The PHR Family: The Role of Extracellular Transglycosylases in Shaping Candida albicans Cells



1
Department of Biosciences, University of Milan, Via Celoria 26, Milan 20133, Italy
2
Department of Microbiology & Immunology, Georgetown University, Washington, DC 20057, USA
*
Author to whom correspondence should be addressed.
J. Fungi 2017, 3(4), 59; https://doi.org/10.3390/jof3040059
Submission received: 2 October 2017
/
Revised: 19 October 2017
/
Accepted: 24 October 2017
/
Published: 29 October 2017
(This article belongs to the Special Issue Fungal Cell Wall)
Abstract
:Candida albicans is an opportunistic microorganism that can become a pathogen causing mild superficial mycosis or more severe invasive infections that can be life-threatening for debilitated patients. In the etiology of invasive infections, key factors are the adaptability of C. albicans to the different niches of the human body and the transition from a yeast form to hypha. Hyphal morphology confers high adhesiveness to the host cells, as well as the ability to penetrate into organs. The cell wall plays a crucial role in the morphological changes C. albicans undergoes in response to specific environmental cues. Among the different categories of enzymes involved in the formation of the fungal cell wall, the GH72 family of transglycosylases plays an important assembly role. These enzymes cut and religate β-(1,3)-glucan, the major determinant of cell shape. In C. albicans, the PHR family encodes GH72 enzymes, some of which work in specific environmental conditions. In this review, we will summarize the work from the initial discovery of PHR genes to the study of the pH-dependent expression of PHR1 and PHR2, from the characterization of the gene products to the recent findings concerning the stress response generated by the lack of GH72 activity in C. albicans hyphae.
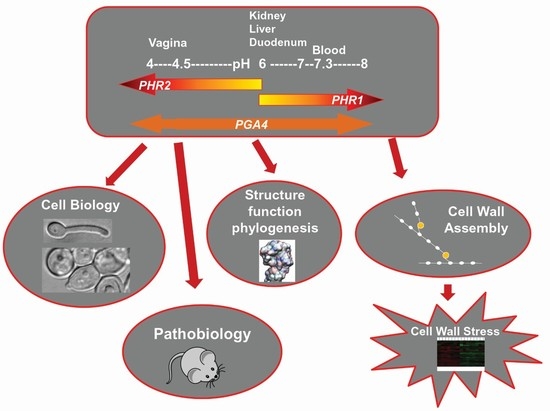
1. Introduction
Candida albicans is routinely found as a benign commensal inhabitant of human skin and various mucosal surfaces of the alimentary and genitourinary tract [1]. However, as an opportunistic fungal pathogen it is a frequent agent of non-invasive infections and life-threatening invasive infections in immune-compromised patients [2]. It has been estimated that C. albicans causes >400,000 life-threatening infections per year worldwide, with mortality rates of 46–75% [3]. Despite being restricted to the human host as an ecological niche, C. albicans is nonetheless exposed to diverse and dynamic microenvironments (mucosae, skin, gastrointestinal tract), which it must sense and to which it must constantly adapt. The interface of this constant exchange is the cell wall. The fungal cell wall is a dynamic structure that changes in response to environmental stresses, including host–pathogen interaction [4,5]. The most overt cell wall changes are manifested in the morphological transitions of C. albicans between yeast, pseudohyphae and hyphae, transitions that are critical to its pathogenicity [6].
From a practical perspective, the fungal cell wall has long been considered a desirable target of antifungal drugs given its essentiality and unique structure and biosynthesis relative to the host. The value of this target was demonstrated with the introduction of the echinocandins, which inhibit glucan synthase, the enzyme responsible for production of the major cell wall polysaccharide β-(1,3)-glucan [7]. This glucan is largely responsible for the integrity and shape of the cell wall. The steps leading from the initial synthesis of β-(1,3)-glucan to its modification, interconnection, and incorporation into the three-dimensional architecture of the cell wall remain poorly defined [8]. However, the GH72 transglycosylases are important players in this complex assembly process. The absence of these enzymes has severe consequences, altering growth, morphology, and, virulence. In C. albicans the GH72 enzymes are encoded by the PHR gene family, some of which are expressed in response to specific environmental stresses. In this review, we will summarize the work from the initial discovery of PHR genes to the study of the pH-dependent expression of PHR1 and PHR2, from the characterization of the gene products to the recent findings concerning the stress response generated by the lack of GH72 activity in C. albicans hyphae. Elucidating these processes will lead to a fuller understanding of cell wall dynamics and may suggest new therapeutic approaches to this problematic pathogen.
2. Identification of the PHR Multigene Family
Discovery of the initial members of the PHR gene family in C. albicans, like the discovery of the orthologous GAS genes in Saccharomyces cerevisiae, was a fortuitous outcome of studies unrelated to fungal cell wall biosynthesis or structure. Gas1p was first identified as a cell-cycle-regulated protein [9], subsequently shown to be glycosylated [10] and one of the predominant glycosylphosphatidyl inositol (GPI)-linked proteins of the yeast plasma membrane [11,12]. In contrast, PHR1 was uncovered in a study designed to identify genes expressed specifically in the yeast or hyphal growth phase [13]. The morphology of C. albicans can be controlled in vitro by altering culture conditions, pH and temperature being two important variables. In a screening for differentially expressed genes, cells were maintained in the yeast morphology by culturing at 25 °C, pH 4.5 and induced to form germ tubes, the initial phase of hyphal outgrowth, at 37 °C, pH 6.5 [13]. Differential hybridization of cDNA prepared from these cultures identified morphology-specific genes as well as one gene regulated specifically in response to the pH of the culture medium [14]. This gene was designated PHR1 (pH responsive). DNA sequence analysis showed PHR1 to be closely related to GAS1 [14] and heterologous complementation of a GAS1 null mutant demonstrated their functional equivalence [15].
The existence in C. albicans of additional PHR1-related genes was suggested by the presence of multiple proteins cross-reactive with anti-Phr1p antisera and the pH-conditional phenotype of phr1 null mutants [14]. One of the presumptive paralogs was identified by PCR using degenerate primers that targeted sequences conserved between PHR1 and GAS1 to amplify related sequences from template DNA of a phr1 deletion mutant [16]. A single PCR product was obtained. Isolation and sequencing of the corresponding gene showed it to encode a protein co-linear with Phr1p and about 53% identical. This gene was designated PHR2. Although multiple primer sets were tested, PHR2 was the only gene identified.
The third member of the PHR family, PHR3, was found by chance sequencing of an EST library clone [17], while two additional family members were identified by in silico means. The latter two, PGA4 and PGA5, were found by an algorithmic search of the C. albicans genome designed to recognize putative GPI-modified proteins [18], thus their names, predicted GPI-anchored proteins [18]. These three genes are more divergent than PHR1 and PHR2, the encoded proteins are only 25–35% identical to Phr1p and Phr2p and to each other [17]. PHR3 is distinguished as the only gene in the family to contain an intron and to encode a protein lacking an identifiable GPI attachment site.
Expression Pattern of PHR Multigene Family
Structural divergence within the PHR family is accompanied by an even greater divergence in expression pattern. As noted above, PHR1 expression varies as a function of culture pH, independent of culture medium composition, temperature, and cell morphology [14]. Expression, as measured by mRNA levels, is greatest in culture medium adjusted to an alkaline pH of 7.5–8.0 and shows a graded decline in parallel with culture pH to undetectable levels at pH 5.0 [14]. Remarkably, expression of PHR2 not only responds to ambient pH, but the response is the precise inverse of PHR1 [16]. Expression is highest at pH 4.0 and is reduced as ambient pH is increased [16]. The pH-conditional expression of these genes is regulated by a zinc-finger containing transcription factor Rim101p, which is proteolytically activated at ambient alkaline pH and induces transcription of PHR1 and represses expression of PHR2 [19,20,21,22,23].
In contrast to PHR1 and PHR2, expression of the other family members is completely independent of ambient pH. PHR3 and PGA5 are constitutive and weakly expressed [17]. Transcript levels of PGA4 are significantly higher and, although pH has no influence, the presence of serum causes a transitory 3- to 4-fold induction [17]. Expression is also increased several fold in a reconstituted human epithelial model and in vivo [17]. These transcriptional differences are directly reflected at the protein level. Phr1p is greatly enhanced in the walls of cells cultured at pH 7, whereas Phr2p is absent, and, conversely, Phr2p is found when cultures are grown at pH 4 [24]. Pga4p is present under both conditions [24].
3. Molecular Features and Localization of Phr Proteins
3.1. Phr Proteins Are C. albicans Representatives of Family GH72
The deduced amino acid sequences of the PHR genes initially provided no clue to function except for a distant relationship with bacterial β-glycosidases [25]. The proteins encoded by PHR1 and PHR2 showed a high degree of amino acid identity (>50%) with Gas1p of S. cerevisiae, the Gel1 protein of the filamentous fungus Aspergillus fumigatus (AfGel1) and Epd1p of C. maltosa (CmEpd1). Gel/Phr/Gas/Epd proteins became the founders of a new group of glycoside hydrolases (GH) classified as family 72 in the database of carbohydrate-active enzymes (CAZy) and whose activity will be described in Section 3.1.1. Family GH72 grew rapidly thanks to the progress of several sequencing projects of yeast and fungal genomes, and currently counts hundreds of members distributed in many species, from Neurospora crassa to Aspergillus sps., from Candida sps. to plant symbiotic fungi such as Tuber melanosporum or phytopathogens such as Magnaporthe oryzae [26]. These enzymes are unique to the Kingdom Fungi. For simplicity throughout this review, GH72 proteins will be indicated by a prefix of the genus and species of origin except Phr of C. albicans and Gas of S. cerevisiae.
Interestingly, GH72 enzymes are characterized (i) by a high degree of redundancy within the same species and (ii) by the existence of orthologs across all species so far examined [27]. The most striking examples of redundancy are: the Gel family of A. fumigatus (AfGel1p to AfGel7p), the Gas family of S. cerevisiae (Gas1p to Gas5p) and the Gel family of N. crassa (NcGel1 to NcGel5). A notable exception is the fungal pathogen Cryptococcus neoformans, and in general Basidiomycota, which have only one GH72-encoding gene [27].
The expansion of gene families encoding GH72 enzymes likely reflects a genetic adaptation to different environmental conditions and ecosystems, in addition to functional specialization of duplicated genes. Cell reshaping occurs during the complex morphological transitions a fungus typically undergoes in its life cycle (yeast/hyphal growth, spore or conidiophore formation) and requires the expression of different paralogs. Well-known examples of a dynamic interplay among paralogs during the fungal life cycle were described for the GAS genes of S. cerevisiae, the gas+ genes of Schizosaccharomyces pombe, and the GELs of Neurospora crassa and of A. fumigatus [28,29,30,31,32]. Gene redundancy occurs not only in microorganisms that underwent whole genome duplication, e.g., S. cerevisiae, but other species as well. The observed co-expression of two or more paralogs during the same stage of the life cycle guarantees a back-up system able to compensate for the loss of one of the paralogs. Taking into consideration the evolutionary cost of keeping duplicated genes in a genome, these observations accentuate the importance of GH72 activity for fungal morphogenesis.
3.1.1. Catalytic Activity of the GH72 Enzymes: AfGel1, Gas1, and Phr1-Phr2 Proteins
In 2000, the Laboratoire des Aspergillus led by Jean Paul Latgé, using laminarin-derived laminarioligosaccharides (β-(1,3)-linked glucose residues) of different length as substrate and HPAEC analysis of the reaction products, demonstrated for the first time that GH72 proteins were catalytically active in processing β-(1,3)-glucan molecules [33]. Recombinant AfGel1p internally cleaved a β-(1,3)-glucan (the donor) and transferred the cleaved product to the non-reducing end of another β-(1,3)-glucan (the acceptor) forming a β-(1,3)-glycosidic linkage at the transfer site. As the reaction proceeded, the elongated product itself became a donor and the reaction culminated in the formation of insoluble glucan [33]. Phr1p, Phr2p, and Gas1p proved to be endowed with the same in vitro activity as AfGel1p [33]. The minimum length of the donor for Phr1p was nine glucose residues and 10 for Phr2p, Gas1p, and AfGel1p. Given this type of endo/transglycosylase activity, these enzymes were collectively called β-(1,3)-glucanosyltransferases or β-(1,3)-glucan elongases [34].
GH72 enzymes are specific for β-(1,3)-glucan both as donor and as acceptor [33,35,36]. By using a laminarioligosaccharide that can only act as an acceptor (rG7) and varying the concentration of the donor substrate (rG11), the Km of the donor site of AfGel1p was measured as 5.3 mM [37]. When substrate concentrations are low, the enzyme will utilize the hydroxyl group of water as the acceptor, resulting in hydrolysis of the glucan. In this respect, GH72 enzymes differ from the “pure” transglycosidases, e.g., plant cell wall xyloglucanosyltransferases (XET), which do not exhibit hydrolytic activity and are also catalytically more efficient [38].
The reaction mechanism of GH72 enzymes is general acid–base catalysis with a proton donor and a nucleophile group. The mechanism of double displacement occurs in two steps involving the general acid/base catalyzed formation and subsequent hydrolysis (or transgycosylation) of a covalent glycosyl-enzyme intermediate (Figure 1). The –COOH group of an acid/base residue (a glutamic acid) and a dissociated carboxylate, –COO−, of another glutamic acid are involved. The reaction can result in an hydrolytic event if the acceptor is H2O or in transglycosylation if the acceptor is the non-reducing end of a laminarioligosaccharide or β-(1,3)-glucan (Figure 1). The two catalytic glutamic acid residues are in different microenvironments: the acid/base group is in a non-polar region in which the dissociation of the side chain is less favored, whereas the nucleophile group is in a polar environment that promotes dissociation.
3.1.2. Domain Composition and Structure of the Phr Family of Proteins
A diagram of the domain structure of the Phr family of proteins is depicted in Figure 2. Phr proteins share a GH72 domain including a linker region but are dissimilar in their C-terminal region. Three are members of the subfamily GH72+ (Phr1p, Phr2p, and Pga5p) and two of the subfamily GH72− (Phr3p and Pga4p).
This division into two subfamilies derives from multiple alignments of the available GH72 sequences [27,40]. All GH72 enzymes share the GH72 domain and a linker region. The sub-family GH72+ contains in addition a carbohydrate binding module of family 43 (CBM43) also annotated as X8 in the Pfam database (Pfam 31.0, EMBL-European Bioinformatics Institute, Wellcome Genome Campus, Hinxton, Cambridgeshire, UK). The GH72 members that lack CBM43/X8 constitute the subfamily GH72− and have a low-complexity tract in the C-terminal region.
In spite of this distinction, GH72+ and GH72− enzymes exhibit the same in vitro transglycosidase activity, which implies a more subtle role of CBM43 in the catalytic activity of GH72+ enzymes (see below Section 3.1.4). In some members, a serine/threonine-rich region (Ser/Thr-box) that is highly O-mannosylated and dispensable for the activity, is present in the C-terminal tract just before the C-terminal GPI-attachment signal [41].
The GH72 domain contains two conserved glutamate residues that are essential for catalysis and three conserved tyrosine residues. Strikingly, GH72 enzymes also share a conserved pattern of regularly spaced cysteine residues that form intramolecular disulfide bonds [42]. In the GH72+ subfamily, two clusters of disulfide bonds are present: Cluster I comprises three disulfide bonds and involves the GH72 domain and a conserved cysteine residue of the linker region, while Cluster II comprises four disulfide bonds connecting cysteine residues located in CBM43/X8. Cluster II is absent in GH72− enzymes. Extensive mutagenesis analysis and unsuccessful attempts at expressing CBM43 domain in isolation, suggested that the GH72 domain and CBM43 constitute an integrated structural and functional unit [42]. In support to this notion, the three dimensional structure of a GH72+ protein (Gas2p) highlighted the existence of a large interaction surface between the GH72 domain and CBM43 [43]. The GH72 domain has a (β/α)8 barrel structure and CBM43 is an α-helical domain. The conserved cysteine residue of the linker is sulfide-bonded to a cysteine residue of the GH72 domain, thus bringing GH72 and CBM43 domains into physical proximity. It has been shown for Gas1p that this disulfide bond is critical for the folding of the entire protein [42]. The two catalytic glutamate residues of the GH72 domain are separated by approximately 100 amino acids in the primary structure but in three-dimensional space are separated by only 5 Å, compatible with catalysis [43]. Based on crystallographic studies, a mechanism of base occlusion was proposed through which the leaving chain does not diffuse away but rather slides along the binding sites that are progressively occupied by the acceptor, limiting the access of water to the active site [43].
Replacement of either active site glutamate residue with a glutamine causes the loss of enzyme activity without affecting protein folding. Consistently, the E169Q or an E270Q mutant of Phr1p fail to complement the corresponding gene inactivation [25]. E159 and E260 are the catalytic residues in Phr2p.
Given the high homology between Phr1-Phr2p and Gas2p (see Section 3.1.3, Figure 4), a molecular model of both proteins was built (see Section 3.1.3). Superimposition of the structures indicated that the stereochemistry of the active site is identical for Phr1p and Phr2p. In Figure 3, the predicted structure of the active site of Phr2p is shown. The catalytic residues are separated by 5 Å and a triad of tyrosines (Y90-Y229-Y292) and N227 surround these residues. Residue Y229, being less than 2 Å from E260, is predicted to greatly influence the dissociation state of its carboxylate. This tyrosine triad and N227 likely create a network of H-bonds and a movement of protons responsible for the protonated state of the catalytic glutamic acid residues. Indeed, replacement of individual tyrosine residues with Phe or Gln in equivalent positions of Gel2p has been shown to affect the transglycosylation reaction [43]. Other residues of Phr2p, R88, R125, and N158 line the active site and are predicted to be involved in substrate binding.
3.1.3. Active, Inactive, and Anomalous Members of the Phr Family of Proteins
Given the medical importance of C. albicans as a fungal pathogen, the redundancy of Phr proteins was explored more deeply to gain information on the activity of the proteins. First, the amino acid sequences of the GH72-Linker domain and of the C-terminal region were compared to the respective sequences of the Gas family of proteins (Figure 4) [36]. As shown in Figure 4, beside the high degree of amino acid identity of Phr1p and Phr2p to Gas1 protein, Pga4p showed a high degree of identity to Gas5p (60%) in the first 360 amino acids (GH72 domain+ linker). Gas1p and Gas5p are expressed during vegetative growth in yeast and are required for cell wall formation with a predominant expression and function of Gas1p over Gas5p [44]. Interestingly, Phr3p and Pga5p are more closely related to the sporulation-specific Gas1-homologs Gas4p and Gas2p, respectively. The Gas2–Gas4 protein pair is specialized in spore-wall formation in yeast [45]. However, C. albicans does not undergo the process of meiosis and sporulation. This suggests that Phr3p and Pga5p, if active, have probably been directed to other processes. However, Phr3p and Pga5p contain various anomalies. The CDS of PHR3 has an unbiased use of codons, an index of low expression and Phr3p lacks an apparent GPI attachment signal. Pga5p has (i) a stretch of 10 consecutive Asn residues (polyAsn, encoded by the cognate codon AAC) and (ii) a stretch of 12 glutamic acid residues (encoded by 10 GAA and two GAG codons). In addition, as mentioned above, PHR3 and PGA5 transcripts are of low abundance.
Furthermore, no phenotypic defects have been associated with deletion of PHR3 or PGA5 (see further Section 4) [27]. Thus, PHR3 and PGA5 appear to be genetically inactive member of the PHR family.
With reference to phylogenetic relationships, PHR1 and PHR2 are orthologs of GAS1 (clade G1), PGA4 is the ortholog of GAS5 (clade alpha), PGA5 is the ortholog of GAS2 (clade G2) and PHR3 is an ortholog of GAS4 (clade gamma) according to a recent evolutionary analysis [27].
