
Detection of Borrelia burgdorferi sensu-lato-Specific Antibodies in Sera of Canine and Equine Origin—A Comparative Study with Two Line Immunoassays

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Serum Samples
2.2. Antibody Detection in the Serum Samples
2.2.1. LIA A
2.2.2. LIA B
2.3. Statistical Methods
3. Results
3.1. Canine Serum Samples
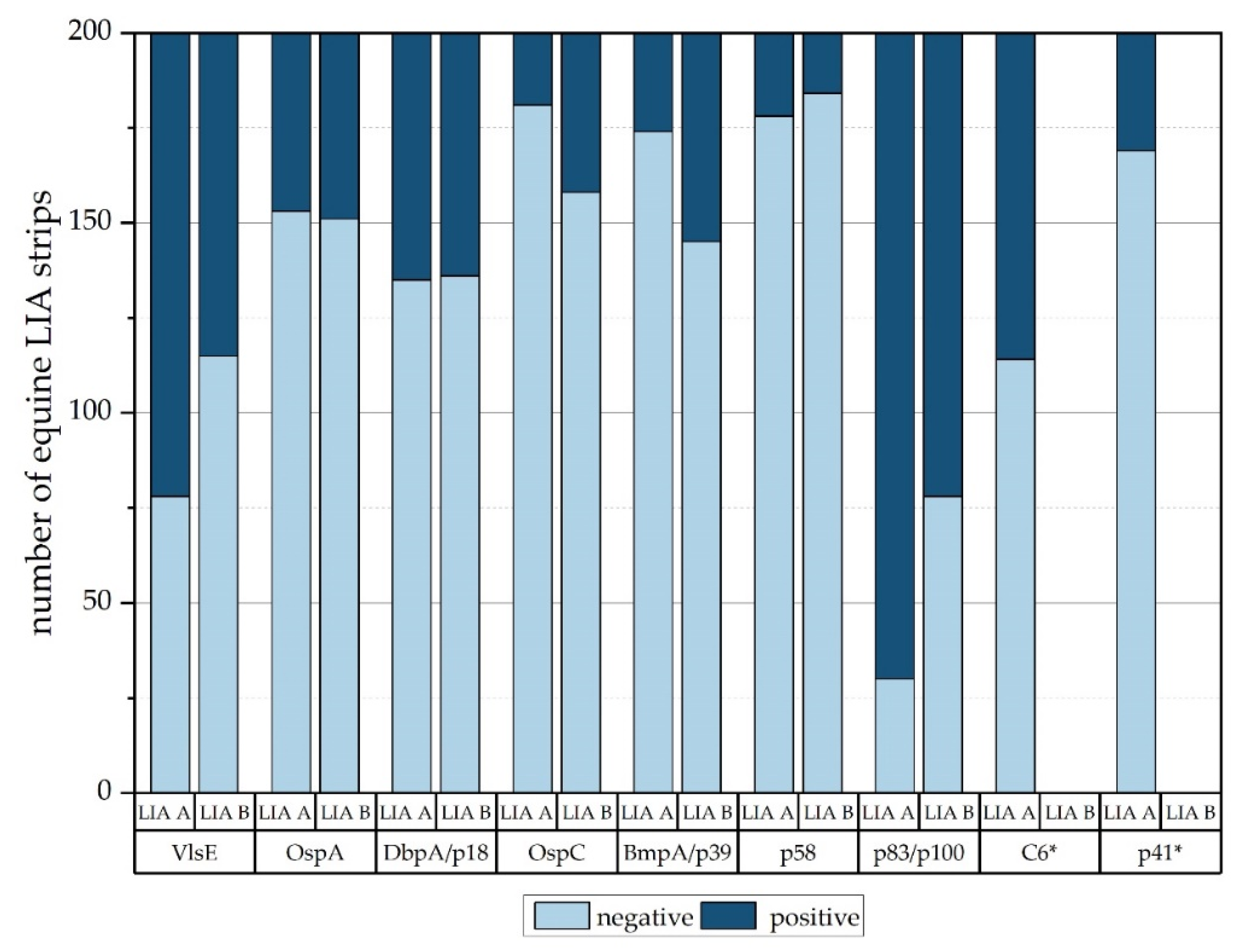
3.2. Equine Serum Samples
4. Discussion
4.1. Sensitivity, Specificity, and Overall Results
4.2. Handling of LIAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Angelou, A.; Gelasakis, A.I.; Verde, N.; Pantchev, N.; Schaper, R.; Chandrashekar, R.; Papadopoulos, E. Prevalence and risk factors for selected canine vector-borne diseases in Greece. Parasites Vectors 2019, 12, 283. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Straubinger, R.K.; Sauter-Louis, C.; Hartmann, K. Prevalence of antibodies against Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum and their clinical relevance in dogs in Munich, Germany. Berl. Münchener Tierärztliche Wochenschr. 2012, 125, 337–344. [Google Scholar]
- Gerber, B.; Eichenberger, S.; Wittenbrink, M.M.; Reusch, C.E. Increased prevalence of Borrelia burgdorferi infections in Bernese Mountain Dogs: A possible breed predisposition. BMC Vet. Res. 2007, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Funk, R.A.; Pleasant, R.S.; Witonsky, S.G.; Reeder, D.S.; Werre, S.R.; Hodgson, D.R. Seroprevalence of Borrelia burgdorferi in horses presented for coggins testing in Southwest Virginia and change in positive test results approximately 1 year later. J. Vet. Intern. Med. 2016, 30, 1300–1304. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.; Erb, H.N. Dogs and horses with antibodies to outer-surface protein C as on-time sentinels for ticks infected with Borrelia burgdorferi in New York State in 2011. Prev. Vet. Med. 2012, 107, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G. Isolation and cultivation of Lyme disease spirochetes. Yale J. Biol. Med. 1984, 57, 521. [Google Scholar]
- Preac-Mursic, V.; Wilske, B.; Reinhardt, S. Culture of Borrelia burgdorferi on six solid media. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wormser, G.P.; Schwartz, I. Borrelia burgdorferi. In Molecular Medical Microbiology; Sussman, M., Ed.; Academic Press: London, UK, 2001; pp. 2059–2092. [Google Scholar]
- Straubinger, R.K. PCR-Based quantification of Borrelia burgdorferi organisms in canine tissues over a 500-Day postinfection period. J. Clin. Microbiol. 2000, 38, 2191–2199. [Google Scholar] [CrossRef]
- Nunes, M.; Parreira, R.; Carreira, T.; Inacio, J.; Vieira, M.L. Development and evaluation of a two-step multiplex TaqMan real-time PCR assay for detection/quantification of different genospecies of Borrelia burgdorferi sensu lato. Ticks Tick Borne Dis. 2018, 9, 176–182. [Google Scholar] [CrossRef]
