
OCT Meets micro-CT: A Subject-Specific Correlative Multimodal Imaging Workflow for Early Chick Heart Development Modeling

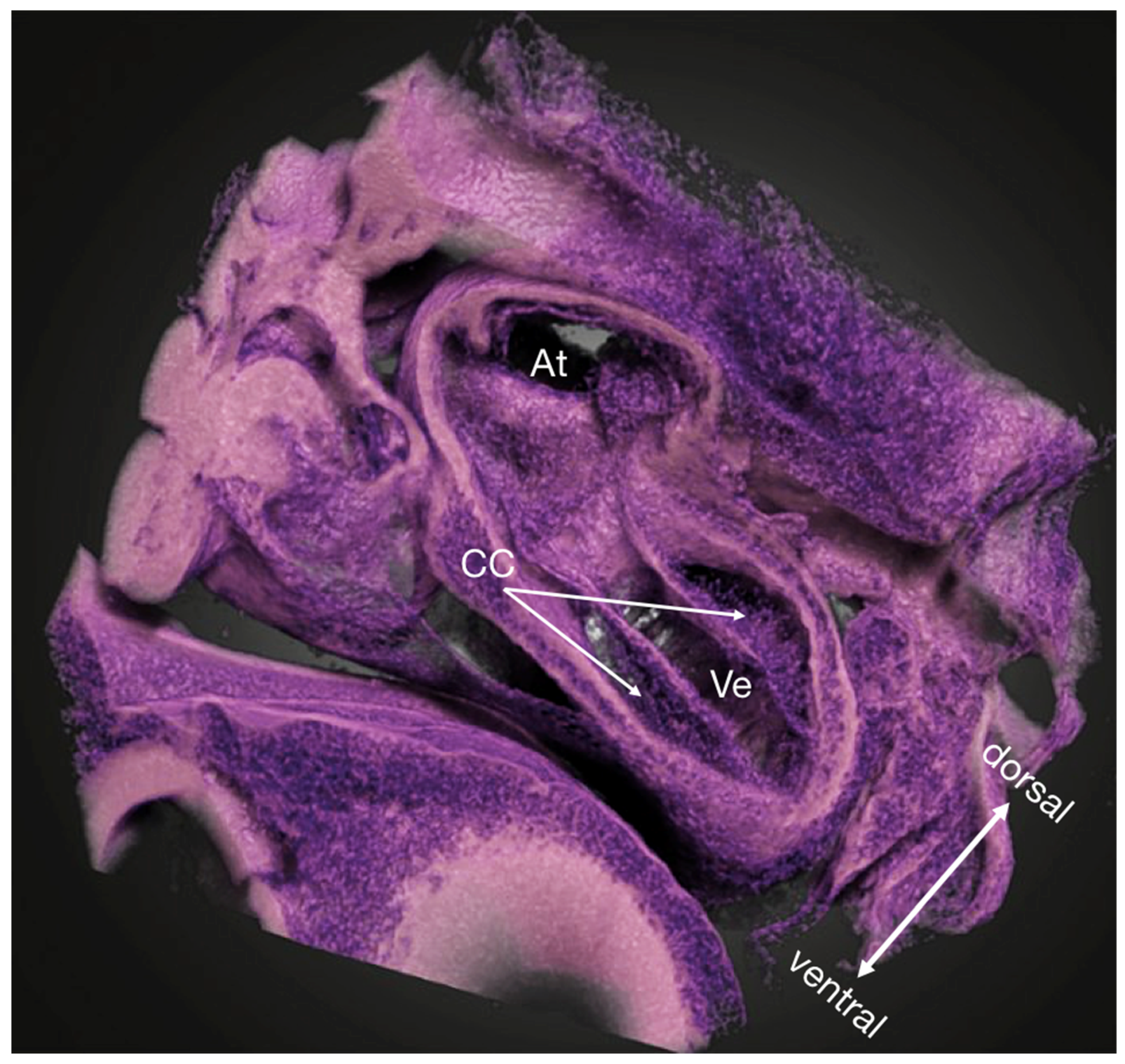{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Embryo Preparation
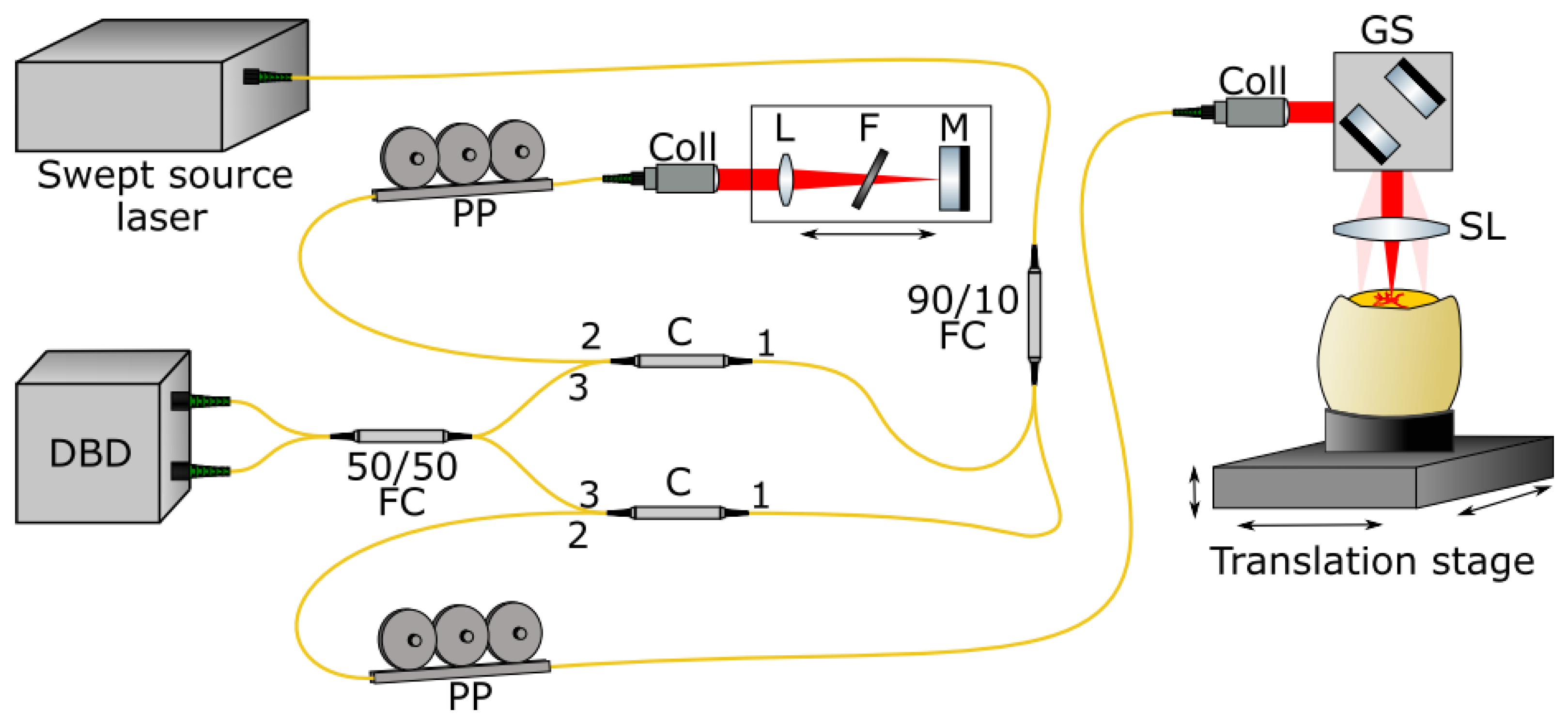
2.2. Optical Coherence Tomography
2.3. Subsequent micro-CT Imaging
2.4. Additional micro-CT Imaging
3. Results
3.1. Embryo Preparation
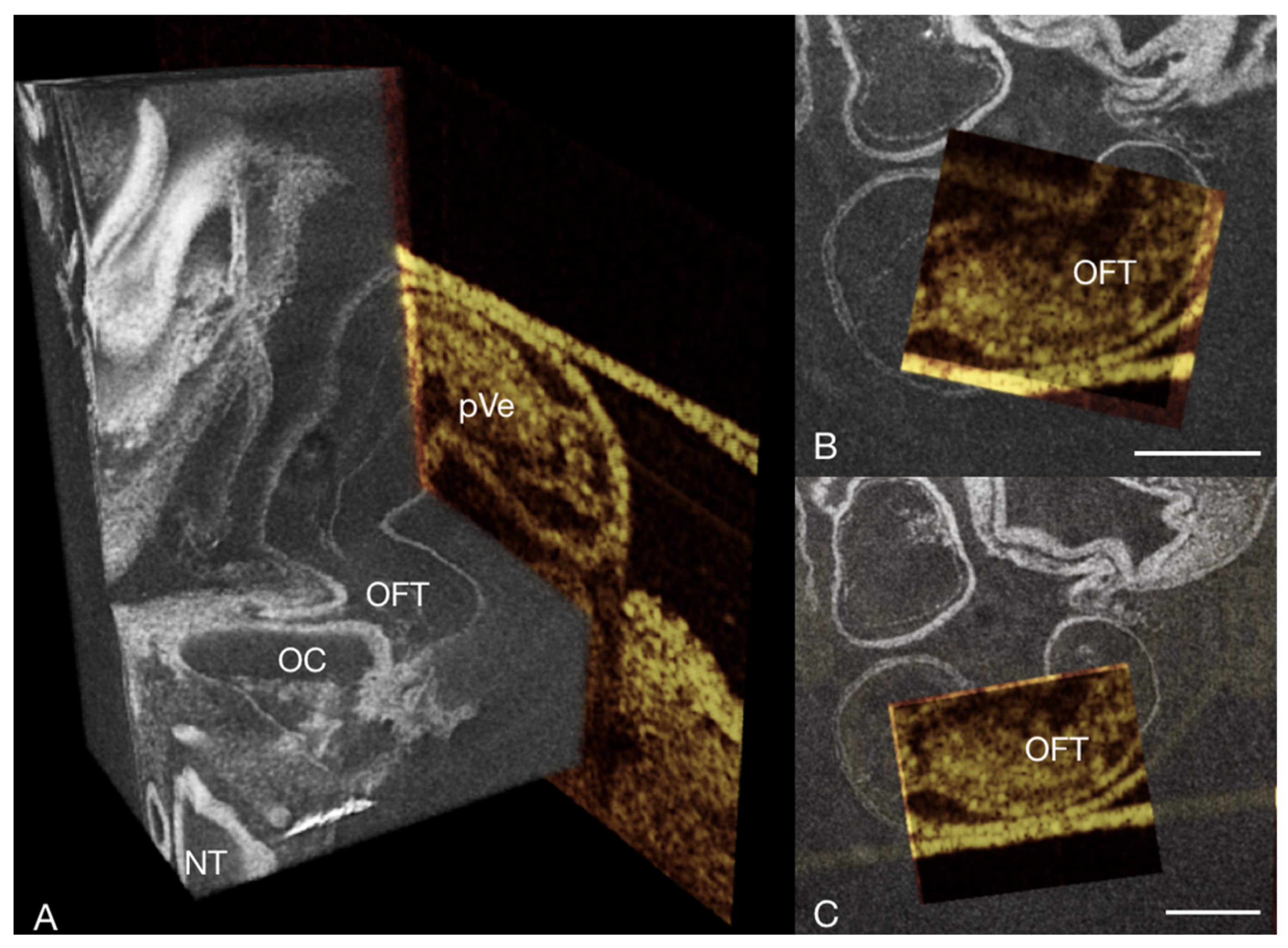
3.2. Image Registration
3.3. Other Findings and Considerations
3.3.1. Staining Used
3.3.2. Using micro-CT for Doppler OCT Based CFD Model Validation
3.3.3. Reproducibility
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lansford, R.; Rugonyi, S. Follow Me! A Tale of Avian Heart Development with Comparisons to Mammal Heart Development. J. Cardiovasc. Dev. Dis. 2020, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeHaan, R.L. Development of form in the embryonic heart an experimental approach. Circulation 1967, 35, 821–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burggren, W.W.; Santin, J.F.; Antich, M.R. Cardio-respiratory development in bird embryos: New insights from a venerable animal model. Rev. Bras. Zootec. 2016, 45, 709–728. [Google Scholar] [CrossRef] [Green Version]
- Midgett, M.; Rugonyi, S. Congenital heart malformations induced by hemodynamic altering surgical interventions. Front. Physiol. 2014, 5, 287. [Google Scholar] [CrossRef] [Green Version]
- Midgett, M.; Thornburg, K.; Rugonyi, S. Blood flow patterns underlie developmental heart defects. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H632–H642. [Google Scholar] [CrossRef]
- Roest, P.A.; van Iperen, L.; Vis, S.; Wisse, L.J.; Poelmann, R.E.; Steegers-Theunissen, R.P.; Molin, D.G.; Eriksson, U.J.; Gittenberger-De Groot, A.C. Exposure of neural crest cells to elevated glucose leads to congenital heart defects, an effect that can be prevented by N-acetylcysteine. Birth Defects Res. Part A: Clin. Mol. Teratol. 2007, 79, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Neeb, Z.; Lajiness, J.D.; Bolanis, E.; Conway, S.J. Cardiac outflow tract anomalies. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 499–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goenezen, S.; Chivukula, V.K.; Midgett, M.; Phan, L.; Rugonyi, S. 4D subject-specific inverse modeling of the chick embryonic heart outflow tract hemodynamics. Biomech. Model. Mechanobiol. 2015, 15, 723–743. [Google Scholar] [CrossRef] [Green Version]
