Abstract
Non-native and invasive tamarisk (Tamarix spp.) and Russian olive (Elaeagnus angustifolia) are common in riparian areas of the Colorado River Basin and are regarded as problematic by many land and water managers. Widespread location data showing current distribution of these species, especially data suitable for remote sensing analyses, are lacking. This dataset contains 3476 species occurrence and absence point records for tamarisk and Russian olive along rivers within the Colorado River Basin in Arizona, California, Colorado, Nevada, New Mexico, and Utah. Data were collected in the field in the summer of 2017 with high-resolution imagery loaded on computer tablets. This dataset includes status (live, dead, defoliated, etc.) of observed tamarisk to capture variability in tamarisk health across the basin, in part attributable to the tamarisk beetle (Diorhabda spp.). For absence points, vegetation or land cover were recorded. These data have a range of applications including serving as a baseline for the current distribution of these species, species distribution modeling, species detection with remote sensing, and invasive species management.
Dataset: Dataset submitted as supplement to this paper.
Dataset License: CC BY 4.0.
1. Summary
Non-native and invasive tamarisk (saltcedar, Tamarix spp.) and Russian olive (Elaeagnus angustifolia) have become some of the most common and abundant woody riparian species across the western United States [1,2]. These species receive particular attention from land and water managers in the Colorado River Basin due to their potential impacts on native riparian vegetation [3,4,5], geomorphology [6], water resources [7], recreation, and wildlife [8,9]. Tamarisk and Russian olive were introduced and widely planted for erosion control, wind breaks, and landscaping in the 19th and 20th centuries and escaped cultivation in the early and mid-20th century [2]. Both species can either form dominant monocultures or can coexist with native riparian vegetation [10,11]. They are generally found in moist areas such as around rivers, irrigation channels, lakes, and reservoirs.
Concerns about the negative impacts of these invasive species have led to their active removal in many areas and the introduction of the tamarisk beetle (Diorhabda spp.) as biocontrol. The tamarisk beetle was first approved for release outside of experimental settings in 2001 and has since become widespread [12]. The tamarisk beetle is now established throughout the Western U.S. ranging from Texas, Oklahoma, and Kansas in the east to California in the west and from Mexico in the south to Oregon, Idaho, and Wyoming in the north [12]. These beetles exhibit a cyclic pattern with tamarisk whereby they defoliate tamarisk initially and then return after tamarisk has recovered to defoliate the plant again. This repeated defoliation within a growing season, or over several growing seasons, has resulted in high tamarisk mortality in some areas, but minimal mortality at other sites [13,14]. Due to this biocontrol, removal efforts, and the spread of tamarisk and Russian olive, their distributions are changing—data documenting their locations and condition are critical for monitoring and mapping efforts.
Broad-scale maps of tamarisk and Russian olive are needed to inform management and our understanding of these invasions [2]. We set out to map tamarisk and Russian olive distributions in the Colorado River Basin, but found existing field data to be inadequate for the task. Existing tamarisk and Russian olive distribution data are limited in their geographic extent—the data are concentrated in some areas and sparse or nonexistent in other areas. Another challenge we encountered is that presence points are frequently incompatible for analyses with remotely-sensed imagery—georegistration errors may be large (e.g., a point is placed on a road adjacent to a tamarisk stand rather than in the tamarisk stand itself) and there may not be a minimum cover or abundance requirement to be included as a presence (e.g., a single tamarisk shoot in an area dominated by other vegetation may be marked as a presence point). These points are well-suited for some applications such as habitat suitability modeling [15,16], but are inadequate when pairing field data with high (e.g., National Agriculture Imagery Program) or moderate-resolution (e.g., Landsat, Sentinel-2) remotely-sensed imagery to detect a specific species. Timing between field data collection and remotely-sensed imagery collection needs to align when mapping species to ensure accurate representation of the current extents of these species and to not confound the analysis with outdated records. Furthermore, tamarisk and/or Russian olive absence points are often not included in datasets but are important for developing models to map species’ locations. The data presented in this article address these issues and are highly suitable for remote sensing analyses which are often not possible with records in existing databases containing invasive species occurrence points (e.g., the Early Detection and Distribution Mapping System (EDDMapS), the Global Biodiversity Information Facility (GBIF)).
To address this need, we designed and implemented a sampling protocol that utilizes high-resolution imagery and computer tablets to efficiently collect data compatible with remote sensing across large areas (i.e., multiple states). We collected 3476 presence and absence locations of tamarisk and Russian olive in the Colorado River Basin across six states. These data are compatible with moderate-resolution satellite sensors like Landsat and Sentinel-2 and could potentially be used to map these species across large areas. This dataset in Supplementary Materials can also serve as a snapshot of tamarisk and Russian olive distribution in the summer of 2017, from which we can better understand change in distribution over time and tamarisk beetle impacts.
