
Challenging the Hypothesis of in Utero Microbiota Acquisition in Healthy Canine and Feline Pregnancies at Term: Preliminary Data
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cesarean Sections and Sample Collection
2.3. Bacterial Culture
2.4. Sequencing of 16S rRNA Gene
2.5. Data Analysis
3. Results
3.1. Canine Feto-Maternal Units: Bacterial Culture
3.2. Feline Feto-Maternal Units: Bacterial Culture
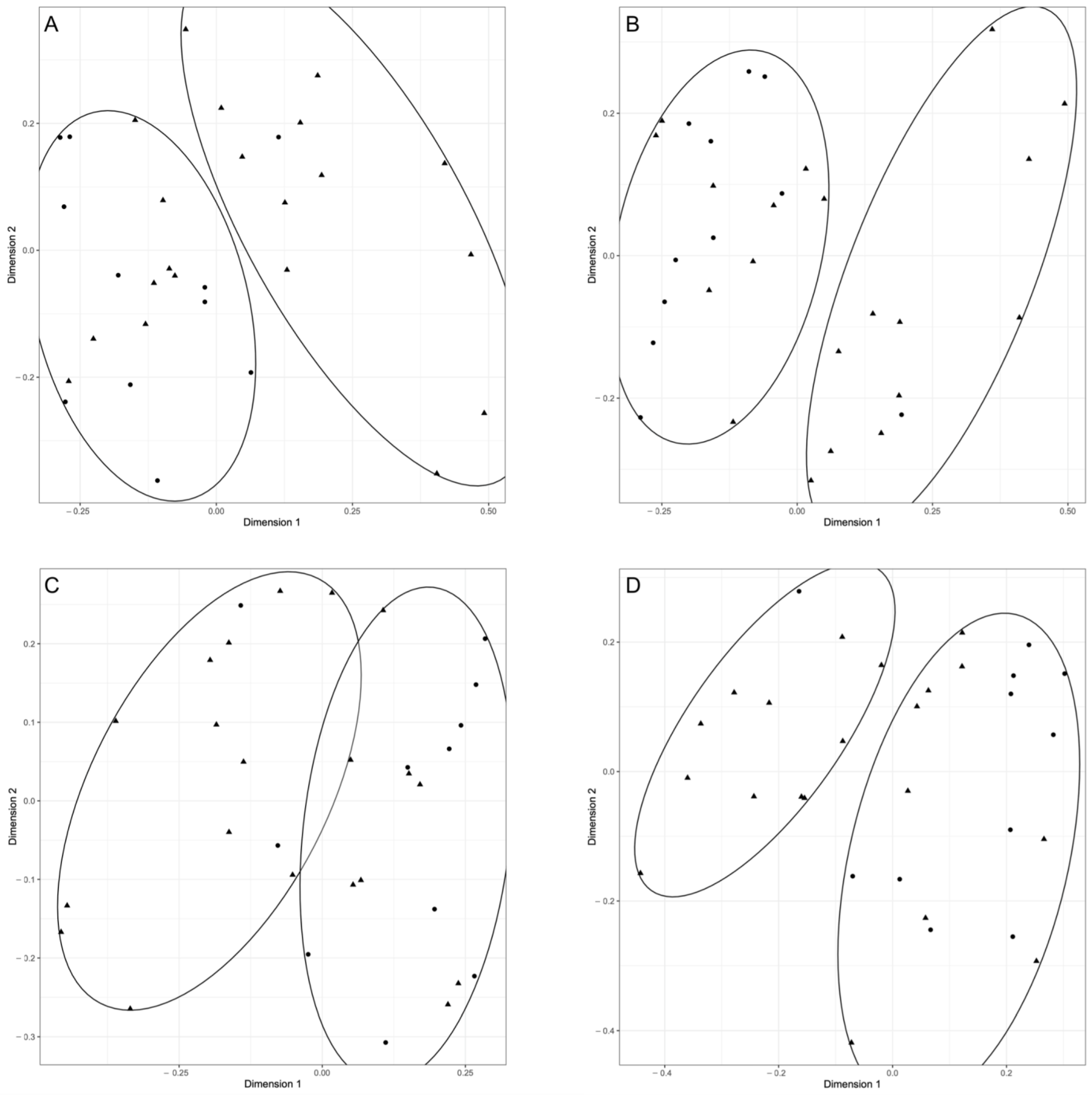
3.3. Canine and Feline Feto-Maternal Units: Sequencing of 16s RNA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Muñoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed]
- Campisciano, G.; Quadrifoglio, M.; Comar, M.; Seta, F.D.; Zanotta, N.; Ottaviani, C.; Stampalija, T. Evidence of bacterial DNA presence in chorionic villi and mniotic fluid in the first and second trimester of pregnancy. Future Microbiol. 2021, 16, 1152. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Kwok, L.-Y.; Xi, X.; Zhong, Z.; Ma, T.; Xu, H.; Meng, H.; Zhao, F.; Zhang, H. The meconium microbiota shares more features with the amniotic fluid microbiota than the maternal fecal and vaginal microbiota. Gut Microbes 2020, 12, 1794266. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S.; Aakko, J.; Isolauri, E.; Salminen, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef]
- Younge, N.; McCann, J.R.; Ballard, J.; Plunkett, C.; Akhtar, S.; Araújo-Pérez, F.; Murtha, A.; Brandon, D.; Seed, P.C. Fetal exposure to the maternal microbiota in humans and mice. JCI Insight 2019, 4, e127806. [Google Scholar] [CrossRef]
- Husso, A.; Lietaer, L.; Pessa-Morikawa, T.; Grönthal, T.; Govaere, J.; Van Soom, A.; Iivanainen, A.; Opsomer, G.; Niku, M. The Composition of the Microbiota in the Full-Term Fetal Gut and Amniotic Fluid: A Bovine Cesarean Section Study. Front. Microbiol. 2021, 12, 626421. [Google Scholar] [CrossRef]
- Rota, A.; Del Carro, A.; Bertero, A.; Del Carro, A.; Cucuzza, A.S.; Banchi, P.; Corrò, M. Does Bacteria Colonization of Canine Newborns Start in the Uterus? Animals 2021, 11, 1415. [Google Scholar] [CrossRef]
- Hemberg, E.; Einarsson, S.; Kútvölgyi, G.; Lundeheim, N.; Bagge, E.; Båverud, V.; Jones, B.; Morrell, J. Occurrence of bacteria and polymorphonuclear leukocytes in fetal compartments at parturition; relationships with foal and mare health in the peripartum period. Theriogenology 2015, 84, 163–169. [Google Scholar] [CrossRef]