- Drexler, W.; Fujimoto, J.G. (Eds.) Optical Coherence Tomography: Technology and Applications, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Leitgeb, R.; Placzek, F.; Rank, E.; Krainz, L.; Haindl, R.; Li, Q.; Liu, M.; Andreana, M.; Unterhuber, A.; Schmoll, T. Enhanced medical diagnosis for dOCTors: A perspective of optical coherence tomography. J. Biomed. Opt. 2021, 26, 100601. [Google Scholar] [CrossRef]
- Carrasco-Zevallos, O.; Keller, B.; Viehland, C.; Shen, L.; Waterman, G.; Todorich, B.; Shieh, C.; Hahn, P.; Farsiu, S.; Kuo, A. Live volumetric (4D) visualization and guidance of in vivo human ophthalmic surgery with intraoperative optical coherence tomography. Sci. Rep. 2016, 6, 31689. [Google Scholar] [CrossRef]
- Gora, M.J.; Suter, M.J.; Tearney, G.J.; Li, X. Endoscopic optical coherence tomography: Technologies and clinical applications. Biomed. Opt. Express 2017, 8, 2405–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placzek, F.; Bautista, E.C.; Kretschmer, S.; Wurster, L.M.; Knorr, F.; González-Cerdas, G.; Erkkilä, M.T.; Stein, P.; Ataman, Ç.; Hermann, G.G. Morpho-molecular ex vivo detection and grading of non-muscle-invasive bladder cancer using forward imaging probe based multimodal optical coherence tomography and Raman spectroscopy. Analyst 2020, 145, 1445–1456. [Google Scholar] [CrossRef] [Green Version]
- Schie, I.W.; Placzek, F.; Knorr, F.; Cordero, E.; Wurster, L.M.; Hermann, G.G.; Mogensen, K.; Hasselager, T.; Drexler, W.; Popp, J. Morpho-molecular signal correlation between optical coherence tomography and Raman spectroscopy for superior image interpretation and clinical diagnosis. Sci. Rep. 2021, 11, 9951. [Google Scholar] [CrossRef]
- Baumann, B. Polarization sensitive optical coherence tomography: A review of technology and applications. Appl. Sci. 2017, 7, 474. [Google Scholar] [CrossRef] [Green Version]
- Leitgeb, R.A.; Baumann, B. Multimodal optical medical imaging concepts based on optical coherence tomography. Front. Phys. 2018, 6, 114. [Google Scholar] [CrossRef] [Green Version]
- Chivukula, V.K.; Goenezen, S.; Liu, A.; Rugonyi, S. Effect of outflow tract banding on embryonic cardiac hemodynamics. J. Cardiovasc. Dev. Dis. 2015, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Walter, A.; Paul-Gilloteaux, P.; Plochberger, B.; Sefc, L.; Verkade, P.; Mannheim, J.G.; Slezak, P.; Unterhuber, A.; Marchetti-Deschmann, M.; Ogris, M. Correlated multimodal imaging in life sciences: Expanding the biomedical horizon. Front. Phys. 2020, 8, 47. [Google Scholar] [CrossRef]