2. Data Description
This dataset in Supplementary Materials provides information on the presence and absence of tamarisk and Russian olive in select tributaries in the Colorado River Basin. The basin extends over about 630,000 km2 in Western North America from the Rocky Mountains to the Gulf of Mexico (Figure 1). Our data span Arizona, California, Colorado, Nevada, New Mexico, and Utah. Data were collected during summer 2017 in the field using high-resolution imagery [17] on a digital collection interface along the Animas, Colorado, Dolores, Escalante, Fremont, Gila, Paria, San Juan, San Miguel, San Pedro, San Rafael, Verde, Virgin, Little Colorado, and Santa Clara Rivers and McElmo Creek. Approximately 3500 presence and absence locations are available as Global Positioning System (GPS) coordinates (“x” and “y” columns) in World Geodetic System 1984 (WGS 84) decimal degrees in three formats: shapefile, Keyhole Markup Language (KMZ), and comma-separated values (CSV). The classification of each point in the “Veg_LandCover” column represents the dominant (>50% of the cover) vegetation or cover type within a roughly 7 m radius circle. This column describes the status (live, dead, defoliated, etc.) of observed tamarisk for tamarisk points, and the vegetation or land cover of locations collected as tamarisk absences. Two additional columns, “TamPresAbs” and “ROPresAbs”, indicate if tamarisk or Russian olive, respectively, are dominant (“present”) or not (“absent”). Any tamarisk point, regardless of the status, is considered as “Present” in the “TamPresAbs” column. Each of these data attributes are described in the dataset’s accompanying metadata.
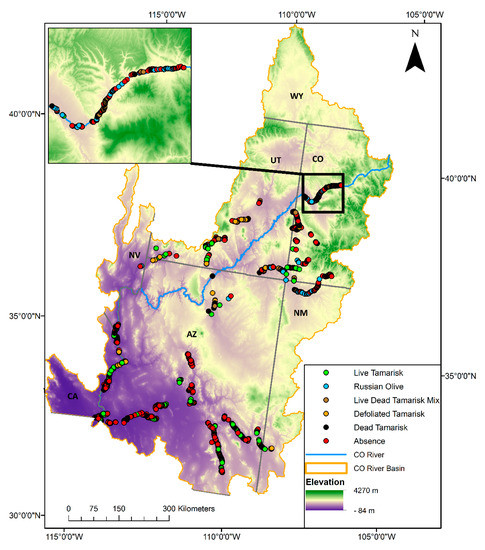
Figure 1.
The extent of the Colorado River Basin in the southwestern United States and locations of the samples included in this dataset. The inset shows data collected along the Colorado River in western Colorado. State labels correspond to the following states: AZ = Arizona; CA = California; CO = Colorado; NV = Nevada; NM = New Mexico; UT = Utah; WY = Wyoming.
3. Methods
We collected these data in July and August of 2017 for use with remotely sensed data to map tamarisk and Russian olive distribution in the Colorado River Basin (Figure 1). The sampling protocol was designed to be compatible with satellite imagery and for efficient data collection across large geographic areas. The most recent high-resolution imagery base maps [17] were loaded on Samsung Galaxy A electronic tablets in the Collector for ArcGIS program [18] and observers placed and classified points on this imagery as they viewed vegetation in the field, typically from a distance. Observers matched vegetation seen in the field with color, structure, and texture patterns observed in the high-resolution imagery to place points in the correct location and classify vegetation. Sampling was conducted opportunistically where land ownership and road access were allowed. Most points were collected from vehicles along roads for efficiency.
Points were classified based on their dominant vegetation to capture locations of tamarisk and Russian olive, and absences (Figure 2). Dominance was defined as occupying greater than 50% of the cover as seen from above within a roughly 7 m radius circle [19]. This plot size was selected based on its use across a broad range of agencies, and its utility in training models fit with remotely sensed imagery [20]. Most of the sampling was done quickly from vehicles, so determination of dominance was based on ocular estimates from a distance. The absence designation indicates that tamarisk and/or Russian olive were absent from or composed a minor portion of the overstory, but should not be interpreted as complete absence from that location since many of these sites were observed from a distance.
