue addition | 31 mg/L | 0.51 mg/L | [50] |
| P. freudenreichii CICC 10019 | Glucose, CSL a | 7 L stirred tankbioreactor | Expanded-bed bioreactor (EBAB) with crop stark hydrolysates | 47.6 mg/L | 0.18 mg/L/h | [51] |
| P. freudenreichii CICC 10019 | Glucose, CSL | 7 L stirred tank bioreactor | EBAB bioreactor | 43.4 mg/L | 0.27 mg/L/h | [52] |
| P. freudenreichii CICC 10019 | Glucose, CSL | 1.5 L stirred tank bioreactor | EBAB bioreactor and DMBI addition | 58.8 mg/L | 0.59 mg/L/h | [53] |
| P. freudenreichii CICC 10019 | Glucose/glycerol, CSL | 5 L stirred tank bioreactor | EBAB bioreactor, glycerol as carbon source and crop stalk hydrolysate as nitrogen source | 43 mg/L | 0.36 mg/L/h | [54] |
| P. freudenreichii DF13 | Supplemented whey permeate | 1 L stirred tank bioreactor | Co-culture with Lactobacillus plantarum SM39 for simultaneous folate and Cbl production | 0.75 mg/L | 0.004 mg/L/h | [55] |
| P. freudenreichii DSM 20271//Lactobacillus brevis ATCC 14869 | Wheat bran dough | n.d. b | Co-fermentation in wheat bran dough for in situ production of Vitamin B12 | 332 ng/g c | n.d. b | [56] |
| P. freudenreichii IFO 12424//Ralstonia eutropha H16 (ATCC17699) | Polypeptone, casein, yeast extract | 5 L stirred tank bioreactor | Cell recycling system and co-culture with Ralstonia eutropha for decreasing propionic acid inhibition | 8 mg/L c | 0.14 mg/L/h c | [57] |
| P. freudenreichii PTCC 1674. | Tryptone, yeast extract, different carbon sources | 100 cm3 | Waste frying sun oil as a carbon source for vitamin B12 production | 2.74 mg/L | 0.02 mg/L/h | [58] |
| P. freudenreichii subsp. shermanii ATCC 13673 | Glucose, yeast extract | 2 L stirred tank bioreactor | Inoculum volume, pH control and substrate concentration optimization | 0.087 mg/L | 0.002 mg/L/h | [59] |
| P. freudenreichii subsp. shermanii CICC 10019 | Glucose, CSL | 100 L fermenter | Addition of DMBI precisely with Ado-Cbl control strategy | 39.15 mg/L | 0.32 mg/L/h | [60] |
| P. freudenreichii subsp. shermanii | Glycerol, tryptone, casein, DMBI | 200 mL shake flask | Media optimization by design of experiments with crude glycerol as the main carbon source | 4.01 mg/L | 0.024 mg/L/h | [61] |
| P. freudenreichii subsp. shermanii | Whey based media | 20 mL tubes | DMBI, Nicotinamide and Riboflavin supplementation | 5.3 mg/L | 0.03 mg/L/h | [62] |
| P. freudenreichii subsp. shermanii | Food-like media (cereal matrices) | n.d. b | Precursor supplementation in different cereal-like matrices | 1.5 mg/Kg | 0.009 mg/Kg/h | [63] |
| P. freudenreichii subsp. shermanii 2067 | Cheese-based propionic media/whey-based liquid media | 50 mL shake flask | Production in food-like conditions without DMBI addition | 0.124 mg/L c | 0.0013 mg/L/h | [64] |
| P. freudenreichii CICC10019 | Glucose, yeast extract, CSL | 100 mL flasks | Media optimization by statistical analysis | 8.32 mg/L | 0.068 mg/L/h | [65] |
| P. freudenreichii CICC10019 | Glucose, CSL | 7 L fermenter | Membrane separation-coupled fed-batch fermentation | 21.6 mg/L | 0.16 mg/L/h | [66] |
| P. denitrificans | Maltose, peptone, betaine | 250 mL shake flask | Addition of rotenone as a respiration inhibitor for enhanced production | 54.7 mg/L | 0.57 mg/L/h | [67] |
| P. denitrificans | Beet molasses, sucrose, betaine | 120 m3 fermenter | Glucose-betaine feeding, pH control strategy | 214.13 mg/L c | 1.27 mg/L/h c | [68] |
| P. denitrificans | Glucose, CSL, betaine | 120 m3 fermenter | Stepwise oxygen uptake rate control strategy | 188 mg/L | 1.12 mg/L/h | [69] |
| P. denitrificans | Glucose, CSL, betaine | 50 L fermenter | Effects of specific oxygen consumption rate on cell morphology and production | 213.1 mg/L | 1.88 mg/L/h | [70] |
| P. denitrificans | Maltose, peptone, betaine | 250 mL shake flask | Betaine supplementation | 58.61 mg/L | 0.48 mg/L/h | [71] |
| P. denitrificans | Maltose syrup, CSL, betaine | 120 m3 fermenter | Maltose syrup and CSL as the main substrates | 198.27 mg/L | 1.10 mg/L/h | [72] |
| P. denitrificans | Glucose, CSL, betaine | 120 m3 fermenter | pO2 stepwise control | 198.80 mg/L | 1.18 mg/L/h | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvillo, Á.; Pellicer, T.; Carnicer, M.; Planas, A. Bioprocess Strategies for Vitamin B12 Production by Microbial Fermentation and Its Market Applications. Bioengineering 2022, 9, 365. https://doi.org/10.3390/bioengineering9080365
Calvillo Á, Pellicer T, Carnicer M, Planas A. Bioprocess Strategies for Vitamin B12 Production by Microbial Fermentation and Its Market Applications. Bioengineering. 2022; 9(8):365. https://doi.org/10.3390/bioengineering9080365
Chicago/Turabian StyleCalvillo, Álvaro, Teresa Pellicer, Marc Carnicer, and Antoni Planas. 2022. "Bioprocess Strategies for Vitamin B12 Production by Microbial Fermentation and Its Market Applications" Bioengineering 9, no. 8: 365. https://doi.org/10.3390/bioengineering9080365