Therefore, recent studies on the catalytic activity of the Phr proteins concentrated on Phr1p, Phr2p and Pga4p. By use of a new fluorescent enzyme microassay, designed in collaboration with Vladimir Farkas of the Laboratory of Glycomics, the basic catalytic properties of recombinant Phr1p, Phr2p, and Pga4p were investigated [36]. Surprisingly, no activity was detected for recombinant Pga4p. Therefore, out of the five members of the Phr family of C. albicans, so far only two genes appear to encode active GH72 isoenzymes (PHR1 and PHR2) and one gene (PGA4) likely encodes a structural mannoprotein. This is different from the situation in S. cerevisiae where, out of five Gas proteins, four are catalytically active and only one does not show activity (Gas3p) [40]. Table 1 summarizes the results of studies on the catalytic properties of the Phr1 and Phr2 proteins.
The retention of only two enzymatically active GH72 isoenzymes in C. albicans may be linked to the commensalism of this opportunistic fungal pathogen that has the human body as a unique host and also to the lack of complex differentiation pathways such as meiosis and sporulation. The human fungal pathogen A. fumigatus is a saprophyte exposed to the natural environment (soil) and spores must be inhaled to produce invasive infections in immunocompromised patients. Moreover, the A. fumigatus life cycle is very complex. This may justify the presence of seven GH72 isoenzymes in this human fungal pathogen.
Interestingly, the gene family encoding catalytic subunits of β-(1,3)-glucan synthase (FKS/GLS) show a similar situation. In S. cerevisiae, three FKS genes are functional. The genome of C. albicans contains three FKS homologs but only one of them, GSC1, is essential, whereas GLS1 encodes a truncated protein and GSL2 expression, if any, is very low. This proved useful in finding inhibitors of β-(1,3)-glucan synthesis that kill C. albicans.
3.1.4. β-(1,3)-glucan Elongation and Branching: Cooperation between Family GH72+ (Gas/Phr/AfGel) and Family GH17 (Bgl/Bgt) Transglycosylases
Although it has long been known that the glucan network is made of branched β-(1,3)-glucan fibers and that lateral branches create anchoring sites for other cell wall components (β-(1,6)-glucan, chitin and mannoproteins), the identification of the branching activities has been a challenge for fungal molecular biologists for many years. A transglycosylase capable of adding β-(1,6)-branches to β-(1,3)-glucan, Bgl2p, was identified in S. cerevisiae, C. albicans, and A. fumigatus (AfBgt1p and AfBgt2p), but the role of these proteins seemed negligible since the corresponding deletion strains did not manifest any remarkable phenotypic trait except higher sensitivity to the chitin synthesis inhibitor Nikkomycin Z and a lower kinetics of biofilm formation of the C. albicans bgl2 null mutant [46,47,48].
Yeast Bgl2p is a 35-kDa glycoprotein of the GH17 family, localized to the cell wall and also secreted in the medium. In vitro Bgl2p/AfBgt1p catalyze the transfer of a disaccharide unit (laminariobiose) from the reducing end of a laminarioligosaccharide (≥ 5 glucose units) to the non-reducing end of an acceptor laminarioligosaccharide (≥4 glucose residues) creating a β-(1,6)-glycosidic linkage at the transfer site [49]. The resulting oligomer is a “kinked” linear β-(1,3)-glucan. Interestingly, another Bgl2-homologous protein identified in A. fumigatus, AfBgt2p, is able to catalyze the transfer of the glucan chain to an internal glucose. Since the internal glucose is linked to the neighboring glucose units by its C-1 and C-3 and to the C-1 of the transferred glucose/glucan chain by its C-6, this activity creates a so-called “branching point”.
Recently, a relevant advance in the field was the finding that (i) GH72 enzymes are endowed with not only elongation, but also branching activity, (ii) GH72 enzymes cooperate with the Bgl/Bgt GH17 proteins in creating the branched core glucan and (iii) the β-(1,3)-glucan of a gas1Δbgl2Δ mutant is devoid of β-(1,6)-branching and exhibits a synthetic sick phenotype [50].
The amount of branching in the cell wall is comparable in yeast, A. fumigatus and C. albicans. The proposed model predicts that Gas1p forms branches either by introducing a β-(1,6)-linkage on an internal glucose unit of a self-created β-(1,3)-linked long glucan or alternatively by catalyzing the transfer of a β-(1,3)-glucan to a C-3 on a kinked linear β-(1,3)-glucan created by Bgl2p. Thus, Gas1p and Bgl2p cooperate in elongation and branching of the cell wall glucan. Bgl2p preferentially uses short chains (≥5 glucose residues) whereas the branching activity of Gas1p first requires the formation of long glucans (≥11). The contribution of Gas1p and Bgl2p in the creation of β-(1,6)-linkages on β-(1,3)-glucan was determined by analyzing the percentage of branching in the deletion mutants. While the bgl2Δ mutant showed a 15% decrease in branching and the gas1Δ mutant exhibited a 70% decrease, the double gas1Δ bgl2Δ mutant had no branching and manifested a synthetic sick phenotype characterized by a slow growth rate, round cell shape, random budding pattern and increased sensitivity to cell wall perturbing agents compared to the wild type or the single bgl2Δ or gas1Δ [50]. Thus, GH72 enzymes represent the major glucan elongation and branching activity and cooperate with GH17 enzymes. No bgl2Δ phr1Δ or bgl2Δ phr2Δ double deletions have been described in C. albicans.
Interestingly only GH72+ proteins (Gas1p, Gas2p, and AfGel4p) proved to be endowed with elongation and branching activity, whereas GH72− enzymes (Gas5p, AfGel1p, and AfGel2p) exhibited the elongation but not the branching activity. This result suggests that CBM43 is essential for proper positioning of the substrate required for the branching activity. This notion provides the first functional distinction between the two sub-families.
In conclusion, the GH72 enzymes work in concert with the plasma membrane β-(1,3)-glucan synthase complex, which synthesizes short chains using Uridine diphosphate glucose (UDP-glucose) as a substrate and extrudes the polymer into the periplasmic space. GH72 enzymes then process these short β-(1,3)-glucan chains in the extracellular space. Given their in vitro activity, the current proposed model predicts a role of GH72 enzymes in creating long glucan chains and in elongating branching points [50]. Figure 5 summarizes the main steps of cell wall assembly.
The in vivo impact of GH72 enzymes on cell wall β-(1,3)-glucan was demonstrated by pulse-chase experiments in C. albicans cells [25] (see Section 4). Another example of the impact of the elongation activity on cell wall assembly in vivo came in 2012. Cabib showed that the lack of Gas1p converts yeast HMW β-(1,3)-glucan into a polydisperse glucan [51]. Studies of A. fumigatus have also demonstrated the in vivo effects [50].
3.2. Where and When Do Phr Proteins Play Their Biological Role in C. albicans Cells?
An important piece in the puzzle of defining the biological role of a protein is its localization. With respect to Phr1, Phr2, and Pga4 proteins, modification by GPI attachment affects their localization. This modification occurs in the ER and, together with N-glycosylation, promotes the transport of proteins through the secretory pathway to cell surface. The GPI moiety is a means of anchoring extracellular proteins to the outer layer of the plasma membrane and also confers lateral mobility to the proteins. In addition, in the majority of Ascomycetes, the glycan moiety of the glycolipid can be processed at the cell surface by a putative transglycosidase that transfers the lipid-less portion of GPI (GPI-remnant) to the cell wall polysaccharide network and consequently covalently cross-links the protein to the cell wall. In this way, a plasma membrane GPI protein is converted into a cell wall protein (CWP), termed GPI-CWP. In C. albicans, the majority of the CWPs are GPI-CWPs and play a crucial role in fitness and virulence [53]. In general, only a minor fraction of GPI proteins are transformed into GPI-CWPs, whereas the majority remain attached to the plasma membrane. However, the proportion of these fractions varies from protein to protein.
Thus, it is not surprising that MS analyses of the C. albicans cell wall proteome identified Phr1p, Phr2p, and Pga4p as GPI-CWPs [24,54,55]. Moreover, in a proteomic study of the cell surface, Phr1p and Pga4p were detected in the material shaved from intact C. albicans cells, indicating their surface location [56]. It is still debated whether the GPI proteins endowed with enzymatic activity retain their activity when they are covalently linked to the cell wall. The GPI remnant is connected to a β-(1,6)-glucan chain and this in turn is linked to β-(1,3)-glucan [53]. The short β-(1,6)-glucan may provide a flexible arm that lets the enzyme operate within a substantial radius of action and at more superficial sites of the cell wall. However, this attractive hypothesis awaits experimental demonstration.
The localization of Phr1p was investigated in detail using an internal GFP tag [57]. In vegetative growth, Phr1p-GFP localized to the presumptive bud site and to the bud periphery, sites where the cell wall is more plastic and new material is incorporated during the polarized growth phase of the cell cycle (Figure 6). During the isotropic growth phase and in particular in cells at cytokinesis, Phr1p-GFP is concentrated at the septum, where it forms a brightly fluorescent band (Figure 6), suggesting a role in the assembly of the secondary septa.
When C. albicans is induced to develop hyphae, Phr1p-GFP localizes to the site of germ tube emergence, to the tip of the germ tube, and to the septum (Figure 7). Later, it localizes to the hyphal apex and also distributes along the lateral walls of the hyphae.
Localization at the site of germ tube emergence, the germ tube tip and the hypha apex, and lateral hyphal walls is dependent on the actin cytoskeleton, whereas microtubules control the localization of Phr1p at the septum [57]
Given that the Phr proteins are located in both the cell membrane and the cell wall, covalent cross-linking to the cell wall might be expected to sequester the protein from membrane turnover. We have previously shown that is possible to specifically localize a GPI-CWP under conditions that induce the degradation of the plasma membrane form [58]. Preliminary experiments indicate that during a shift from pH 7.5 to 4.5, a condition that switches off the expression of PHR1, Phr1p-GFP totally disappears in a couple of hours from the cell contour (Popolo and Degani, unpublished data). This suggests that all of the Phr1p-GFP is subjected to protein turnover and no covalently cross-linked cell wall forms are detectable. This differs from Gas1p-GFP in yeast, which was shown to be cross-linked to the wall of the bud scars, where brightly fluorescent rings overlapped with the chitin rings on the cell surface [29].
Finally, the recruitment of Phr1p-GFP to hypha-damaged sites caused by neutrophil attack in vitro led to the proposal of a potential repair role of Phr1p [59]. This suggests that the synthesis of a new wall induces a lateral mobility of pre-existing Phr1p molecules to the damaged site or that more molecules are recycled by endocytosis and trafficking to the plasma membrane upon injury of subapical hyphal compartments.
4. PHR Family in Morphogenesis and Virulence
PHR1 and PHR2 are the most thoroughly studied members of the PHR family and exhibit the more overt mutant phenotypes. Cell morphology is dramatically affected in phr1 and phr2 deletion mutants [14,16]. In accord with their transcriptional patterns, the morphological deficits are pH-conditional. A phr1 null mutant grows with a normal budding pattern and typical ellipsoidal yeast shape in acidic culture conditions. At pH 8, 37° C, conditions that promote germ tube formation, the mutant fails to form germ tubes and instead forms rounded outgrowths with extremely wide septa [14]. At 25° C, which promotes growth as yeast, the initial daughter cells of the mutant have a flattened appearance, more wide than long, with wide bud necks [14]. After several generations at alkaline pH, at either temperature, the mutant forms aggregates of enlarged, rounded cells with wide bud necks [14], a phenotype similar to that of S. cerevisiae Gas1 mutants [60]. Mutants lacking PHR2 show similar morphological abnormalities, but the defects appear under acidic culture conditions [16]. One notable difference between PHR1 and PHR2 mutants is that phr1 null mutants continue to grow at alkaline pH, albeit with a reduced growth rate, but phr2 deletion mutants cease growth within two generations at acidic pH [16]. The essential nature of PHR2 under acidic growth conditions was also observed in tetracycline-conditional expression mutants of PHR2 [61]. Despite this difference, forced expression of PHR1 complements the morphological and growth defects of a phr2 mutant and vice versa, demonstrating that they encode analogous activities [16].
The morphological changes in the mutants are accompanied by biochemical changes in the cell wall. When cultured at alkaline pH, the cell wall glucan content of a phr1 null mutant is either unaltered [25,62] or reduced by about 25% [63], depending perhaps on the method of analysis. However, the alkali-soluble fraction is increased [25,62], suggesting a defect in proper cross-linking of nascent β-(1,3)-glucans and consistent with the in vitro enzymatic activities. At the same time, there is a 5- to 6-fold increase in the amount of zymolase-insoluble glucan, β-(1,6)-glucan, cross-linked to chitin, and a similar increase in chitin content [25,62], further indicating a deficiency in β-(1,3)-glucan processing. In line with these changes, a phr1 mutant is more sensitive to calcofluor white, nikkomycin Z, and SDS, but more resistant to caspofungin [62,63]. Analysis of the cell wall of phr2 mutants is more limited, but a similar accumulation of chitin-linked β-(1,6)-glucan occurs at the restrictive pH [25]. These biochemical changes manifest as ultrastructural alterations in the cell wall of both yeast and hyphae [63,64].
Given the impact of PHR1 and PHR2 on the cell wall and the importance of this structure in mediating host–pathogen interactions, loss of PHR1 or PHR2 compromises virulence of C. albicans. Moreover, the differential expression of these two genes results in niche-specific attenuation of virulence. PHR1 null mutants are avirulent in a mouse model of systemic infection employing either BALB/c or CD2F1 mice [65,66]. Although initial colonization of brain, liver, and kidney tissues is comparable to wild-type cells, subsequent proliferation of the mutant is greatly reduced and histological sections demonstrate a lack of hyphae and the presence of enlarged rounded cells similar to those seen in vitro [65,66]. However, in a rat model of vaginal candidiasis, a phr1 null mutant is fully virulent [65]. In contrast, deletion of PHR2 imparts the inverse virulence phenotypes. The phr2 null mutant is fully virulent in the systemic model, but is strongly attenuated in the vaginitis model [65].
Results of the systemic model correlate well with the pH-dependent expression pattern of these genes, given the slightly alkaline pH (7.3) of blood and tissues and the alkaline-induced expression of PHR1. Indeed, expression of PHR1 has been demonstrated in vitro in blood and in vivo in various tissues and infection models including samples from patients with oropharyngeal candidiasis [67] and a mouse model of oropharyngeal candidiasis [68], during intraperitoneal infection and liver invasion [69], in infected rabbit kidneys [70], in the cecum during intestinal colonization of mice [71], and in a zebrafish infection model [72]. Results of the vaginitis model are less readily explained as the reported pH of the rat vagina averages around 6.95 [73]. Acidification of the vaginal environment during infection might account for the PHR2 requirement or other environmental variables of the vagina might influence PHR2 expression.
The virulence attributes impacted by PHR1 and PHR2 are not completely defined, but include adhesion, immune system interactions, and biofilm formation. Cells lacking PHR1 are unable to penetrate or invade reconstituted human epithelia, which correlates with the mutant’s inability to form hyphae and a gross reduction in adherence to epithelial cells and polystyrene [74]. The adherence defect may reflect altered expression of adhesion proteins or failure to properly incorporate them into the cell wall. It is known that the architecture of cell surface mannoproteins is disrupted in phr2 mutants as evidenced by the aberrant exposure of the glucan layer in these cells [75]. Exposure of the glucan layer may contribute to the loss of virulence of PHR null mutants as the mannan layer appears to shield the cell’s glucan layer from immune detection via Dectin-1 and the ensuing inflammatory response [75,76].
The formation of biofilms, a matrix-embedded community of cells, plays a significant role in the biology and pathogenicity of C. albicans [77]. An important component of the biofilm matrix is β-(1,3)-glucan [78]. Expression of PHR1 is increased in biofilm versus planktonic cells [79,80] and cells lacking PHR1 produce nearly 10-fold less matrix glucan [81]. In contrast, another study showed that overexpression of PHR2, but not PHR1, enhanced biofilm occupancy, but not adherence or biofilm mass [82]. These differences likely reflect variations in media composition and pH used for biofilm formation but nonetheless implicate these enzymes in the process. A further indicator is the reduced expression of PHR1 when biofilm formation is blocked by mutation of SUN41 [83].
Another biologically important role of biofilms is their implication in mating. C. albicans cells are typically heterozygous a/α at the mating-type locus. Cells that become homozygous a/a or α/α can undergo a reversible switch between white-phase and opaque-phase cells, which differ in size, morphology, and virulence properties [84]. Opaque cells are competent for mating and α-factor, the pheromone secreted by α/α cells, promotes biofilm formation by white cells in the population [85]. The pheromone response entails upregulation of a number of genes encoding cell wall proteins including PHR1 and PHR2, both of which contain a white pheromone-regulated element (WPRE) within their promoter regions [85]. Furthermore, the switching process itself, from white to opaque, is sensitive to pH [86]. Deletion of PHR1 did not prevent the pH effect, but PHR2 was required to maintain cells in the opaque phase at acidic pH [86].
In contrast to PHR1 and PHR2, few functional studies have been conducted for PHR3, PGA4, or PGA5. Mutants lacking PHR3, PGA4, or PGA5 show no significant alterations in growth, morphology, or virulence [17,87]. Nor do the mutations alter sensitivity to cell wall perturbing agents such as calcofluor white, Congo red, or SDS [17]. Deletion of PGA4 does, however, appear to influence cell wall structure as the mutant has enhanced resistance to caspofungin and anidulofungin, antifungals that inhibit glucan synthase [63]. Furthermore, the mutant has a slightly reduced content of β-(1,3)-glucan in the cell wall and is more sensitive to osmotic stress when cultured with glucose vs. lactose as the carbon source [63,88,89]. Expression of PGA4 is downregulated in ACE2 mutants, which are defective in filamentation under hypoxic conditions [90]. The only phenotype reported for a PHR3 mutant is a reduction in agar invasion ability in a heterozygous transposon insertion mutant [91]. No phenotypes have been associated with mutation of PGA5.
5. The Adaptive Response to Cell Wall Stress Induced by the Lack of Phr1p
5.1. The Response to Cell Wall Stress
Among the stress conditions encountered by C. albicans, immune insults or drug-induced damage can harm cell wall integrity. In general, the response to stress involves a sensing apparatus that activates signaling pathways and conveys/amplifies the input signal. The cellular changes range from gene expression to chromatin modifications, from regulation of protein activity by post-translational modifications to degradation of selected proteins. The combination of these responses induces an adaptation of the cell to the new condition and the consequent downregulation of the signaling pathways.
The response to cell wall stress conditions has been intensely studied both in S. cerevisiae and in C. albicans using different “stressors” such as mutations in cell-wall-related genes, inhibitors of β-(1,3)-glucan synthesis (echinocandin, caspofungin, and mycofungin), cell wall perturbing agents [Calcofluor, a dye that binds nascent chitin chains or Congo red, a compound that binds β-(1,3)-glucan] or treatments with β-(1,3)-glucanases that hydrolyze the cell wall glucan (e.g., Zymolyase). Most of these stressors can be used either in acute or chronic administration. Other widely used stressors are SDS or temperature, although they act indirectly. According to the type of stressor, the activation of signaling pathways can be different and vary in kinetics. In this section, we will limit discussion to the cell wall stress generated by alterations of β-(1,3)-glucan synthesis or assembly (absence of Phr1p). Figure 8 recapitulates the signaling pathways of cell wall stress from the surface to the effectors.
Loss of the mechanical resistance of the cell wall is a relevant stress signal. The associated stretching or warping of the plasma membrane alters the intracellular concentration of Ca++, a crucial second messenger in eukaryotes. This occurs through the action of mechanosensitive-membrane channels (Figure 8) or release of Ca++ from intracellular stores such as that of the ER, an exquisite sensor of cell stress in eukaryotes, or vacuoles [93] (not shown in Figure 8). Specific sensors have been identified as belonging to the Wscp/Mid/Mtl families of type I transmembrane glycoproteins. These proteins have an integrin-like structural organization and detect alterations in chemical/physical properties of the cell wall through their ectodomain. Similar to Wsc proteins, mucin-like proteins or other highly glycosylated surface proteins also contribute to monitoring the status of the cell wall.
