Investigation of PAG2 mRNA Expression in Water Buffalo Peripheral Blood Mononuclear Cells and Polymorphonuclear Leukocytes from Maternal Blood at the Peri-Implantation Period

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Pregnancy Diagnosis
2.3. PAG-2 Radioimmunoassay
2.4. Isolation of PBMC and PMN
2.5. RNA Isolation Reverse Transcription and qPCR
2.6. Statistical Analysis
3. Results
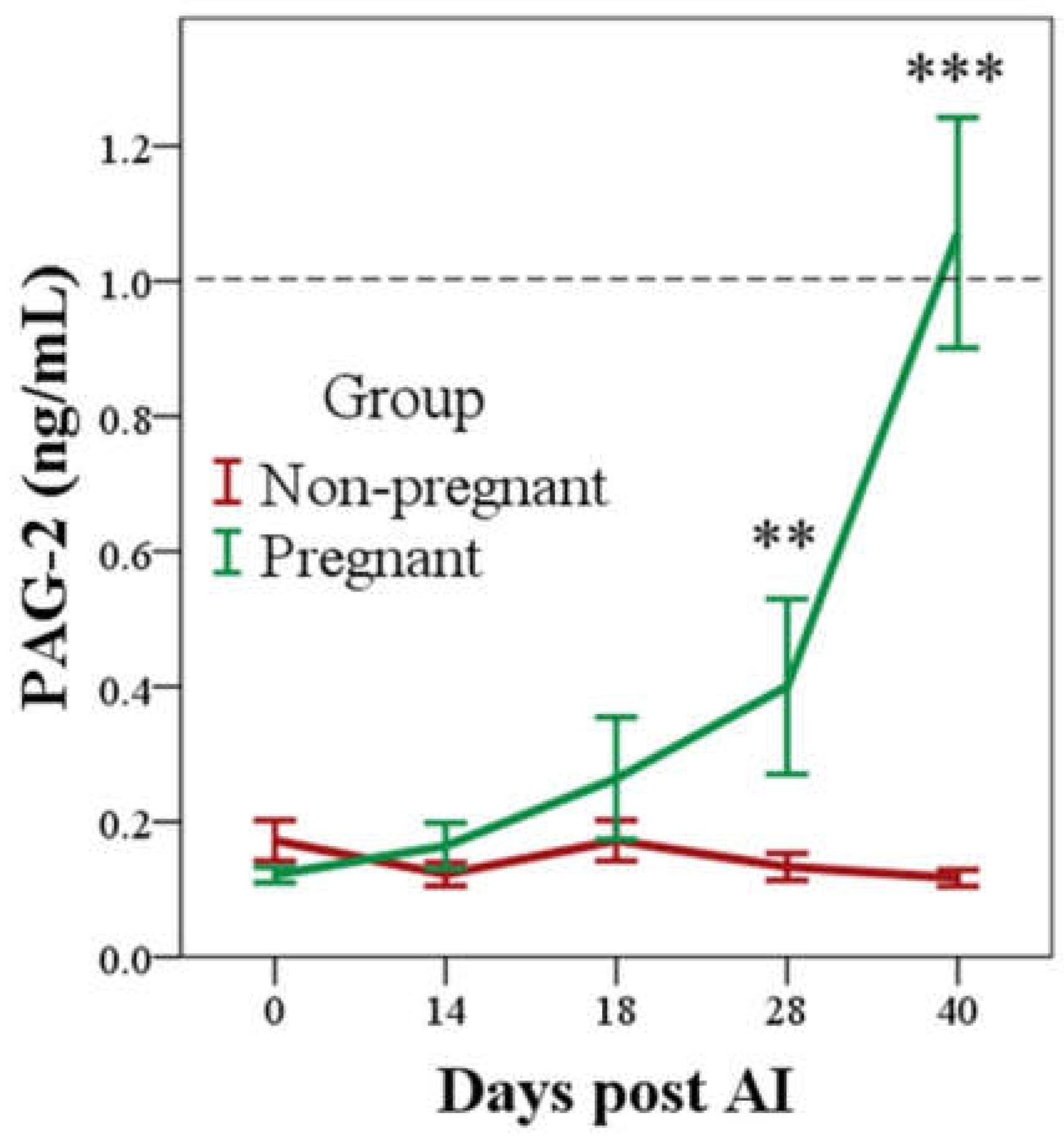
3.1. PAG-2 Concentrations Determined by RIA System in Pregnant and Non-Pregnant Buffaloes
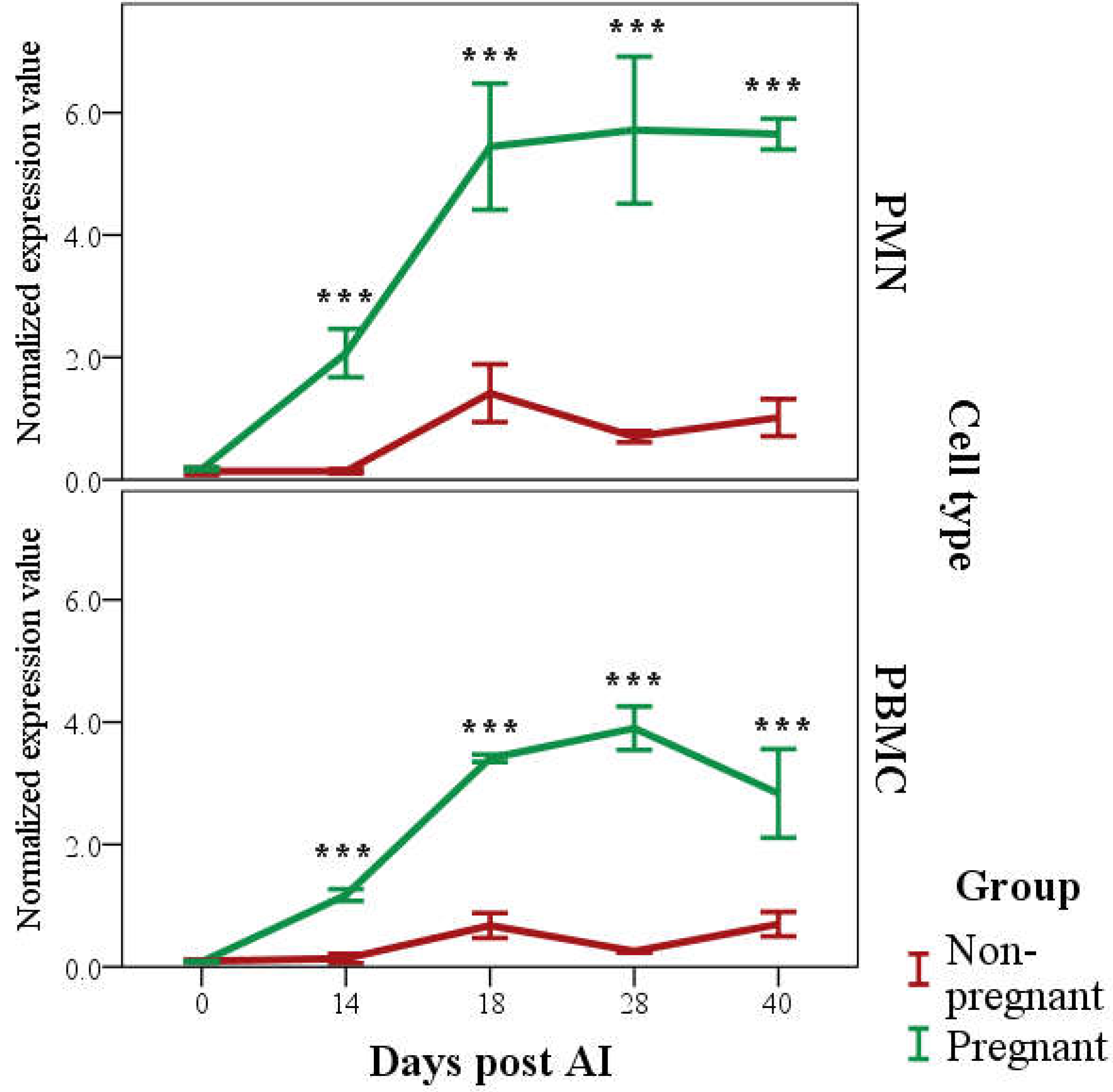
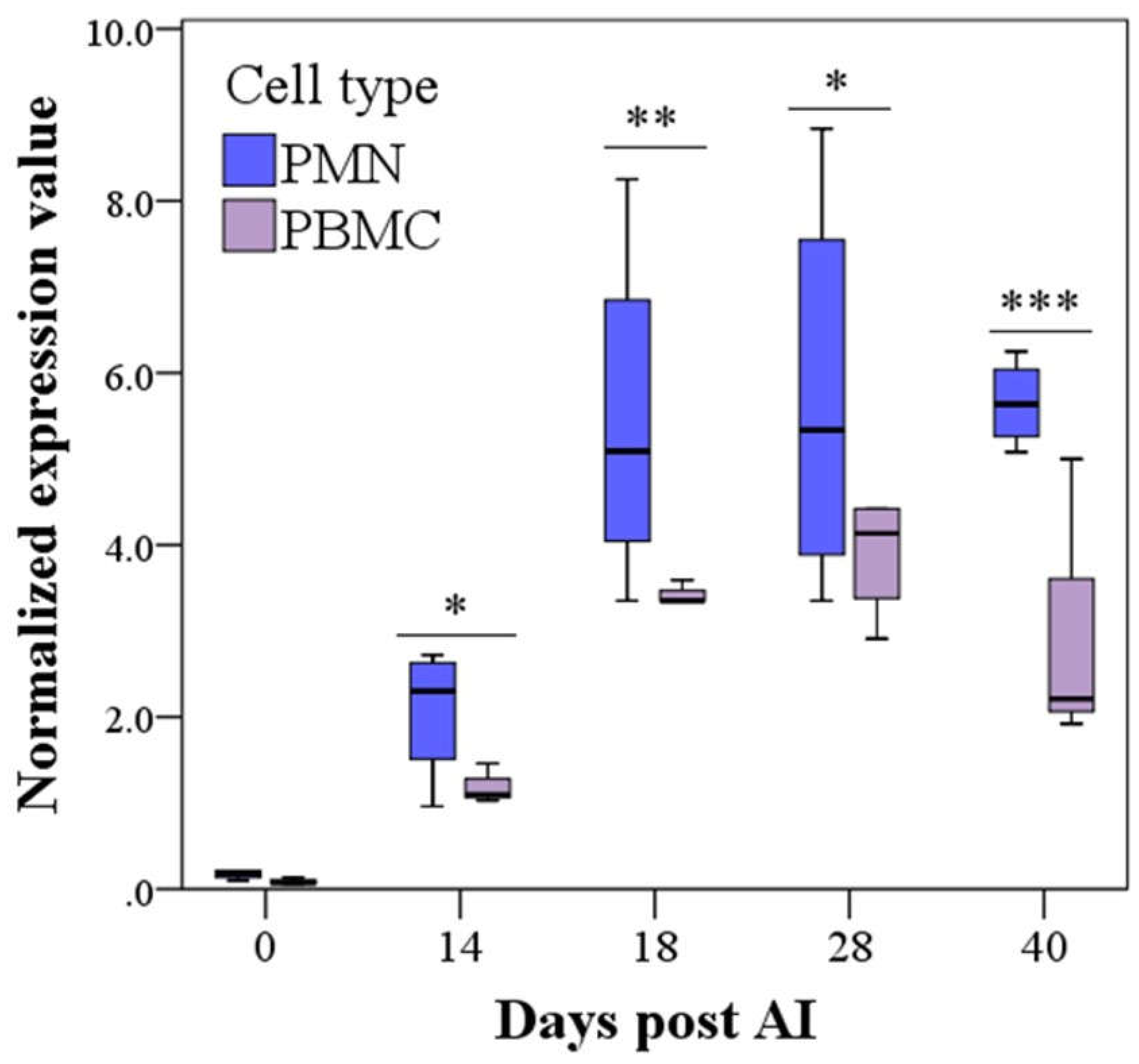
3.2. PAG2 Expression in PMN and PBMC in Pregnant and Non-Pregnant Buffaloes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zoli, A.P.; Beckers, J.F.; Wouters-Ballman, P.; Closset, J.; Falmagne, P.; Ectors, F. Purification and characterization of a bovine pregnancy-associated glycoprotein. Boil. Reprod. 1991, 45, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Beckers, J.F.; Roberts, R.M.; Zoli, A.P.; Ectors, F.; Derivaux, J. Molécules de la familles des proteases aspartique dans la placenta des Runinats: Hormones ou proteins? Bull. Acad. Méd. Belg. 1994, 143, 355–367. [Google Scholar]
- Xie, S.; Low, B.G.; Nagel, R.J.; Beckers, J.F.; Roberts, R.M. A novel glycoprotein of the aspartic proteinase gene family expressed in bovine placental trophectoderm. Boil. Reprod. 1994, 51, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Szafranska, B.; Xie, S.; Green, J.; Roberts, R.M. Porcine Pregnancy-Associated Glycoproteins: New members of the aspartic Proteinase gene family expressed in the trophectoderm. Boil. Reprod. 1995, 53, 21–28. [Google Scholar] [CrossRef]
- Xie, S.; Green, J.; Bao, B.; Beckers, J.F.; Valdez, K.E.; Hakami, L.; Roberts, R.M. Multiple pregnancy-associated glycoproteins are secreted by day 100 ovine placental tissue. Boil. Reprod. 1997, 57, 1384–1393. [Google Scholar] [CrossRef] [Green Version]
- Garbayo, J.M.; Remy, B.; Alabart, J.L.; Folch, J.; Wattiez, R.; Falmagne, P.; Beckers, J.F. Isolation and partial characterization of a pregnancy-associated glycoprotein family from the goat placenta. Boil. Reprod. 1998, 58, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Garbayo, J.M.; Green, J.; Manikkam, M.; Beckers, J.F.; Kiesling, D.; Ealy, A.D.; Roberts, R.M. Caprine Pregnancy-associated glycoproteins (PAG): Their cloning, expression and evolutionary relationship to other PAG. Mol. Reprod. Dev. 2000, 57, 311–322. [Google Scholar] [CrossRef]