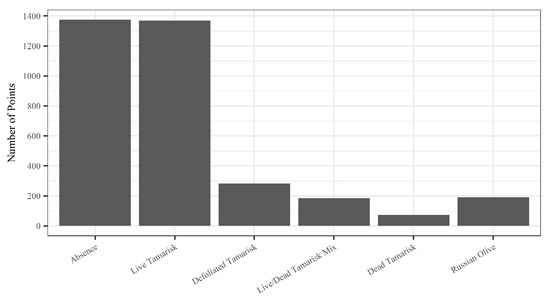
Figure 2.
The number of points by vegetation class for a total of 3476 plots.
Tamarisk health varies broadly across the Colorado River Basin, largely due to defoliation by the tamarisk beetle [12]. We collected information to classify the predominant status of tamarisk as either live, dead (leafless with no obvious evidence of live growth as determined from distance sampling), a mixture of live and dead, or defoliated (having a reddish appearance from tamarisk beetle defoliation and desiccation; Figure 3). We classified the absence points to indicate the dominant vegetation or land cover (i.e., water, bare ground, etc.) at that location. Most points are classified to genus, but some are identified to the species, family, or land cover type. Our distance sampling method did not allow for the identification of tamarisk to the species level, however Tamarix ramosissima and T. chinensis and their hybrids are the most common species in the western U.S. [1,2,21]. Additional species known to be established in the U.S. that we may have encountered in our sampling include T. aphylla, T. parviflora, T. canariensis, and T. gallica [1,2,21]. At the beginning of sampling, points not classified as tamarisk or Russian olive were not labeled with the vegetation/land cover—these points were labeled as an “unclassified absence”. Sample points were placed in vegetation and land cover types representative of the area being sampled, but sampling frequency was not necessarily proportional to the area covered by a given vegetation or land cover. Data were checked in the office and the “Veg_LandCover” column was edited so that naming conventions were consistent for equivalent vegetation and cover types. This dataset in Supplementary Materials provides valuable information on the 2017 distribution and status of tamarisk and Russian olive in the Colorado River Basin.

Figure 3.
An example of reddish tamarisk in the foreground that has been impacted by tamarisk beetles. It would be classified as “defoliated” in this dataset. Photo credit: Dr. Amanda West.
Supplementary Materials
The following are available online at http://www.mdpi.com/2306-5729/3/4/42/s1, Tamarisk and Russian olive occurrence and absence point data in three formats: shapefile, KMZ, and CSV.
Author Contributions
Conceptualization and methodology, A.G.V., B.D.W., A.M.W., N.E.Y. and P.H.E.; data acquisition, A.G.V., A.M.W., R.G.S. and T.J.M.; data curation, A.G.V., A.M.W. and R.K.G.; writing—original draft preparation, A.G.V. and B.D.W.; writing—review & editing, all authors; visualization, B.D.W. and A.M.W.; funding acquisition, P.H.E.
Funding
This research was funded by the Walton Family Foundation and the U.S. Geological Survey, grant number 006194-00002.
Acknowledgments
The authors thank Shannon Hatch, Nate Higginson, Melissa McMaster, Patrice Mutchnick, Anna Schrenk, David Varner, and Mike Wight for their helpful site descriptions that helped guide field sampling.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Friedman, J.M.; Auble, G.T.; Shafroth, P.B.; Scott, M.L.; Merigliano, M.F.; Freehling, M.D.; Griffin, E.R. Dominance of non-native riparian trees in western USA. Biol. Invasions 2005, 7, 747–751. [Google Scholar] [CrossRef]
- Nagler, P.L.; Glenn, E.P.; Jarnevich, C.S.; Shafroth, P.B. Distribution and abundance of Saltcedar and Russian Olive in the Western United States. Crit. Rev. Plant Sci. 2011, 30, 508–523. [Google Scholar] [CrossRef]
- Carman, J.G.; Brotherson, J.D. Comparisons of Sites Infested and Not Infested with Saltcedar (Tamarix pentandra) and Russian Olive (Elaeagnus angustifolia). Weed Sci. 1982, 30, 360–364. [Google Scholar] [CrossRef]
- Sher, A.A.; Marshall, D.L.; Taylor, J.P. Establishment patterns of native Populus and Salix in the presence of invasive nonnative Tamarix. Ecol. Appl. 2002, 12, 760–772. [Google Scholar] [CrossRef]
