Inter-Specimen Imbalance of Mitochondrial Gene Copy Numbers Predicts Clustering of Pneumocystis jirovecii Isolates in Distinct Subgroups


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Samples
2.3. Magnetic Labelling and Separation
2.4. DNA/RNA Extraction
2.5. Quantitative PCR
2.6. Pneumogenius® Assay
2.7. Statistical Analyses
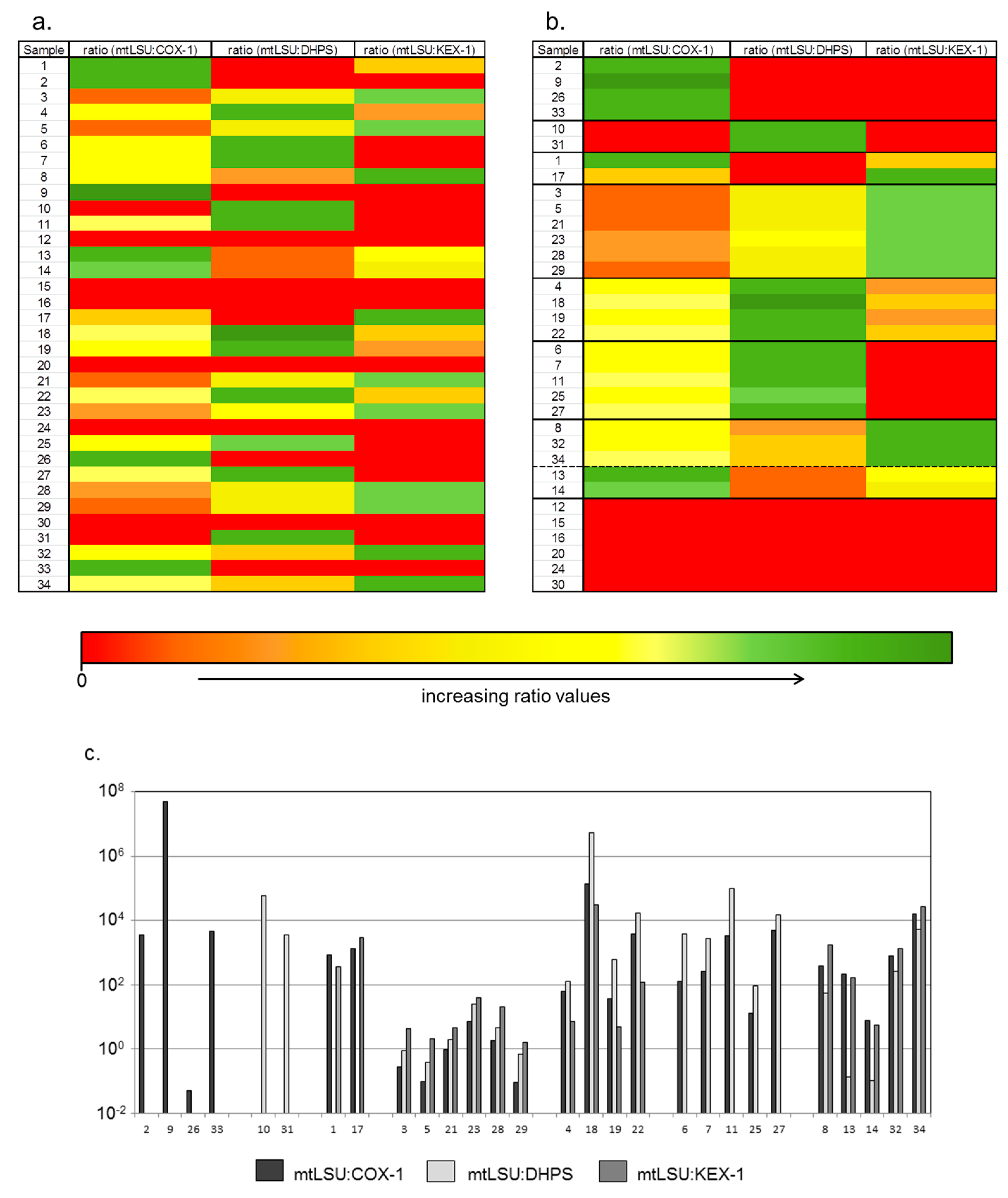
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
| Sample No. | mtLSU (Copies/mL) | COX1 (Copies/mL) | KEX1 (Copies/ mL) | DHPS (Copies/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| Pre-Separation | Post-Separation | Pre-Purification | Post-Purification | Pre-Purification | Post-Purification | Pre-Purification | Post-Purification | |
| 1 | 4.33 × 108 | negative | 5.22 × 105 | negative | 1.21 × 106 | negative | negative | negative |
| 2 | 2.16 × 107 | negative | 5.93 × 103 | negative | negative | negative | negative | negative |
| 3 | 9.02 × 108 | 1.30 × 107 | 3.33 × 109 | 3.14 × 108 | 2.08 × 108 | 2.19 × 109 | 9.97 × 108 | 7.04 × 107 |
| 4 | 9.52 × 107 | 3.30 × 104 | 1.53 × 106 | 1.82 × 105 | 1.30 × 107 | negative | 7.41 × 105 | negative |
| 5 | 7.99 × 109 | 2.20 × 105 | 8.12 × 1010 | 5.18 × 107 | 3.77 × 109 | 1.40 × 108 | 2.04 × 1010 | 9.44 × 106 |
| 6 | 1.99 × 109 | 8.20 × 104 | 1.56 × 107 | 1.14 × 106 | n/a | negative | 5.21 × 105 | 2.73 × 101 |
| 7 | 3.50 × 107 | negative | 1.36 × 105 | negative | n/a | negative | 1.30 × 104 | 4.22 × 103 |
| 8 | 2.62 × 107 | negative | 6.56 × 104 | 9.51 × 104 | 1.47 × 104 | negative | 4.71 × 105 | negative |
| 9 | 2.33 × 109 | 6.20 × 104 | 4.79 × 101 | negative | negative | negative | negative | negative |
| 10 | 1.47 × 108 | negative | negative | negative | negative | negative | 2.54 × 103 | negative |
| 11 | 2.11 × 109 | 1.70 × 1011 | 6.26 × 105 | 1.89 × 105 | negative | negative | 2.16 × 104 | negative |
| 12 | 5.79 × 108 | negative | negative | negative | negative | negative | negative | negative |
| 13 | 3.32 × 1012 | n/a | 1.49 × 1010 | n/a | 2.05 × 1010 | n/a | 2.52 × 1013 | n/a |
| 14 | 1.03 × 1010 | 1.30 × 107 | 1.37 × 109 | 1.75 × 109 | 1.84 × 109 | 8.41 × 109 | 1.00 × 1011 | 4.04 × 109 |
| 15 | 1.13 × 108 | negative | negative | negative | negative | 5.71 × 107 | negative | negative |
| 16 | 4.33 × 109 | negative | negative | negative | negative | negative | negative | negative |
| 17 | 5.99 × 108 | 3.60 × 1011 | 4.52 × 105 | 2.90 × 104 | 2.06 × 105 | negative | negative | negative |
| 18 | 1.61 × 1014 | n/a | 1.14 × 109 | n/a | 5.04 × 109 | n/a | 3.04 × 107 | n/a |