- Placzek, F.; Micko, A.; Sentosa, R.; Fonollà, R.; Winklehner, M.; Hosmann, A.; Andreana, M.; Höftberger, R.; Drexler, W.; Leitgeb, R.A. Towards ultrahigh resolution OCT based endoscopical pituitary gland and adenoma screening: A performance parameter evaluation. Biomed. Opt. Express 2020, 11, 7003–7018. [Google Scholar] [CrossRef]
- Binding, J.; Arous, J.B.; Léger, J.-F.; Gigan, S.; Boccara, C.; Bourdieu, L. Brain refractive index measured in vivo with high-NA defocus-corrected full-field OCT and consequences for two-photon microscopy. Opt. Express 2011, 19, 4833–4847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scully, D.M.; Larina, I.V. Mouse embryo phenotyping with optical coherence tomography. Front. Cell Dev. Biol. 2022, 10, 1894. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Withers, P.J.; Bouman, C.; Carmignato, S.; Cnudde, V.; Grimaldi, D.; Hagen, C.K.; Maire, E.; Manley, M.; Du Plessis, A.; Stock, S.R. X-ray computed tomography. Nat. Rev. Methods Prim. 2021, 1, 18. [Google Scholar] [CrossRef]
- Leyssens, L.; Pestiaux, C.; Kerckhofs, G. A Review of Ex Vivo X-ray Microfocus Computed Tomography-Based Characterization of the Cardiovascular System. Int. J. Mol. Sci. 2021, 22, 3263. [Google Scholar] [CrossRef] [PubMed]
- Novo Matos, J.; Garcia-Canadilla, P.; Simcock, I.C.; Hutchinson, J.C.; Dobromylskyj, M.; Guy, A.; Arthurs, O.J.; Cook, A.C.; Luis Fuentes, V. Micro-computed tomography (micro-CT) for the assessment of myocardial disarray, fibrosis and ventricular mass in a feline model of hypertrophic cardiomyopathy. Sci. Rep. 2020, 10, 20169. [Google Scholar] [CrossRef] [PubMed]
- Dejea, H.; Bonnin, A.; Cook, A.C.; Garcia-Canadilla, P. Cardiac multi-scale investigation of the right and left ventricle ex vivo: A review. Cardiovasc. Diagn. Ther. 2020, 10, 1701–1717. [Google Scholar] [CrossRef]
- Karimi, A.; Navidbakhsh, M.; Rezaee, T.; Hassani, K. Measurement of the circumferential mechanical properties of the umbilical vein: Experimental and numerical analyses. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 1418–1426. [Google Scholar] [CrossRef]
- Siddiqui, H.B.; Dogru, S.; Lashkarinia, S.S.; Pekkan, K. Soft-Tissue Material Properties and Mechanogenetics during Cardiovascular Development. J. Cardiovasc. Dev. Dis. 2022, 9, 64. [Google Scholar] [CrossRef]