- Okeyo, M.; Hartberger, C.; Margos, G.; Straubinger, R.K.; Sing, A.; Fingerle, V. Comparison of methods for economic and efficient tick and Borrelia DNA purification. Ticks Tick Borne Dis. 2019, 10, 1041–1045. [Google Scholar] [CrossRef]
- Steere, A.C.; Grodzicki, R.L.; Kornblatt, A.N.; Craft, J.E.; Barbour, A.G.; Burgdorfer, W.; Schmid, G.P.; Johnson, E.; Malawista, S.E. The spirochetal etiology of Lyme disease. N. Engl. J. Med. 1983, 308, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Dressler, F.; Whalen, J.A.; Reinhardt, B.N.; Steere, A.C. Western blotting in the serodiagnosis of Lyme disease. J. Infect. Dis. 1993, 167, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme disease–A tick-borne spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Bartenhagen, N.H.; Craft, J.E.; Hutchinson, G.J.; Newman, J.H.; Rahn, D.W.; Sigal, L.H.; Spieler, P.N.; Stenn, K.S.; Malawista, S.E. The early clinical manifestations of Lyme disease. Ann. Intern. Med. 1983, 99, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.W. Immunodiagnostic tests for Lyme disease. Yale J. Biol. Med. 1984, 57, 567–572. [Google Scholar] [PubMed]
- Gomes-Solecki, M.J.; Wormser, G.P.; Schriefer, M.; Neuman, G.; Hannafey, L.; Glass, J.D.; Dattwyler, R.J. Recombinant assay for serodiagnosis of Lyme disease regardless of OspA vaccination status. J. Clin. Microbiol. 2002, 40, 193–197. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Flavell, R.A.; Padula, S.J.; Anderson, J.F.; Fikrig, E. Serologic diagnosis of canine and equine borreliosis: Use of recombinant antigens in enzyme-linked immunosorbent assays. J. Clin. Microbiol. 1997, 35, 169–173. [Google Scholar] [CrossRef]
- Hulinska, D. Diagnosis of Lyme borreliosis with western blotting. Epidemiol. Mikrobiol. Imunol. 1997, 46, 3–8. [Google Scholar]
- Wilske, B.; Fingerle, V.; Herzer, P.; Hofmann, A.; Lehnert, G.; Peters, H.; Pfister, H.W.; Preac-Mursic, V.; Soutschek, E.; Weber, K. Recombinant immunoblot in the serodiagnosis of Lyme borreliosis. Comparison with indirect immunofluorescence and enzyme-linked immunosorbent assay. Med. Microbiol. Immunol. 1993, 182, 255–270. [Google Scholar] [CrossRef]
- Gauthier, D.T.; Mansfield, L.S. Western immunoblot analysis for distinguishing vaccination and infection status with Borrelia burgdorferi (Lyme disease) in dogs. J. Vet. Diagn. Investig. 1999, 11, 259–265. [Google Scholar] [CrossRef]