The cell wall stress response appears to be common, at least in part, to other yeast and human pathogenic fungi and involves evolutionarily conserved modules of Mitogen-Activated Protein (MAP) kinases comprised of a MAP kinase kinase kinase (MKKK) and a MAP kinase kinase (MKK) that catalyzes the dual phosphorylation of a MAPK, which in turn phosphorylates crucial substrates [94]. In C. albicans, the key MAPK sensitive to cell wall stress is Mkc1p, a homolog of Mpk1p/Slt2p of S. cerevisiae. Rho1p, a small GTPase, activates Protein kinase C (Pkc1p), an evolutionarily conserved Ser/Thr protein kinase that is essential for preventing cell lysis in yeast and regulates Fks1p, the catalytic subunit of glucan synthase.
In C. albicans, the PKC1-Mkc1p pathway, also called the PKC cell wall integrity (CWI) pathway, is transiently activated by hypo-osmotic shock, cell wall stress, oxidative stress, and physical contact [95,96]. In cell wall stress, the transcriptional response effected by Mkc1p is dominated by the activation of the transcription factor (TF) Rlm1p. In addition, the MAP kinases Cek1p, required for filamentation, and Hog1p, typically activated by high osmolarity, oxidative stress, and clamydospore formation, are both triggered [96,97]. Finally, through binding of calcium to calmodulin, activation of the calcineurin/Crz1p pathway leads to dephosphorylation of a latent TF, Crz1p, which translocates from cytoplasm to the nucleus and activates transcription [98]. Recent work in yeast revealed the involvement of the cAMP-dependent protein kinase pathway (cAMP-PKA), typically implicated in nutritional signaling in yeast and essential for viability. Loss of Gas1p function or treatment with caspofungin induces the CWI pathway but represses the cAMP-PKA pathway [99,100]. Inhibition of PKA signaling induces a decrease in telomeric silencing and an increase in rDNA silencing, both changes mediated by Sir2p, a NAD+-dependent histone deacetylase and a subunit of the rDNA silencing complex RENT [100]. Whereas it is unquestionable that the lack of Gas1p or treatments with caspofungin can cause important changes in chromatin silencing [100,101,102], it seems difficult to reconcile the involvement of nuclear Gas1p molecules in the direct control of Sir2p-mediated transcriptional silencing, as was proposed by some authors [103]. Several lines of evidence argue against this: first, Gas1p was never detected in the nucleus using internally tagged GFP fusions versus C-terminal fusions at the GPI-anchor signal, second, the lack of biochemical evidence that Gas1p can transfer a β-(1,3)-glucan to a protein (Sir2p) and third, the chance of fortuitous in vitro interaction between Gas1p domains expressed in bacteria, an unsuitable host for folding of GH72 enzymes, and Sir2p.
Repression of the cAMP-PKA pathway also triggers the accumulation of the TFs Msn2p/Msn4p in the nucleus. In yeast, Msn2p/Msn4p bind the STRE element that is present in the promoters of a set of genes regulated by cell wall stress [104]. A recent link with transcriptional silencing was evidenced by the presence of a STRE element in PNC1, encoding a nicotinaminidase, which provides a connection between the level of nicotinamide and inhibition of Sir2p [100]. Regarding the cAMP-PKA pathway in C. albicans, it is essential for the dimorphic transition, but not for growth. No data are available on the potential repression of the cAMP-PKA pathway in response to cell wall stress in C. albicans. In addition to these pathways, studies of cell wall stress induced by caspofungin treatment recently revealed that phosphatidylinositol(4,5)-bisphosphate and the septin ring also play a role in cell wall integrity [105].
5.2. The Adaptive Response in C. albicans Cells Lacking Phr1p: Protecting Cell Integrity by Preventing Cell Polarization
Our interest in GH72 enzymes motivated a study on the cell wall stress generated by the lack of this activity in C. albicans. The pH-conditional phenotype of phr1∆ cells offered the advantage of simply switching on cell wall stress by a pH up-shift. As described in Section 4, the round cell shape and the widening of the mother–daughter neck of phr1∆ cells reflect the severe weakening of the cell wall that does not efficiently counteract the expanding force caused by the high intracellular turgor pressure, similarly to the condition of hypo-osmotic shock. As expected from the well-known phenomenon of Chs3p-mediated hyper-accumulation of chitin in response to cell wall stress, a mutant deficient in PHR1 and CHS3 showed abnormal enlargement and extensive lysis when cultured as yeast at alkaline pH at 25 °C (Figure 9). The stretching resistance of chitin fibrils likely creates a mechanical barrier to the expanding force coming from inside [34]. The other chitin synthases had no significant effect on morphology or viability in this condition.
We characterized the genome-wide transcriptional profile of phr1Δ cells during hyphal development because this is the stage of maximum adherence to substrates and capacity for penetration of tissues. After the induction of hyphal growth, in the wild-type cells Mkc1p, Cek1p, and Hog1p were transiently activated, whereas in a phr1∆ mutant these MAP kinases remained persistently hyper-stimulated for at least 7 h, indicative of the severe cell wall stress and defective morphogenesis experienced by phr1∆ cells [106].
Three different time points, 1, 3, and 5 h after induction of hyphal growth, were examined for transcriptome analysis [106]. No transcriptionally modulated cross-compensation by the other PHR paralogs was detected. The functional categories of genes more transcribed in the mutant than in the wild type and those more repressed in the mutant than in the wild type were many. Changes in the “Cell wall” category were the most relevant at all three time points. Genes in the functional category of “RNA and ribosome” maturation were among the most represented repressed genes at 1 h, and this is probably related to the abovementioned increase in silencing of rDNA.
The “Cell wall” category included:
- a set of genes encoding mannoproteins PGA23, orf19.750, RBR1, PGA13, PGA54, RBT4, ECM331 and PGA6, suggesting a qualitative and quantitative change in the pattern of cell wall mannoproteins.
- CHS2 and CHS8 encoding the Class I Chitin synthases, Chs2p and Chs8p, and reported to be subject to Mkc1p and Hog1p-mediated transcriptional increase in the presence of cell wall stress.
- CHS7 encoding the ER-chaperone for Chitin synthase 3 (Chs3p), a post-translational regulated Class IV Chitin synthase.
- CRH11, a homolog of S. cerevisiae CRH1 and a member of C. albicans family GH16 trans-glycosylases together with UTR2/CSF4 and CRH12. GH16 proteins cross-link chitin with glucans by transferring the reducing end of chitin to either the non-reducing end of a β-(1,3)-glucan chain or to a β-(1,3)-glucose side-branch of a β-(1,6)-glucan chain [107] (Figure 5).
The existence of an additional Class I chitin synthase in C. albicans, CHS8, stimulated our interest in the physiological role of chitin synthases in the response to CW stress. This was examined in a set of double mutants lacking PHR1 and one or more of the chitin synthase genes. Here we summarize the most relevant findings. The combined deletion of PHR1 and inactivation of a CHSX (X = CHS2, CHS8 or CHS2, CHS8) gene was examined during filamentous growth. The observed phenotypes depended on ambient pH and the composition and type of medium (liquid or solid). In M199, at the less restrictive pH (pH 7.5) at 37 °C, only phr1∆ chs3∆ cells displayed a worsening of the phr1∆ mutant phenotype with progressive swelling and cell lysis. At the more restrictive pH 8, phr1∆ chs3∆ cells were highly aggregated and, after sonication, appeared arrested at the onset or early germ tube emergence with a majority of lysed cells. Although Chs2p and Chs8p contributed to the chitin increase in the phr1∆ mutant, they were not crucial for viability [106].
On M199 agar-solidified plates the phr1∆ chs3∆ mutant did not geminate at either pH 7.5 or pH 8 and the phenotype was not by-passed by inclusion of 0.8 M sorbitol in the plates. The parental strains were unaffected in their capacity to germinate and formed colonies. On the same medium, at pH 8 only, the phr1∆ chs8∆ mutant also displayed an interesting phenotype. The double mutant germinated but cells did not progress further giving rise to abortive microcolonies visible only under the stereomicroscope. This phenotype was not sorbitol-remediable. No effects of the CHSX gene deletions were visible in the wild-type background at pH 8.
These results indicate that Chs3p and Chs8p have two different execution points in phr1∆ cells, Chs3p acts before Chs8p in protecting the site of germinating phr1∆ cells (Figure 10). Notably, these phenotypes also suggest the existence of a strict coordination among Phr1p, Chs3p, Chs8p, and the polarity machinery required at the onset of germ tube emergence and during elongation.
In order to apply these studies to the identification of news drugs, it is important to sort out those transcriptional responses essential for adaptation. Thus, limiting our discussion to the cell wall components identified by several studies as part of the cell wall response, the coordinated increase of chitin, of mannoproteins and of the enzyme that cross-links them to glucan or chitin (Crh11p) might be crucial for viability. However, in S. cerevisiae the gas1Δ crh1Δ mutant (CRH1 is equivalent to CRH11 of C. albicans) exhibits a phenotype similar to gas1Δ suggesting that CRH1 is not crucial for survival to cell wall stress. No data are available on the phenotype of a combined phr1Δ crh11Δ mutant of C. albicans. On the contrary, the deletion of GAS1 with KRE6, the first gene of the β-(1,6)-glucan synthesis process, is synthetic lethal suggesting that β-(1,6)-glucan is crucial for survival. Chitin and β-(1,6)-glucan synthesis likely compensate for defects in the synthesis/assembly of the core β-(1,3)-glucan mesh.
Among other genes with increased transcription in phr1∆ cells, it is worth mentioning: (i) CPP1, encoding a dual specificity protein phosphatase homologous to Msg5p of S. cerevisiae, which is of interest for its potential role in downregulation of the MAPK pathways, given the promiscuity of action of these protein phosphatases; (ii) SSK2, encoding the only MAPKKK of the Hog1p pathway in C. albicans and (iii) FLC2, in the category of “Transport”, encoding a putative ER membrane transporter that in S. cerevisiae is involved in the release of Ca++ in response to hyposmotic shock [108].
6. Conclusions and Future Outlook
Overall research on the PHR multigene family has highlighted the important role of PHR1 and PHR2 in C. albicans biology and pathogenesis. In a wider vision of human fungal pathogens, GH72 enzymes are essential for virulence and in some instances even for fungus viability. The latter is observed for GEL4 of A. fumigatus [109] and for GAS1-GAS2 of C. glabrata [110], whereas PHR1 and/or PHR2 produce a conditional synthetic sick/lethal phenotype [16,61,111].
These findings encourage further studies of the PHR family both for increasing basic knowledge of C. albicans and also to potentially exploit at Phr1p and Phr2p as molecular targets of new antifungal agents for prophylaxis, therapy of invasive infections, or lock therapy. The armamentarium of antifungal drugs is still limited to three classes: polyenes, azoles, and echinocandin. The advantages of the GH72 enzymes are (i) they have no counterpart in humans; (ii) their extracellular location makes them directly accessible to drugs; (iii) 3D molecular models are available based on the structure of homologous proteins (yeast Gas2p); and (iv) advanced knowledge of the cell wall stress response may provide a means to better monitor drugs’ efficacy or define effective drug target combinations.
The transfer of knowledge from basic biology to therapeutic applications requires an interdisciplinary approach relying on the cooperation of experts in enzymology, glycobiology, structural biology, and bioinformatics. Potential inhibitors would undermine cell wall assembly and in this respect would be the fungal equivalent of penicillin. It is worth mentioning that PHR genes/proteins may find applications in medical mycology for species identification by rapid PCR-based tests [112] and for blood tests, since GAS/GEL proteins are potent fungal antigens that can discriminate patients affected by candidiasis or aspergillosis from healthy individuals and may also have an impact on vaccinations [113,114].
Author Contributions
L.P. and W.F. conceived and designed the structure of the review, wrote and edited the manuscript. G.D. performed the bibliography review, collected the data for the tables, prepared the figures and assisted in drafting the manuscript. C.C. built and analyzed the homology-derived molecular models of Phr1 and Phr2 proteins and prepared the figure on Phr2p. The manuscript has been read and approved by all named authors.
Acknowledgments
We apologize to those whom we were unable to reference due to space limitations. Genny Degani is the recipient of a fellowship financed by the University of Milan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Odds, F.C. Candida and Candidosis: A Review and Bibliography, 2nd ed.; Bailliere Tindal: London, UK, 1988; p. 468. [Google Scholar]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Latge, J.P. The cell wall: A carbohydrate armour for the fungal cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Latge, J.P.; Beauvais, A. Functional duality of the cell wall. Curr. Opin. Microbiol. 2014, 20, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Douglas, C.M.; D’Ippolito, J.A.; Shei, G.J.; Meinz, M.; Onishi, J.; Marrinan, J.A.; Li, W.; Abruzzo, G.K.; Flattery, A.; Bartizal, K.; et al. Identification of the fks1 gene of Candida albicans as the essential target of 1,3-β-d-glucan synthase inhibitors. Antimicrob. Agents Chemother. 1997, 41, 2471–2479. [Google Scholar] [PubMed]
- Orlean, P. Architecture and biosynthesis of the Saccharomyces cerevisiae cell wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed]
- Popolo, L.; Alberghina, L. Identification of a labile protein involved in the g1-to-s transition in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1984, 81, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Popolo, L.; Vai, M.; Alberghina, L. Identification of a glycoprotein involved in cell cycle progression in yeast. J. Biol. Chem. 1986, 261, 3479–3482. [Google Scholar] [PubMed]
- Vai, M.; Popolo, L.; Grandori, R.; Lacana, E.; Alberghina, L. The cell cycle modulated glycoprotein gp115 is one of the major yeast proteins containing glycosylphosphatidylinositol. Biochim. Biophys. Acta 1990, 1038, 277–285. [Google Scholar] [CrossRef]
- Conzelmann, A.; Riezman, H.; Desponds, C.; Bron, C. A major 125-kd membrane glycoprotein of Saccharomyces cerevisiae is attached to the lipid bilayer through an inositol-containing phospholipid. EMBO J. 1988, 7, 2233–2240. [Google Scholar] [PubMed]
- Birse, C.E.; Irwin, M.Y.; Fonzi, W.A.; Sypherd, P.S. Cloning and characterization of ece1, a gene expressed in association with cell elongation of the dimorphic pathogen Candida albicans. Infect. Immun. 1993, 61, 3648–3655. [Google Scholar] [PubMed]
- Saporito-Irwin, S.M.; Birse, C.E.; Sypherd, P.S.; Fonzi, W.A. Phr1, a pH-regulated gene of Candida albicans, is required for morphogenesis. Mol. Cell. Biol. 1995, 15, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Vai, M.; Orlandi, I.; Cavadini, P.; Alberghina, L.; Popolo, L. Candida albicans homologue of ggp1/gas1 gene is functional in Saccharomyces cerevisiae and contains the determinants for glycosylphosphatidylinositol attachment. Yeast 1996, 12, 361–368. [Google Scholar] [CrossRef]
- Mühlschlegel, F.A.; Fonzi, W.A. Phr2 of Candida albicans encodes a functional homolog of the pH-regulated gene phr1 with an inverted pattern of pH-dependent expression. Mol. Cell. Biol. 1997, 17, 5960–5967. [Google Scholar] [CrossRef] [PubMed]
- Eckert, S.E.; Heinz, W.J.; Zakikhany, K.; Thewes, S.; Haynes, K.; Hube, B.; Muhlschlegel, F.A. Pga4, a gas homologue from Candida albicans, is up-regulated early in infection processes. Fungal. Genet. Biol. 2007, 44, 368–377. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.W.; Hellingwerf, K.J.; Klis, F.M. Genome-wide identification of fungal GPI proteins. Yeast 2003, 20, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Ramon, A.M.; Porta, A.; Fonzi, W.A. Effect of environmental ph on morphological development of Candida albicans is mediated via the PACC-related transcription factor encoded by prr2. J. Bacteriol. 1999, 181, 7524–7530. [Google Scholar] [PubMed]
- Ramon, A.M.; Fonzi, W.A. Diverged binding specificity of rim101p, the Candida albicans ortholog of pacc. Eukaryot. Cell 2003, 2, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Porta, A.; Wang, Z.; Ramon, A.M.; Mühlschlegel, F.A.; Fonzi, W.A. Spontaneous second site suppressors of the filamentation defect of prr1d mutants define a critical domain of rim101p. Mol. Genet. Genom. 2001, 266, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.; Wilson, R.B.; Mitchell, A.P. Rim101-dependent and-independent pathways govern ph responses in Candida albicans. Mol. Cell. Biol. 2000, 20, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.U.; Martin, S.J.; Davis, D.A. Evidence for novel ph-dependent regulation of Candida albicans rim101, a direct transcriptional repressor of the cell wall β-glycosidase phr2. Eukaryot. Cell 2006, 5, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Sosinska, G.J.; de Koning, L.J.; de Groot, P.W.; Manders, E.M.; Dekker, H.L.; Hellingwerf, K.J.; de Koster, C.G.; Klis, F.M. Mass spectrometric quantification of the adaptations in the wall proteome of Candida albicans in response to ambient ph. Microbiology 2011, 157, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Fonzi, W.A. Phr1 and phr2 of Candida albicans encode putative glycosidases required for proper cross-linking of β-1,3- and β-1,6-glucans. J. Bacteriol. 1999, 181, 7070–7079. [Google Scholar] [PubMed]
- Samalova, M.; Melida, H.; Vilaplana, F.; Bulone, V.; Soanes, D.M.; Talbot, N.J.; Gurr, S.J. The β-1,3-glucanosyltransferases (gels) affect the structure of the rice blast fungal cell wall during appressorium-mediated plant infection. Cell Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Sillo, F.; Gissi, C.; Chignoli, D.; Ragni, E.; Popolo, L.; Balestrini, R. Expression and phylogenetic analyses of the gel/gas proteins of tuber melanosporum provide insights into the function and evolution of glucan remodeling enzymes in fungi. Fungal. Genet. Biol. 2013, 53, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Free, S.J. Genetic and biochemical characterization of the gh72 family of cell wall transglycosylases in neurospora crassa. Fungal. Genet. Biol. 2017, 101, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Rolli, E.; Ragni, E.; de Medina-Redondo, M.; Arroyo, J.; de Aldana, C.R.; Popolo, L. Expression, stability, and replacement of glucan-remodeling enzymes during developmental transitions in Saccharomyces cerevisiae. Mol. Biol. Cell. 2011, 22, 1585–1598. [Google Scholar] [CrossRef] [PubMed]
- De Medina-Redondo, M.; Arnaiz-Pita, Y.; Clavaud, C.; Fontaine, T.; del Rey, F.; Latge, J.P.; Vazquez de Aldana, C.R. β(1,3)-glucanosyl-transferase activity is essential for cell wall integrity and viability of schizosaccharomyces pombe. PLoS ONE 2010, 5, e14046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Medina-Redondo, M.; Arnaiz-Pita, Y.; Fontaine, T.; Del Rey, F.; Latge, J.P.; Vazquez de Aldana, C.R. The β-1,3-glucanosyltransferase GAS4P is essential for ascospore wall maturation and spore viability in schizosaccharomyces pombe. Mol. Microbiol. 2008, 68, 1283–1299. [Google Scholar] [CrossRef] [PubMed]
- Mouyna, I.; Morelle, W.; Vai, M.; Monod, M.; Lechenne, B.; Fontaine, T.; Beauvais, A.; Sarfati, J.; Prevost, M.C.; Henry, C.; et al. Deletion of gel2 encoding for a β(1–3)glucanosyltransferase affects morphogenesis and virulence in Aspergillus fumigatus. Mol. Microbiol. 2005, 56, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Mouyna, I.; Fontaine, T.; Vai, M.; Monod, M.; Fonzi, W.A.; Diaquin, M.; Popolo, L.; Hartland, R.P.; Latge, J.P. Glycosylphosphatidylinositol-anchored glucanosyltransferases play an active role in the biosynthesis of the fungal cell wall. J. Biol. Chem. 2000, 275, 14882–14889. [Google Scholar] [CrossRef] [PubMed]
