Genotypic Diversity Is Independent of Pathogenicity in Colombian Strains of Cryptococcus neoformans and Cryptococcus gattii in Galleria mellonella


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolates
2.2. Macroscopic Morphology
2.3. Mating Type Determination
2.4. Invertebrate Model Galleria mellonella
2.5. Cellular and Capsular Size Determination
2.6. Statistical Analysis
3. Results
3.1. Macroscopic Morphology
3.2. Mating Type Determination
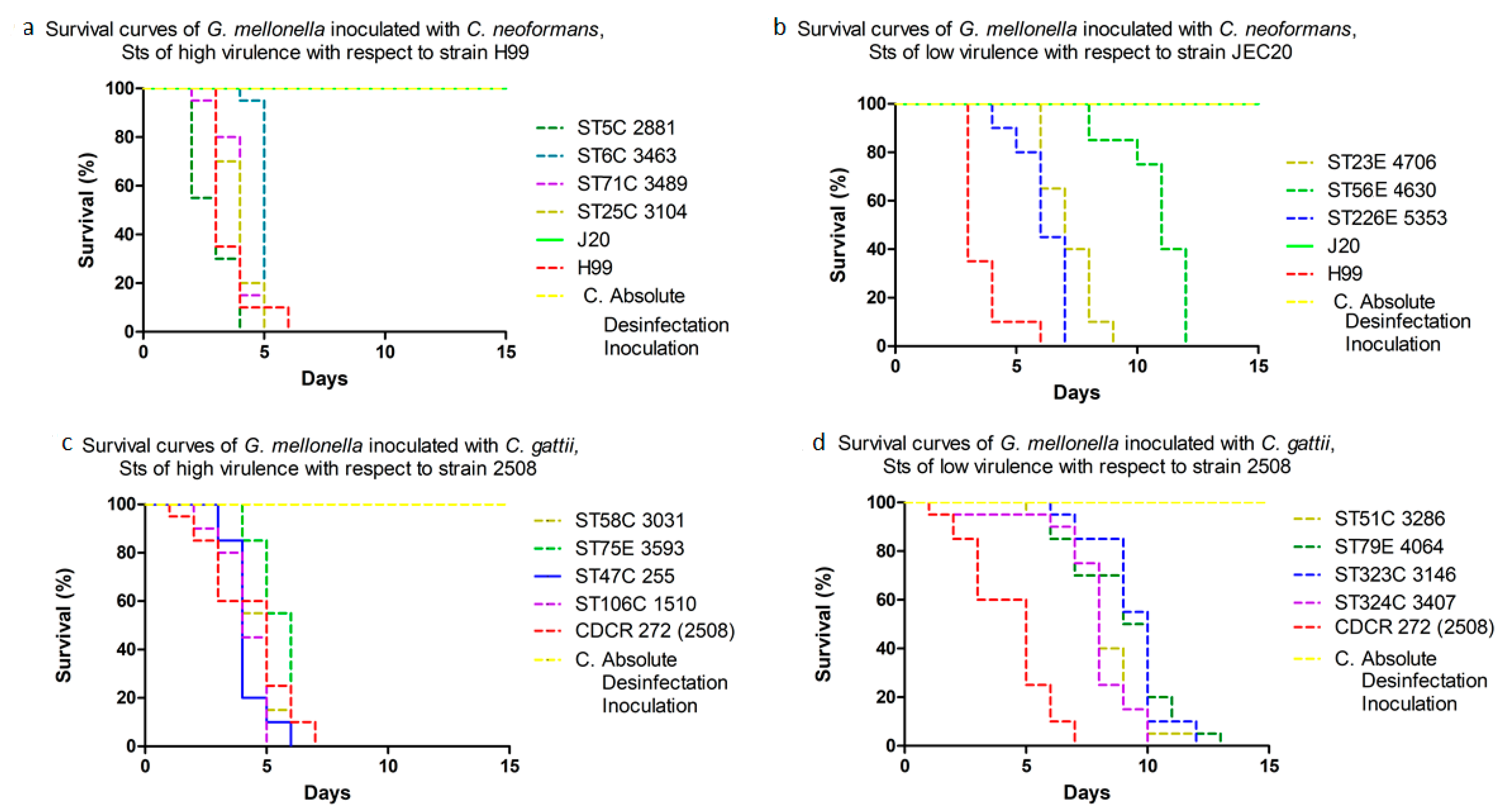
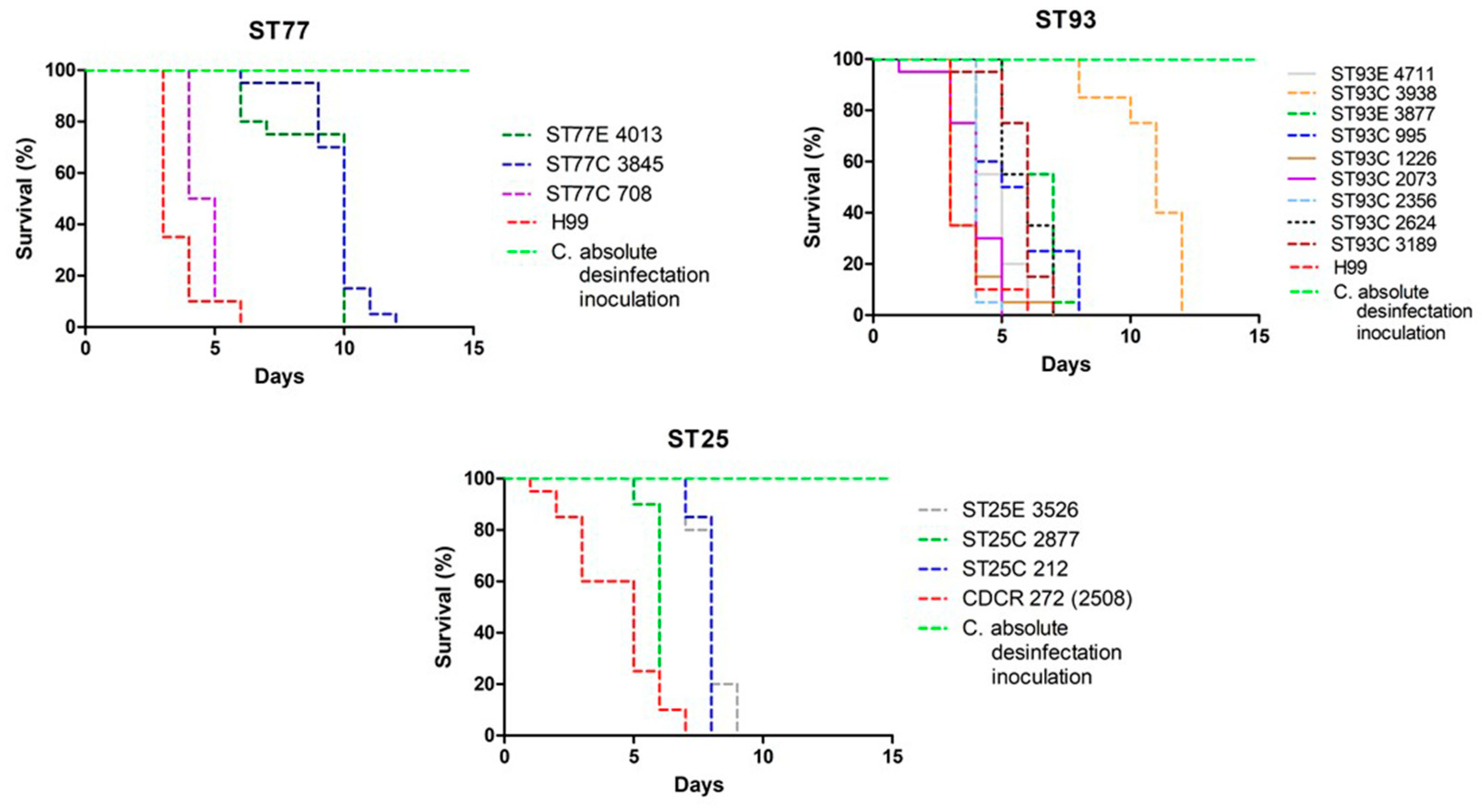
3.3. Invertebrate Model Survival Curves
3.4. Cellular and Capsular Size Determination Pre- and Post-Inoculation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lin, Y.Y.; Shiau, S. Risk factors for invasive Cryptococcus neoformans diseases: A case-control study. PLoS ONE 2015, 10, e0119090. [Google Scholar] [CrossRef] [PubMed]
- Mesquita da Costa, M.; Teixeira, F.M.; Schalcher, T.R.; Magalhães de Brito, F.; Silva, E.; Chagas, M. Cryptococcosis, a risk for immunocompromised and immunocompetent individuals. OpenEpi 2013, 6, 9–17. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E.; Wickes, B.L.; Meyer, W.; Cuomo, C.A.; Wollenburg, K.R.; Bicanic, T.A.; Castañeda, E.; Chang, Y.C.; Chen, J.; et al. The Case for Adopting the “Species Complex” Nomenclature for the Etiologic Agents of Cryptococcosis. mSphere 2017, 11. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.B.; O’Brien, E.; Khoury, J.B.; Mylonakis, E. Methods for using Galleria mellonella as a model host to study fungal pathogenesis. Virulence 2010, 1, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Moreno, R.; El Khoury, J.B.; Idnurm, A.; Heitman, J.; Calderwood, S.B.; Ausubel, F.M.; Diener, A. Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 2005, 73, 3842–3850. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; McArthur, J. Developing Galleria mellonella as a model host for human pathogens. Virulence 2013, 4, 350–353. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.; Thomas, D.; Whiteway, M.; Kavanagh, K. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med. Microbiol. 