- Wilske, B.; Fingerle, V.; Preac-Mursic, V.; Jauris-Heipke, S.; Hofmann, A.; Loy, H.; Pfister, H.W.; Rossler, D.; Soutschek, E. Immunoblot using recombinant antigens derived from different genospecies of Borrelia burgdorferi sensu lato. Med. Microbiol. Immunol. 1994, 183, 43–59. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, S.M.; Shoop, E.; Johnson, R.C. Immunoblot interpretation criteria for serodiagnosis of early Lyme disease. J. Clin. Microbiol. 1995, 33, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Zoller, L.; Cremer, J.; Faulde, M. Western blot as a tool in the diagnosis of Lyme borreliosis. Electrophoresis 1993, 14, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Hauser, U.; Lehnert, G.; Wilske, B. Diagnostic value of proteins of three Borrelia species (Borrelia burgdorferi sensu lato) and implications for development and use of recombinant antigens for serodiagnosis of Lyme borreliosis in Europe. Clin. Diagn. Lab. Immunol. 1998, 5, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Hauser, U.; Wilske, B. Enzyme-linked immunosorbent assays with recombinant internal flagellin fragments derived from different species of Borrelia burgdorferi sensu lato for the serodiagnosis of Lyme neuroborreliosis. Med. Microbiol. Immunol. 1997, 186, 145–151. [Google Scholar] [CrossRef]
- Marangoni, A.; Moroni, A.; Accardo, S.; Cevenini, R. Borrelia burgdorferi VlsE antigen for the serological diagnosis of Lyme borreliosis. Eur. J. Clin. Microbiol. Infect. Dis. 2008, 27, 349–354. [Google Scholar] [CrossRef]
- Eicken, C.; Sharma, V.; Klabunde, T.; Lawrenz, M.B.; Hardham, J.M.; Norris, S.J.; Sacchettini, J.C. Crystal structure of Lyme disease variable surface antigen VlsE of Borrelia burgdorferi. Biol. Chem. 2002, 277, 21691–21696. [Google Scholar] [CrossRef]
- Zhang, J.R.; Norris, S.J. Genetic variation of the Borrelia burgdorferi gene VlsE involves cassette-specific, segmental gene conversion. Infect. Immun. 1998, 66, 3698–3704. [Google Scholar] [CrossRef]
- Shin, S.J.; Chang, Y.-F.; Jacobson, R.H.; Shaw, E.; Lauderdale, T.-L.; Appel, M.J.; Lein, D.H. Cross-reactivity between B. burgdorferi and other spirochetes affects specificity of serotests for detection of antibodies to the Lyme disease agent in dogs. Vet. Microbiol. 1993, 36, 161–174. [Google Scholar] [CrossRef]
- Jacobson, R.H.; Chang, Y.F.; Shin, S.J. Lyme disease: Laboratory diagnosis of infected and vaccinated symptomatic dogs. Semin. Vet. Med. Surg. (Small Anim.) 1996, 11, 172–182. [Google Scholar] [CrossRef]
- Chang, Y.F.; Appel, M.J.; Jacobson, R.H.; Shin, S.J.; Harpending, P.; Straubinger, R.; Patrican, L.A.; Mohammed, H.; Summers, B.A. Recombinant OspA protects dogs against infection and disease caused by Borrelia burgdorferi. Infect. Immun. 1995, 63, 3543–3549. [Google Scholar] [CrossRef] [PubMed]
- Barth, C.; Straubinger, R.K.; Krupka, I.; Muller, E.; Sauter-Louis, C.; Hartmann, K. Comparison of different diagnostic assays for the detection of Borrelia burgdorferi-specific antibodies in dogs. Vet. Clin. Pathol. 2014, 43, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Straubinger, R.K.; Chang, Y.F.; Jacobson, R.H.; Appel, M.J. Sera from OspA-vaccinated dogs, but not those from tick-infected dogs, inhibit in vitro growth of Borrelia burgdorferi. J. Clin. Microbiol. 1995, 33, 2745–2751. [Google Scholar] [CrossRef] [PubMed]