- Green, J.A.; Xie, S.; Quan, X.; Bao, B.; Gan, X.; Mathialagan, N.; Beckers, J.F.; Roberts, R.M. Pregnancy-associated bovine and ovine glycoproteins exhibit spatially and temporally distinct expression patterns during pregnancy. Boil. Reprod. 2000, 62, 1624–1631. [Google Scholar] [CrossRef]
- El Amiri, B.; Remy, B.; De Sousa, N.M.; Beckers, J.F. Isolation and characterization of eight pregnancy-associated glycoproteins present at high levels in the ovine placenta between day 60 and day 100 of gestation. Reprod. Nutr. Dev. 2004, 44, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, G.A.; Parks, T.E.; Killian, G.; Ealy, A.D.; Green, J.A. A cloning and expression analysis of pregnancy-associated glycoproteins expressed in trophoblast of the white-tail deer placenta. Mol. Reprod. Dev. 2007, 74, 1355–1362. [Google Scholar] [CrossRef]
- Barbato, O.; Sousa, N.; Klisch, K.; Clerget, E.; Debenedetti, A.; Barile, V.; Malfatti, A.; Beckers, J. Isolation of new pregnancy-associated glycoproteins from water buffalo (bubalus bubalis) placenta by Vicia villosa affinity cromatography. Res. Vet. Sci. 2008, 85, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Kiewisz, K.; Sousa, N.M.; Beckers, J.F.; Panasiewics, G.; Gizejewski, Z.; Szafranska, B. Identification of multiple pregnancy-associated glycoproteins (PAGs) purified from European bison (Eb; Bison bison) placenta. Anim. Reprod. Sci. 2009, 112, 229–250. [Google Scholar] [CrossRef] [PubMed]
- Telugu, B.P.; Walker, A.M.; Green, J.A. Characterization of the bovine pregnancy-associated glycoprotein gene family—Analysis of gene sequences, regulatory regions within the promoter and expression of selected genes. BMC Genom. 2009, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.H.; Barbato, O.; Bui, X.N.; Beckers, J.F.; Melo de Sousa, N. Assessment of pregnancy-associated glycoprotein (PAG) concentrations in swamp buffalo samples from fetal and maternal origins using interspecies antisera. Anim. Sci. J. 2012, 83, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Barbato, O.; Melo de Sousa, N.; Barile, V.L.; Canali, C.; Beckers, J.F. Purification of pregnancy-associated glycoproteins from late-pregnancy Bubalus bubalis placentas and development of a radioimmunoassay for pregnancy diagnosis in water buffalo females. BMC Vet. Res. 2013, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Beriot, M.; Tchimbou, A.F.; Barbato, O.; Beckers, J.F.; Sousa, N.M. Identification of pregnancy-associated glycoproteins and alpha-fetoprotein in in fallow deer (Dama dama) placenta. Acta Vet. Scand. 