- Handschuh, S.; Glösmann, M. Mouse embryo phenotyping using X-ray microCT. Front. Cell Dev. Biol. 2022, 10, 949184. [Google Scholar] [CrossRef]
- Degenhardt, K.; Wright, A.C.; Horng, D.; Padmanabhan, A.; Epstein, J.A. Rapid 3D phenotyping of cardiovascular development in mouse embryos by micro-CT with iodine staining. Circ. Cardiovasc. Imaging 2010, 3, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.T.; Hansen, M.S.; Wu, I.; Healy, L.J.; Johnson, C.R.; Jones, G.M.; Capecchi, M.R.; Keller, C. Virtual histology of transgenic mouse embryos for high-throughput phenotyping. PLoS Genet. 2006, 2, e61. [Google Scholar] [CrossRef]
- Nagase, T.; Sasazaki, Y.; Kikuchi, T.; Machida, M. Rapid 3-dimensional imaging of embryonic craniofacial morphology using microscopic computed tomography. J. Comput. Assist. Tomogr. 2008, 32, 816–821. [Google Scholar] [CrossRef]
- Tobita, K.; Liu, X.; Lo, C.W. Imaging modalities to assess structural birth defects in mutant mouse models. Birth Defects Res. Part C Embryo Today Rev. 2010, 90, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.C.; Dover, S.D. X-ray microtomography. J. Microsc. 1982, 126, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Neues, F.; Epple, M. X-ray Microcomputer Tomography for the Study of Biomineralized Endo- and Exoskeletons of Animals. Chem. Rev. 2008, 108, 4734–4741. [Google Scholar] [CrossRef] [PubMed]
- de Bournonville, S.; Vangrunderbeeck, S.; Kerckhofs, G. Contrast-Enhanced MicroCT for Virtual 3D Anatomical Pathology of Biological Tissues: A Literature Review. Contrast Media Mol. Imaging 2019, 2019, 8617406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metscher, B.D. MicroCT for developmental biology: A versatile tool for high-contrast 3D imaging at histological resolutions. Dev. Dyn. 2009, 238, 632–640. [Google Scholar] [CrossRef]
- Aminu, A.J.; Chen, W.; Yin, Z.; Kuniewicz, M.; Walocha, J.; Perde, F.; Molenaar, P.; Iaizzo, P.A.; Dobrzynski, H.; Atkinson, A.J. Novel micro-computed tomography contrast agents to visualise the human cardiac conduction system and surrounding structures in hearts from normal, aged, and obese individuals. Transl. Res. Anat. 2022, 27, 100175. [Google Scholar] [CrossRef]
- Aslanidi, O.V.; Nikolaidou, T.; Zhao, J.; Smaill, B.H.; Gilbert, S.H.; Holden, A.V.; Lowe, T.; Withers, P.J.; Stephenson, R.S.; Jarvis, J.C.; et al. Application of Micro-Computed Tomography With Iodine Staining to Cardiac Imaging, Segmentation, and Computational Model Development. IEEE Trans. Med. Imaging 2013, 32, 8–17. [Google Scholar] [CrossRef]