- Straubinger, R.K.; Dharma Rao, T.; Davidson, E.; Summers, B.A.; Jacobson, R.H.; Frey, A.B. Protection against tick-transmitted Lyme disease in dogs vaccinated with a multiantigenic vaccine. Vaccine 2001, 20, 181–193. [Google Scholar] [CrossRef]
- Summers, B.A.; Straubinger, A.F.; Jacobson, R.H.; Chang, Y.F.; Appel, M.J.; Straubinger, R.K. Histopathological studies of experimental Lyme disease in the dog. J. Comp. Pathol. 2005, 133, 1–13. [Google Scholar] [CrossRef]
- Knödlseder, J.M.; Fell, S.F.; Straubinger, R.K. A study with a commercial vaccine against Lyme borreliosis in horses using two different vaccination schedules: Characterization of the humoral immune response. Vaccine 2019, 37, 7207–7212. [Google Scholar] [CrossRef]
- Cohen, J. Weighted kappa: Nominal scale agreement with provision for scaled disagreement or partial credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef]
- Day, M.J. One health: The importance of companion animal vector-borne diseases. Parasites Vectors 2011, 4, 49. [Google Scholar] [CrossRef]
- Lindgren, E.; Gustafson, R. Tick-borne encephalitis in Sweden and climate change. Lancet 2001, 358, 16–18. [Google Scholar] [CrossRef]
- Breu, D.; Müller, E. Heterogeneity of canine immune responses to Borrelia burgdorferi in a line immunoassay comprising recombinant VlsE and C6 peptide. Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2017, 45, 295–300. [Google Scholar] [CrossRef]
- Divers, T.J.; Gardner, R.B.; Madigan, J.E.; Witonsky, S.G.; Bertone, J.J.; Swinebroad, E.L.; Schutzer, S.E.; Johnson, A.L. Borrelia burgdorferi Infection and Lyme Disease in North American Horses: A Consensus Statement. J. Vet. Intern. Med. 2018, 32, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Browning, A.; Carter, S.D.; Barnes, A.; May, C.; Bennett, D. Lameness associated with Borrelia burgdorferi infection in the horse. Vet. Rec. 1993, 132, 610–611. [Google Scholar] [CrossRef] [PubMed]
- James, F.M.; Engiles, J.B.; Beech, J. Meningitis, cranial neuritis, and radiculoneuritis associated with Borrelia burgdorferi infection in a horse. J. Am. Vet. Med. Assoc. 2010, 237, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Bhide, M.; Yilmaz, Z.; Golcu, E.; Torun, S.; Mikula, I. Seroprevalence of anti-Borrelia burgdorferi antibodies in dogs and horses in Turkey. Ann. Agric. Environ. Med. 2008, 15, 85–90. [Google Scholar] [PubMed]
- Durrani, A.Z.; Goyal, S.M. Seroprevalence of Borrelia burgdorferi in horses in Minnesota. J. Anim. Plant Sci. 2011, 21, 8–11. [Google Scholar]
- Käsbohrer, A.; Schönberg, A. Serologic studies of the occurrence of Borrelia burgdorferi in domestic animals in Berlin (West). Berl. Münchener Tierärztliche Wochenschr. 1990, 103, 374–378. [Google Scholar]