2002, 34, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martínez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol. 2013, 51, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Thomaz, L.; Garcia-Rodas, R.; Guimaraes, A.; Taborda, C.P.; Zaragoza, O.; Nosanchuk, J.D. Galleria mellonella as a model host to study Paracoccidioides lutzii and Histoplasma capsulatum. Virulence 2013, 4, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Firacative, C.; Shuyao, D.; Meyer, W. Galleria mellonella model identifies highly virulent strains among all major molecular types of Cryptococcus gattii. PLoS ONE 2014, 9, e1050576. [Google Scholar] [CrossRef] [PubMed]
- Escandón, P.; Sánchez, A.; Martínez, M.; Meyer, W.; Castañeda, E. Molecular epidemiology of clinical and environmental isolates of the Cryptococcus neoformans species complex reveals a high genetic diversity and the presence of the molecular type VGII mating type a in Colombia. FEMS Yeast Res. 2006, 6, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.; Castañeda, A.; Jackson, S.; Huynh, M.; Castañeda, E. Molecular typing of Ibero American Cryptococcus neoformans isolates. Emerg. Infect. Dis. 2003, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Latouche, A.; Huynh, M.; Sorrell, T.; Meyer, W. PCR-restriction fragment length polymorphism analysis of the phospholipase B (PLB1) gene for subtyping of Cryptococcus neoformans isolates. Appl. Environ. Microbiol. 2003, 69, 2080–2086. [Google Scholar] [CrossRef] [PubMed]
- Meyer, W.; Aanensen, D.M.; Boekhout, T.; Cogliati, M.; Diaz, M.R.; Esposto, M.C.; Fisher, M.; Gilgado, F.; Hagen, F.; Kaocharoen, S. Consensus multi-locus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med. Mycol. 2009, 47, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.; Giles, S.; Wenink, E.; Geunes-Boyer, S.G.; Wright, J.R.; Diezmann, S.; Allen, A.; Stajich, J.E.; Dietrich, F.S.; Perfect, J.R.; et al. Same-sex mating and the origin of the Vancouver Island Cryptococcus gattii outbreak. Nature 2005, 437, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.R., 3rd; Albert, N.; Hodge, G.; Wilson, M.D.; Sykes, J.E.; Bays, D.J.; Firacative, C.; Meyer, W.; Kontoyiannis, D.P. Phenotypic differences of Cryptococcus molecular types: Implications for virulence in a Drosophila model of infection. Infect. Immun. 2014, 82, 3058–3065. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Escandón, P.; Castañeda, E. In vitro determination of virulence factors activity associated with several Cryptococcus neoformans clinical isolates. Rev. Iberoam. Micol. 2008, 25, 145–149. [Google Scholar] [CrossRef]