- Lesage, G.; Bussey, H. Cell wall assembly in saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Mazan, M.; Ragni, E.; Popolo, L.; Farkas, V. Catalytic properties of the gas family β-(1,3)-glucanosyltransferases active in fungal cell-wall biogenesis as determined by a novel fluorescent assay. Biochem. J. 2011, 438, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kovacova, K.; Degani, G.; Stratilova, E.; Farkas, V.; Popolo, L. Catalytic properties of phr family members of cell wall glucan remodeling enzymes: Implications for the adaptation of Candida albicans to ambient ph. FEMS Yeast Res 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Hartland, R.P.; Fontaine, T.; Debeaupuis, J.P.; Simenel, C.; Delepierre, M.; Latge, J.P. A novel β-(1–3)-glucanosyltransferase from the cell wall of Aspergillus fumigatus. J. Biol. Chem. 1996, 271, 26843–26849. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, H.; Denman, S.E.; Campuzano, I.D.; Ademark, P.; Master, E.R.; Teeri, T.T.; Brumer, H., 3rd. N-linked glycosylation of native and recombinant cauliflower xyloglucan endotransglycosylase 16a. Biochem. J. 2003, 375, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Raich, L.; Borodkin, V.; Fang, W.; Castro-Lopez, J.; van Aalten, D.M.; Hurtado-Guerrero, R.; Rovira, C. A trapped covalent intermediate of a glycoside hydrolase on the pathway to transglycosylation. Insights from experiments and quantum mechanics/molecular mechanics simulations. J. Am. Chem. Soc. 2016, 138, 3325–3332. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Fontaine, T.; Gissi, C.; Latge, J.P.; Popolo, L. The gas family of proteins of saccharomyces cerevisiae: Characterization and evolutionary analysis. Yeast 2007, 24, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Gatti, E.; Popolo, L.; Vai, M.; Rota, N.; Alberghina, L. O-linked oligosaccharides in yeast glycosyl phosphatidylinositol-anchored protein gp115 are clustered in a serine-rich region not essential for its function. J. Biol. Chem. 1994, 269, 19695–19700. [Google Scholar] [PubMed]
- Popolo, L.; Ragni, E.; Carotti, C.; Palomares, O.; Aardema, R.; Back, J.W.; Dekker, H.L.; de Koning, L.J.; de Jong, L.; de Koster, C.G. Disulfide bond structure and domain organization of yeast β(1,3)-glucanosyltransferases involved in cell wall biogenesis. J. Biol. Chem. 2008, 283, 18553–18565. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Guerrero, R.; Schuttelkopf, A.W.; Mouyna, I.; Ibrahim, A.F.; Shepherd, S.; Fontaine, T.; Latge, J.P.; van Aalten, D.M. Molecular mechanisms of yeast cell wall glucan remodelling. J. Biol. Chem. 2008, 284, 8461–8469. [Google Scholar] [CrossRef] [PubMed]
- Rolli, E.; Ragni, E.; Rodriguez-Pena, J.M.; Arroyo, J.; Popolo, L. Gas3, a developmentally regulated gene, encodes a highly mannosylated and inactive protein of the gas family of Saccharomyces cerevisiae. Yeast 2010, 27, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Coluccio, A.; Rolli, E.; Rodriguez-Pena, J.M.; Colasante, G.; Arroyo, J.; Neiman, A.M.; Popolo, L. Gas2 and gas4, a pair of developmentally regulated genes required for spore wall assembly in Saccharomyces cerevisiae. Eukaryot. Cell 2007, 6, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Sarthy, A.V.; McGonigal, T.; Coen, M.; Frost, D.J.; Meulbroek, J.A.; Goldman, R.C. Phenotype in Candida albicans of a disruption of the Bgl2 gene encoding a 1,3-β-glucosyltransferase. Microbiology 1997, 143, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, R.; Takada, A.; Iwatani, S.; Oka, C.; Kitamoto, T.; Kajiwara, S. The role of bgl2p in the transition to filamentous cells during biofilm formation by Candida albicans. Mycoses 2017, 60, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Mouyna, I.; Hartland, R.P.; Fontaine, T.; Diaquin, M.; Simenel, C.; Delepierre, M.; Henrissat, B.; Latge, J.P. A 1,3-β-glucanosyltransferase isolated from the cell wall of Aspergillus fumigatus is a homologue of the yeast bgl2p. Microbiology 1998, 144, 3171–3180. [Google Scholar] [CrossRef] [PubMed]
- Goldman, R.C.; Sullivan, P.A.; Zakula, D.; Capobianco, J.O. Kinetics of β-1,3 glucan interaction at the donor and acceptor sites of the fungal glucosyltransferase encoded by the bgl2 gene. Eur. J. Biochem. 1995, 227, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Aimanianda, V.; Simenel, C.; Garnaud, C.; Clavaud, C.; Tada, R.; Barbin, L.; Mouyna, I.; Heddergott, C.; Popolo, L.; Ohya, Y.; et al. The dual activity responsible for the elongation and branching of β-(1,3)-glucan in the fungal cell wall. MBIO 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E.; Blanco, N.; Arroyo, J. Presence of a large β(1–3)glucan linked to chitin at the Saccharomyces cerevisiae mother-bud neck suggests involvement in localized growth control. Eukaryot. Cell 2012, 11, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Ecker, M.; Deutzmann, R.; Lehle, L.; Mrsa, V.; Tanner, W. Pir proteins of Saccharomyces cerevisiae are attached to β-1,3-glucan by a new protein-carbohydrate linkage. J. Biol. Chem. 2006, 281, 11523–11529. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Sosinska, G.J.; de Groot, P.W.; Brul, S. Covalently linked cell wall proteins of Candida albicans and their role in fitness and virulence. FEMS Yeast Res. 2009, 9, 1013–1028. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.W.; de Boer, A.D.; Cunningham, J.; Dekker, H.L.; de Jong, L.; Hellingwerf, K.J.; de Koster, C.; Klis, F.M. Proteomic analysis of Candida albicans cell walls reveals covalently bound carbohydrate-active enzymes and adhesins. Eukaryot. Cell 2004, 3, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.J.; Sorgo, A.G.; Mohammadi, S.; Sosinska, G.J.; de Koster, C.G.; Brul, S.; de Koning, L.J.; Klis, F.M. Surface stress induces a conserved cell wall stress response in the pathogenic fungus Candida albicans. Eukaryot. Cell 2013, 12, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Marin, E.; Parra-Giraldo, C.M.; Hernandez-Haro, C.; Hernaez, M.L.; Nombela, C.; Monteoliva, L.; Gil, C. Candida albicans shaving to profile human serum proteins on hyphal surface. Front. Microbiol. 2015, 6, 1343. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Calderon, J.; Fascio, U.; Sipiczki, M.; Fonzi, W.A.; Popolo, L. Phr1p, a glycosylphosphatidylinsitol-anchored β(1,3)-glucanosyltransferase critical for hyphal wall formation, localizes to the apical growth sites and septa in Candida albicans. Fungal. Genet. Biol. 2011, 48, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Rolli, E.; Ragni, E.; Calderon, J.; Porello, S.; Fascio, U.; Popolo, L. Immobilization of the glycosylphosphatidylinositol-anchored gas1 protein into the chitin ring and septum is required for proper morphogenesis in yeast. Mol. Biol. Cell 2009, 20, 4856–4870. [Google Scholar] [CrossRef] [PubMed]
- Hopke, A.; Nicke, N.; Hidu, E.E.; Degani, G.; Popolo, L.; Wheeler, R.T. Neutrophil attack triggers extracellular trap-dependent Candida cell wall remodeling and altered immune recognition. PLoS Pathog. 2016, 12, e1005644. [Google Scholar] [CrossRef] [PubMed]
- Popolo, L.; Vai, M.; Gatti, E.; Porello, S.; Bonfante, P.; Balestrini, R.; Alberghina, L. Physiological analysis of mutants indicates involvement of the Saccharomyces cerevisiae gpi-anchored protein gp115 in morphogenesis and cell separation. J. Bacteriol. 1993, 175, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.M.; Kauffman, S.J.; Hauser, M.; Huang, L.; Lin, M.; Sillaots, S.; Jiang, B.; Xu, D.; Roemer, T. Pathway analysis of Candida albicans survival and virulence determinants in a murine infection model. Proc. Natl. Acad. Sci. USA 2010, 107, 22044–22049. [Google Scholar] [CrossRef] [PubMed]
- Popolo, L.; Vai, M. Defects in assembly of the extracellular matrix are responsible for altered morphogenesis of a Candida albicans phr1 mutant. J. Bacteriol. 1998, 180, 163–166. [Google Scholar] [PubMed]
- Plaine, A.; Walker, L.; Da Costa, G.; Mora-Montes, H.M.; McKinnon, A.; Gow, N.A.; Gaillardin, C.; Munro, C.A.; Richard, M.L. Functional analysis of Candida albicans gpi-anchored proteins: Roles in cell wall integrity and caspofungin sensitivity. Fungal. Genet. Biol. 2008, 45, 1404–1414. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Piberger, H.; Neupert, C.; Garcia-Cantalejo, J.; Popolo, L.; Arroyo, J.; Aebi, M.; Strahl, S. The genetic interaction network of ccw12, a Saccharomyces cerevisiae gene required for cell wall integrity during budding and formation of mating projections. BMC Genom. 2011, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- De Bernardis, F.; Mühlschlegel, F.A.; Cassone, A.; Fonzi, W.A. The ph of the host niche controls gene expression in and virulence of Candida albicans. Infect. Immun. 1998, 66, 3317–3325. [Google Scholar] [PubMed]
- Ghannoum, M.A.; Spellberg, B.; Saporito-Irwin, S.M.; Fonzi, W.A. Reduced virulence of Candida albicans phr1 mutants. Infect. Immun. 1995, 63, 4528–4530. [Google Scholar] [PubMed]
- Zakikhany, K.; Naglik, J.R.; Schmidt-Westhausen, A.; Holland, G.; Schaller, M.; Hube, B. In vivo transcript profiling of Candida albicans identifies a gene essential for interepithelial dissemination. Cell. Microbiol. 2007, 9, 2938–2954. [Google Scholar] [CrossRef] [PubMed]
- Fanning, S.; Xu, W.; Solis, N.; Woolford, C.A.; Filler, S.G.; Mitchell, A.P. Divergent targets of Candida albicans biofilm regulator bcr1 in vitro and in vivo. Eukaryot. Cell 2012, 11, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Thewes, S.; Kretschmar, M.; Park, H.; Schaller, M.; Filler, S.G.; Hube, B. In vivo and ex vivo comparative transcriptional profiling of invasive and non-invasive Candida albicans isolates identifies genes associated with tissue invasion. Mol. Microbiol. 2007, 63, 1606–1628. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.A.; Maccallum, D.M.; Bertram, G.; Gow, N.A.; Odds, F.C.; Brown, A.J. Genome-wide analysis of Candida albicans gene expression patterns during infection of the mammalian kidney. Fungal. Genet. Biol. 2009, 46, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Rosenbach, A.; Dignard, D.; Pierce, J.V.; Whiteway, M.; Kumamoto, C.A. Adaptations of Candida albicans for growth in the mammalian intestinal tract. Eukaryot. Cell 2010, 9, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Hsu, P.C.; Jen, C.F.; Chen, I.H.; Wang, C.H.; Chan, H.C.; Tsai, P.W.; Tung, K.C.; Wang, C.H.; Lan, C.Y.; et al. Zebrafish as a model host for Candida albicans infection. Infect. Immun. 2010, 78, 2512–2521. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.; Markovetz, A.J.; Galask, R.P. The bacterial flora of the female rat genital tract. Proc. Soc. Exp. Biol. Med. 1976, 151, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Calderon, J.; Zavrel, M.; Ragni, E.; Fonzi, W.A.; Rupp, S.; Popolo, L. Phr1, a ph-regulated gene of Candida albicans encoding a glucan remodeling enzyme, is required for adhesion and invasion. Microbiology 2010, 156, 2484–2494. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.T.; Fink, G.R. A drug-sensitive genetic network masks fungi from the immune system. PLoS Pathog. 2006, 2. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.T.; Kombe, D.; Agarwala, S.D.; Fink, G.R. Dynamic, morphotype-specific Candida albicans β-glucan exposure during infection and drug treatment. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microb. Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Al-Fattani, M.A.; Douglas, L.J. Biofilm matrix of Candida albicans and Candida tropicalis: Chemical composition and role in drug resistance. J. Med. Microbiol. 2006, 55, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Murillo, L.A.; Newport, G.; Lan, C.Y.; Habelitz, S.; Dungan, J.; Agabian, N.M. Genome-wide transcription profiling of the early phase of biofilm formation by Candida albicans. Eukaryot. Cell 2005, 4, 1562–1573. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.E.; Lepak, A.J.; Marchillo, K.; Andes, D.R. Time course global gene expression analysis of an in vivo Candida biofilm. J. Infect. Dis. 2009, 200, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Taff, H.T.; Nett, J.E.; Zarnowski, R.; Ross, K.M.; Sanchez, H.; Cain, M.T.; Hamaker, J.; Mitchell, A.P.; Andes, D.R. A Candida biofilm-induced pathway for matrix glucan delivery: Implications for drug resistance. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Cabral, V.; Znaidi, S.; Walker, L.A.; Martin-Yken, H.; Dague, E.; Legrand, M.; Lee, K.; Chauvel, M.; Firon, A.; Rossignol, T.; et al. Targeted changes of the cell wall proteome influence Candida albicans ability to form single- and multi-strain biofilms. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norice, C.T.; Smith, F.J., Jr.; Solis, N.; Filler, S.G.; Mitchell, A.P. Requirement for Candida albicans sun41 in biofilm formation and virulence. Eukaryot. Cell 2007, 6, 2046–2055. [Google Scholar] [CrossRef] [PubMed]
- Soll, D.R. Why does Candida albicans switch? FEMS Yeast Res. 2009, 9, 973–989. [Google Scholar] [CrossRef] [PubMed]
- Sahni, N.; Yi, S.; Daniels, K.J.; Srikantha, T.; Pujol, C.; Soll, D.R. Genes selectively up-regulated by pheromone in white cells are involved in biofilm formation in Candida albicans. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cao, C.; Jia, W.; Tao, L.; Guan, G.; Huang, G. Ph regulates white-opaque switching and sexual mating in Candida albicans. Eukaryot. Cell 2015, 14, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; French, S.; Kohn, L.A.; Chen, V.; Johnson, A.D. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 2010, 42, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Suwunnakorn, S.; Wakabayashi, H.; Rustchenko, E. Chromosome 5 of human pathogen Candida albicans carries multiple genes for negative control of caspofungin and anidulafungin susceptibility. Antimicrob. Agents Chemother. 2016, 60, 7457–7467. [Google Scholar] [PubMed]
- Ene, I.V.; Adya, A.K.; Wehmeier, S.; Brand, A.C.; MacCallum, D.M.; Gow, N.A.; Brown, A.J. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell. Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulhern, S.M.; Logue, M.E.; Butler, G. Candida albicans transcription factor ace2 regulates metabolism and is required for filamentation in hypoxic conditions. Eukaryot. Cell 2006, 5, 2001–2013. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Fung, E.; Schlecht, U.; Davis, R.W.; Giaever, G.; St Onge, R.P.; Deutschbauer, A.; Nislow, C. Gene annotation and drug target discovery in Candida albicans with a tagged transposon mutant collection. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.A.; Gow, N.A.; Munro, C.A. Fungal echinocandin resistance. Fungal. Genet. Biol. 2010, 47, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, K.W. Acidic calcium stores of Saccharomyces cerevisiae. Cell Calcium 2011, 50, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, K.; Samantaray, S.; Wagener, J. Cell wall integrity signalling in human pathogenic fungi. Cell Microbiol. 2016, 18, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, C.A. A contact-activated kinase signals Candida albicans invasive growth and biofilm development. Proc. Natl. Acad. Sci. USA 2005, 102, 5576–5581. [Google Scholar] [CrossRef] [PubMed]
- De Dios, C.H.; Roman, E.; Monge, R.A.; Pla, J. The role of mapk signal transduction pathways in the response to oxidative stress in the fungal pathogen Candida albicans: Implications in virulence. Curr. Protein Pept. Sci. 2010, 11, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Eisman, B.; Alonso-Monge, R.; Roman, E.; Arana, D.; Nombela, C.; Pla, J. The cek1 and hog1 mitogen-activated protein kinases play complementary roles in cell wall biogenesis and chlamydospore formation in the fungal pathogen Candida albicans. Eukaryot. Cell 2006, 5, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hou, Y.; Liu, W.; Lu, C.; Wang, W.; Sun, S. Components of the calcium-calcineurin signaling pathway in fungal cells and their potential as antifungal targets. Eukaryot. Cell 2015, 14, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.; Bravo, E.; Diez-Muniz, S.; Nombela, C.; Rodriguez-Pena, J.M.; Arroyo, J. A novel connection between the cell wall integrity and the pka pathways regulates cell wall stress response in yeast. Sci. Rep. 2017, 7, 5703. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.W.; Kim, K.; Chang, Y.J.; Kim, B.; Huh, W.K. The β-1,3-glucanosyltransferase gas1 regulates sir2-mediated rdna stability in Saccharomyces cerevisiae. Nucleic Acids Res. 2014, 42, 8486–8499. [Google Scholar] [CrossRef] [PubMed]
- Ai, W.; Bertram, P.G.; Tsang, C.K.; Chan, T.F.; Zheng, X.F. Regulation of subtelomeric silencing during stress response. Mol. Cell. 2002, 10, 1295–1305. [Google Scholar] [CrossRef]
- Sanz, A.B.; Garcia, R.; Rodriguez-Pena, J.M.; Diez-Muniz, S.; Nombela, C.; Peterson, C.L.; Arroyo, J. Chromatin remodeling by the SWI/SNF complex is essential for transcription mediated by the yeast cell wall integrity mapk pathway. Mol. Biol. Cell. 2012, 23, 2805–2817. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.R.; Pillus, L. The glucanosyltransferase gas1 functions in transcriptional silencing. Proc. Natl. Acad. Sci. USA 2009, 106, 11224–11229. [Google Scholar] [CrossRef] [PubMed]
- Lagorce, A.; Hauser, N.C.; Labourdette, D.; Rodriguez, C.; Martin-Yken, H.; Arroyo, J.; Hoheisel, J.D.; Francois, J. Genome-wide analysis of the response to cell wall mutations in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 20345–20357. [Google Scholar] [CrossRef] [PubMed]
- Badrane, H.; Nguyen, M.H.; Clancy, C.J. Highly dynamic and specific phosphatidylinositol 4,5-bisphosphate, septin, and cell wall integrity pathway responses correlate with caspofungin activity against Candida albicans. Antimicrob. Agents Chemother. 2016, 60, 3591–3600. [Google Scholar] [CrossRef] [PubMed]
- Degani, G.; Ragni, E.; Botias, P.; Ravasio, D.; Calderon, J.; Pianezzola, E.; Rodriguez-Pena, J.M.; Vanoni, M.A.; Arroyo, J.; Fonzi, W.A.; et al. Genomic and functional analyses unveil the response to hyphal wall stress in Candida albicans cells lacking β(1,3)-glucan remodeling. BMC Genom. 2016, 17, 482. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.; Farkas, V.; Sanz, A.B.; Cabib, E. Strengthening the fungal cell wall through chitin-glucan cross-links: Effects on morphogenesis and cell integrity. Cell Microbiol. 2016, 18, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Rigamonti, M.; Groppi, S.; Belotti, F.; Ambrosini, R.; Filippi, G.; Martegani, E.; Tisi, R. Hypotonic stress-induced calcium signaling in Saccharomyces cerevisiae involves trp-like transporters on the endoplasmic reticulum membrane. Cell Calcium 2015, 57, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Gastebois, A.; Fontaine, T.; Latge, J.P.; Mouyna, I. Β(1–3)glucanosyltransferase gel4p is essential for Aspergillus fumigatus. Eukaryot. Cell 2010, 9, 1294–1298. [Google Scholar] [CrossRef] [PubMed]
- Weig, M.; Haynes, K.; Rogers, T.R.; Kurzai, O.; Frosch, M.; Muhlschlegel, F.A. A gas-like gene family in the pathogenic fungus Candida glabrata. Microbiology 2001, 147, 2007–2019. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Jiang, B.; Davison, J.; Ketela, T.; Veillette, K.; Breton, A.; Tandia, F.; Linteau, A.; Sillaots, S.; Marta, C.; et al. Large-scale essential gene identification in Candida albicans and applications to antifungal drug discovery. Mol. Microbiol. 2003, 50, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Kurzai, O.; Heinz, W.J.; Sullivan, D.J.; Coleman, D.C.; Frosch, M.; Muhlschlegel, F.A. Rapid pcr test for discriminating between Candida albicans and Candida dubliniensis isolates using primers derived from the ph-regulated phr1 and phr2 genes of c. Albicans. J. Clin. Microbiol. 1999, 37, 1587–1590. [Google Scholar] [PubMed]
- Arroyo, J.; Sarfati, J.; Baixench, M.T.; Ragni, E.; Guillen, M.; Rodriguez-Pena, J.M.; Popolo, L.; Latge, J.P. The gpi-anchored gas and crh families are fungal antigens. Yeast 2007, 24, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Bozza, S.; Clavaud, C.; Giovannini, G.; Fontaine, T.; Beauvais, A.; Sarfati, J.; D’Angelo, C.; Perruccio, K.; Bonifazi, P.; Zagarella, S.; et al. Immune sensing of Aspergillus fumigatus proteins, glycolipids, and polysaccharides and the impact on th immunity and vaccination. J. Immunol. 2009, 183, 2407–2414. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Double-displacement reaction for GHs of family 72. In the first step (glycosylation), a covalent glycosyl–enzyme intermediate (GEI) is formed and the leaving group is released. In the second step (deglycosylation), an acceptor molecule reacts with the GEI generating two different products depending on the nature of the acceptor, either a water molecule (R = H, hydrolysis product) or a sugar acceptor (R = sugar, transglycosylation product). The acid/base residue (E176) and the nucleophile group (E275) are those of Gas2p. Reprinted with permission from [39]. Journal of the American Chemical Society, Copyright 2016, American Chemical Society.