2014, 56, 4. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Green, J.A.; Piontkivska, H.; Robetrs, R.M. Adaptive diversification within a large family of recently duplicated, placentally expressed genes. Proc. Natl. Acad. Sci. USA 2000, 97, 3119–3323. [Google Scholar] [CrossRef]
- Hughes, A.L.; Green, J.A.; Piontkivska, H.; Robetrs, R.M. Aspartic Proteinase Phylogenetic and the Origin of Pregnancy-Associated Glycoproteins. Mol. Biol. Evol. 2013, 20, 1940–1945. [Google Scholar] [CrossRef]
- Wooding, F.B.; Roberts, R.M.; Green, J.A. Light and electron microscope immunocytochemical studies of the distribution of pregnancy-associated glycoproteins (PAGs) throughout pregnancy in the cow: Possible functional implications. Placenta 2005, 26, 807–827. [Google Scholar] [CrossRef]
- Garbayo, J.M.; Serrano, B.; Lopez-Gatius, F. Identification of novel pregnancy-associated glycoproteins (PAG) expressed by the perimplantion conceptus of domestic ruminants. Anim. Reprod. Sci. 2008, 103, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.M.; Pohler, K.J.; Smith, M.F.; Green, J.A. Placental PAGs: Gene origins, expression patterns, and use as markers of pregnancy. Reproduction 2015, 149, R115–R126. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.M.; Remy, B.; El Amiri, B.; De Figueiredo, J.R.; Banga-Mboko, H.; Goncalves, P.B.D.; Beckers, J.F. Characterization of pregnancy-associated glycoproteins extracted from zebu (Bos indicus) placentas removed at different gestational period. Reprod. Nutr. Dev. 2002, 42, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Touzard, E.; Reinaud, P.; Dubois, O.; Guyader-Joly, C.; Humblot, P. Specific expression patterns and cell distribution of ancient and modern PAG in bovine placenta during pregnancy. Reproduction 2013, 146, 347–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoli, A.P.; Guibault, L.A.; Delahaut, P.; Benitez Ortiz, W.; Beckers, J.F. Radioimmunoassay of a bovine pregnancy-associated glycoprotein in serum: Its application for pregnancy diagnosis. Boil. Reprod. 1992, 46, 83–92. [Google Scholar] [CrossRef]