- Wang, H.; Yang, G.X.; Hu, Y.; Lam, P.; Sangha, K.; Siciliano, D.; Swenerton, A.; Miller, R.; Tilley, P.; Von Dadelszen, P.; et al. Comprehensive human amniotic fluid metagenomics supports the sterile womb hypothesis. Sci. Rep. 2022, 12, 6875. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Zhu, B.; Zhao, H.; Ai, Q.; Tong, Y.; Qin, S.; Feng, Y.; Wang, Y.; Wang, S.; et al. Midtrimester amniotic fluid from healthy pregnancies has no microorganisms using multiple methods of microbiologic inquiry. Am. J. Obstet. Gynecol. 2020, 223, 248.e1–248.e21. [Google Scholar] [CrossRef]
- Turunen, J.; Tejesvi, M.V.; Paalanne, N.; Hekkala, J.; Lindgren, O.; Kaakinen, M.; Tapiainen, T. 2021 Presence of distinctive microbiome in the first-pass meconium of newborn infants. Sci. Rep. 2021, 11, 19449. [Google Scholar] [CrossRef]
- Kennedy, K.M.; Gerlach, M.J.; Adam, T.; Heimesaat, M.M.; Rossi, L.; Surette, M.G. Fetal meconium does not have a detectable microbiota before birth. Nat. Microbiol. 2021, 6, 865–873. [Google Scholar] [CrossRef]
- Malmuthuge, N.; Griebel, P.J. Fetal environment and fetal intestine are sterile during the third trimester of pregnancy. Vet. Immunol. Immunopathol. 2018, 204, 59–64. [Google Scholar] [CrossRef]
- Theis, K.R.; Romero, R.; Greenberg, J.M.; Winters, A.D.; Garcia-Flores, V.; Motomura, K.; Ahmad, M.M.; Galaz, J.; Arenas-Hernandez, M.; Gomez-Lopez, N. No Consistent Evidence for Microbiota in Murine Placental and Fetal Tissues. mSphere 5 2020, 5, e00933-19. [Google Scholar] [CrossRef]
- Winters, A.D.; Romero, R.; Greenberg, J.M.; Galaz, J.; Shaffer, Z.D.; Garcia-Flores, V. Does the Amniotic Fluid of Mice Contain a Viable Microbiota? Front. Immunol. 2022, 13, 820366. [Google Scholar] [CrossRef]
- Kennedy, K.M.; de Goffau, M.C.; Perez-Muñoz, M.E.; Arrieta, M.-C.; Bäckhed, F.; Bork, P.; Braun, T.; Bushman, F.D.; Dore, J.; de Vos, W.M.; et al. Questioning the fetal microbiome illustrates pitfalls of low-biomass microbial studies. Nature 2023, 613, 639–649. [Google Scholar] [CrossRef]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef]
- Zakošek Pipan, M.; Kajdič, L.; Kalin, A.; Plavec, T.; Zdovc, I. Do newborn puppies have their own microbiota at birth? Influence of type of birth on newborn puppy microbiota. Theriogenology 2020, 152, 18–28. [Google Scholar] [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. Isolating “uncultivable” microorganisms in pure culture in a simulated natural environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef]
