
The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling


1
Department of Microbiology & Molecular Genetics, Michigan State University, East Lansing, MI 48824, USA
2
Sashi Bhusan Rath Government Autonomous Women’s College, Brahmapur 760001, India
3
Center for Plant Science Innovation, University of Nebraska-Lincoln, Lincoln, NE 68588, USA
4
Department of Plant, Soil and Microbial Science, Michigan State University, East Lansing, MI 48824, USA
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2024, 10(1), 13; https://doi.org/10.3390/ncrna10010013
Submission received: 11 December 2023
/
Revised: 5 February 2024
/
Accepted: 6 February 2024
/
Published: 7 February 2024
(This article belongs to the Special Issue Non-Coding RNA and Their Regulatory Roles in Plant)
Abstract
:Plant species utilize a variety of regulatory mechanisms to ensure sustainable productivity. Within this intricate framework, numerous non-coding RNAs (ncRNAs) play a crucial regulatory role in plant biology, surpassing the essential functions of RNA molecules as messengers, ribosomal, and transfer RNAs. ncRNAs represent an emerging class of regulators, operating directly in the form of small interfering RNAs (siRNAs), microRNAs (miRNAs), long noncoding RNAs (lncRNAs), and circular RNAs (circRNAs). These ncRNAs exert control at various levels, including transcription, post-transcription, translation, and epigenetic. Furthermore, they interact with each other, contributing to a variety of biological processes and mechanisms associated with stress resilience. This review primarily concentrates on the recent advancements in plant ncRNAs, delineating their functions in growth and development across various organs such as root, leaf, seed/endosperm, and seed nutrient development. Additionally, this review broadens its scope by examining the role of ncRNAs in response to environmental stresses such as drought, salt, flood, heat, and cold in plants. This compilation offers updated information and insights to guide the characterization of the potential functions of ncRNAs in plant growth, development, and stress resilience in future research.
Keywords:
non-coding RNA; root growth; leaf development; seed; endosperm; seed nutrient development; drought; salt; flood; heat; cold stress1. Introduction
Plants exhibit a highly intricate mechanism to synchronize various biological processes and adapt to a spectrum of stress conditions. The gene regulatory mechanism operates at multiple levels, including transcriptional, post-transcriptional, translational, and epigenetic, playing a pivotal role in growth, development, and stress-responsive signaling. This orchestration significantly contributes to the sustainable productivity of plants [1]. Advances in sequencing technologies, tissue-specific and single-cell RNA-sequencing, mass spectrometry-based proteomics, and bioinformatics tools have greatly shifted the focus from gene coding to non-coding elements, maintaining ‘cellular homeostasis’ amid ever-changing environmental conditions [2,3,4].
Conventionally, genomic DNA serves as the template for RNA transcription; however, this process does not necessarily result in protein production. The portion of the genome that does not contribute to a gene and hence does not translate into a protein is commonly referred to as “non-coding DNA,” also known as “experimental artifacts” or “junk DNA” [5]. Almost 90% of the genome acts as an RNA template, with only a minor portion coding for messenger RNA (mRNA). Many RNAs are transcribed from the DNA by RNA polymerases and remain functional even if they do not code. These are referred to as non-protein-coding RNAs and are involved in diverse levels of gene regulation, such as epigenetic, transcriptional, and post-transcriptional [6].