| 19 | 1.59 × 1010 | 1.40 × 106 | 4.20 × 108 | 2.22 × 108 | 3.24 × 109 | 9.40 × 108 | 2.59 × 107 | 2.25 × 108 |
| 20 | 2.70 × 107 | 2.10 × 102 | negative | negative | negative | negative | negative | negative |
| 21 | 2.98 × 1011 | 8.60 × 106 | 3.05 × 1011 | 1.03 × 109 | 6.45 × 1010 | 9.55 × 109 | 1.53 × 1011 | 7.11 × 109 |
| 22 | 1.05 × 1012 | 1.90 × 106 | 2.74 × 108 | 1.96 × 108 | 8.51 × 109 | 9.07 × 108 | 6.29 × 107 | 3.14 × 108 |
| 23 | 1.01 × 1011 | 3.50 × 107 | 1.44 × 1010 | 4.82 × 108 | 2.52 × 109 | 5.89 × 108 | 3.91 × 109 | 3.47 × 106 |
| 24 | 9.92 × 108 | 4.00 × 1014 | negative | negative | negative | negative | negative | 1.21 × 104 |
| 25 | 2.11 × 107 | 3.70 × 105 | 1.67 × 106 | 8.37 × 104 | negative | negative | 2.34 × 105 | negative |
| 26 | 1.09 × 1010 | 6.50 × 108 | 2.17 × 1011 | 1.05 × 1010 | n/a | 5.64 × 1010 | n/a | 2.25 × 109 |
| 27 | 1.65 × 108 | negative | 3.24 × 104 | negative | negative | negative | 1.11 × 104 | 9.39 × 103 |
| 28 | 1.07 × 108 | n/a | 5.92 × 107 | n/a | 5.23 × 106 | n/a | 2.39 × 107 | n/a |
| 29 | 1.04 × 109 | n/a | 1.17 × 1010 | n/a | 6.56 × 108 | n/a | 1.55 × 109 | n/a |
| 30 | 2.88 × 107 | 5.70 × 103 | negative | negative | negative | negative | negative | 4.66 × 103 |
| 31 | 1.79 × 107 | n/a | negative | n/a | negative | n/a | 4.83 × 103 | n/a |
| 32 | 9.90 × 108 | negative | 1.21 × 106 | negative | 7.41 × 105 | negative | 3.79 × 106 | negative |
| 33 | 2.80 × 108 | negative | 6.20 × 104 | negative | negative | negative | negative | 2.08 × 104 |
| 34 | 1.98 × 1010 | 6.20 × 105 | 1.21 × 106 | 5.00 × 106 | 7.41 × 105 | 2.66 × 107 | 3.79 × 106 | 2.85 × 106 |
References
- Chagas, C. Nova Tripanosomiae Humana; Memorias do Instituto Oswaldo Cruz: Rio de Janeiro, Brazil, 1909; pp. 159–218. [Google Scholar]
- Delanoe, P. Sur les rapports des kystes de Carini du poumon des rats avec le Trypanosoma lewisi. CR Acad. Sci. Paris 1912, 155, 658–661. (In French) [Google Scholar]
- Dvorak, J.; Jirovec, O. Diagnosis and therapy of atypic parasitic pneumonia in children caused by Pneumocystis carinii. Pediatr. Listy 1952, 7, 264–267. [Google Scholar] [PubMed]