- Stinson, L.F.; Boyce, M.C.; Payne, M.S.; Keelan, J.A. The Not-so-Sterile Womb: Evidence That the Human Fetus Is Exposed to Bacteria Prior to Birth. Front. Microbiol. 2019, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- Ekenstedt, K.J.; Crosse, K.R.; Risselada, M. Canine Brachycephaly: Anatomy, Pathology, Genetics and Welfare. J. Comp. Pathol. 2020, 176, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Schrank, M.; Sozzi, M.; Mollo, A. Prevalence of cesarean sections in swiss Bernese Mountain Dogs (2001–2020) and identification of risk factors. Acta Vet. Scand. 2022, 64, 42. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Artusi, E.; Drigo, M.; Mateus, L.; Siena, G.; Gelli, D.; Falomo, M.E.; Romagnoli, S. Ultrasonographic analysis of fetal gastrointestinal motility during the peripartum period in the dog. Anim. Reprod. Sci. 2020, 219, 106514. [Google Scholar] [CrossRef]
- Nöthling, J.O.; De Cramer, K.G.M. Comparing the values of progesterone in the blood of bitches as measured with an chemiluminescence immunoassay and a radioimmunoassay. Reprod. Domest. Anim. 2018, 53, 1136–1141. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef]
- Du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Tsuchida, S.; Umemura, H.; Nakayama, T. Current Status of Matrix-Assisted Laser Desorption/Ionization-Time-of-Flight Mass Spectrometry (MALDI-TOF MS) in Clinical Diagnostic Microbiology. Molecules 2020, 25, 4775. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Hartigan, J.A.; Wong, M.A. Algorithm AS 136: A K-means clustering algorithm. Appl. Stat. 1979, 28, 100–108. [Google Scholar] [CrossRef]
- Dargère, S.; Cormier, H.; Verdon, R. Contaminants in blood cultures: Importance, implications, interpretation and prevention. Clin. Microbiol. Infect. 2018, 24, 964–969. [Google Scholar] [CrossRef]
- Tega, L.; Raieta, K.; Ottaviani, D.; Russo, G.L.; Blanco, G.; Carraturo, A. Catheter-related bacteremia and multidrug-resistant Acinetobacter lwoffii. Emerg. Infect. Dis. 2007, 13, 355–356. [Google Scholar] [CrossRef]
- Urban, C.; Segal-Maurer, S.; Rahal, J.J. Considerations in control and treatment of nosocomial infections due to multidrug-resistant Acinetobacter baumannii. Clin. Infect. Dis. 2003, 36, 1268–1274. [Google Scholar] [CrossRef]
- Crone, S.; Vives-Flórez, M.; Kvich, L.; Saunders, A.M.; Malone, M.; Nicolaisen, M.H.; Martínez-García, E.; Rojas-Acosta, C.; Gomez-Puerto, M.C.; Calum, H.; et al. The environmental occurrence of Pseudomonas aeruginosa. APMIS 2020, 128, 220–231. [Google Scholar] [CrossRef]
- Wong, V.; Levi, K.; Baddal, B.; Turton, J.; Boswell, T.C. Spread of Pseudomonas fluorescens due to contaminated drinking water in a bone marrow transplant unit. J. Clin. Microbiol. 2011, 49, 2093–2096. [Google Scholar] [CrossRef]