- Bae, J.M. Horses as a Potential Reservoir of Lyme Borreliosis in Jeju-do, Korea. J. Prev. Med. Public Health 2018, 51, 213–214. [Google Scholar] [CrossRef]
- Bruckbauer, H.R.; Preac-Mursic, V.; Fuchs, R.; Wilske, B. Cross-reactive proteins of Borrelia burgdorferi. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 224–232. [Google Scholar] [CrossRef]
- Wilske, B.; Preac-Mursic, V.; Fuchs, R.; Schierz, G. Diagnostik der Lyme-Borreliose. Diagnose & Labor 1990, 40, 24–36. [Google Scholar]
- Keir, A.A.; Stämpfli, H.R.; Crawford, J. Outbreak of acute colitis on a horse farm associated with tetracycline-contaminated sweet feed. Can. Vet. J. 1999, 40, 718–720. [Google Scholar]
- Barr, B.S.; Waldridge, B.M.; Morresey, P.R.; Reed, S.M.; Clark, C.; Belgrave, R.; Donecker, J.M.; Weigel, D.J. Antimicrobial-associated diarrhoea in three equine referral practices. Equine Vet. J. 2013, 45, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.P.; de Lacerda Neto, J.C. Jugular thrombophlebitis in horses: A review of fibrinolysis, thrombus formation, and clinical management. Can. Vet. J. 2013, 54, 65–71. [Google Scholar] [PubMed]
- Fawcett, P.T.; Rose, C.D.; Budd, S.M.; Gibney, K.M. Effect of immunization with recombinant OspA on serologic tests for Lyme borreliosis. Clin. Diagn. Lab. Immunol. 2001, 8, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.H.F.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia burgdorferi and ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Goldberg, M.S.; Popova, T.G.; Schoeler, G.B.; Wikel, S.K.; Hagman, K.E.; Norgard, M.V. Interdependence of environmental factors influencing reciprocal patterns of gene expression in virulent Borrelia burgdorferi. Mol. Microbiol. 2000, 37, 1470–1479. [Google Scholar] [CrossRef]
- Purser, J.E.; Norris, S.J. Correlation between plasmid content and infectivity in Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2000, 97, 13865–13870. [Google Scholar] [CrossRef]
- O’Connor, T.P.; Esty, K.J.; Hanscom, J.L.; Shields, P.; Philipp, M.T. Dogs vaccinated with common Lyme disease vaccines do not respond to IR6, the conserved immunodominant region of the VlsE surface protein of Borrelia burgdorferi. Clin. Diagn. Lab. Immunol. 2004, 11, 458–462. [Google Scholar]
- Liang, F.T.; Steere, A.C.; Marques, A.R.; Johnson, B.J.; Miller, J.N.; Philipp, M.T. Sensitive and specific serodiagnosis of Lyme disease by enzyme-linked immunosorbent assay with a peptide based on an immunodominant conserved region of Borrelia burgdorferi VlsE. J. Clin. Microbiol. 1999, 37, 3990–3996. [Google Scholar] [CrossRef]
- Topfer, K.; Straubinger, R.K. Characterization of the humoral immune response in dogs after vaccination against the Lyme borreliosis agent–A study with five commercial vaccines using two different vaccination schedules. Vaccine 2007, 25, 314–326. [Google Scholar] [CrossRef]