Figure 1.
Double-displacement reaction for GHs of family 72. In the first step (glycosylation), a covalent glycosyl–enzyme intermediate (GEI) is formed and the leaving group is released. In the second step (deglycosylation), an acceptor molecule reacts with the GEI generating two different products depending on the nature of the acceptor, either a water molecule (R = H, hydrolysis product) or a sugar acceptor (R = sugar, transglycosylation product). The acid/base residue (E176) and the nucleophile group (E275) are those of Gas2p. Reprinted with permission from [39]. Journal of the American Chemical Society, Copyright 2016, American Chemical Society.
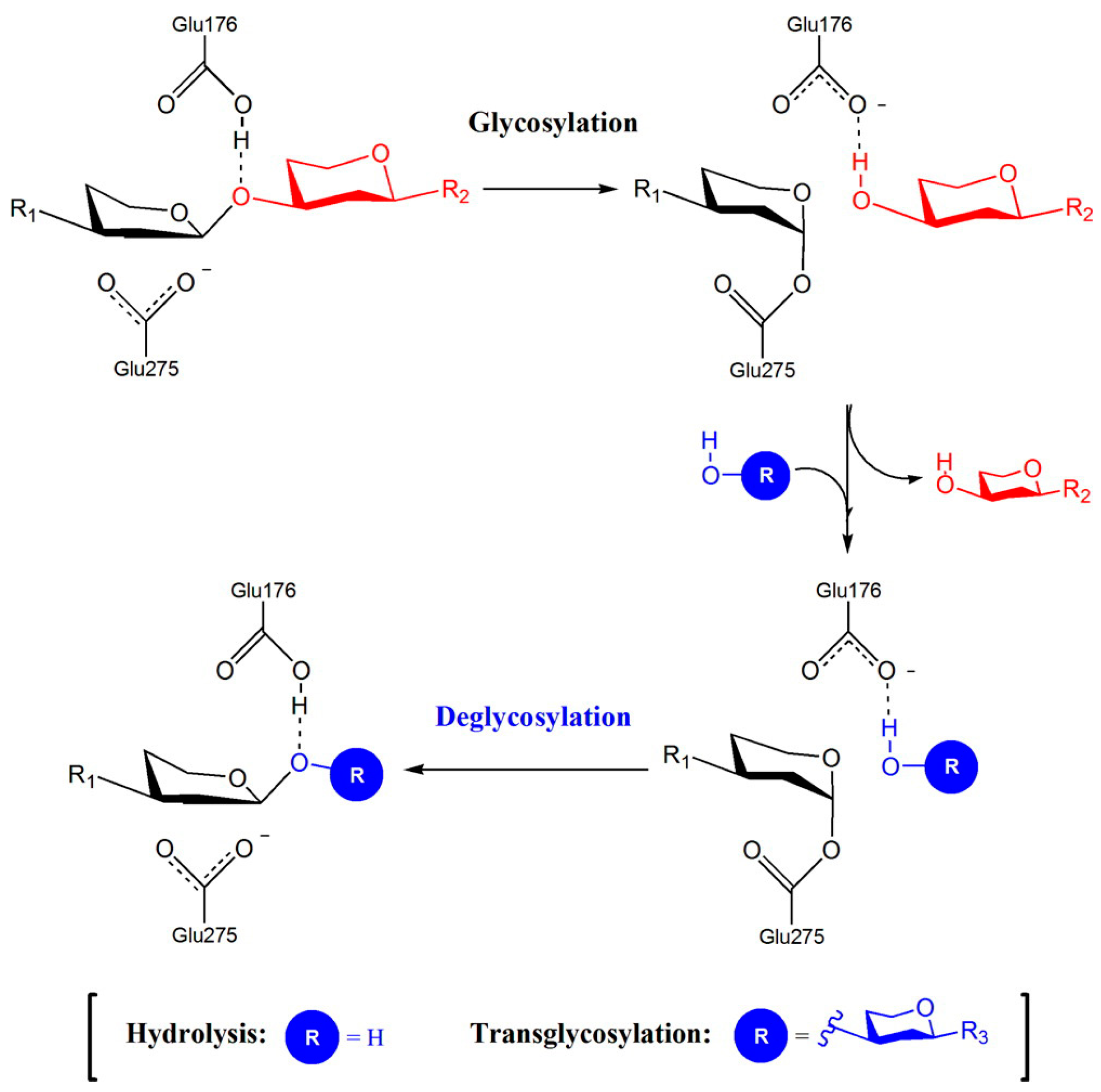
Figure 2.
The Phr family of proteins includes members of the GH72+ and GH72− subfamilies. The asterisk (*) indicates the conserved cysteine residue of the linker region (L). The presence of an X8/CBM43 domain defines the GH72+ subfamily. The catalytic glutamic acid residues and a triad of conserved tyrosines are indicated.
Figure 2.
The Phr family of proteins includes members of the GH72+ and GH72− subfamilies. The asterisk (*) indicates the conserved cysteine residue of the linker region (L). The presence of an X8/CBM43 domain defines the GH72+ subfamily. The catalytic glutamic acid residues and a triad of conserved tyrosines are indicated.
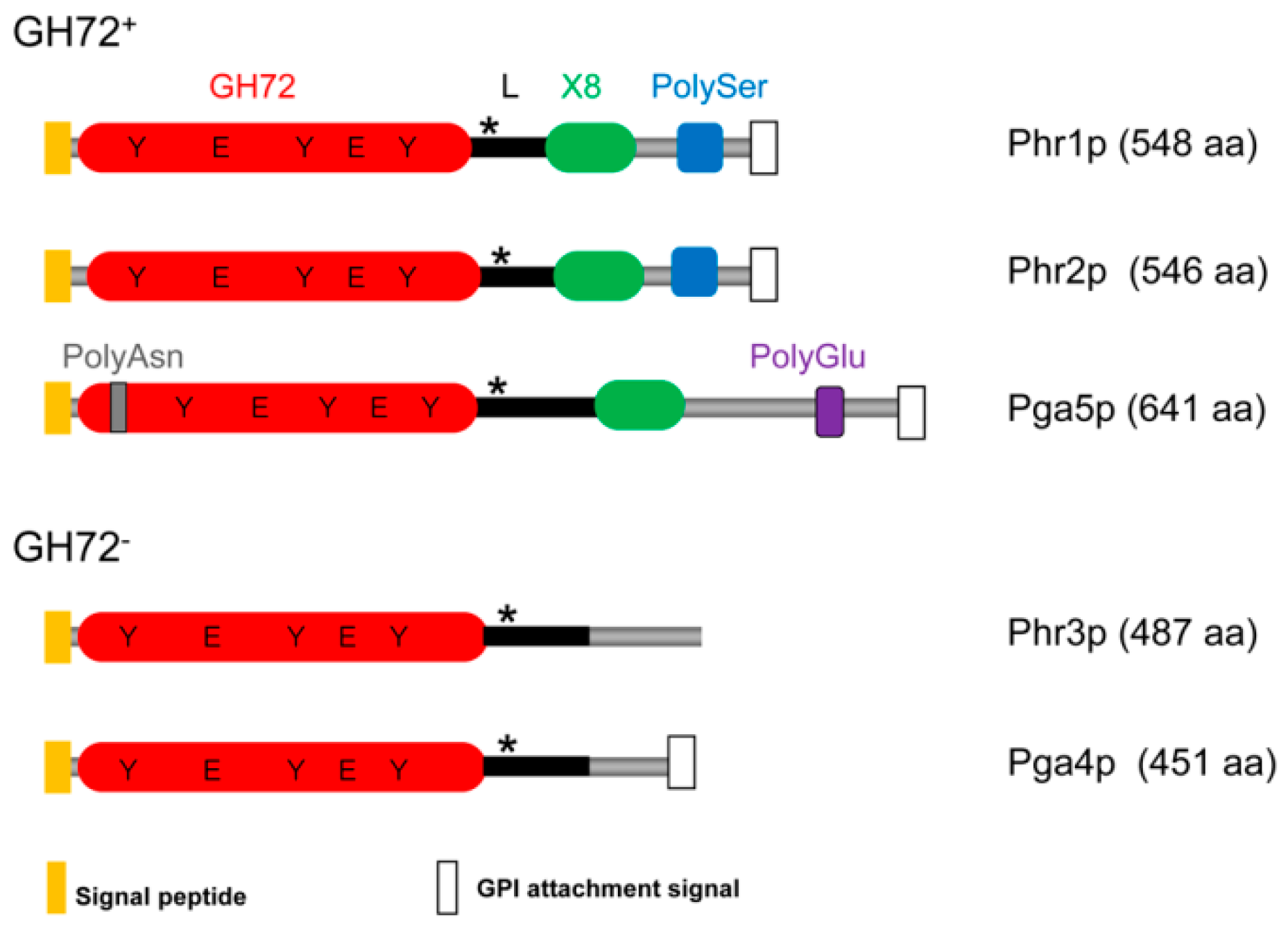
Figure 3.
Molecular model of the three-dimensional structure of Phr2p of C. albicans. On the left is the full enzyme. On the right is the active site: E159, the acid/base group, and E260, the nucleophile (in ball-and-stick representation). Y90, Y292, Y229, and N227 are predicted to affect the dissociation state of the catalytic residues. R288, R125, and N158 are involved in shaping the active site pocket.
Figure 3.
Molecular model of the three-dimensional structure of Phr2p of C. albicans. On the left is the full enzyme. On the right is the active site: E159, the acid/base group, and E260, the nucleophile (in ball-and-stick representation). Y90, Y292, Y229, and N227 are predicted to affect the dissociation state of the catalytic residues. R288, R125, and N158 are involved in shaping the active site pocket.
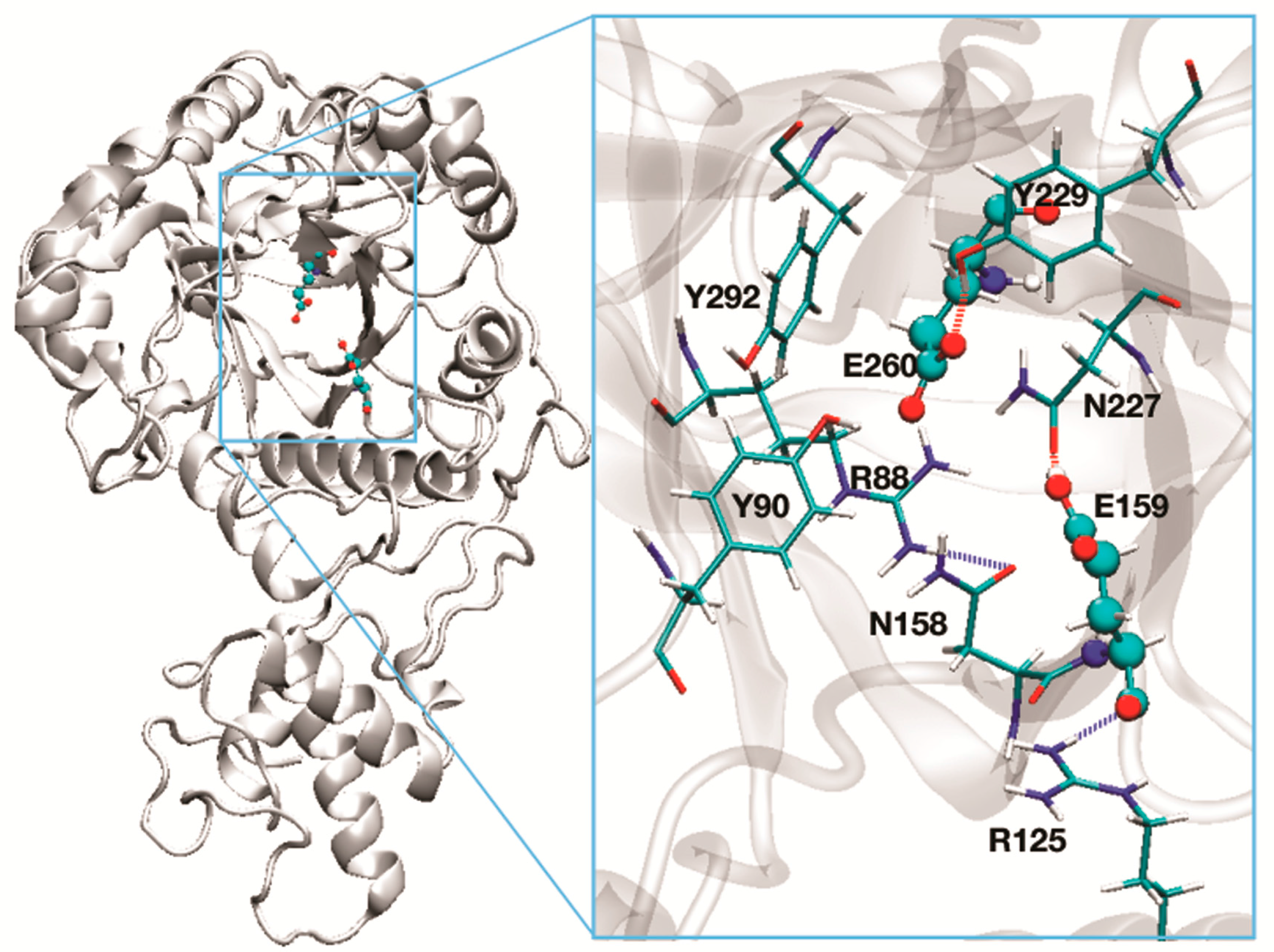
Figure 4.
Comparison of amino acid sequences of PHR and GAS families. Colored blocks indicate C. albicans proteins most closely related to functionally defined S. cerevisiae proteins. Yellow: proteins active in cell wall assembly during vegetative growth of S. cerevisiae. Light blue: proteins active in spore wall assembly in S. cerevisiae. * indicates the presence of a Poly(Glu) tract in the Pga5p C-terminal region.
Figure 4.
Comparison of amino acid sequences of PHR and GAS families. Colored blocks indicate C. albicans proteins most closely related to functionally defined S. cerevisiae proteins. Yellow: proteins active in cell wall assembly during vegetative growth of S. cerevisiae. Light blue: proteins active in spore wall assembly in S. cerevisiae. * indicates the presence of a Poly(Glu) tract in the Pga5p C-terminal region.
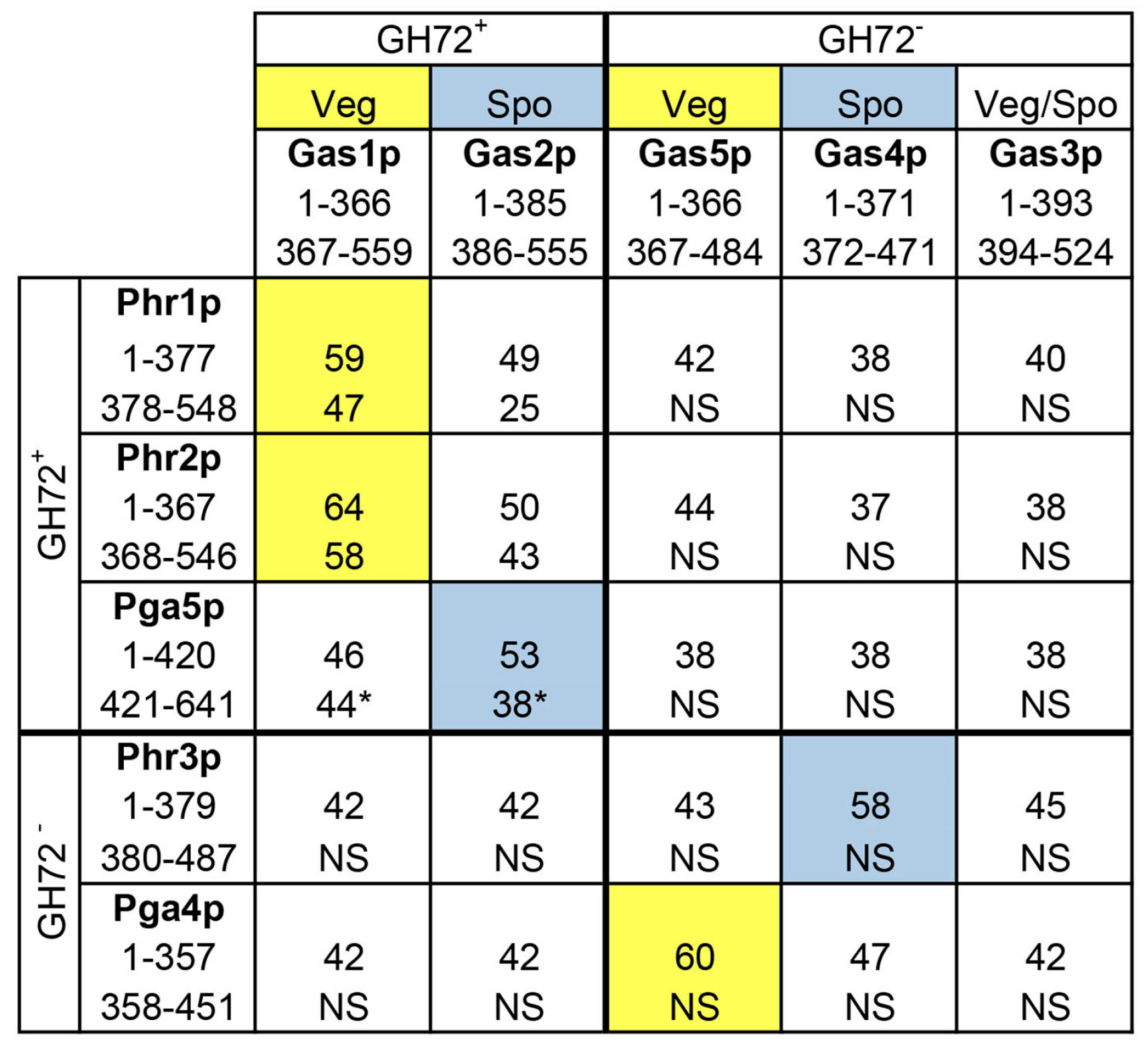
Figure 5.