- Szenci, O.; Beckers, J.F.; Sulon, J.; Sasser, G.; Taverne, M.A.M.; Varga, J.; Borzsony, L.; Hanzen, C.; Schekk, G. Comparison of ultrasonography, bovine pregnancy specific B and bovine pregnancy-associated glycoprotein 1 test for pregnancy detection in dairy cows. Theriogenology 1998, 50, 77–88. [Google Scholar] [CrossRef]
- Barbato, O.; Sousa, N.M.; Debenedetti, A.; Canali, C.; Todini, L.; Beckers, J.F. Validation of a new pregnancy-associated glycoprotein radioimmunoassay method for the detection of early pregnancy in ewes. Theriogenology 2009, 72, 993–1000. [Google Scholar] [CrossRef]
- El Amiri, B.; Sousa, N.M.; Alvarez Oxiley, A.; Hadarbach, D.; Beckers, J.F. Pregnancy-associated glycoprotein (PAG) concentration in plasma and milk samples for early pregnancy diagnosis in Lacaune dairy sheep. Res. Vet. Sci. 2015, 99, 30–36. [Google Scholar] [CrossRef]
- Barbato, O.; Chiardia, E.; Barile, V.L.; Pierri, F.; Sousa, M.N.; Terracina, L.; Canali, C.; Avellini, L. Investigation into omocysteine, vitamin E and malondialdeyde as indicators of successful artificial insemination in synchronized buffalo cows (Bubalus bubalis). Res. Vet. Sci. 2016, 104, 100–105. [Google Scholar] [CrossRef]
- Barbato, O.; Menchetti, L.; Sousa, N.M.; Malfatti, A.; Brecchia, G.; Canali, C.; Beckers, J.F.; Barile, V.L. Pregnancy-associated glycoproteins (PAGs) concentrations in water buffaloes (Bubalus bubalis) during gestation and the postpartum period. Theriogenology 2017, 97, 73–77. [Google Scholar] [CrossRef]
- Barbato, O.; Menchetti, L.; Sousa, N.M.; Malfatti, A.; Brecchia, G.; Canali, C.; Beckers, J.F.; Barile, V.L. Correlation of two radioimmunoassay systems for measuring plasma pregnancy-associated glycoproteins concentrations during early pregnancy and postpartum periods in water buffalo. Reprod. Domest. Anim. 2018, 53, 1483–1490. [Google Scholar] [CrossRef]
- Garcia-Ispierto, B.; Almeria, S.; Serrano-Perez, B.; Sousa, N.M.; Beckers, J.F.; López-Gatius, F. Plasma Concentrations of Pregnancy-Associated Glycoproteins measured using anti-bovine PAG-2 antibodies on day 120 of gestation predict abortion in dairy cows naturally infected with Neospora caninum. Reprod. Domest. Anim. 2013, 48, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ispierto, B.; Serrano-Pérez, S.; Martínez-Bello, D.; Tchimbou, A.F.; Sousa, N.M.; Beckers, J.F.; López-Gatius, F. Effects of crossbreeding