- Luft, B.J.; Mudri, S.; Jiang, W.; Dattwyler, R.J.; Gorevic, P.D.; Fischer, T.; Munoz, P.; Dunn, J.J.; Schubach, W.H. The 93-kilodalton protein of Borrelia burgdorferi: An immunodominant protoplasmic cylinder antigen. Infect. Immun. 1992, 60, 4309–4321. [Google Scholar] [CrossRef]
- Eiffert, H.; Schlott, T.; Hoppert, M.; Lotter, H.; Thomssen, R. Identification of an endoflagellar associated protein in Borrelia burgdorferi. J. Med. Microbiol. 1992, 36, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Rasiah, C.; Rauer, S.; Gassmann, G.S.; Vogt, A. Use of a hybrid protein consisting of the variable region of the Borrelia burgdorferi flagellin and part of the 83-kDa protein as antigen for serodiagnosis of Lyme disease. J. Clin. Microbiol. 1994, 32, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Rössler, D.; Eiffert, H.; Jauris-Heipke, S.; Lehnert, G.; Preac-Mursic, V.; Teepe, J.; Schlott, T.; Soutschek, E.; Wilske, B. Molecular and immunological characterization of the p83/100 protein of various Borrelia burgdorferi sensu lato strains. Med. Microbiol. Immunol. 1995, 184, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Ditton, H.J.; Neuss, M.; Zoller, L. Evidence that Borrelia burgdorferi immunodominant proteins p100, p94 and p83 are identical. FEMS Microbiol. Lett. 1992, 73, 217–220. [Google Scholar] [CrossRef]
- Embers, M.E.; Jacobs, M.B.; Johnson, B.J.; Philipp, M.T. Dominant epitopes of the C6 diagnostic peptide of Borrelia burgdorferi are largely inaccessible to antibody on the parent VlsE molecule. Clin. Vaccine Immunol. 2007, 14, 931–936. [Google Scholar] [CrossRef]
- Guder, W.G.; Ehret, W.; Da Fonseca-Wollheim, F.; Heil, W.; Müller-Plathe, O.; Schmitt, Y.; Töpfer, G.; Wisser, H.; Zawta, B. Die Qualität diagnostischer Proben. Lab. Medizin 2002, 26, 267–283. [Google Scholar] [CrossRef]
- Sturgeon, C.M.; Viljoen, A. Analytical error and interference in immunoassay: Minimizing risk. Ann. Clin. Biochem. 2011, 48, 418–432. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Seropositive | Equivocal | Seronegative | Vaccinated | |
|---|---|---|---|---|
| Canine test sera (n = 150) | 50 | 50 | 50 | |
| Canine control sera—positive/vaccination/negative (n = 50) * | 12 | 19 | 19 | |
| Equine test sera (n = 150) | 50 | 50 | 50 | |
| Equine control sera—vaccination (n = 50) ** | 50 |
| Antigen | LIA A | LIA B |
|---|---|---|
| VlsE | Ba | Bbss, Bg |
| OspA | Ba | Ba, Bg, Bbss |
| DbpA/p18 | Bbss, Bg, Ba, Bs | Bg, Bbav, Ba |
| OspC/p23 | Ba, Bbss, Bs, Bg, BgII | Ba, Bbav, Bbss |
| BmpA/p39 | Ba | Ba |
| p58 | Bg | Bbav |
| p83 | np | Ba |
| C6 | Bg | np |
| p41 | Bbss | np |
| p100 | Ba | np |
| Allocation Categories | Canine Sera | Equine Sera |
|---|---|---|
| neg | 0–1 AG lines ≥ COC except OspA or VlsE/C6 AG line | 0–2 AG lines ≥ COC except OspA or VlsE/C6 AG line |