- Casali, A.K.; Goulart, L.; Rosa e Silva, L.K.; Ribeiro, A.M.; Amaral, A.A.; Alves, S.H.; Schrank, A.; Meyer, W.; Vainstein, M.H. Molecular typing of clinical and environmental Cryptococcus neoformans isolates in the Brazilian state Rio Grande do Soul. FEMS Yeast Res. 2003, 3, 405–415. [Google Scholar] [CrossRef]
- Halliday, C.L.; Bul, T.; Krockenberger, M.; Malik, R.; Ellis, D.; Carter, D. Presence of α and a Mating types in environmental and clinical collections of Cryptococcus neoformans var. gattii strains from Australia. J. Clin. Microbiol. 1999, 37, 2920–2926. [Google Scholar] [PubMed]
- Franzot, S.; Mukherjee, J.; Cherniak, R.; Chen, L.C.; Hamdan, J.S.; Casadevall, A. Microevolution of a standard strain of Cryptococcus neoformans resulting in differences in virulence and other phenotypes. Infect. Immun. 1998, 66, 89–97. [Google Scholar] [PubMed]
- Daw, B.; Trapp, R. Bioestadística Medica, 4th ed.; Manual Moderno: Mexico City, Mexico, 2005; ISBN 9789707291348. [Google Scholar]
- Fries, B.; Goldman, D.; Cherniak, R.; Ju, R.; Casadevall, A. Phenotypic switching in Cryptococcus neoformans results in changes in cellular morphology and glucoronoxylomannan structure. Infect. Immun. 1999, 67, 6076–6083. [Google Scholar] [PubMed]
- Feldmesser, M.; Kress, Y.; Casadevall, A. Dynamic changes in the morphology of Cryptococcus neoformans during murine pulmonary infection. Microbiology 2001, 147, 2355–2365. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O.; Rodrigues, M.; De Jesus, M.; Frases, S.; Dadachova, E.; Caadevall, A. The capsule of the fungal pathogen Cryptococcus neoformans. Adv. Appl. Microbiol. 2009, 68, 133–216. [Google Scholar] [PubMed]
- García-Rodas, R.; Casadevall, A.; Rodríguez-Tudela, J.; Cuenca-Estrella, M.; Zaragoza, O. Cryptococcus neoformans Capsular Enlargement and Cellular Gigantism during Galleria mellonella Infection. PLoS ONE 2011, 6, e24485. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual cycle of Cryptococcus neoformans var. grubii and virulence of congenic a and alpha isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [PubMed]
- Kwon-Chung, K.; Edman, J.; Wickes, B. Genetic association of mating types and virulence in Cryptococcus neoformans. Infect. Immun. 1992, 60, 602–605. [Google Scholar] [PubMed]