- Mols, K.L.; Boe-Hansen, G.B.; Mikkelsen, D.; Bryden, W.L.; Cawdell-Smith, A.J. Prenatal establishment of the foal gut microbiota: A critique of the in utero colonisation hypothesis. Anim. Prod. Sci. 2020, 60, 2080–2092. [Google Scholar] [CrossRef]
- Dreikausen, L.; Blender, B.; Trifunovic-Koenig, M.; Salm, F.; Bushuven, S.; Gerber, B.; Henke, M. Analysis of microbial contamination during use and reprocessing of surgical instruments and sterile packaging systems. PLoS ONE 2023, 18, e0280595. [Google Scholar] [CrossRef]
- Cuscó, A.; Sánchez, A.; Altet, L.; Ferrer, L.; Francino, O. Individual Signatures Define Canine Skin Microbiota Composition and Variability. Front. Vet. Sci. 2017, 4, 6. [Google Scholar] [CrossRef]
- Older, C.E.; Diesel, A.B.; Lawhon, S.D.; Queiroz, C.R.R.; Henker, L.C.; Rodrigues Hoffmann, A. The feline cutaneous and oral microbiota are influenced by breed and environment. PLoS ONE 2019, 14, e0220463. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Wade, W. Unculturable bacteria--the uncharacterized organisms that cause oral infections. J. R. Soc. Med. 2002, 95, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Naidu, P.; Shokoples, S.; Martin, I.; Zelyas, N.; Singh, A. Evaluation of 5 commercial assays for the detection of Mycoplasma genitalium and other Urogenital Mycoplasmas. Med. Microbiol. Immunol. 2021, 210, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Mijten, P.; van den Bogaard, A.E.; Hazen, M.J.; de Kruif, A. Bacterial contamination of fetal fluids at the time of cesarean section in the cow. Theriogenology 1997, 48, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Lagier, J.C.; Raoult, D.; Khelaifia, S. Bacterial culture through selective and non-selective conditions: The evolution of culture media in clinical microbiology. New Microbes New Infect. 2019, 34, 100622. [Google Scholar] [CrossRef]


{kind=link}
| Age Years Mean ± SD | Weight kg Median (Range) | Litter Size Mean ± SD | Breed | |
|---|---|---|---|---|
| Bitches | 3.8 ± 2.2 | 38 (3.2–63) | 5 ± 1.4 | Boston Terrier, Chihuahua, Dogue de Bordeaux, English Staffordshire Bull Terrier, French Bulldog |
| Queens | 4.5 ± 2 | 3.9 (3.3–4.3) | 2.5 ± 0.5 | Russian blue (n = 2), Scottish fold |
| Uterus a | Amniotic Fluid b | Meconium c | |||||||
|---|---|---|---|---|---|---|---|---|---|
| FMU | Result | Bacteria | Growth * | Result | Bacteria | Growth * | Result | Bacteria | Growth * |
| C-FMU-1 | neg | neg | neg | ||||||
| C-FMU-2 | + | Bacillus spp. X | High | + | Bacillus spp. | High | + | Coagulase-negative Staphylococcus (S. hominis) X Acinetobacter baumannii X | High High |
| C-FMU-3 | neg | neg | + | Coagulase-negative Staphylococci (S. epidermidis) | Low | ||||