| equiv | 2–3 AG lines ≥ COC except OspA or VlsE/C6 AG line | VlsE/C6 AG line and 0–2 AG lines ≥ COC or 3 AG lines except VlsE AG line ≥ COC |
| pos | VlsE/C6 AG line or ≥ 4 AG lines (except OspA AG line) ≥ COC | VlsE/C6 AG line and ≥ 3 AG lines ≥ COC or ≥ 4 AG lines ≥ COC or VlsE/C6 AG line and p18 AG line and ≥ 1 AG line ≥ COC |
| vac | OspA AG line and ≥ 0 AG lines (except VlsE/C6) ≥ COC | - |
| vac + pos | OspA AG line and VlsE/ C6 AG line and ≥0 AG lines ≥ COC | - |
| Allocation Categories | Canine Serum | Equine Serum |
|---|---|---|
| neg | 0 AG lines or AG lines ≤ COC or VlsE AG line = COC or 0–1 AG line (except VlsE and OspA) ≥ COC | 0 AG lines or AG lines ≤ COC or 0–2 AG line (except VlsE) ≥ COC |
| equiv | 2–3 AG lines ≥ COC (except VlsE and OspA) | 3 AG line (except VlsE) ≥ COC or VlsE AG line and 0–2 AG lines ≥ COC |
| pos | VlsE > COC or VlsE ≥ COC and 1 AG line (except OspA) or ≥ 4 AG lines (except VlsE and OspA) | VlsE AG line and ≥ 3 AG lines ≥ COC or VlsE AG line and DbpA AG line and 1 AG line or ≥ 4 AG lines ≥ COC (except VlsE) |
| vac | OspA AG line isolated or OspA AG line and ≥ 1 AG line (beside VlsE) or OspA AG line and VlsE AG line isolated = COC | - |
| vac + pos | OspA AG line and VlsE AG line isolated ≥ COC or OspA AG line and VlsE AG line and ≥ 1 AG line | - |
| Sample ID | Group | LIA A | LIA B | KELA Values |
|---|---|---|---|---|
| RKS-B-5365-C | Negative | neg | pos | 138.9 |
| RKS-B-7240-C | Negative | pos | neg | 151.7 |
| RKS-B-7279-C | Negative | vac | neg | 125.6 |
| RKS-B-8564-C | Negative | neg | pos | 127.0 |
| RKS-B-5119-C | Positive | pos | vac + pos | 394.6 |
| RKS-B-7996-C | Positive | pos | equiv | 103.7 |
| RKS-B-8113-C | Positive | pos | equiv | 243.1 |
| Hanka | Vaccinated | vac + pos | vac | 613.6 |
| RKS-B-5512-C | Vaccinated | neg | vac | 234.9 |
| RKS-B-8324-C | Vaccinated | vac + pos | vac | 580.9 |
| S98-5/1 * | Control sera | pos | neg | 63.3 |
| A93-3/3 * | Control sera | neg | vac | 383.2 |
| A93-3/4 * | Control sera | vac | neg | 311.2 |
| Groups | Positive (n = 50) | Negative (n = 50) | Vaccinated (n = 50) | Control Sera (n = 50) 1 | |
|---|---|---|---|---|---|
| LIA Results | |||||
| LIA A | pos | 47 | 2 | 0 | 12 |
| equiv | 0 | 0 | 0 | 0 | |
| neg | 3 | 47 | 0 | 20 | |
| vac | 0 | 1 | 50 ** | 18 | |
| LIA B | pos | 45 * | 2 | 0 | 12 |
| equiv | 2 | 0 | 0 | 0 | |
| neg | 3 | 48 | 0 | 20 | |
| vac | 0 | 0 | 50 *** | 18 |
| Antigen Group | VlsE | OspA | DbpA/p18 | OspC | BmpA/p39 | p58 |
|---|---|---|---|---|---|---|
| canneg | 0.236 | 0.111 | 0.331 | 0.123 | 0.560 | 0.580 |
| canpos | 0.852 | 0.147 | 0.156 | 0.645 | 0.850 | 0.574 |
| canvacc | 0.411 | 0.460 | −0.084 | 0.772 | 0.577 | 0.464 |
| cancon | 0.777 | 0.956 | 0.096 | 0.805 | 0.785 | 0.508 |
| equneg | 0.575 | 0.123 | 0.572 | 0.254 | 0.169 | 0.414 |