- Dunmore-Buyze, P.J.; Tate, E.; Xiang, F.l.; Detombe, S.A.; Nong, Z.; Pickering, J.G.; Drangova, M. Three-dimensional imaging of the mouse heart and vasculature using micro-CT and whole-body perfusion of iodine or phosphotungstic acid. Contrast Media Mol. Imaging 2014, 9, 383–390. [Google Scholar] [CrossRef]
- Helfenstein-Didier, C.; Taïnoff, D.; Viville, J.; Adrien, J.; Maire, É.; Badel, P. Tensile rupture of medial arterial tissue studied by X-ray micro-tomography on stained samples. J. Mech. Behav. Biomed. Mater. 2018, 78, 362–368. [Google Scholar] [CrossRef]
- Nierenberger, M.; Rémond, Y.; Ahzi, S.; Choquet, P. Assessing the three-dimensional collagen network in soft tissues using contrast agents and high resolution micro-CT: Application to porcine iliac veins. Comptes Rendus Biol. 2015, 338, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Handschuh, S.; Okada, C.T.C.; Walter, I.; Aurich, C.; Glosmann, M. An optimized workflow for microCT imaging of formalin-fixed and paraffin-embedded (FFPE) early equine embryos. Anat. Histol. Embryol. 2022, 51, 611–623. [Google Scholar] [CrossRef]
- Kraus, N.; Metscher, B. Anuran development: A reinvestigation of the conus arteriosus and gill formation in Bufo bufo throughout metamorphosis using micro-CT. Anat. Rec. 2022, 305, 1100–1111. [Google Scholar] [CrossRef] [PubMed]
- Busse, M.; Muller, M.; Kimm, M.A.; Ferstl, S.; Allner, S.; Achterhold, K.; Herzen, J.; Pfeiffer, F. Three-dimensional virtual histology enabled through cytoplasm-specific X-ray stain for microscopic and nanoscopic computed tomography. Proc. Natl. Acad. Sci. USA 2018, 115, 2293–2298. [Google Scholar] [CrossRef] [Green Version]
- Dullin, C.; Ufartes, R.; Larsson, E.; Martin, S.; Lazzarini, M.; Tromba, G.; Missbach-Guentner, J.; Pinkert-Leetsch, D.; Katschinski, D.M.; Alves, F. μCT of ex-vivo stained mouse hearts and embryos enables a precise match between 3D virtual histology, classical histology and immunochemistry. PLoS ONE 2017, 12, e0170597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabner, S.; Böck, P.; Fink, D.; Glösmann, M.; Handschuh, S. The visible skeleton 2.0: Phenotyping of cartilage and bone in fixed vertebrate embryos and foetuses based on X-ray microCT. Development 2020, 147, dev187633. [Google Scholar] [CrossRef]