on endocrine patterns determined in beef/dairy cows naturally infected with Neospora caninum. Theriogenology 2015, 83, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ispierto, B.; Ressello-Visa, M.A.; Serrano-Pérez, B.; Mur-Nuvales, R.; Sousa, N.M.; Beckers, J.F.; López-Gatius, F. Plasma concentration of pregnancy-associated glycoproteins I and II and progesterone on day 28 post AI as marker of twin pregnancy in dairy cattle. Livest. Sci. 2016, 192, 44–47. [Google Scholar] [CrossRef]
- Mur-Novales, R.; López-Gatius, F.; Serrano-Pérez, B.; Garcia-Ispierto, B.; Darwich, L.; Cabezon, O.; Sousa, N.M.; Beckers, J.F.; Almeria, S. Experimental Neospora Caninum infection in pregnant dairy heifers raises voncentrations of pregnancy-associated glycoproteins 1 and 2 in in foetal fluids. Reprod. Domest. Anim. 2016, 51, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Almeria, S.; Serrano-Pérez, B.; López-Gatius, F. Immune response in bovine neosporis: Protection or contribution to the pathogenesis of abortion. Microb. Pathog. 2017, 109, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Barbato, O.; Guelfi, G.; Barile, V.L.; Menchetti, L.; Tortiello, C.; Canali, C.; Brecchia, G.; Traina, G.; Beckers, J.F.; Sousa, N.M. Using real-time PCR to identify pregnancy-associated glycoprotein 2 (PAG-2) in water buffalo (Bubalus bubalis) blood in early pregnany. Theriogenology 2017, 97, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Singh, J.K.; Khanna, S.; Phulia, S.K.; Sarkar, S.K.; Singh, I. Fetal age determination in Murrah buffaloes from days 22 through 60 with ultrasonography. Indian J. Anim. Sci. 2012, 82, 374–376. [Google Scholar]
- Serrano-Pérez, B.; Hansen, P.J.; Mur-Novales, R.; Garcia-Ispierto, B.; Sousa, N.M.; Beckers, J.F.; Almeria, S.; López-Gatius, F. Crosstalk between uterine serpin (SERPINA14) and pregnancy-associated glycoproteins at the fetal-maternal interface in pregnant dairy heifer experimentally infected with Neospora caninum. Theriogenology 2016, 86, 824–830. [Google Scholar] [CrossRef]
- Beckers, J.F.; Dewulf, M.; Verstegen, J.; Wouters-Ballman, P.; Ectors, F. Isolation of a bovine chorionic gonadotrophin (bCG). Theriogenology 1988, 29, 218. [Google Scholar] [CrossRef]
- Vaitukaitis, J.; Robbins, J.B.; Nieschlag, E.; Ross, G.T. A method for producing specific antisera with small doses of immonogen. J. Clin. Endocrinol. Metab. 1971, 33, 988–991. [Google Scholar] [CrossRef]