| equpos | 0.636 | 0.084 | 0.358 | 0.315 | 0.565 | 0.445 |
| equvacc | 0.665 | 0.668 | 0.015 | 0.559 | 0.523 | 0.234 |
| equequiv | 0.367 | 0.083 | 0.186 | 0.560 | 0.551 | 0.459 |
| Sample ID | Group | LIA A with OspA * | LIA A without OspA ** | LIA B | KELA Values |
|---|---|---|---|---|---|
| S4-8 1 | Control sera—vaccinated | equiv | neg | neg | 654.3 |
| S4-32 1 | Control sera—vaccinated | pos | equiv | equiv | 735.0 |
| S4-180 1 | Control sera—vaccinated | pos | equiv | equiv | 688.6 |
| S7-43 1 | Control sera—vaccinated | equiv | neg | neg | 728.9 |
| S7-44 1 | Control sera—vaccinated | pos | equiv | neg | 633.8 |
| S7-58 1 | Control sera—vaccinated | equiv | neg | neg | 725.4 |
| S7-61 1 | Control sera—vaccinated | pos | equiv | neg | 694.8 |
| S1-5 1 | Control sera—vaccinated | equiv | neg | pos | 568.4 |
| Sample ID | Group | LIA A | LIA B | KELA Values |
|---|---|---|---|---|
| S4-5 1 | Control sera—vaccinated | pos | pos | 584.0 |
| S4-26 1 | Control sera—vaccinated | pos | pos | 659.9 |
| S4-30 1 | Control sera—vaccinated | pos * | pos | 709.4 |
| S4-31 1 | Control sera—vaccinated | pos | pos | 697.8 |
| S4-176 1 | Control sera—vaccinated | pos ** | pos | 722.4 |
| S7-86 1 | Control sera—vaccinated | pos * | pos | 731.8 |
| S1-5 1 | Control sera—vaccinated | equiv | pos | 568.4 |
| S4-28 1 | Control sera—vaccinated | neg | pos | 661.2 |
| S5-31 1 | Control sera—vaccinated | neg | pos | 646.9 |
| Groups | Positive (n = 50) | Equivocal (n = 50) | Negative (n = 50) | Control Sera-Vaccinated 1 (n = 50) | |
|---|---|---|---|---|---|
| LIA Results | |||||
| LIA A | pos | 29 | 25 | 5 | 0 |
| equiv | 20 | 22 | 6 | 1 | |
| neg | 1 | 3 | 38 | 2 | |
| vac | 0 | 0 | 0 | 47 * | |
| LIA B | pos | 17 | 6 | 1 | 0 |
| equiv | 28 | 23 | 2 | 1 | |
| neg | 4 | 21 | 47 | 1 | |
| vac | 1 * | 0 | 0 | 48 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doff, S.C.; Wenderlein, J.; Wiesinger, A.; Hiereth, S.; Ulrich, S.; Straubinger, R.K. Detection of Borrelia burgdorferi sensu-lato-Specific Antibodies in Sera of Canine and Equine Origin—A Comparative Study with Two Line Immunoassays. Vet. Sci. 2022, 9, 633. https://doi.org/10.3390/vetsci9110633
Doff SC, Wenderlein J, Wiesinger A, Hiereth S, Ulrich S, Straubinger RK. Detection of Borrelia burgdorferi sensu-lato-Specific Antibodies in Sera of Canine and Equine Origin—A Comparative Study with Two Line Immunoassays. Veterinary Sciences. 2022; 9(11):633. https://doi.org/10.3390/vetsci9110633
Chicago/Turabian StyleDoff, Sophie Charlotte, Jasmin Wenderlein, Anna Wiesinger, Stephanie Hiereth, Sebastian Ulrich, and Reinhard K. Straubinger. 2022. "Detection of Borrelia burgdorferi sensu-lato-Specific Antibodies in Sera of Canine and Equine Origin—A Comparative Study with Two Line Immunoassays" Veterinary Sciences 9, no. 11: 633. https://doi.org/10.3390/vetsci9110633