Treatment of Infections Due to Aspergillus terreus Species Complex


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Pathogen and Fungal Epidemiology
3. Patients at Risk Suffering from Aspergillus terreus Species Complex
4. Treatment Recommendations
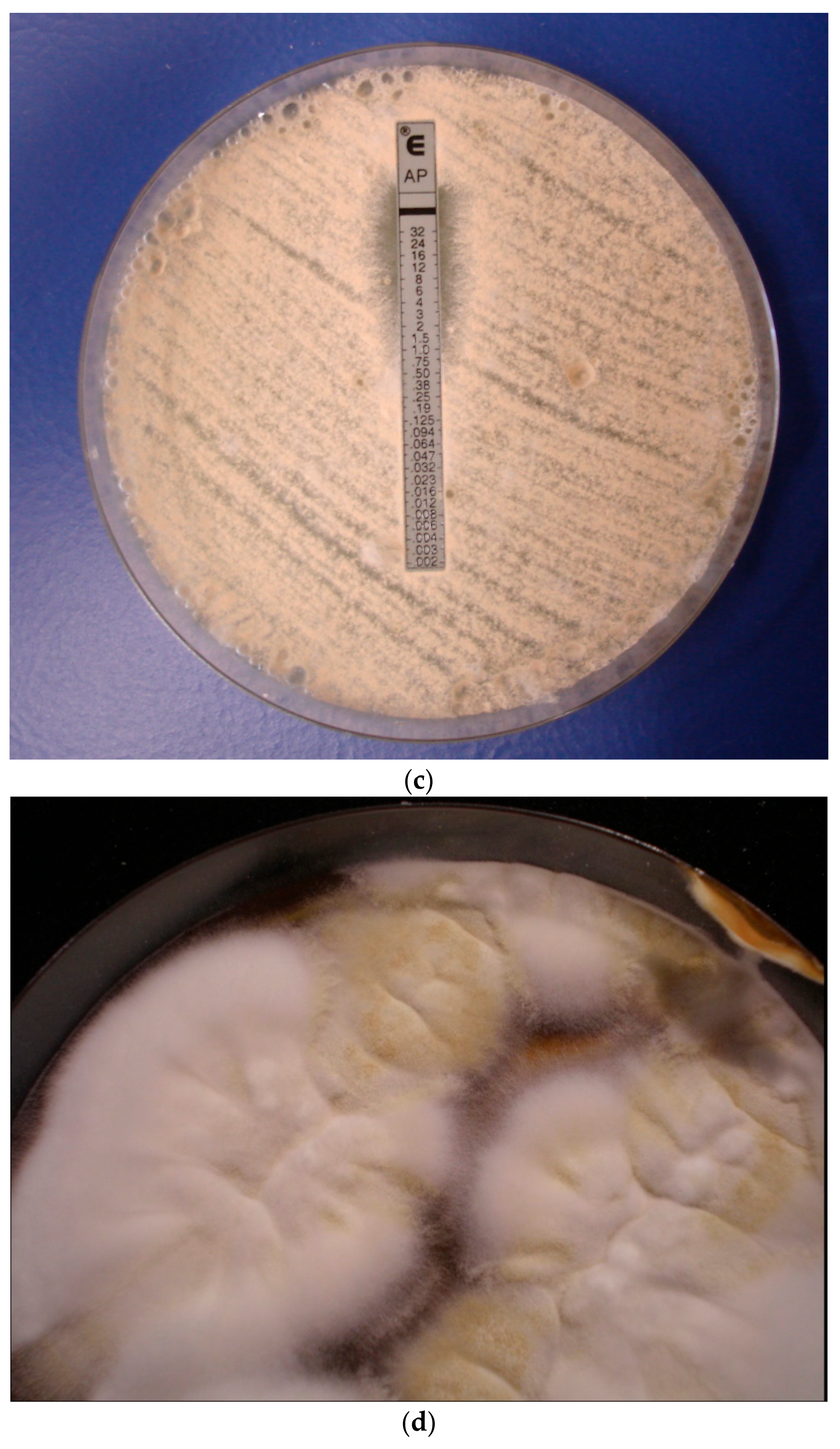
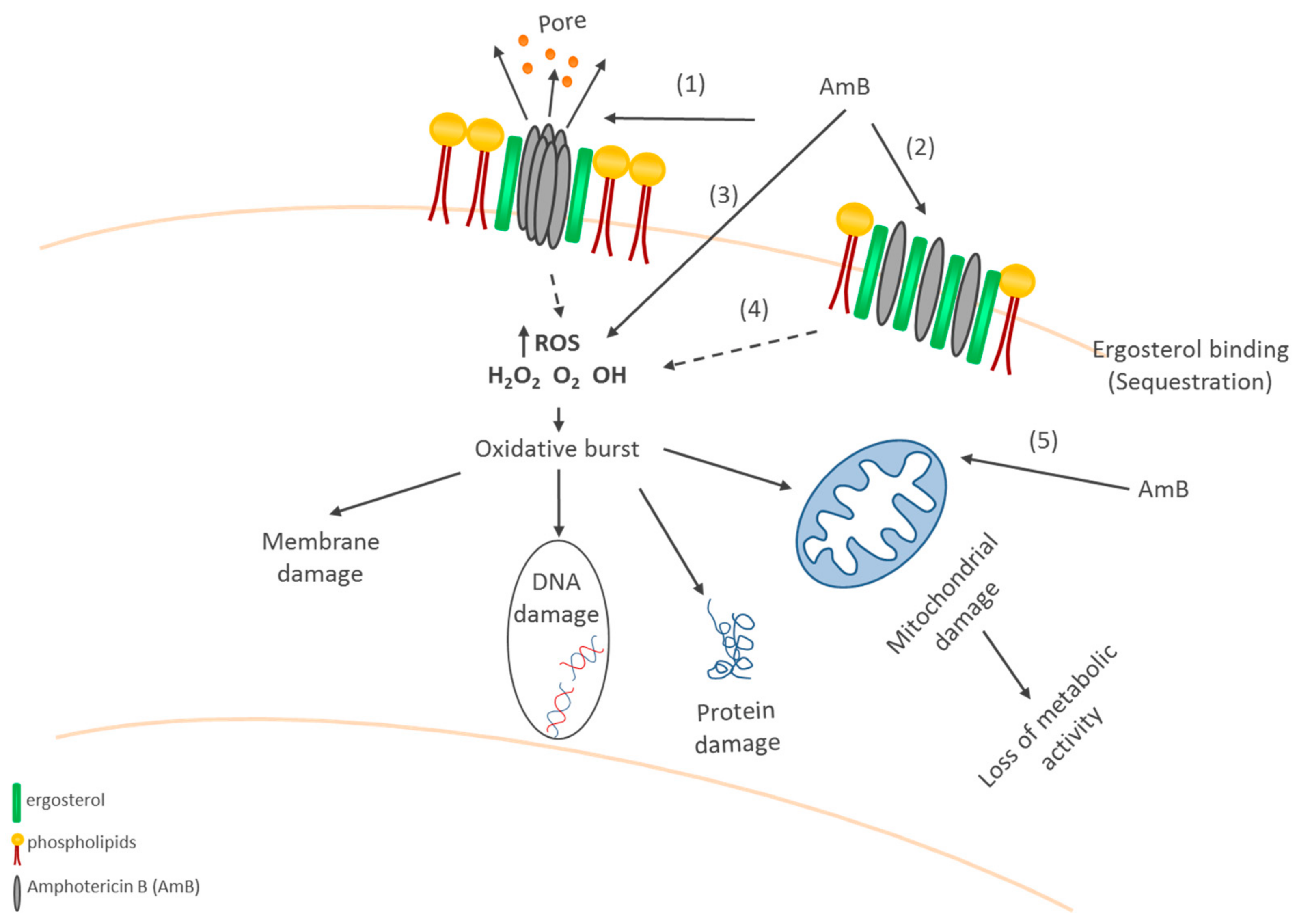
5. Amphotericin B
6. Azoles
7. Conclusions
Funding
Conflicts of Interest
References
- Iwen, P.C.; Rupp, M.E.; Langnas, A.N.; Reed, E.C.; Hinrichs, S.H. Invasive pulmonary aspergillosis due to Aspergillus terreus: 12-year experience and review of the literature. Clin. Infect. Dis. 1998, 26, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Peterson, S.W.; Friswad, J.C.; Varga, J. New species in Aspergillus section Terrei. Stud. Mycol. 2011, 69, 39–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lass-Flörl, C.; Grif, K.; Kontoyiannis, D.P. Molecular typing of Aspergillus terreus isolates collected in Houston, Texas, and Innsbruck, Austria: Evidence of great genetic diversity. J. Clin. Microbiol. 2007, 45, 2686–2690. [Google Scholar] [CrossRef] [PubMed]
- Blum, G.; Perkhofer, S.; Haas, H.; Schrettl, M.; Würzner, R.; Dierich, M.P.; Lass-Flörl, C. Potential basis for amphotericin B resistance in Aspergillus terreus. Antimicrob. Agents Chemother. 2008, 52, 1553–1555. [Google Scholar] [CrossRef] [PubMed]
- Pastor, F.J.; Guarro, J. Treatment of Aspergillus terreus infections: A clinical problem not yet solved. Int. J. Antimicrob. Agents 2014, 44, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C.; Griff, K.; Mayr, A.; Petzer, A.; Gastl, G.; Bonatti, H.; Freund, M.; Kropshofer, G.; Dierich, M.P.; Nachbaur, D. Epidemiology and outcome of infections due to Aspergillus terreus: 10-year single centre experience. Br. J. Haematol. 