- Masur, H.; Michelis, M.A.; Greene, J.B.; Onorato, I.; Stouwe, R.A.; Holzman, R.S.; Wormser, G.; Brettman, L.; Lange, M.; Murray, H.W.; et al. An outbreak of community-acquired Pneumocystis carinii pneumonia: Initial manifestation of cellular immune dysfunction. N. Engl. J. Med. 1981, 305, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Pixley, F.J.; Wakefield, A.E.; Banerji, S.; Hopkin, J.M. Mitochondrial gene sequences show fungal homology for Pneumocystis carinii. Mol. Microbiol. 1991, 5, 1347–1351. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.C.; Kovacs, J.A.; Masur, H.; Santi, D.V.; Elwood, H.J.; Sogin, M.L. Ribosomal RNA sequence shows Pneumocystis carinii to be a member of the fungi. Nature 1988, 334, 519. [Google Scholar] [CrossRef] [PubMed]
- Stringer, S.L.; Hudson, K.; Blase, M.A.; Walzer, P.D.; Cushion, M.T.; Stringer, J.R. Sequence from ribosomal RNA of Pneumocystis carinii compared to those of four fungi suggests an ascomycetous affinity. J. Protozool. 1989, 36. [Google Scholar] [CrossRef]
- Stringer, J.R.; Cushion, M.T.; Wakefield, A.E. New nomenclature for the genus Pneumocystis. J. Eukaryot. Microbiol. 2001. [Google Scholar] [CrossRef]
- Helweg-Larsen, J.; Lundgren, B.; Lundgren, J.D. Heterogeneity and compartmentalization of Pneumocystis carinii f. sp. hominis genotypes in autopsy lungs. J. Clin. Microbiol. 2001, 39, 3789–3792. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Mansharamani, N.G.; Garland, R.; Delaney, D.; Koziel, H. Management and outcome patterns for adult Pneumocystis carinii pneumonia, 1985 to 1995: Comparison of HIV-associated cases to other immunocompromised states. Chest 2000, 118, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.A.; Morris, A. Pneumocystis infection and the pathogenesis of chronic obstructive pulmonary disease. Immunol. Res. 2011, 50, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Sing, A.; Trebesius, K.; Roggenkamp, A.; Russmann, H.; Tybus, K.; Pfaff, F.; Bogner, J.R.; Emminger, C.; Heesemann, J. Evaluation of diagnostic value and epidemiological implications of PCR for Pneumocystis carinii in different immunosuppressed and immunocompetent patient groups. J. Clin. Microbiol. 2000, 38, 1461–1467. [Google Scholar] [PubMed]
- Botterel, F.; Cabaret, O.; Foulet, F.; Cordonnier, C.; Costa, J.M.; Bretagne, S. Clinical significance of quantifying Pneumocystis jirovecii DNA by using real-time PCR in bronchoalveolar lavage fluid from immunocompromised patients. J. Clin. Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Larsen, H.H.; Masur, H.; Kovacs, J.A.; Gill, V.J.; Silcott, V.A.; Kogulan, P.; Maenza, J.; Smith, M.; Lucey, D.R.; Fischer, S.H. Development and evaluation of a quantitative, touch-down, real-time PCR assay for diagnosing Pneumocystis carinii pneumonia. J. Clin. Microbiol. 