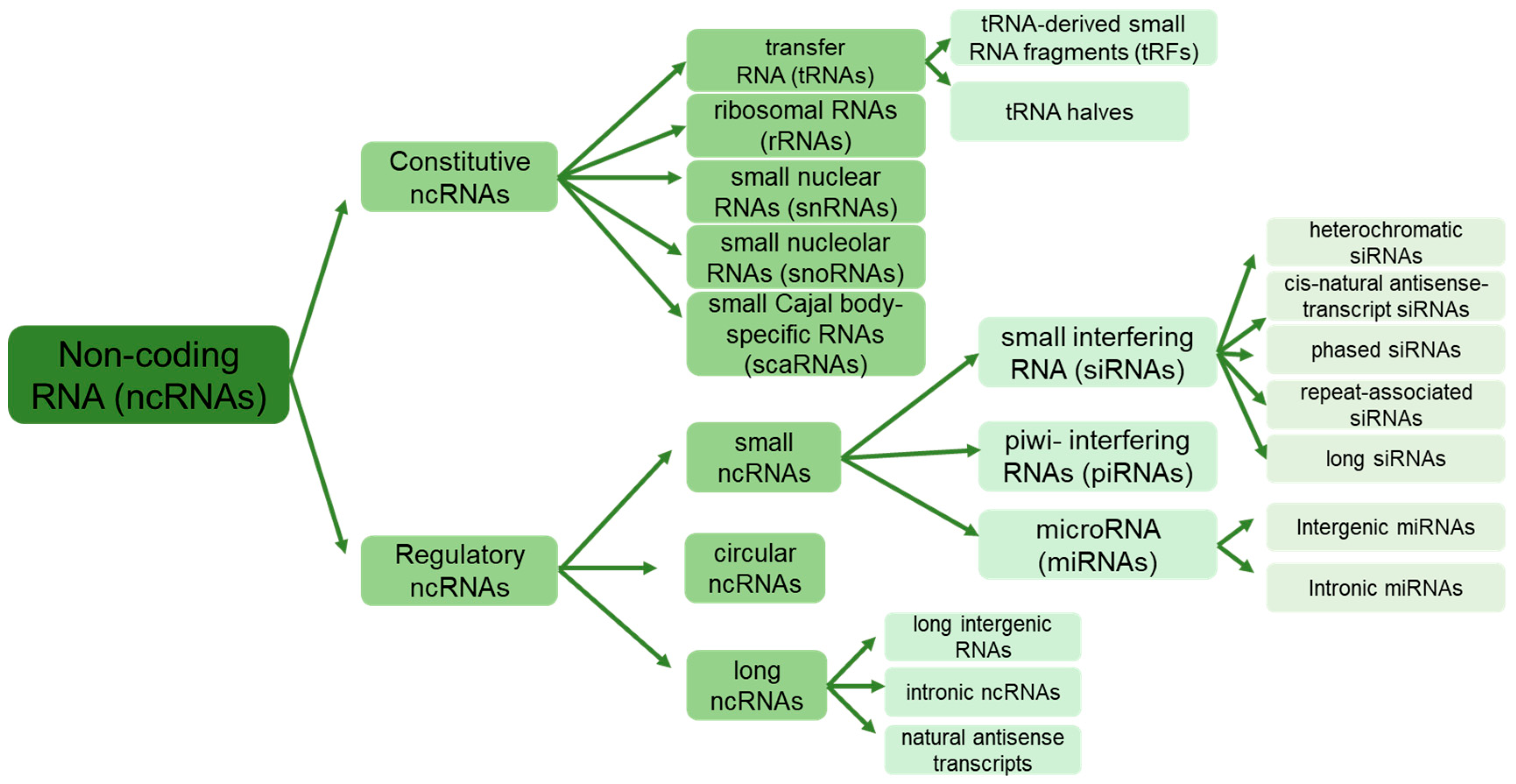
Non-protein-coding RNAs, or non-coding RNAs (ncRNAs), constitute a diverse family of RNA molecules transcribed by RNA polymerase but lack codons for protein synthesis [7]. They differ from regular RNAs in their origin and ways of action on target genes and are classified into two categories: regular/housekeeping/constitutive ncRNAs and regulatory ncRNAs (Figure 1). Constitutive ncRNAs, such as ribosomal RNAs (rRNAs), transfer RNAs (tRNAs), small nuclear RNAs (snRNAs), small nucleolar RNAs (snoRNAs), and small Cajal body-specific RNAs (scaRNAs), are constantly expressed in the cells [8]. These ncRNAs typically range from 50 to 200 nucleotides long, facilitating various cellular and ribosomal functions. Furthermore, recent investigations have unveiled complex populations of tRNA-derived small RNA fragments (tRFs) formed by cleaving tRNA at various positions and typically ranging from 17 to 26 nucleotides [9].