- Byrnes, E.; Bildfell, R.; Frank, S.; Mitchell, T.G.; Marr, K.A.; Heitman, J. Molecular evidence that the range of the Vancouver Island outbreak of Cryptococcus gattii infection has expanded into the Pacific Northwest in the United States. J. Infect. Dis. 2009, 199, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Kaufman, L.; Maekura, S.; Teramura, K.; Satou, T.; Hashimoto, S. Pulmonary cryptococcosis due to a capsule-deficient strain confused with metastatic lung cancer. Mycopathologia 2007, 140, 65–68. [Google Scholar] [CrossRef]
- Laurenson, I.; Ross, J.; Milne, L. Microscopy and latex antigen negative cryptococcal meningitis. J. Infect. 1998, 36, 329–331. [Google Scholar] [CrossRef]
- Moser, S.; Friedman, L.; Varraux, A. Atypical isolate of Cryptococcus neoformans cultured from sputum of a patient with pulmonary cancer and blastomycosis. J. Clin. Microbiol. 1978, 7, 316–318. [Google Scholar] [PubMed]
- Ro, J.; Lee, S.; Ayala, A. Advantage of Fontana-Masson stains in capsule-deficient cryptococcal infection. Arch. Pathol. Lab. Med. 1987, 111, 53–57. [Google Scholar] [PubMed]
- Harding, S.; Scheld, W.; Feldman, P.; Sande, M. Pulmonary infection with capsule-deficient Cryptococcus neoformans. Virchows Arch. A Pathol. Anat. Histol. 1979, 382, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Salkowski, C.; Balish, E. Susceptibility of congenitally immunodeficient mice to a nonencapsulated strain of Cryptococcus neoformans. Can. J. Microbiol. 1991, 37, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Sabiiti, W.; Robertson, E.; Beale, M.; Johnston, S.A.; Brouwer, A.E.; Loyse, A.; Jarvis, J.N.; Gilbert, A.S.; Fisher, M.C.; Harrison, T.S.; et al. Efficient phagocytosis and laccase activity affect the outcome of HIV-associated cryptococcosis. J. Clin. Investig. 2014, 124, 2000–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouklas, T.; Navarro, E.; Wang, X.; Fenster, M.; Fries, B.C. Characterization of the virulence of Cryptococcus neoformans strains in an insect model. Virulence 2015, 6, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Beale, A.; Sabiiti, W.; Robertson, E.; Fuentes-Cabrejo, K.M.; O’Hanlon, S.J.; Jarvis, J.N.; Loyse, A.; Meintjes, G.; Harrison, T.S.; May, R.C.; et al. Genotypic diversity is associated with clinical outcome and phenotype in cryptococcal meningitis across Southern Africa. PLoS Negl. Trop. Dis. 2015, 9, e0003847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litvintseva, A.; Mitchell, T. Most environmental isolates of Cryptococcus neoformans var. grubii (Serotype A) are not lethal for mice. Infect. Immun. 2009, 77, 3188–3195. [Google Scholar] [CrossRef] [PubMed]