2002, 40, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Damiani, C.; Le Gal, S.; Da Costa, C.; Virmaux, M.; Nevez, G.; Totet, A. Combined quantification of pulmonary Pneumocystis jirovecii DNA and serum (1→3)-β-d-glucan for differential diagnosis of Pneumocystis pneumonia and Pneumocystis colonization. J. Clin. Microbiol. 2013, 51. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Desoubeaux, G.; Sarfati, C.; Hamane, S.; Bergeron, A.; Azoulay, E.; Molina, J.M.; Derouin, F.; Menotti, J. Real-time PCR assay-based strategy for differentiation between active Pneumocystis jirovecii pneumonia and colonization in immunocompromised patients. Clin. Microbiol. Infect. 2011, 17, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Prickartz, A.; Lüsebrink, J.; Khalfaoui, S.; Schildgen, O.; Schildgen, V.; Windisch, W.; Brockmann, M. Low Titer Pneumocystis jirovecii Infections: More than Just Colonization? J. Fungi 2016, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Kessl, J.J.; Hill, P.; Lange, B.B.; Meshnick, S.R.; Meunier, B.; Trumpower, B.L. Molecular basis for atovaquone resistance in Pneumocystis jirovecii modeled in the cytochrome bc1 complex of Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 2817–2824. [Google Scholar] [CrossRef] [PubMed]
- Kutty, G.; Kovacs, J.A. A single-copy gene encodes Kex1, a serine endoprotease of Pneumocystis jiroveci. Infect. Immun. 2003, 71, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Russian, D.A.; Andrawis-Sorial, V.; Goheen, M.P.; Edman, J.C.; Vogel, P.; Turner, R.E.; Klivington, D.L.; Angus, C.W.; Kovacs, J.A. Characterization of a multicopy family of genes encoding a surface-expressed serine endoprotease in rat Pneumocystis carinii. Proc. Assoc. Am. Physicians 1999, 111, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Schildgen, V.; Wulfert, M.; Gattermann, N. Impaired mitochondrial gene transcription in myelodysplastic syndromes and acute myeloid leukemia with myelodysplasia-related changes. Exp. Hematol. 2011, 39, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Fauchier, T.; Hasseine, L.; Gari-Toussaint, M.; Casanova, V.; Marty, P.M.; Pomares, C. Detection of Pneumocystis jirovecii by Quantitative PCR To Differentiate Colonization and Pneumonia in Immunocompromised HIV-Positive and HIV-Negative Patients. J. Clin. Microbiol. 