Regulatory ncRNAs, on the other hand, are divided into two classes based on their length: long-non-coding RNAs (lncRNAs) exceeding 200 nucleotides and small non-coding RNAs (sRNAs), including microRNAs (miRNAs) of 19–25 nucleotides, small interfering RNAs (siRNAs) of 20–24 nucleotides, and piwi-interacting RNAs (piRNAs) with a length of 24–32 nucleotides [10,11]. The ncRNAs are further categorized based on chromosomal location, sequence/structure, ways of splicing, and biological functions [12,13]. Recently, a unique group of endogenous ncRNAs called “circular” RNAs (circRNAs) with a closed-loop structure has been identified [14,15]. circRNAs are ubiquitous and abundant in all eukaryotes and even in prokaryotic archaea [15].
ncRNAs play a wide range of crucial roles in many cellular and biological processes, such as plant growth, development, senescence, and responses to biotic and abiotic stressors, despite their inability to undergo protein translation [10]. These roles are often associated with processes such as RNA splicing, chromatin remodeling, epigenetic memory, turnover, editing, transcription, and post-transcriptional regulation of gene expression [16,17]. The regulatory pathways facilitated by ncRNAs are interconnected, controlling the expression of target genes either synergistically or antagonistically. This intricate regulation protects the genome from both external and internal factors [18]. For example, miRNAs act as negative regulators of target gene expression at the post-transcriptional level, while lncRNAs may compete with miRNAs to build a regulatory network that coordinates the expression of target genes [19,20].
Approximately 35–40 years ago, a single ncRNA was typically identified using a chemical or enzymatic sequencing-based approach. This process involved the size-separation of total RNA on a denaturing gel, followed by visualization and excision of a specific band. In subsequent years, complementary DNA (cDNA) libraries, tilling arrays, and microarrays gained popularity for identifying ncRNAs, each with its own set of limitations and concerns [21]. Recent improvements in computational methods and next-generation sequencing technologies, notably in the realms of transcriptome profiling and single-cell RNA sequencing, have facilitated the identification of thousands of new cell-type-specific ncRNAs. Furthermore, deep learning models are now being developed to predict ncRNA identification and classification [22]. The expression of ncRNAs in the lab can typically be validated using reverse transcription polymerase chain reaction (RT-PCR), and their functionality can be assessed using high-precision single-base CRISPR/Cas9 technology. This allows the creation of ncRNA-related mutants to effectively explore the relationship between ncRNAs and their target genes.
The first ncRNA discovered in baker’s yeast was alanine-tRNA, which played a pivotal role in understanding genetic regulation. Subsequently, the regulatory role of ncRNAs emerged as a significant revolution in other kingdoms (animals, fungi) as compared to plants [23]. In this review, we emphasized an overview of ncRNAs over the past 20 years (2003–2023) that regulate the target expression of genes and their involvement in growth, development, and stress resilience, contributing to improved agronomic traits for sustainable yield production in many model plant species in the face of future climate change.
2. Biogenesis of Housekeeping and Regulatory ncRNAs
2.1. Transfer RNA Derived Fragments (tRFs)
tRNAs represent the second most abundant RNA type after rRNAs. They are typically transcribed from precursor tRNA in the nucleus by RNA polymerase III, resulting in the generation of mature tRNA after the removal of 5′ leader and 3′ trailer sequences, as well as the addition of a CCA tail at the 3′ end. The cleavage of 5′ and 3′ ends usually results in transfer RNA-derived fragments (tRF)-5s and tRF-3s, respectively, which range in length from 13 to 20 nucleotides [9]. Mature tRNA with an anticodon loop can be cleaved to produce 5′ or 3′ tRNA halves, spanning in length from 31 to 40 nucleotides. tRFs regulate gene expression through transcription inhibition, RNA degradation, and translation regulation. This regulatory function was evident in processes such as the symbiotic relationship between microbes and soybean roots [24] and the development of pollen grains in Arabidopsis [25].