- Müller, M.; Kimm, M.A.; Ferstl, S.; Allner, S.; Achterhold, K.; Herzen, J.; Pfeiffer, F.; Busse, M. Nucleus-specific X-ray stain for 3D virtual histology. Sci. Rep. 2018, 8, 17855. [Google Scholar] [CrossRef] [Green Version]
- Metscher, B. A simple nuclear contrast staining method for microCT-based 3D histology using lead(II) acetate. J. Anat. 2021, 238, 1036–1041. [Google Scholar] [CrossRef]
- Ermakova, O.; Orsini, T.; Fruscoloni, P.; Chiani, F.; Gambadoro, A.; Putti, S.; Cirilli, M.; Mezzi, A.; Kaciulis, S.; Pasquini, M.; et al. Three-Dimensional X-ray Imaging of beta-Galactosidase Reporter Activity by Micro-CT: Implication for Quantitative Analysis of Gene Expression. Brain Sci. 2021, 11, 746. [Google Scholar] [CrossRef]
- Metscher, B.D. X-ray microtomographic imaging of intact vertebrate embryos. Cold Spring Harb. Protoc. 2011, 2011, 1462–1471. [Google Scholar] [CrossRef]
- Tyler, M.S. Developmental Biology: A Guide for Experimental Study; Sinauer Associates: Sunderland, MA, USA, 1994. [Google Scholar]
- Stekelenburg-de Vos, S.; Steendijk, P.; Ursem, N.T.; Wladimiroff, J.W.; Delfos, R.; Poelmann, R.E. Systolic and diastolic ventricular function assessed by pressure-volume loops in the stage 21 venous clipped chick embryo. Pediatr. Res. 2005, 57, 16–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Männer, J.; Thrane, L.; Norozi, K.; Yelbuz, T.M. High-resolution in vivo imaging of the cross-sectional deformations of contracting embryonic heart loops using optical coherence tomography. Dev. Dyn. 2008, 237, 953–961. [Google Scholar] [CrossRef] [PubMed]
- McQuinn, T.C.; Bratoeva, M.; DeAlmeida, A.; Remond, M.; Thompson, R.P.; Sedmera, D. High-frequency ultrasonographic imaging of avian cardiovascular development. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 3503–3513. [Google Scholar] [CrossRef] [PubMed]
- Metscher, B.D. MicroCT for comparative morphology: Simple staining methods allow high-contrast 3D imaging of diverse non-mineralized tissues. BMC Physiol. 2009, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metscher, B. Fiji (ImageJ) plugin to read Xradia XRM files. Zenodo 2022. [Google Scholar] [CrossRef]
- Anbara, T.; Sharifi, M.; Aboutaleb, N. Endothelial to mesenchymal transition in the cardiogenesis and cardiovascular diseases. Curr. Cardiol. Rev. 2020, 16, 306–314. [Google Scholar] [CrossRef]
- Kovacic, J.C.; Dimmeler, S.; Harvey, R.P.; Finkel, T.; Aikawa, E.; Krenning, G.; Baker, A.H. Endothelial to Mesenchymal Transition in Cardiovascular Disease. J. Am. Coll. Cardiol. 2019, 73, 190–209. [Google Scholar] [CrossRef]