- Thorell, J.I.; Johansson, B.G. Enzymatic iodination of polypeptides with 125I to high specific activity. Biochim. Biophys. Acta 1971, 251, 363–369. [Google Scholar] [CrossRef]
- Diverio, S.; Guelfi, G.; Barbato, O.; Di Mari, W.; Egidi, G.; Santoro, M.M. Non-invasive assessment of animal exercise stress: Real-time PCR of GLUT4, COX2, SOD1 and HSP70 in avalanche military dog saliva. Animal 2015, 5, 774–792. [Google Scholar] [CrossRef] [PubMed]
- Greiner, M.; Pfeiffer, D.; Smith, R.D. Principles and practical application of the receiver-operating characteristic analysis for diagnostic tests. Prev. Vet. Med. 2000, 45, 23–41. [Google Scholar] [CrossRef]
- Yankey, S.J.; Hicks, B.A.; Carnahan, K.G.; Assiri, A.M.; Sinor, S.J.; Kodali, K.; Stellflug, J.N.; Ott, T.L. Expression of the antiviral protein Mx in peripheral blood mononuclear cells of pregnant and bred, non-pregnant ewes. J. Endocrinol. 2001, 170, R7–R11. [Google Scholar] [CrossRef]
- Gifford, C.A.; Racicot, K.; Clark, D.S.; Austin, K.J.; Hansen, T.R.; Lucy, M.C.; Davies, C.J.; Ott, T.L. Regulation of interferon-stimulated genes in peripheral blood leukocytes in pregnant and bred, non pregnant dairy cows. J. Dairy Sci. 2007, 90, 274–280. [Google Scholar] [CrossRef]
- Pugliesi, G.; Miagawa, B.T.; Paiva, Y.N.; Franca, M.R.; Silva, L.A.; Binelli, M. Conceptus-induced changes in the Gene Expression of Blood Immune Cells and the ultrasound-accessed luteal function in beef-cattle: How early can we detect pregnancy? Boil. Reprod. 2014, 91, 95. [Google Scholar] [CrossRef] [PubMed]
- Chethan, S.G.; Singh, S.K.; Nongsiej, J.; Rakesh, H.B.; Singh, R.P.; Kumar, N.; Agarwal, S.K. IFN-τ acts in a dose-dependent manner on prostaglandin production by buffalo endometrial stromal cells cultured in vitro. Reprod. Domest. Anim. 2014, 49, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Burghardt, R.C.; Johnson, G.A.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Anim. Reprod. Sci. 2004, 82–83, 537–550. [Google Scholar] [CrossRef]
- Parent, J.; Chapdelaine, P.; Sirois, J.; Fortier, M.A. Expression of microsomal prostaglandin E synthase in bovine endometrium: Coexpression with cyclooxygenase type 2 and regulation by interferon-τ. Endocrinology 2002, 143, 2936–2943. [Google Scholar] [CrossRef]