2016, 54, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Togashi, Y.; Masago, K.; Ito, Y.; Sakamori, Y.; Okuda, C.; Fukuhara, A.; Nagai, H.; Kim, Y.H.; Mishima, M. Pneumocystis jiroveci pneumonia and colonization in patients with advanced lung cancer. Oncol. Lett. 2013, 5, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Eddens, T.; Kolls, J.K. Lung pathology associated with Pneumocystis colonization in infants. Clin. Infect. Dis. 2013, 56, 180–181. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Olivi, M.; Cabaret, O.; Foulet, F.; Bellanger, A.P.; Millon, L.; Berceanu, A.; Cordonnier, C.; Costa, J.M.; Bretagne, S. Correlation Between Pneumocystis jirovecii Mitochondrial Genotypes and High and Low Fungal Loads Assessed by Single Nucleotide Primer Extension Assay and Quantitative Real-Time PCR. J. Eukaryot. Microbiol. 2015, 62, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Ronit, A.; Klitbo, D.M.; Kildemoes, A.O.; Benfield, T.; Gerstoft, J.; Vestbo, J.; Jensen, J.S.; Kurtzhals, J.; Nielsen, S.D. Absence of Pneumocystis jirovecii Colonization in Human Immunodeficiency Virus-Infected Individuals With and Without Airway Obstruction and With Undetectable Viral Load. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2016; Volume 3. [Google Scholar]
- Schildgen, V.; Mai, S.; Khalfaoui, S.; Lusebrink, J.; Pieper, M.; Tillmann, R.L.; Brockmann, M.; Schildgen, O. Pneumocystis jirovecii can be productively cultured in differentiated CuFi-8 airway cells. MBio 2014, 5, e01186-01114. [Google Scholar] [CrossRef] [PubMed]
- Valero, C.; Buitrago, M.J.; Gits-Muselli, M.; Benazra, M.; Sturny-Leclère, A.; Hamane, S.; Guigue, N.; Bretagne, S.; Alanio, A. Copy Number Variation of Mitochondrial DNA Genes in Pneumocystis jirovecii According to the Fungal Load in BAL Specimens. Front. Microbiol. 2016, 7, 1403. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Huang, D.W.; Cuomo, C.A.; Sykes, S.; Fantoni, G.; Das, B.; Sherman, B.T.; Yang, J.; Huber, C.; Xia, Y.; et al. Sequencing and characterization of the complete mitochondrial genomes of three Pneumocystis species provide new insights into divergence between human and rodent Pneumocystis. FASEB J. 2013, 27, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Keely, S.P.; Stringer, J.R. Molecular and phenotypic description of Pneumocystis wakefieldiae sp. nov., a new species in rats. Mycologia 2004, 96, 429–438. [Google Scholar] [CrossRef] [PubMed]

| mtLSU | PneumoGenius® | mtLSU | PneumoGenius® | ||||
|---|---|---|---|---|---|---|---|
| mtLSU | DHPS | mtLSU | DHPS | ||||
| 1 | 2.52 × 105 | + | + | 26 | 6.48 × 103 | + | ⌀ |
| 2 | 7.55 × 104 | + | + | 27 | 2.45 × 106 | + | + |
| 3 | 1.36 × 106 | + | ⌀ | 28 | 1.05 × 106 | + | + |