2.2. Small Interfering RNAs (siRNAs)
Small interfering RNAs (siRNAs), typically 21–24 nucleotides long, are ubiquitous in both animal and plant kingdoms. The generation of siRNA is primarily dependent on one of six RNA-dependent RNA polymerases (RDR1–6) that copy single-stranded RNA (ssRNA) to generate double-stranded RNA (dsRNA) [26]. This dsRNA is then precisely processed into small RNA (sRNA) duplexes by RNase III Dicer-like (DCL1-4) proteins. DCL1 predominantly produces 18–21 nucleotide-long sRNA, whereas DCL2, DCL3, and DCL4 generate 22-, 24-, and 21-nucleotide-long sRNA, respectively. After dicing, sRNA duplexes either remain in the nucleus for chromatin-level activities or are exported to the cytoplasm, where they are then assembled into ARGONAUTE (AGO) protein by the RNA-induced Gene Silencing Complex (RISC) for post-transcriptional gene silencing (PTGS) [27].
siRNA can originate both exogenously (from viral RNA and transgenes) and endogenously from repeat-rich genomic regions, transposables, and retro-elements [28]. Depending on their origin and processing enzyme involved, endogenous siRNAs are categorized into various types, including cis-acting siRNAs (casiRNAs) or heterochromatic siRNAs (hcsiRNAs; 24 nucleotides), cis-natural antisense-transcript siRNAs (cis-NATs; 24 nucleotides), phased siRNAs (phasiRNAs; 21 or 24 nucleotides), repeat-associated siRNAs (rasiRNAs; 24 nucleotides), and long siRNAs (lsiRNAs; 30–40 nucleotides) [29,30].
hcsiRNAs are the most abundant, contributing nearly 70%, and their biogenesis depends on RDR2 and DCL3. They serve an essential function in plant reproduction as well as in maintaining genome integrity and epigenetic modification [31]. cis-NATs, typically present in 6–16%, are usually amplified and processed by RDR6 and DCL1. They also play an important role in plant development and responses to environmental cues [32]. phasiRNAs, a type of secondary siRNA, are typically processed by miRNA-mediated cleavage of RNA transcripts and generated from PHAS loci. These loci, located within the protein-coding region, include many transcription factors (MYBs, NAC), hormone-responsive genes (AUXIN RESPONSE FACTORs; ARFs), and nucleotide-binding leucine-rich repeat (NLR) genes. These ncRNAs function as negative regulators in many biological processes [33]. In addition, PHAS loci are found within noncoding regions, producing lncRNAs, which then generate trans-acting siRNAs (tasiRNAs) through TAS loci [34]. tasiRNAs function in a trans manner and are involved in post-transcriptional regulation, particularly in anther development [35]. rasiRNAs, derived from transposons and repetitive genomic regions, mediate transcriptional gene silencing to maintain genome stability. Distinct rasiRNAs were discovered between two maize-inbred lines (B73 and Mo17) as well as their hybrid, contributing to a high degree of heterosis [36]. lsiRNAs, usually processed by DCL1, are induced either in response to pathogen infection or under specific growth conditions. For instance, in Arabidopsis, AtlsiRNA-1 to AtlsiRNA-5 was induced by Pseudomonas syringae carrying effector avrRpt2, which silences RAP-domain proteins involved in disease resistance [37].
2.3. MicroRNAs (miRNAs)
Plant microRNAs (miRNAs) are endogenous ncRNAs present in a diverse range of organisms, including both animal and plant kingdoms. They typically consist of approximately 20–24 nucleotides. The first miRNA was discovered in Caenorhabditis elegans, where it orchestrates developmental timing by binding to specific target mRNAs [38].
miRNAs typically originate from a stem-loop structure and can be classified as either ‘intergenic’ or ‘intronic’. They are transcribed by miRNA (MIR) genes through RNA polymerase II (RNAPII), resulting in single-stranded primary miRNA transcripts (pri-miRNAs). Subsequently, aided by HYPONASTIC LEAVES 1 and SERRATE, pri-miRNA undergoes two cleavage events and is processed into a double-stranded miRNA through DICER-LIKE (DCL1). The processed miRNA is then incorporated into the AGO1 protein, exported to the cytoplasm, and binds to target mRNAs. This initiates PTGS by assembling the miRNA-induced silencing complex (miRISC) [39]. The AGO4-mediated RISC usually generates a 24-nucleotide miRNA and is involved in epigenetic modification by targeting DNA methylation at the transcriptional level [34].
miRNAs typically inhibit the expression of target transcripts via two main mechanisms: transcript cleavage and translational inhibition. Specifically, transcript cleavage is facilitated by exoribonuclease activity, while translation inhibition happens through targeting the 5′ untranslated region to prevent ribosome recruitment [40]. Notably, a type of peptide known as miRNA-encoded peptides (miPEPs) is translated from short open reading frames (sORFs) in the 5′ leader sequence of pri-miRNAs [41,42]. These peptides promote the transcription of their corresponding pri-miRNAs, thus regulating many biological processes [43]. For instance, miPEP165a and miPEP171b suppress the target genes associated with lateral root development and stimulate main root growth in Arabidopsis and Medicago, respectively [43]. Furthermore, vvi-miPEP171d1 has been shown to regulate adventitious root formation in grapevine [44], while miPEP858a altered plant development and induced secondary metabolite accumulation by influencing the expression of genes involved in phenylpropanoid pathways and auxin signaling in Arabidopsis [45].