- Spencer, T.E.; Bazer, F.W. Biology of progesterone action during pregnancy recognition and maintenance of pregnancy. Front. Biosci. 2002, 7, 1879–1898. [Google Scholar] [CrossRef]
- Mamo, S.; Mehta, J.P.; McGettigan, P.; Fair, T.; Spencer, T.E.; Bazer, F.W.; Lonergan, P. RNA sequencing reveals novel gene clusters in bovine conceptuses associated with maternal recognition of pregnancy and implantation. Biol. Reprod. 2011, 85, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.M.; Cerri, R.L.; Kim, I.H.; Ealy, A.D.; Hansen, P.J.; Staples, C.R.; Thatcher, W.W. Effects of lactation and pregnancy on metabolic and hormonal responses and expression of selected conceptus and endometrial genes of Holstein dairy cattle. J. Dairy Sci. 2012, 95, 5645–5656. [Google Scholar] [CrossRef]
- Hue, I.; Degrelle, S.A.; Turenne, N. Conceptus elongation in cattle: Genes, models and questions. Anim. Reprod. Sci. 2012, 134, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, R.P.; Sutherland, W.D.; Sasse, R.G. Effect of pregnancy-specific protein B on luteal cell progesterone, prostaglandin, and oxytocin production during two stages of the bovine estrous cycle. J. Anim. Sci. 1995, 73, 2662–2668. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, R.P.; Sutherland, W.D.; Sasser, R.G. Bovine luteal cell production in vitro of prostaglandin E2, oxytocin and progesterone in response to pregnancy-specific protein B and prostaglandin F2α. J. Reprod. Fertil. 1996, 107, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Weems, S.Y.; Lammoglia, M.A.; Vera-Avila, H.R.; Randel, R.D.; King, C.; Sasser, R.G.; Weems, C.W. Effect of luteinizing hormone (LH), PGE2, 8-EPI-PGE1, 8-EPI-PGE2, trichosanthin, and pregnancy specific protein B (PSPB) on secretion of progesterone in vitro by corpora lutea (CL) from non pregnant and pregnant cows. Prostaglandins 1998, 55, 27–42. [Google Scholar] [CrossRef]
- Austin, K.J.; King, L.P.; Vierk, J.E.; Spencer, R.C.; Hauren, T.R. Pregnancy-specific-protein B induces release on an alpha chemokine in bovine endometrium. Endocrinology 1999, 140, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Hoeben, D.; Burvenicha, C.; Massart-Leena, A.; Lenjou, M.; Nij, G. In vitro effect of ketone bodies, glucocorticosteroids and bovine pregnancy-associated glycoprotein on cultures of bone marrow progenitor cells of cows and calves. Vet. Immunol. Immunopathol. 1999, 68, 229–240. [Google Scholar] [CrossRef]