- Trevijano-Contador, N.; de Oliveira, H.C.; García-Rodas, R.; Rossi, S.A.; Llorente, I.; Zaballos, Á.; Janbob, G.; Ariño, J.; Zaragoza, Ó. Cryptococcus neoformans can form titan-like cells in vitro in response to multiple signals. PLoS Pathog. 2018, 14, e1007007. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species Complex | ST | Strain (HOO58-I-) | Colony Morphology | Cellular and Capsular Size in µm | ||||
|---|---|---|---|---|---|---|---|---|
| Texture | Appearance | Pre-Inoculation | Post-Inocultion | |||||
| Cellular | Capsular | Cellular | Capsular | |||||
| C. neoformans | Clinical | |||||||
| 2 | 3746 | Mucoid | Smooth | 7.53 | 0.21 | 0.52 | 4.49 | |
| 2 | 3852 | 5.34 | 0.48 | 1.60 | 6.98 | |||
| 5 | 2881 | 5.57 | 0.52 | 1.76 | 4.58 | |||
| 6 | 3463 | 5.68 | 0.50 | 2.88 | 4.05 | |||
| 25 | 3104 | 5.49 | 0.75 | 0.49 | 6.14 | |||
| 32 | 2340 | Non-mucoid | 5.87 | 0.48 | 0.56 | 5.42 | ||
| 40 | 3589 | Mucoid | 6.20 | 0.48 | 1.63 | 4.40 | ||
| 63 | 2503 | 5.75 | 0.52 | 0.87 | 4.48 | |||
| 69 | 3099 | 5.87 | 0.45 | 1.81 | 5.96 | |||
| 71 | 3489 | 3.30 | 0.34 | 1.60 | 5.22 | |||
| 77 | 708 | 6.71 | 1.14 | 1.03 | 6.40 | |||
| 77 | 3845 | 5.72 | 0.67 | 1.60 | 4.35 | |||
| 93 | 2624 | Non-mucoid | Wrinkled | 6.53 | 0.45 | 1.61 | 11.88 | |
| 93 | 3189 | Mucoid | Smooth | 6.72 | 0.59 | 1.25 | 11.51 | |
| 93 | 995 | 5.54 | 0.59 | 3.08 | 9.18 | |||
| 93 | 1226 | 5.99 | 0.57 | 0.86 | 6.39 | |||
| 93 | 2073 | 4.73 | 1.22 | 1.02 | 7.98 | |||
| 93 | 2356 | Non-mucoid | 6.09 | 0.55 | 1.10 | 6.84 | ||
| 93 | 3938 | Mucoid | 6.15 | 0.48 | 0.90 | 4.40 | ||
| 199 | 714 | 5.90 | 0.93 | 3.42 | 9.78 | |||
| 307 | 707 | 5.85 | 0.92 | 2.55 | 7.97 | |||
| 307 | 727 | 5.44 | 0.83 | 1.32 | 8.00 | |||
| 307 | 2087 | 6.19 | 0.85 | 3.24 | 10.33 | |||
| 307 | 2274 | 6.12 | 0.89 | 2.25 | 9.16 | |||
| Environmental | ||||||||
| 15 | 4419 | Mucoid | Smooth | 5.69 | 0.65 | 1.12 | 6.04 | |
| 23 | 4706 | 4.29 | 0.25 | 1.34 | 6.80 | |||
| 56 | 4630 | 5.33 | 0.36 | 2.56 | 4.85 | |||
| 77 | 4013 | 5.74 | 0.67 | 2.05 | 4.37 | |||
| 93 | 3877 | 6.12 | 0.50 | 0.91 | 4.23 | |||