- Menon, V.; Eberth, J.; Goodwin, R.; Potts, J. Altered Hemodynamics in the Embryonic Heart Affects Outflow Valve Development. J. Cardiovasc. Dev. Dis. 2015, 2, 108–124. [Google Scholar] [CrossRef]
- Von Gise, A.; Pu, W.T. Endocardial and epicardial epithelial to mesenchymal transitions in heart development and disease. Circ. Res. 2012, 110, 1628–1645. [Google Scholar] [CrossRef]
- Hsu, C.W.; Kalaga, S.; Akoma, U.; Rasmussen, T.L.; Christiansen, A.E.; Dickinson, M.E. High Resolution Imaging of Mouse Embryos and Neonates with X-ray Micro-Computed Tomography. Curr. Protoc. Mouse Biol. 2019, 9, e63. [Google Scholar] [CrossRef]
- Courchaine, K.; Rugonyi, S. Optical coherence tomography for in vivo imaging of endocardial to mesenchymal transition during avian heart development. Biomed. Opt. Express 2019, 10, 5989. [Google Scholar] [CrossRef] [PubMed]
- Handschuh, S.; Beisser, C.J.; Ruthensteiner, B.; Metscher, B.D. Microscopic dual-energy CT (microDECT): A flexible tool for multichannel ex vivo 3D imaging of biological specimens. J. Microsc. 2017, 267, 3–26. [Google Scholar] [CrossRef]
- Liu, A.; Wang, R.K.; Thornburg, K.L.; Rugonyi, S. Dynamic variation of hemodynamic shear stress on the walls of developing chick hearts: Computational models of the heart outflow tract. Eng. Comput. 2009, 25, 73–86. [Google Scholar] [CrossRef]
- Yin, X.; Liu, A.; Thornburg, K.L.; Wang, R.K.; Rugonyi, S. Extracting cardiac shapes and motion of the chick embryo heart outflow tract from four-dimensional optical coherence tomography images. J. Biomed. Opt. 2012, 17, 096005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tearney, G.J.; Boppart, S.A.; Bouma, B.E.; Brezinski, M.E.; Weissman, N.J.; Southern, J.F.; Fujimoto, J.G. Scanning single-mode fiber optic catheter–endoscope for optical coherence tomography. Opt. Lett. 1996, 21, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Westphal, V.; Yazdanfar, S.; Rollins, A.M.; Izatt, J.A. Real-time, high velocity-resolution color Doppler optical coherence tomography. Opt. Lett. 2002, 27, 34–36. [Google Scholar] [CrossRef] [PubMed]
- Zvyagin, A.V.; Silva, K.D.; Alexandrov, S.A.; Hillman, T.R.; Armstrong, J.J.; Tsuzuki, T.; Sampson, D.D. Refractive index tomography of turbid media by bifocal optical coherence refractometry. Opt. Express 2003, 11, 3503–3517. [Google Scholar] [CrossRef] [Green Version]