- Sandra, O.; Constanta, C.; Carvalho, A.V.; Eozénoua, C.; Valoura, D.; Mauffréa, V.; Huea, I.; Charpignya, G. Maternal organism and embryo biosensoring: Insights from ruminants. J. Reprod. Immunol. 2015, 108, 105–113. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Exons Connected | Amplicon |
|---|---|---|---|
| Pregnancy-Associated Glycoprotein 2 (PAG2) | BOS | 9-9 | 127 bp |
| TaqMan Bt03292796_gH | NM_176614.1 | ||
| Actin beta(ACTB) | BOS | 4-5 | 144 bp |
| TaqMan Bt03279175_g1 | NM_173979.3 |
| Day | Diagnosis | Outcome 1 | Sensitivity | Specificity | Accuracy | |
|---|---|---|---|---|---|---|
| Non-Pregnant (n = 18) | Pregnant (n = 14) | |||||
| 14 | Non-Pregnant | 18 (100.0%) | 14 (100.0%) | 0.00% | 100.00% | 56.30% |
| Pregnant | 0 (0.0%) | 0 (0.0%) | ||||
| 18 | Non-Pregnant | 18 (100.0%) | 13 (92.9%) | 7.10% | 100.00% | 59.40% |
| Pregnant | 0 (0.0%) | 1 (7.1%) | ||||
| 28 | Non-Pregnant | 18 (100.0%) | 11 (78.6%) | 21.40% | 100.00% | 65.60% |
| Pregnant | 0 (0.0%) | 3 (21.4%) | ||||
| 40 | Non-Pregnant | 18 (100.0%) | 3 (21.4%) | 78.60% | 100.00% | 90.60% |
| Pregnant | 0 (0.0%) | 11 (78.6%) | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbato, O.; Guelfi, G.; Menchetti, L.; Brecchia, G.; Melo de Sousa, N.; Canali, C.; Grandoni, F.; Scatà, M.C.; De Matteis, G.; Casano, A.B.; et al. Investigation of PAG2 mRNA Expression in Water Buffalo Peripheral Blood Mononuclear Cells and Polymorphonuclear Leukocytes from Maternal Blood at the Peri-Implantation Period. Vet. Sci. 2019, 6, 8. https://doi.org/10.3390/vetsci6010008
Barbato O, Guelfi G, Menchetti L, Brecchia G, Melo de Sousa N, Canali C, Grandoni F, Scatà MC, De Matteis G, Casano AB, et al. Investigation of PAG2 mRNA Expression in Water Buffalo Peripheral Blood Mononuclear Cells and Polymorphonuclear Leukocytes from Maternal Blood at the Peri-Implantation Period. Veterinary Sciences. 2019; 6(1):8. https://doi.org/10.3390/vetsci6010008
Chicago/Turabian StyleBarbato, Olimpia, Gabriella Guelfi, Laura Menchetti, Gabriele Brecchia, Noelita Melo de Sousa, Claudio Canali, Francesco Grandoni, Maria Carmela Scatà, Giovanna De Matteis, Anna Beatrice Casano, and et al. 2019. "Investigation of PAG2 mRNA Expression in Water Buffalo Peripheral Blood Mononuclear Cells and Polymorphonuclear Leukocytes from Maternal Blood at the Peri-Implantation Period" Veterinary Sciences 6, no. 1: 8. https://doi.org/10.3390/vetsci6010008