2005, 131, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Deak, E.; Nelson, M.; Hernández-Rodríguez, Y.; Gade, L.; Baddley, J.; Momany, M.; Steele, C.; Balajee, S.A. Aspergillus terreus accessory conidia are multinucleated, hyperpolarizing structures that display differential dectin staining and can induce heightened inflammatory responses in a pulmonary model of aspergillosis. Virulence 2011, 2, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Risslegger, B.; Zoran, T.; Lackner, M.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. A prospective international Aspergillus terreus survey: An EFISG, ISHAM and ECMM joint study. Clin. Microbiol. Infect. 2017, 23, 776.e1–776.e5. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M.; Coassin, S.; Haun, M.; Binder, U.; Kronenberg, F.; Haas, H.; Jank, M.; Maurer, E.; Meis, J.F.; Hagen, F.; et al. Geographically predominant genotypes of Aspergillus terreus species complex in Austria: A microsatellite typing study. Clin. Microbiol. Infect. 2016, 22, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Hachem, R.; Ribeiro Gomes, M.Z.; el Helou, G.; el Zakhem, A.; Kassis, C.; Ramos, E.; Jiang, Y.; Chaftari, A.; Raad, I.I. Invasive aspergillosis caused by Aspergillus terreus: An emerging opportunistic infection with poor outcome independent of azole therapy. J. Antimicrob. Chemother. 2014, 69, 3148–3155. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of Aspergillus diseases: Executive summary of the 2017 ESCMID-ECMM-ERS guideline. Clin. Microbiol. Infect. 2018, 24, e1–e38. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Marr, K.A.; Anaissie, E.J.; Azie, N.; Quan, S.; Meier-Kriesche, H.; Apewokin, S.; Horn, D.L. Clinical epidemiology of 960 patients with invasive aspergillosis from the PATH Alliance registry. J. Infect. 2012, 65, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Albert, N.P.; Liao, G.; Wan, W.; Prince, R.A.; Kontoyiannis, D.P. High-dose induction liposomal amphotericin B followed by de-escalation is effective in experimental Aspergillus terreus pneumonia. J. Antimicrob. Chemother. 