Scheme for the assembly of cell wall components in the extracellular space. In red are indicated the family of transglycosylases described in the text; in light blue: β-(1,3)-glucan; in orange β-(1,6)-glucose-linked residues. GH72 and GH17 cooperate in creating the core glucan structure and the anchoring sites for the other cell wall components. Pir are proteins with internal repeats that are directly linked to β-(1,3)-glucan chains through alkali labile ester linkages [52].
Figure 5.
Scheme for the assembly of cell wall components in the extracellular space. In red are indicated the family of transglycosylases described in the text; in light blue: β-(1,3)-glucan; in orange β-(1,6)-glucose-linked residues. GH72 and GH17 cooperate in creating the core glucan structure and the anchoring sites for the other cell wall components. Pir are proteins with internal repeats that are directly linked to β-(1,3)-glucan chains through alkali labile ester linkages [52].
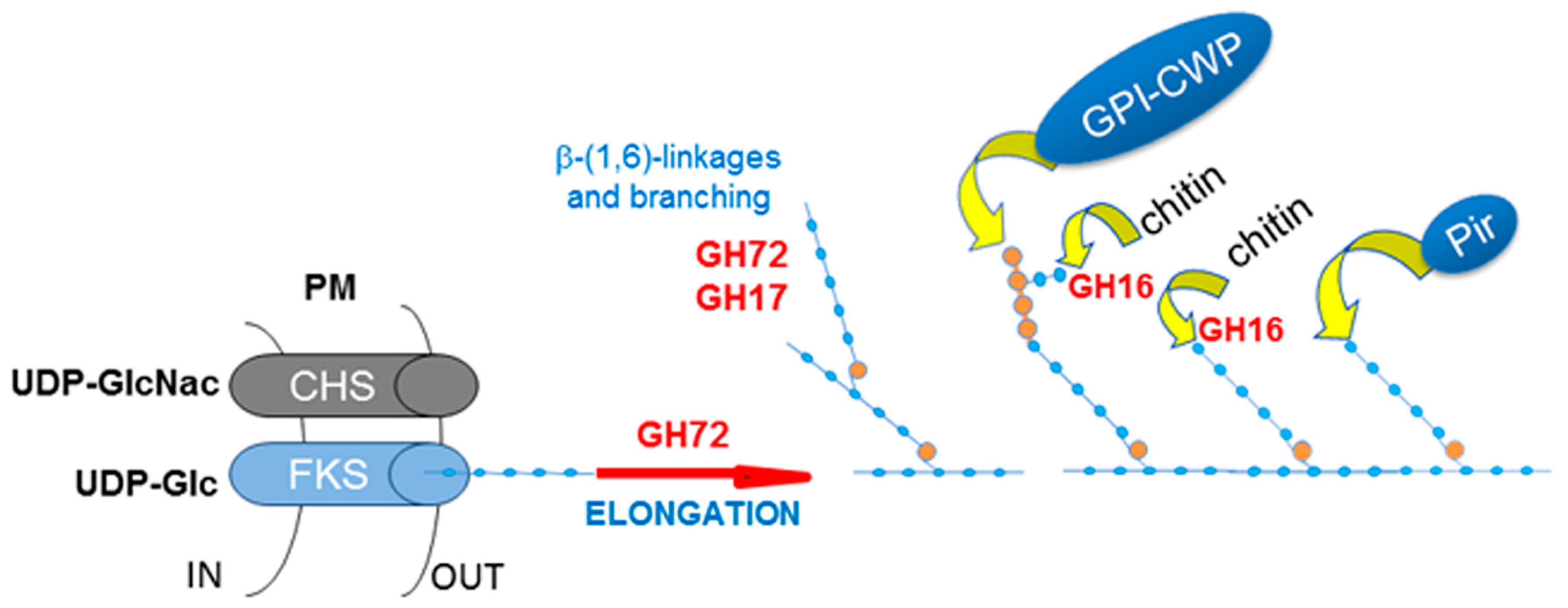
Figure 6.
Localization of Phr1p-GFP in C. albicans. Cells growing in YPD-150 mM HEPES pH 7.5 at 25 °C.
Figure 6.
Localization of Phr1p-GFP in C. albicans. Cells growing in YPD-150 mM HEPES pH 7.5 at 25 °C.
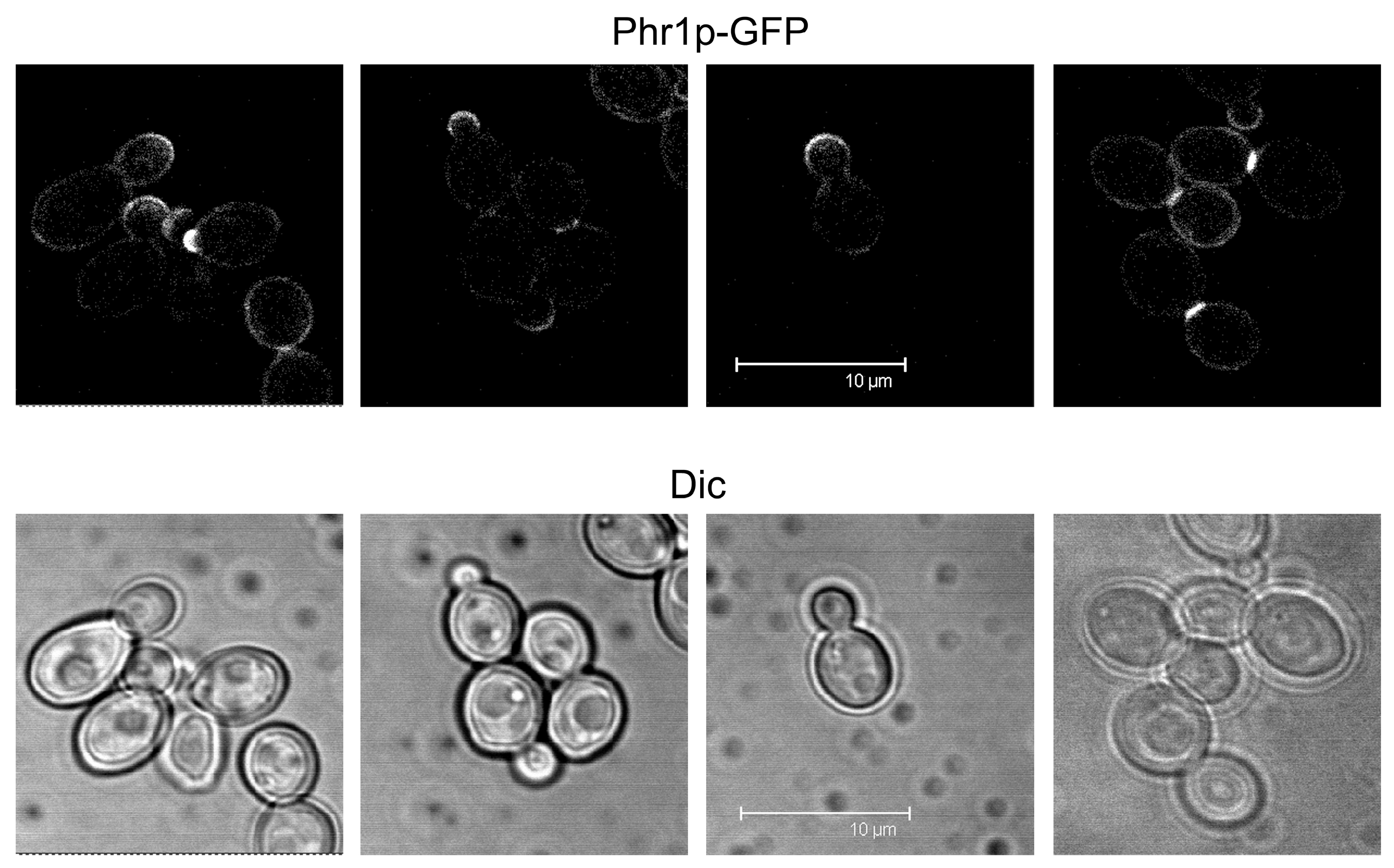
Figure 7.
Localization of Phr1p-GFP in C. albicans. Phr1p-GFP localization in conditions of induction of hyphal growth in M199–150 mM HEPES, pH 7.5 at 37 °C, Bar: 5 µm. Modified from [57].
Figure 7.
Localization of Phr1p-GFP in C. albicans. Phr1p-GFP localization in conditions of induction of hyphal growth in M199–150 mM HEPES, pH 7.5 at 37 °C, Bar: 5 µm. Modified from [57].
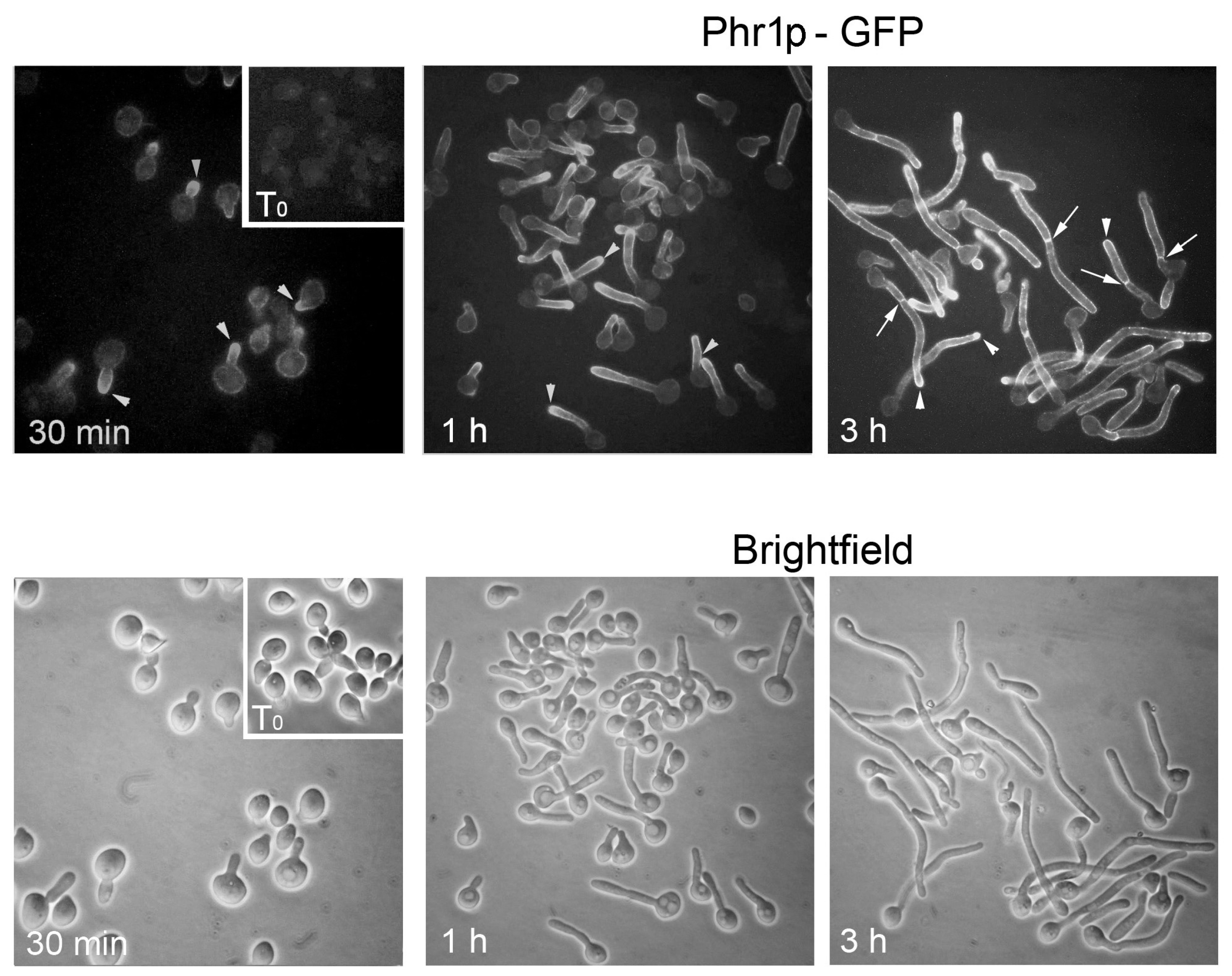
Figure 8.
Diagram of input signal and signaling pathways activated in C. albicans by cell wall stress. On the right (in the black box), the inhibitors of calcineurin and in white the target proteins. P in the black box represents the phosphate group that calcineurin removes from Crz1p. The question mark on Psk1p indicates that cell wall damage activates Psk1p in an unknown manner. The question mark on CaCas5 indicates an unknown activator of this TF that regulates the cell wall stress-response. Reproduced from [92].
Figure 8.
Diagram of input signal and signaling pathways activated in C. albicans by cell wall stress. On the right (in the black box), the inhibitors of calcineurin and in white the target proteins. P in the black box represents the phosphate group that calcineurin removes from Crz1p. The question mark on Psk1p indicates that cell wall damage activates Psk1p in an unknown manner. The question mark on CaCas5 indicates an unknown activator of this TF that regulates the cell wall stress-response. Reproduced from [92].

Figure 9.
Phenotype of the indicated mutants in liquid YPD-150 mM HEPES, pH 8 at 25 °C (vegetative growth). Cells were sonicated for three cycles of 6 s. The percentages of lysed cells were: WT: 2%, phr1∆: 14%, chs3∆: 55% and phr1∆ chs3∆: 85%. The arrows indicate fractures of the cell wall and release of cellular material, Bar: 5 µm. Unpublished results.
Figure 9.
Phenotype of the indicated mutants in liquid YPD-150 mM HEPES, pH 8 at 25 °C (vegetative growth). Cells were sonicated for three cycles of 6 s. The percentages of lysed cells were: WT: 2%, phr1∆: 14%, chs3∆: 55% and phr1∆ chs3∆: 85%. The arrows indicate fractures of the cell wall and release of cellular material, Bar: 5 µm. Unpublished results.
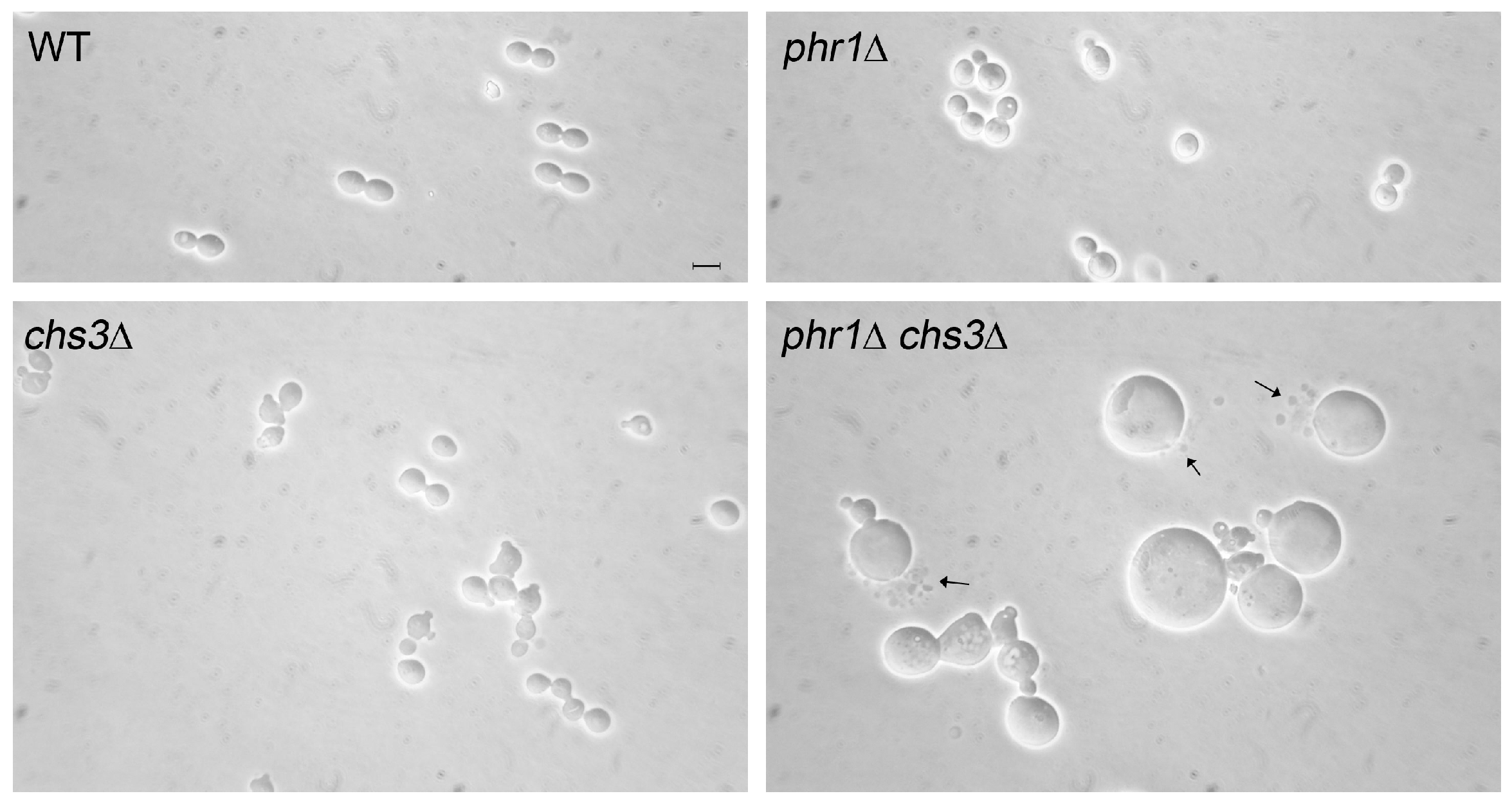
Figure 10.
Model of the sequential involvement of the chitin synthases in the adaptation to cell wall stress of a phr1 cell during induction of hyphal growth in M199 at a restrictive pH. First, at the time of germ tube emergence Chs3p is crucial in liquid media and, if not present, phr1Δ cells arrest at early germ tube emergence, swell and die. On solid surface, Chs3p is crucial for the onset of germination and if absent phr1Δ cells do not produce colonies. Chs8p is crucial for the development of the germ tube. Once the germ tube has emerged, Chs8p acts to protect the tip. If Chs8p is absent during growth on a solid medium, phr1Δ cells give rise to abortive microcolonies containing dead cells.
Figure 10.
Model of the sequential involvement of the chitin synthases in the adaptation to cell wall stress of a phr1 cell during induction of hyphal growth in M199 at a restrictive pH. First, at the time of germ tube emergence Chs3p is crucial in liquid media and, if not present, phr1Δ cells arrest at early germ tube emergence, swell and die. On solid surface, Chs3p is crucial for the onset of germination and if absent phr1Δ cells do not produce colonies. Chs8p is crucial for the development of the germ tube. Once the germ tube has emerged, Chs8p acts to protect the tip. If Chs8p is absent during growth on a solid medium, phr1Δ cells give rise to abortive microcolonies containing dead cells.