| 4 | 1.38 × 106 | + | ⌀ | 29 | 3.12 × 106 | + | + |
| 5 | 4.41 × 104 | ⌀ | + | 30 | 4.15 × 106 | + | + |
| 6 | 3.22 × 105 | + | ⌀ | 31 | 7.84 × 106 | + | + |
| 7 | 4.02 × 104 | + | + | 32 | 7.50 × 104 | ⌀ | ⌀ |
| 8 | 2.20 × 102 | + | ⌀ | 33 | 1.95 × 105 | + | + |
| 9 | 1.21 | + | ⌀ | 34 | 2.21 × 106 | + | + |
| 10 | 9.26 × 106 | + | + | 35 | 2.22 × 106 | + | + |
| 11 | 7.63 × 106 | + | + | 36 | 3.36 × 104 | + | + |
| 12 | 1.12 × 106 | + | ⌀ | 37 | 8.76 × 106 | + | + |
| 13 | 4.52 × 106 | + | + | 38 | 2.87 × 106 | + | + |
| 14 | 6.92 × 103 | + | + | 39 | 1.08 × 106 | + | + |
| 15 | 1.02 × 104 | + | + | 40 | 1.00 × 106 | ⌀ | + |
| 16 | 5.71 × 105 | + | ⌀ | 41 | 6.73 × 106 | + | ⌀ |
| 17 | 9.77 × 106 | + | + | 42 | 1.65 × 106 | ⌀ | + |
| 18 | 4.33 × 105 | + | + | 43 | 2.78 × 105 | + | + |
| 19 | 1.52 × 106 | + | + | 44 | 2.60 × 106 | + | + |
| 20 | 1.43 × 105 | + | + | 45 | 1.55 × 105 | ⌀ | + |
| 21 | 3.19 × 103 | ⌀ | ⌀ | 46 | 4.29 × 106 | + | ⌀ |
| 22 | 4.60 × 103 | + | + | 47 | 1.12 × 103 | + | + |
| 23 | 1.58 × 104 | ⌀ | + | 48 | 1.42 × 104 | ⌀ | + |
| 24 | 3.77 × 103 | + | + | 49 | 8.35 × 103 | + | ⌀ |
| 25 | 9.27 × 103 | + | ⌀ | 50 | 6.92 × 103 | + | ⌀ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunaiski, C.M.; Janssen, L.; Erzinger, H.; Pieper, M.; Damaschek, S.; Schildgen, O.; Schildgen, V. Inter-Specimen Imbalance of Mitochondrial Gene Copy Numbers Predicts Clustering of Pneumocystis jirovecii Isolates in Distinct Subgroups. J. Fungi 2018, 4, 84. https://doi.org/10.3390/jof4030084
Dunaiski CM, Janssen L, Erzinger H, Pieper M, Damaschek S, Schildgen O, Schildgen V. Inter-Specimen Imbalance of Mitochondrial Gene Copy Numbers Predicts Clustering of Pneumocystis jirovecii Isolates in Distinct Subgroups. Journal of Fungi. 2018; 4(3):84. https://doi.org/10.3390/jof4030084
Chicago/Turabian StyleDunaiski, Cara Mia, Lena Janssen, Hannah Erzinger, Monika Pieper, Sarah Damaschek, Oliver Schildgen, and Verena Schildgen. 2018. "Inter-Specimen Imbalance of Mitochondrial Gene Copy Numbers Predicts Clustering of Pneumocystis jirovecii Isolates in Distinct Subgroups" Journal of Fungi 4, no. 3: 84. https://doi.org/10.3390/jof4030084
APA StyleDunaiski, C. M., Janssen, L., Erzinger, H., Pieper, M., Damaschek, S., Schildgen, O., & Schildgen, V. (2018). Inter-Specimen Imbalance of Mitochondrial Gene Copy Numbers Predicts Clustering of Pneumocystis jirovecii Isolates in Distinct Subgroups. Journal of Fungi, 4(3), 84. https://doi.org/10.3390/jof4030084