2013, 68, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.J.; Petraitis, V.; Petraitiene, R.; Field-Ridley, A.; Sutton, D.; Ghannoum, M.; Sein, T.; Schaufele, R.; Peter, J.; Bacher, J.; et al. Experimental pulmonary aspergillosis due to Aspergillus terreus: Pathogenesis and treatment of an emerging fungal pathogen resistant to amphotericin B. J. Infect. Dis. 2003, 188, 305–319. [Google Scholar] [CrossRef] [PubMed]
- Graybill, J.R.; Hernandez, S.; Bocanegra, R.; Najvar, L.K. Antifungal therapy of murine Aspergillus terreus infections. Antimicrob. Agents Chemother. 2004, 48, 3715–3719. [Google Scholar] [CrossRef] [PubMed]
- Blum, G.; Hörtnagl, C.; Jukic, E.; Erbeznik, T.; Pümpel, T.; Dietrich, H.; Nagl, M.; Speth, C.; Rambach, G.; Lass-Flörl, C. New insights into amphotericin B resistance in Aspergillus terreus. Antimicrob. Agents Chemother. 2013, 57, 1538–1588. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Diekema, D.J.; Fothergill, A.; Johnson, E.; Pelaez, T.; Pfaller, M.A.; Rinaldi, M.G.; Canton, E.; Turnidge, J. Wild-type MIC distributions and epidemiological cutoff values for the triazoles and six Aspergillus spp. for the CLSI broth microdilution method (M38-A2 document). J. Clin. Microbiol. 2010, 48, 3251–3257. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Benjamin, D.K.; Kontoyiannis, D.P.; Perfect, J.R.; Lutsar, I.; Marr, K.A.; Lionakis, M.S.; Torres, H.A.; Jafri, H.; Walsh, T.J. Infections due to Aspergillus terreus: A multicenter retrospective analysis of 83 cases. Clin. Infect. Dis. 2004, 39, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing. Antifungal Susceptibility Testing (AFST). EUCAST, 2012. Available online: http://www.eucast.org/ (accessed on 27 March 2018).