- Butcher, J.T.; Sedmera, D.; Guldberg, R.E.; Markwald, R.R. Quantitative volumetric analysis of cardiac morphogenesis assessed through micro-computed tomography. Dev. Dyn. 2007, 236, 802–809. [Google Scholar] [CrossRef]
- Wong, M.D.; Spring, S.; Henkelman, R.M. Structural Stabilization of Tissue for Embryo Phenotyping Using Micro-CT with Iodine Staining. PLoS ONE 2013, 8, e84321. [Google Scholar]
- Dawood, Y.; Hagoort, J.; Siadari, B.; Ruijter, J.; Gunst, Q.; Lobe, N.; Strijkers, G.; de Bakker, B.; van den Hoff, M. Reducing soft-tissue shrinkage artefacts caused by staining with Lugol’s solution. Sci Rep. 2021, 11, 19781. [Google Scholar] [CrossRef]
- Jenkins, M.; Adler, D.; Gargesha, M.; Huber, R.; Rothenberg, F.; Belding, J.; Watanabe, M.; Wilson, D.; Fujimoto, J.; Rollins, A. Ultrahigh-speed optical coherence tomography imaging and visualization of the embryonic avian heart using a buffered Fourier Domain Mode Locked laser. Opt. Express 2007, 15, 6251–6267. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.J.; Francis, R.; Liu, X.; Devine, W.A.; Ramirez, R.; Anderton, S.J.; Wong, L.Y.; Faruque, F.; Gabriel, G.C.; Leatherbury, L. Microcomputed tomography provides high accuracy congenital heart disease diagnosis in neonatal and fetal mice. Circ. Cardiovasc. Imaging 2013, 6, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, C.M.; Zambelli, V.; Botta, G.; Moltrasio, F.; Cattoretti, G.; Lucchini, V.; Fesslova, V.; Cuttin, M.S. Postmortem microcomputed tomography (micro-CT) of small fetuses and hearts. Ultrasound Obstet. Gynecol. 2014, 44, 600–609. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraus, N.; Placzek, F.; Metscher, B. OCT Meets micro-CT: A Subject-Specific Correlative Multimodal Imaging Workflow for Early Chick Heart Development Modeling. J. Cardiovasc. Dev. Dis. 2022, 9, 379. https://doi.org/10.3390/jcdd9110379
Kraus N, Placzek F, Metscher B. OCT Meets micro-CT: A Subject-Specific Correlative Multimodal Imaging Workflow for Early Chick Heart Development Modeling. Journal of Cardiovascular Development and Disease. 2022; 9(11):379. https://doi.org/10.3390/jcdd9110379
Chicago/Turabian StyleKraus, Nina, Fabian Placzek, and Brian Metscher. 2022. "OCT Meets micro-CT: A Subject-Specific Correlative Multimodal Imaging Workflow for Early Chick Heart Development Modeling" Journal of Cardiovascular Development and Disease 9, no. 11: 379. https://doi.org/10.3390/jcdd9110379