- Meinhardt, K.A.; Gehring, C.A. Disrupting mycorrhizal mutualisms: A potential mechanism by which exotic tamarisk outcompetes native cottonwoods. Ecol. Appl. 2012, 22, 532–549. [Google Scholar] [CrossRef] [PubMed]
- Stromberg, J.C. Functional equivalency of saltcedar (Tamarix chinensis) and fremont cottonwood (Populus fremontii) along a free-flowing river. Wetlands 1998, 18, 675–686. [Google Scholar] [CrossRef]
- Nagler, P.L.; Morino, K.; Didan, K.; Erker, J.; Osterberg, J.; Hultine, K.R.; Glenn, E.P. Wide-area estimates of saltcedar (Tamarix spp.) evapotranspiration on the lower Colorado River measured by heat balance and remote sensing methods. Ecohydrology 2009, 2, 18–33. [Google Scholar] [CrossRef]
- Fischer, R.A.; Valente, J.J.; Guilfoyle, M.P.; Michael, D.; Jackson, S.S.; Ratti, J.T. Bird Community Response to Vegetation Cover and Composition in Riparian Habitats Dominated by Russian Olive (Elaeagnus angustifolia). Northwest Sci. 2012, 86, 39–52. [Google Scholar] [CrossRef]
- Nelson, S.M.; Wydoski, R. Butterfly Assemblages Associated with Invasive Tamarisk (Tamarix spp.) Sites: Comparisons with Tamarisk Control and Native Vegetation Reference Sites. J. Insects 2013, 2013, 10. [Google Scholar] [CrossRef]
- Katz, G.L.; Shafroth, P.B. Biology, ecology and management of Elaeagnus angustifolia L. (Russian olive) in western North America. Wetlands 2003, 23, 763–777. [Google Scholar] [CrossRef]
- Merritt, D.M.; Leroy Poff, N. Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol. Appl. 2010, 20, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Bloodworth, B.R.; Shafroth, P.B.; Sher, A.A.; Manners, R.B.; Bean, D.W.; Johnson, M.J.; Hinojosa-Huerta, O. Tamarisk beetle (Diorhabda spp.) in the Colorado River Basin: Synthesis of an Expert Panel Forum; Scientific and Technical Report No. 1; Colorado Mesa University Ruth Powell Hutchins Water Center: Grand Junction, CO, USA, 2016. [Google Scholar]
- Hultine, K.R.; Dudley, T.L.; Koepke, D.F.; Bean, D.W.; Glenn, E.P.; Lambert, A.M. Patterns of herbivory-induced mortality of a dominant non-native tree/shrub (Tamarix spp.) in a southwestern US watershed. Biol. Invasions 2015, 17, 1729–1742. [Google Scholar] [CrossRef]
- Kennard, D.; Louden, N.; Gemoets, D.; Ortega, S.; González, E.; Bean, D.; Cunningham, P.; Johnson, T.; Rosen, K.; Stahlke, A. Tamarix dieback and vegetation patterns following release of the northern tamarisk beetle (Diorhabda carinulata) in western Colorado. Biol. Control 2016, 101, 114–122. [Google Scholar] [CrossRef]
- Morisette, J.T.; Jarnevich, C.S.; Ullah, A.; Cai, W.; Pedelty, J.A.; Gentle, J.E.; Stohlgren, T.J.; Schnase, J.L. A tamarisk habitat suitability map for the continental United States. Front. Ecol. Environ. 2006, 4, 11–17. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Reynolds, L.V. Challenges of predicting the potential distribution of a slow-spreading invader: A habitat suitability map for an invasive riparian tree. Biol. Invasions 2010, 13, 153–163. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ESRI). World Imagery; ESRI: Redlands, CA, USA, 2017. [Google Scholar]
- Environmental Systems Research Institute (ESRI). Collector for ArcGIS Desktop; ESRI: Redlands, CA, USA, 2016. [Google Scholar]
- Stohlgren, T.J.; Barnett, D.T.; Simonson, S.E. Beyond North American Weed Management Association Standards. 2010. Available online: http://irmaservices.nps.gov/datastore/v4/rest/DownloadFile/434136?accessType=DOWNLOAD (accessed on 17 October 2018).
- West, A.M.; Evangelista, P.H.; Jarnevich, C.S.; Kumar, S.; Swallow, A.; Luizza, M.; Chignell, S. Using multi-date satellite imagery to monitor invasive grass species distribution in post-wildfire landscapes; an iterative, adaptable approach that employs open-source data and software. Int. J. Appl. Earth Obs. Geoinf. 2017, 59, 135–146. [Google Scholar] [CrossRef]
- Gaskin, J.F.; Schaal, B.A. Hybrid Tamarix widespread in U.S. invasion and undetected in native Asian range. Proc. Natl. Acad. Sci. USA 2002, 99, 11256–11259. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).