- Blatzer, M.; Blum, G.; Jukic, E.; Posch, W.; Gruber, P.; Nagl, M.; Binder, U.; Maurer, E.; Sarg, B.; Lindner, H.; et al. Blocking Hsp70 enhances the efficiency of amphotericin B treatment against resistant Aspergillus terreus strains. Antimicrob. Agents Chemother. 2015, 59, 3778–3788. [Google Scholar] [CrossRef] [PubMed]
- Blatzer, M.; Jukic, E.; Posch, W.; Schöpf, B.; Binder, U.; Steger, M.; Blum, G.; Hackl, H.; Gnaiger, E.; Cornelia, L.F.; et al. Amphotericin B resistance in Aspergillus terreus is overpowered by coapplication of pro-oxidants. Antioxid. Redox Signal. 2015, 23, 1424–1438. [Google Scholar] [CrossRef] [PubMed]
- Posch, W.; Blatzer, M.; Wilflingseder, D.; Lass-Flörl, C. Aspergillus terreus: Novel lessons learned on amphotericin B resistance. Med. Mycol. 2018, 56, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Maurer, E.; Lackner, M.; Lass-Flörl, C. Effect of reduced oxygen on the antifungal susceptibility of clinically relevant aspergilli. Antimicrob. Agents Chemother. 2015, 59, 1806–1810. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Jukic, E.; Blatzer, M.; Binder, U.; Mayr, L.; Lass-Flörl, C.; Lackner, M. Impact of morphological sectors on antifungal susceptibility testing and virulence studies. Antimicrob. Agents Chemother. 2017, 61, e00755-17. [Google Scholar] [CrossRef] [PubMed]
- Trifilio, S.; Ortiz, R.; Pennick, G.; Verma, A.; Pi, J.; Stosor, V.; Zembower, T.; Mehta, J. Voriconazole therapeutic drug monitoring in allogeneic hematopoietic stem cell transplant recipients. Bone Marrow Transplant. 2005, 35, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, V.; Pastor, F.J.; Sutton, D.A.; Calvo, E.; Mayayo, E.; Fothergill, A.W.; Rinaldi, M.G.; Guarro, J. MIC values of voriconazole are predictive of treatment results in murine infections by Aspergillus terreus species complex. Antimicrob. Agents Chemother. 2013, 57, 1532–1534. [Google Scholar] [CrossRef] [PubMed]
- Risslegger, B.; Zoran, T.; Lackner, M.; Aigner, M.; Sánchez-Reus, F.; Rezusta, A.; Chowdhary, A.; Taj-Aldeen, S.J.; Arendrup, M.C.; Oliveri, S.; et al. Azole-resistance in Aspergillus terreus and related species: An emerging problem or a rare phenomenon? Front. Micorbiol. 2018, 9, 516. [Google Scholar]
- Arendrup, M.C.; Jensen, R.H.; Grif, K.; Skov, M.; Pressler, T.; Johansen, H.K.; Lass-Flörl, C. In vivo emergence of Aspergillus terreus with reduced azole susceptibility and a Cyp51a M217l alteration. J. Infect. Dis. 2012, 206, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Azole-resistant aspergillosis: Epidemiology, molecular mechanisms, and treatment. J. Infect. Dis. 2017, 216, S436–S444. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lass-Flörl, C. Treatment of Infections Due to Aspergillus terreus Species Complex. J. Fungi 2018, 4, 83. https://doi.org/10.3390/jof4030083
Lass-Flörl C. Treatment of Infections Due to Aspergillus terreus Species Complex. Journal of Fungi. 2018; 4(3):83. https://doi.org/10.3390/jof4030083
Chicago/Turabian StyleLass-Flörl, Cornelia. 2018. "Treatment of Infections Due to Aspergillus terreus Species Complex" Journal of Fungi 4, no. 3: 83. https://doi.org/10.3390/jof4030083
APA StyleLass-Flörl, C. (2018). Treatment of Infections Due to Aspergillus terreus Species Complex. Journal of Fungi, 4(3), 83. https://doi.org/10.3390/jof4030083