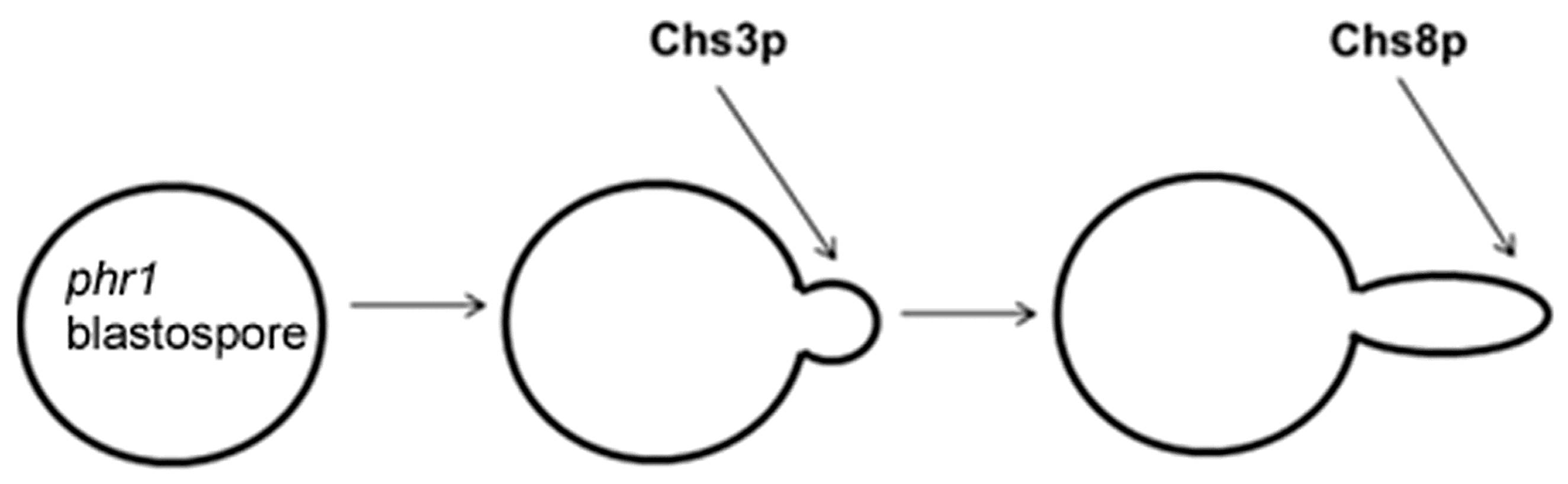

{kind=link}
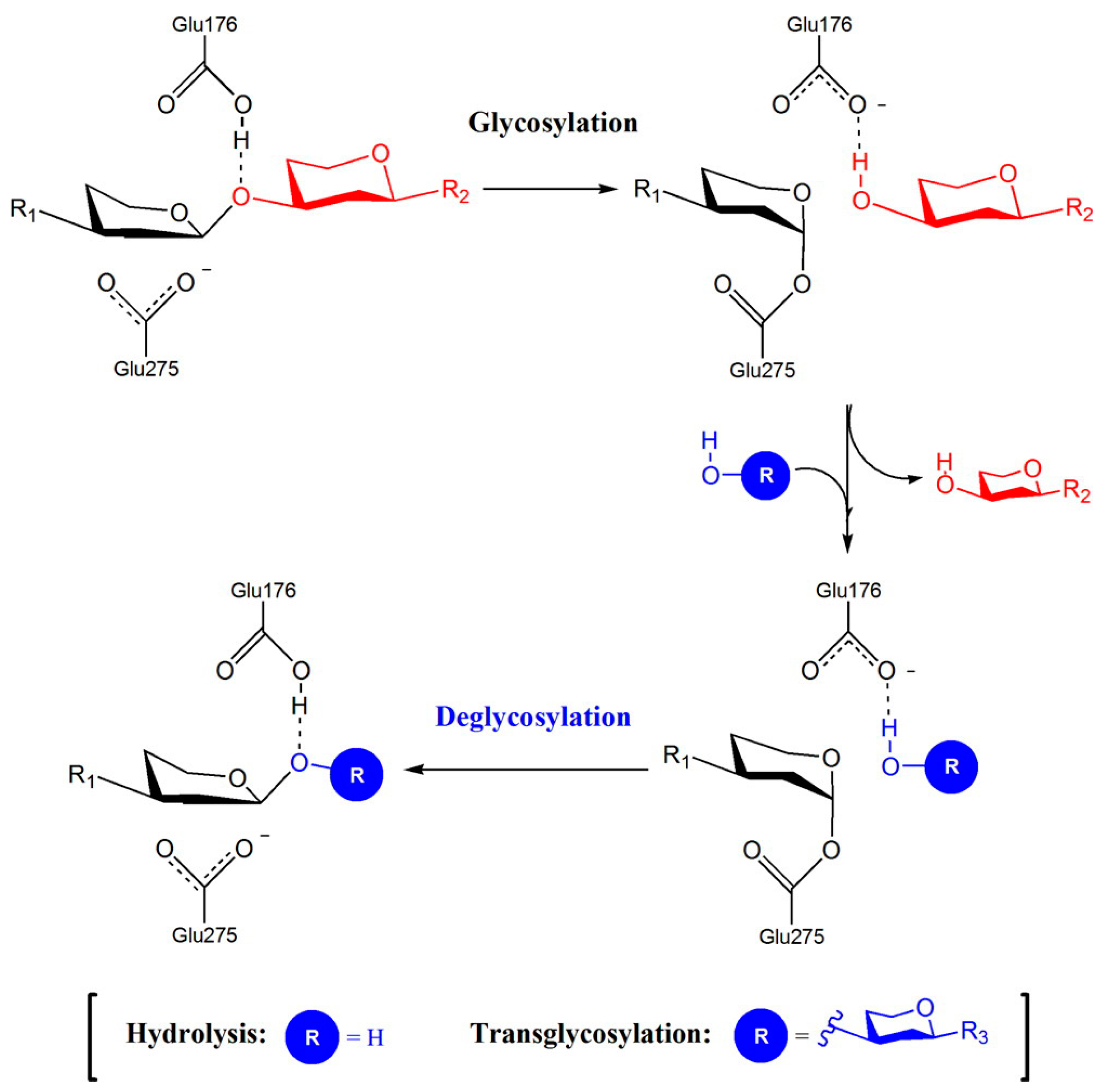{kind=link}
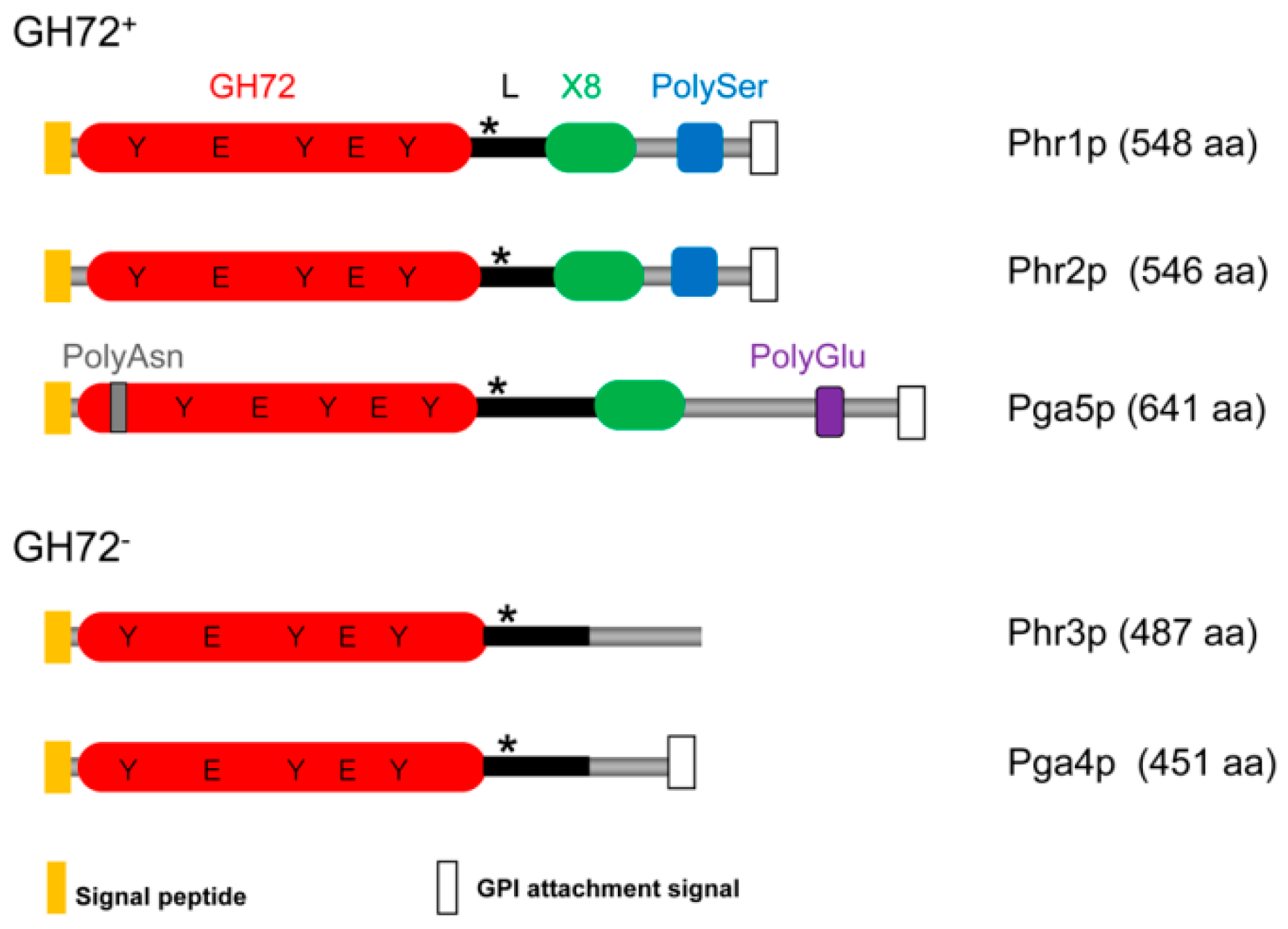{kind=link}
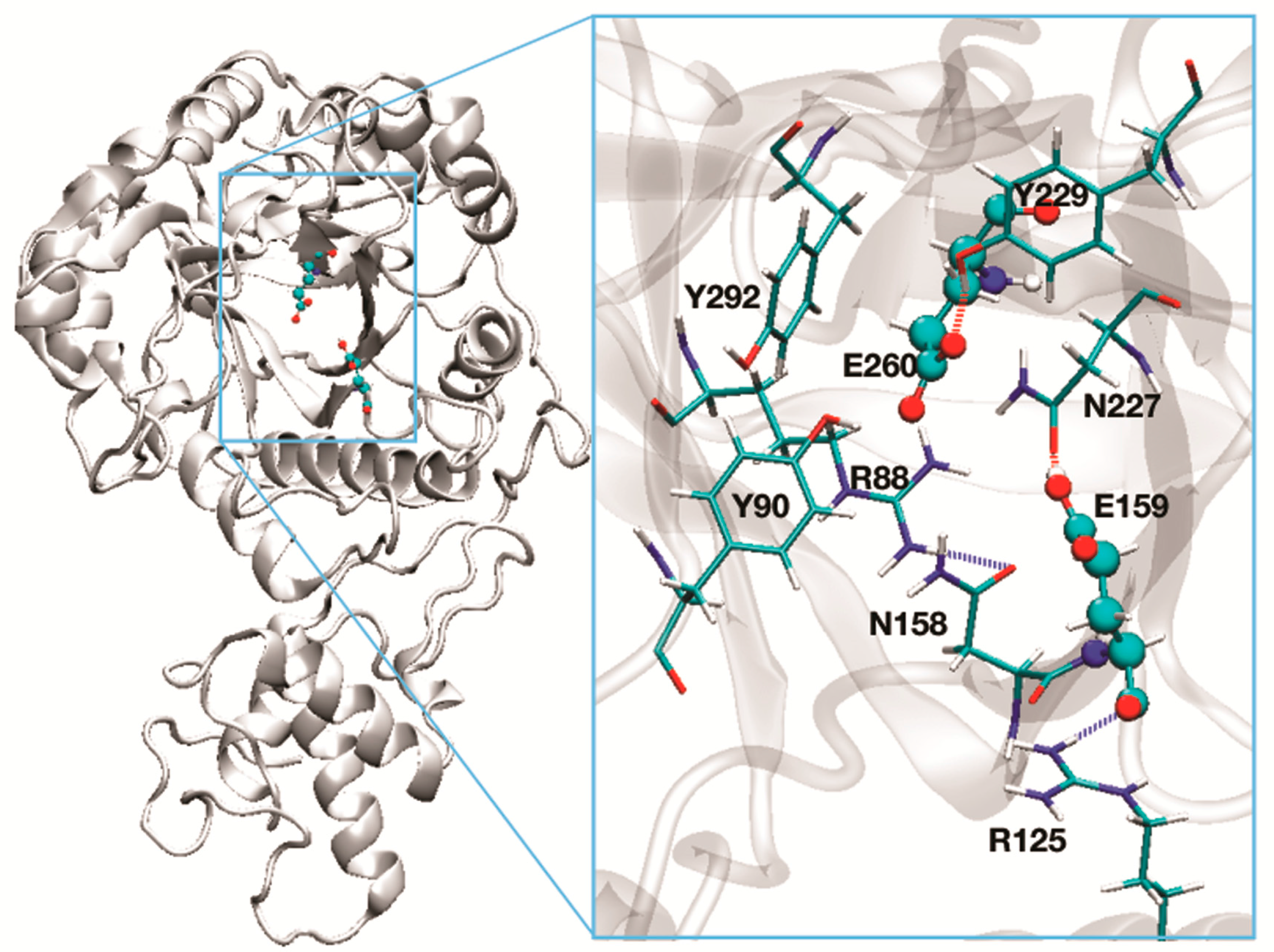{kind=link}
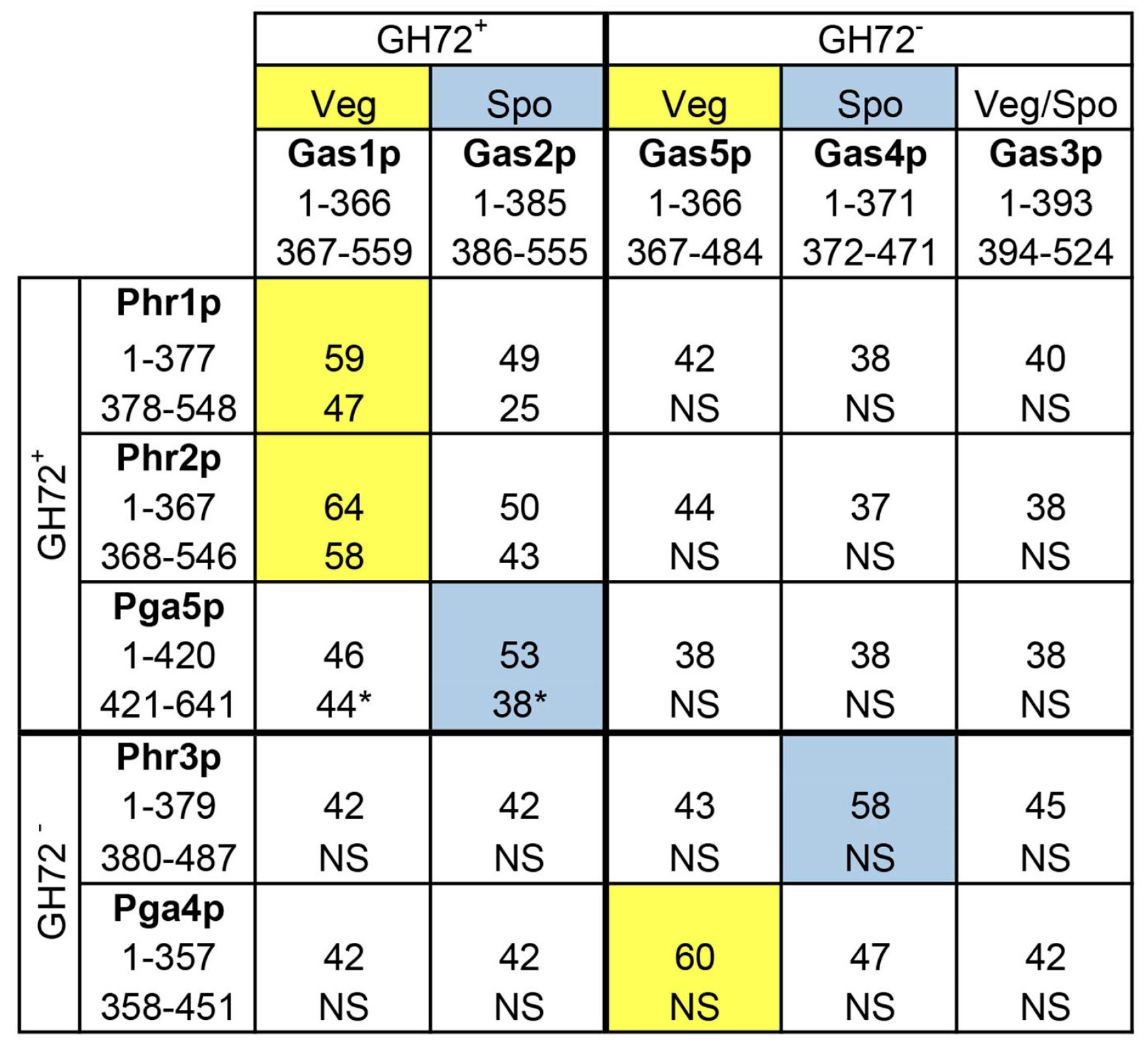{kind=link}
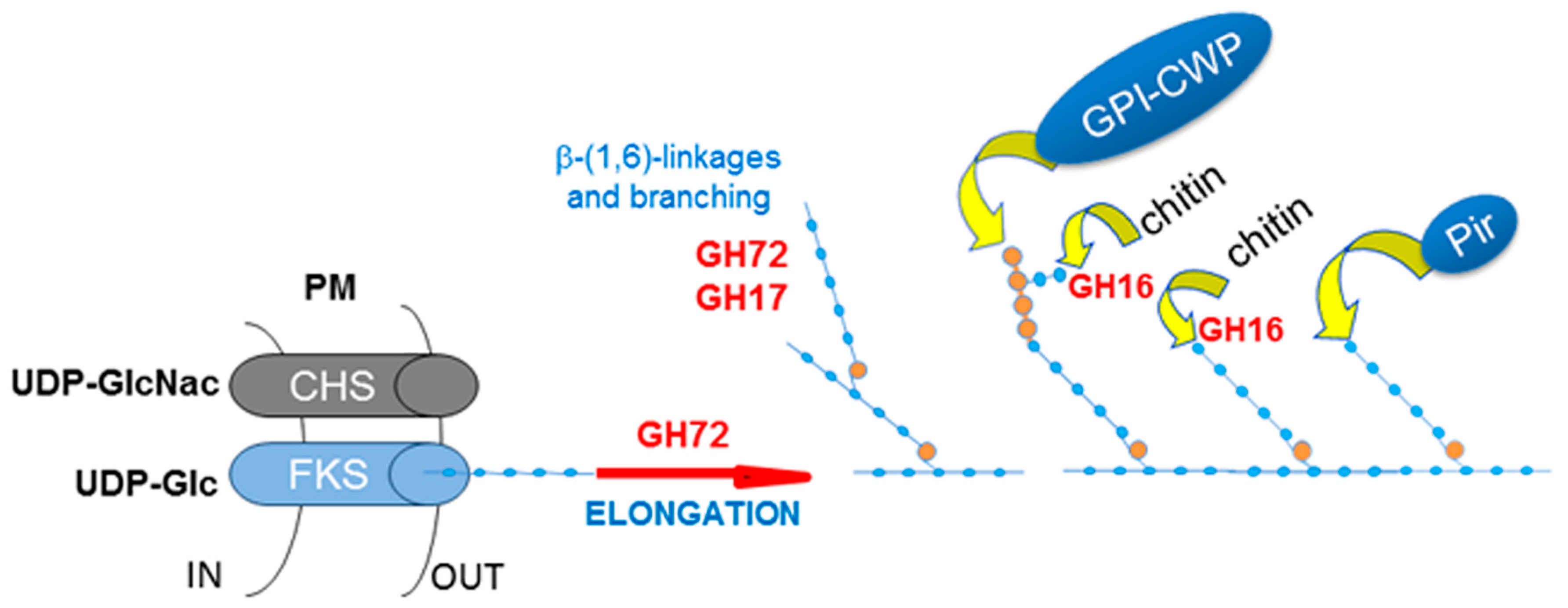{kind=link}
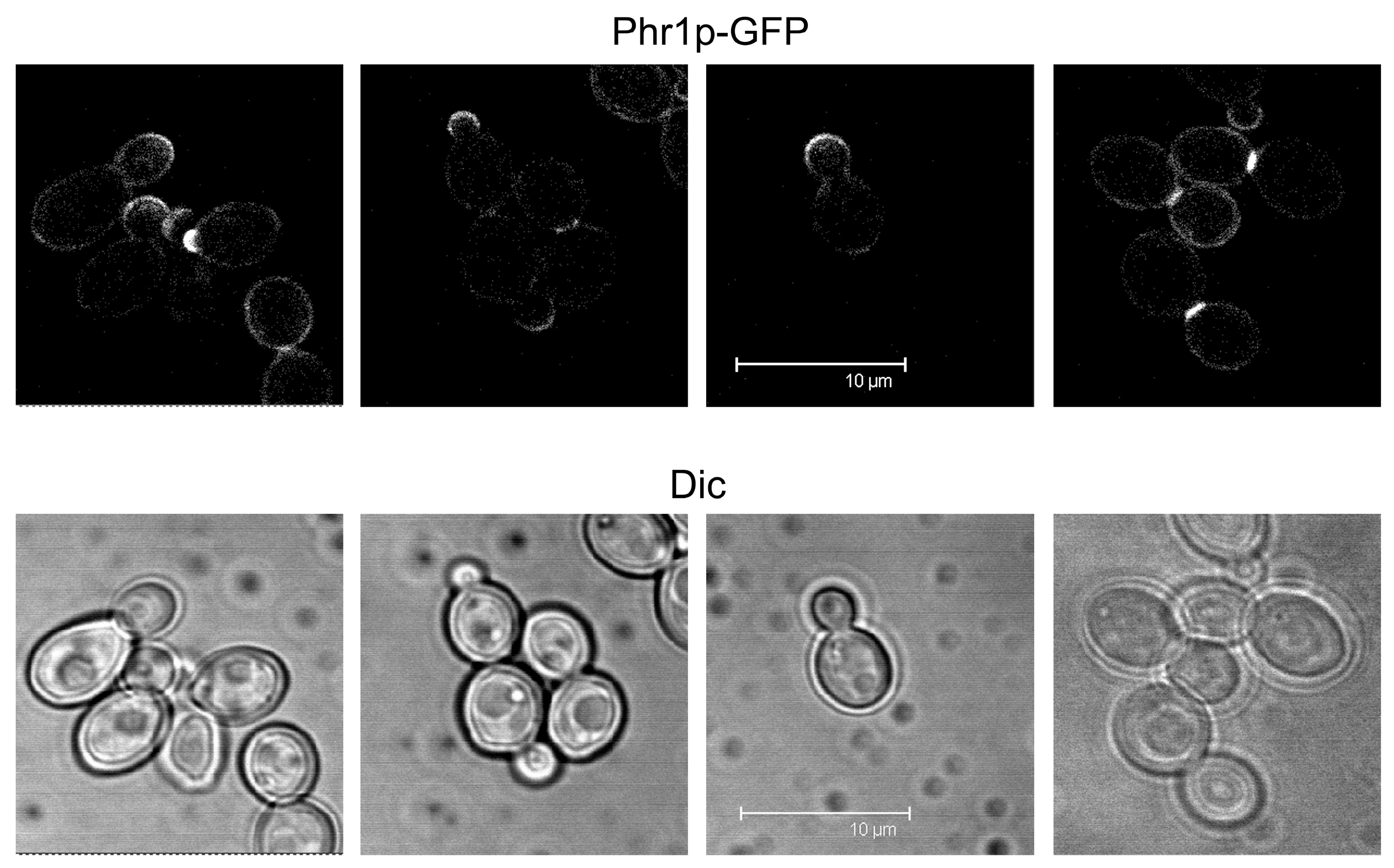{kind=link}
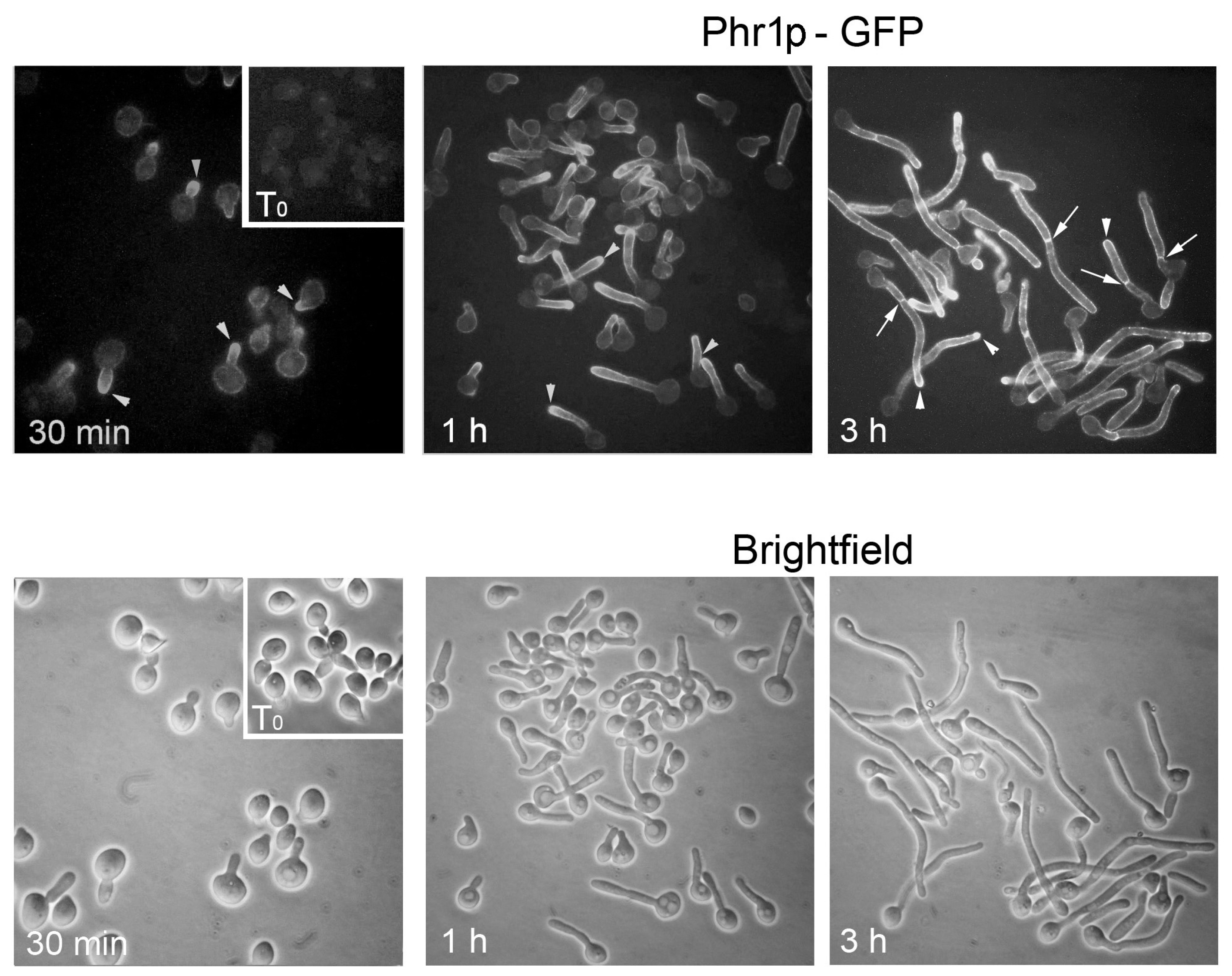{kind=link}
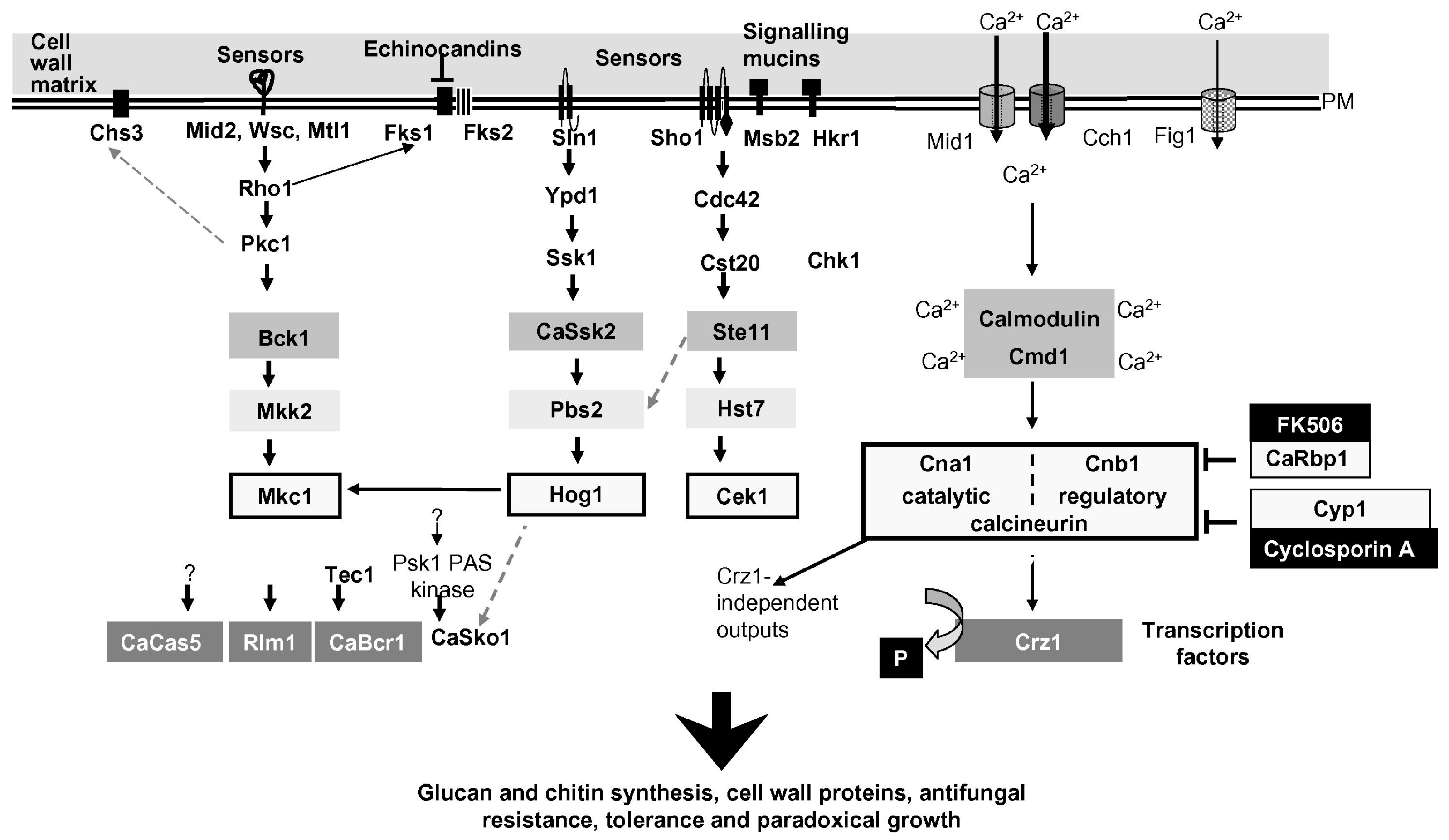{kind=link}
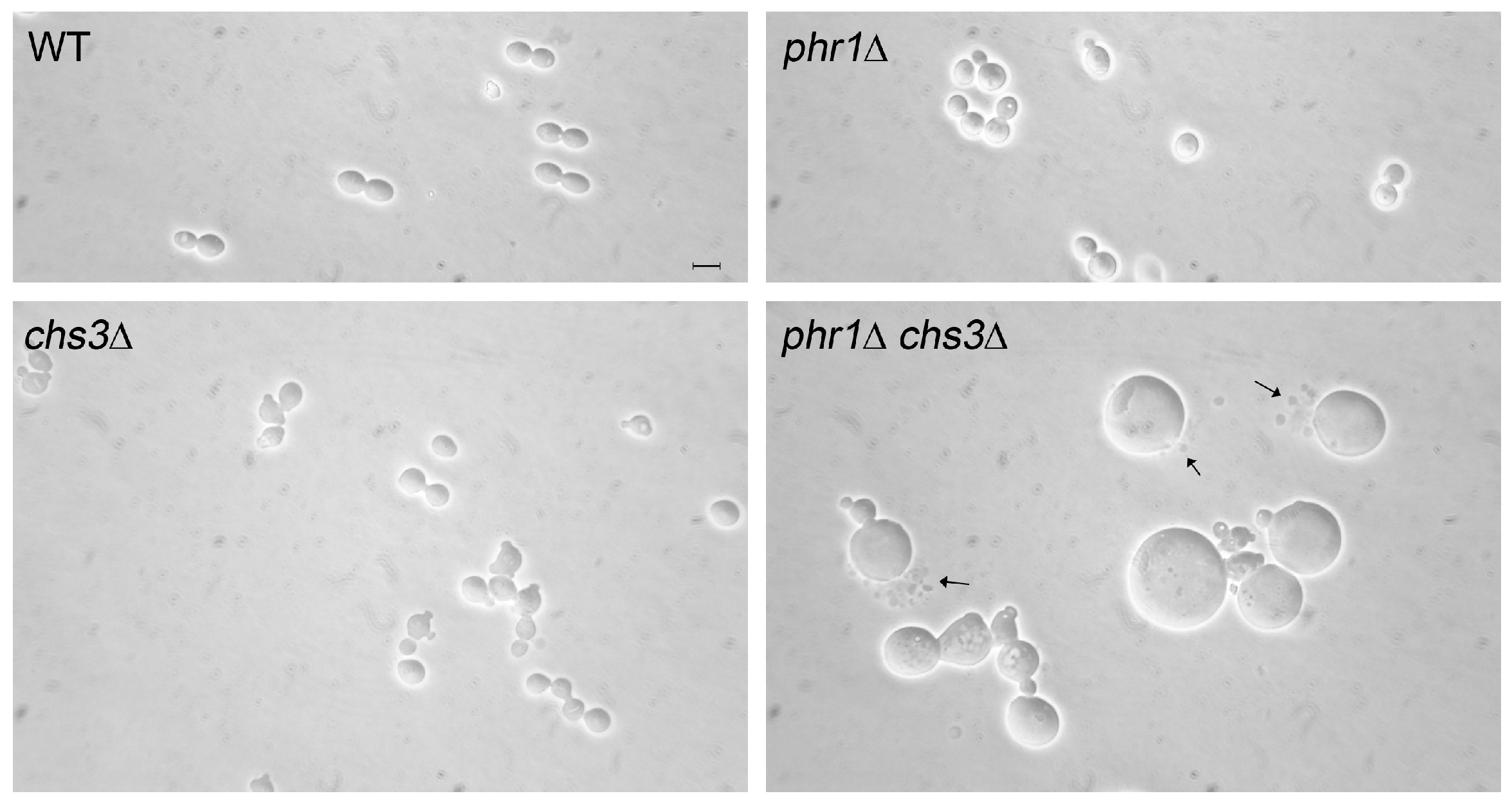{kind=link}
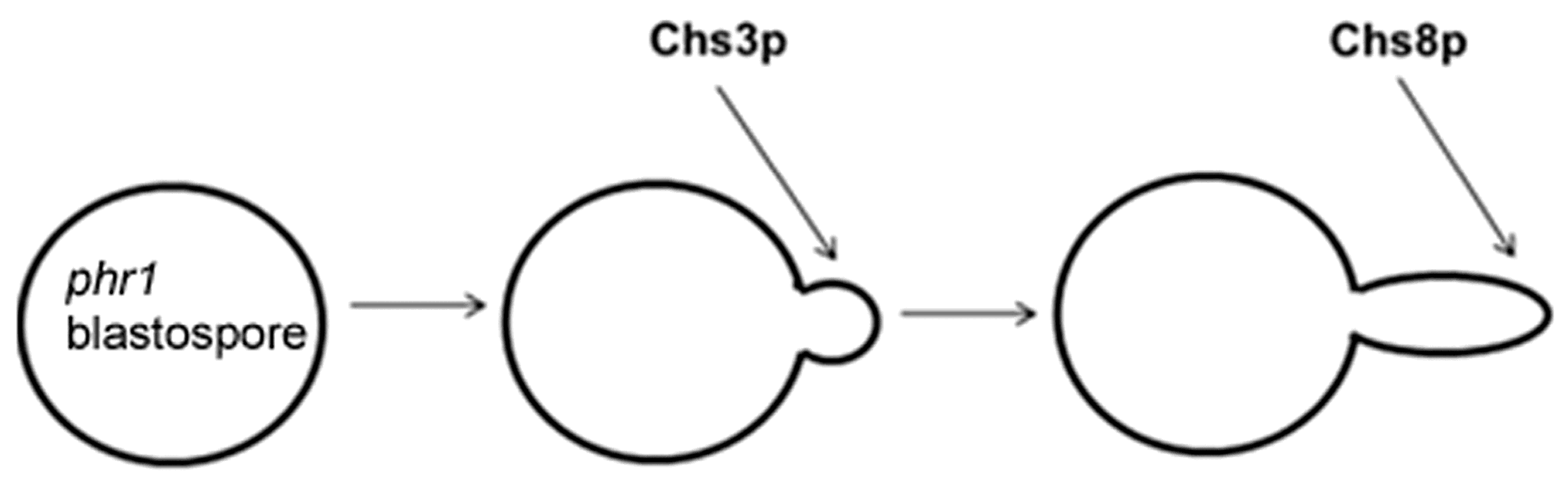{kind=link}
Table 1.
Catalytic properties of Phr proteins of Candida albicans 1.
| Phr1p | Phr2p | |
|---|---|---|
| Activity in vitro | Endo β-(1,3)-glucanase/transglysosylase | Endo β-(1,3)-glucanase/transglysosylase |
| Temperature optimum | 30 °C | 30 °C |
| pH optimum | 5.8 | 3 |
| Half-time of activity decay at 95 °C | 10 min | 15 min |
| Minimum length of the donor 2 | rG9 | rG10 |
| Preferred acceptor length | ≥L7 | ≥L7 |
| Residues essential for catalysis 3 | Glu169 and Glu270 | Glu159 and Glu260 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Popolo, L.; Degani, G.; Camilloni, C.; Fonzi, W.A. The PHR Family: The Role of Extracellular Transglycosylases in Shaping Candida albicans Cells. J. Fungi 2017, 3, 59. https://doi.org/10.3390/jof3040059
AMA Style
Popolo L, Degani G, Camilloni C, Fonzi WA. The PHR Family: The Role of Extracellular Transglycosylases in Shaping Candida albicans Cells. Journal of Fungi. 2017; 3(4):59. https://doi.org/10.3390/jof3040059
Chicago/Turabian StylePopolo, Laura, Genny Degani, Carlo Camilloni, and William A. Fonzi. 2017. "The PHR Family: The Role of Extracellular Transglycosylases in Shaping Candida albicans Cells" Journal of Fungi 3, no. 4: 59. https://doi.org/10.3390/jof3040059
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.