| C-FMU-4 | neg | neg | neg | ||||||
| C-FMU-5 | + | Pseudomonas spp. (p. fluorescens) | Low | neg | + | Acinetobacter lwoffiiX | High | ||
| Uterus a | Amniotic Fluid b | Meconium c | |||||||
|---|---|---|---|---|---|---|---|---|---|
| FMU | Results | Bacteria | Growth * | Result | Bacteria | Growth * | Result | Bacteria | Growth * |
| F-FMU-1 | neg | neg | neg | ||||||
| F-FMU-2 | + | Coagulase-negative Staphylococcus (S. epidermidis) | ** Very low * | + | Pseudomonas aeruginosaX | ** Very low | neg | ||
| F-FMU-3 | + | Pseudomonas aeruginosaX | High | neg | + | Psychrobacter sanguinis | ** Very low | ||
| Grouping Method | K 1 | Order p-Value | Family p-Value | Genus p-Value | Species p-Value |
|---|---|---|---|---|---|
| Animal species | (K = 2) | 0.017 a ** 0.008 b *** | 0.033 a ** 0.020 b ** | 0.062 a 0.057 b | 0.033 a ** 0.020 b ** |
| Feto-maternal unit | Cats (K = 3) | 0.45 a 0.68 b | 0.73 a 1 b | 0.38 a 0.63 b | 0.34 a 0.59 b |
| Dogs (K = 5) | 0.59 a 0.68 b | 0.52 a 0.60 b | 0.58 a 0.71 b | 0.61 a 0.82 b | |
| Feto-maternal tissues vs. controls | Cats (K = 2) | 0.82 a 0.54 b | 1 a 1 b | 1 a 1 b | 1 a 1 b |
| Dogs (K = 2) | 0.51 a 0.53 b | 0.51 a 0.53 b | 0.51 a 0.53 b | 0.43 a 0.31 b | |
| Tissues (uterus, amniotic fluid, meconium, controls) | Cats (K = 4) | 0.46 a 0.77 b | 0.47 a 0.78 b | 0.34 a 0.58 b | 0.22 a 0.18 b |
| Dogs (K = 4) | 0.11 a 0.25 b | 0.035 a ** 0.09 b | 0.07 a 0.08 b | 0.35 a 0.34 b | |
| Surgical facility | (K = 2) | 0.90 a 0.70 b | 0.74 a 0.67 b | 1 a 1 b | 0.73 a 0.67 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banchi, P.; Colitti, B.; Del Carro, A.; Corrò, M.; Bertero, A.; Ala, U.; Del Carro, A.; Van Soom, A.; Bertolotti, L.; Rota, A. Challenging the Hypothesis of in Utero Microbiota Acquisition in Healthy Canine and Feline Pregnancies at Term: Preliminary Data. Vet. Sci. 2023, 10, 331. https://doi.org/10.3390/vetsci10050331
Banchi P, Colitti B, Del Carro A, Corrò M, Bertero A, Ala U, Del Carro A, Van Soom A, Bertolotti L, Rota A. Challenging the Hypothesis of in Utero Microbiota Acquisition in Healthy Canine and Feline Pregnancies at Term: Preliminary Data. Veterinary Sciences. 2023; 10(5):331. https://doi.org/10.3390/vetsci10050331
Chicago/Turabian StyleBanchi, Penelope, Barbara Colitti, Andrea Del Carro, Michela Corrò, Alessia Bertero, Ugo Ala, Angela Del Carro, Ann Van Soom, Luigi Bertolotti, and Ada Rota. 2023. "Challenging the Hypothesis of in Utero Microbiota Acquisition in Healthy Canine and Feline Pregnancies at Term: Preliminary Data" Veterinary Sciences 10, no. 5: 331. https://doi.org/10.3390/vetsci10050331