| 93 | 4711 | 6.39 | 0.59 | 0.91 | 4.35 | |||
| 226 | 5353 | 1.70 | 0.29 | 0.91 | 4.35 | |||
| C. gattii | Clinical | |||||||
| 25 | 212 | Non-mucoid | Smooth | 4.49 | 0.68 | 11.10 | 2.25 | |
| 47 | 255 | Mucoid | 5.40 | 2.88 | 9.40 | 1.85 | ||
| 85 | 792 | 3.57 | 0.66 | 8.90 | 2.21 | |||
| 106 | 1510 | 6.93 | 2.76 | 7.60 | 0.90 | |||
| 25 | 2877 | 5.74 | 0.46 | 5.53 | 1.20 | |||
| 51 | 3286 | 5.64 | 0.33 | 4.27 | 2.10 | |||
| 58 | 3031 | 4.98 | 0.47 | 6.07 | 1.10 | |||
| 323 | 3146 | 6.45 | 0.83 | 6.69 | 6.10 | |||
| 324 | 3407 | 4.40 | 1.39 | 4.29 | 1.40 | |||
| Environmental | ||||||||
| 25 | 3526 | Mucoid | Smooth | 3.08 | 0.25 | 5.69 | 1.20 | |
| 75 | 3593 | 2.45 | 0.65 | 5.15 | 1.90 | |||
| 79 | 4064 | 1.86 | 0.29 | 4.26 | 1.60 | |||
| 79 | 3080 | 1.89 | 0.25 | 4.30 | 1.60 | |||
| 79 | 3874 | 3.09 | 0.29 | 4.28 | 1.50 | |||
| Species Complex | Isolates | In Galleria mellonella | Mean Survival Days | |||||
|---|---|---|---|---|---|---|---|---|
| ST | Clinical | Environmental | Pre-Inoculation | Post-Inoculation | ||||
| Mean Cellular Size | Mean Capsular Size | Mean Cellular Size | Mean Capsular Size | |||||
| µm | ||||||||
| C. neoformans | 93 | 7 | 2 | 6.03 | 0.62 | 7.42 | 1.29 | 3.67 |
| 307 | 4 | 0 | 5.90 | 0.87 | 8.87 | 2.34 | 3.25 | |
| C. gattii | 25 | 2 | 1 | 4.44 | 0.46 | 7.43 | 1.55 | 4.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velez, N.; Alvarado, M.; Parra-Giraldo, C.M.; Sánchez-Quitian, Z.A.; Escandón, P.; Castañeda, E. Genotypic Diversity Is Independent of Pathogenicity in Colombian Strains of Cryptococcus neoformans and Cryptococcus gattii in Galleria mellonella. J. Fungi 2018, 4, 82. https://doi.org/10.3390/jof4030082
Velez N, Alvarado M, Parra-Giraldo CM, Sánchez-Quitian ZA, Escandón P, Castañeda E. Genotypic Diversity Is Independent of Pathogenicity in Colombian Strains of Cryptococcus neoformans and Cryptococcus gattii in Galleria mellonella. Journal of Fungi. 2018; 4(3):82. https://doi.org/10.3390/jof4030082
Chicago/Turabian StyleVelez, Norida, Maira Alvarado, Claudia Marcela Parra-Giraldo, Zilpa Adriana Sánchez-Quitian, Patricia Escandón, and Elizabeth Castañeda. 2018. "Genotypic Diversity Is Independent of Pathogenicity in Colombian Strains of Cryptococcus neoformans and Cryptococcus gattii in Galleria mellonella" Journal of Fungi 4, no. 3: 82. https://doi.org/10.3390/jof4030082