MIR genes are intergenic and rarely arranged in tandem, although clustering does not seem uncommon in some plants, such as soybeans [46]. As of 5 February 2024, nearly 326 MIR gene families have been deposited in the miRbase registry for Arabidopsis thaliana, 174 for Zea mays, 122 for Triticum aestivum, 604 for Oryza sativa, 247 for Physcomitrella patens, and 58 for Selaginella moellendorffii (http://microrna.sanger.ac.uk/sequences, accessed on 5 February 2024) [26]. Interestingly, in Arabidopsis, nearly 100 MIR gene families represent an evolutionary fluid set of molecules that arose after the split between land plants and mosses but before the monocot/dicot divergence. Some miRNA families are conserved in mosses, indicating a very ancient origin. These include miR156, miR160, miR319, and miR390, all of which regulate ancestral transcription factors responsible for specifying basic meristem functions, organ polarity and separation, cell division, and hormonal control [47,48].
2.4. Long Non-Coding RNAs (lncRNAs)
Long non-coding RNAs (lncRNAs) are another type of ncRNA that exceed 200 nucleotides in length and lack discernible coding potential. They are found ubiquitously in plants, animals, fungi, and prokaryotes. Although most lncRNAs are primarily located in the nucleus and associate with chromatin, they can function in both nuclear and cytoplasmic compartments [49]. lncRNA can be broadly classified into three categories: long intergenic RNAs (lincRNAs), intronic ncRNAs (incRNAs), and natural antisense transcripts (NATs) [11]. incRNAs are transcribed in any orientation relative to coding genes, while NATs are transcribed from the antisense strand of genes.
Eukaryotes deploy three conserved multi-subunit RNA polymerases (Pol I, II, and III) to transcribe their nuclear genomes into various coding and noncoding transcripts [11]. All three of these RNA polymerases contribute to lncRNA production. Pol I and Pol III are specifically dedicated to lncRNA production, while Pol II, despite its primary function in mRNA biogenesis, also produces a range of lncRNAs. Notably, in plants, two additional enzymes, Pol IV and Pol V, are utilized to produce lncRNAs, which play a crucial role in recognizing and silencing transposable elements [50].
In terms of gene regulation, lncRNAs exhibit diverse mechanisms, operating at multiple levels through both simple and complex mechanisms. They can function in cis or trans, operate through sequence complementarity to RNA or DNA, and are recognized via specific sequence motifs or secondary/tertiary structures [51]. Furthermore, lncRNAs may compete with mRNAs for miRNA molecules, regulating miRNA-mediated target repression through competing endogenous RNA (ceRNA) [52]. Overall, lncRNAs can serve as precursors of miRNAs and siRNAs and act as endogenous target mimics (eTM), competing for various miRNAs [53].
In recent years, the rapid development of various technologies has led to the discovery of different types of lncRNAs. The first identified plant lncRNA, ENOD40, was isolated from alfalfa and demonstrated to modify the subcellular localization of the nuclear RNA-binding protein MtRBP1 (RNA-binding protein 1) [54,55]. Subsequently, several lncRNAs involved in many developmental and stress resilience processes have been identified. For instance, lncRNA SVALKA is associated with freezing response [56], seed dormancy through the Delay of Germination 1 gene (DOG1 antisense) [57], and FLOWERING LOCUS C (FLC) gene silencing during vernalization through COLD ASSISTED INTRONIC NONCODING RNA (COLDAIR) [58], COLDWRAP [59], COLD INDUCED LONG ANTISENSE INTRAGENIC RNAs (COOLAIR) [60], and COLD INDUCED lncRNA 1 (CIL1) [61].
Many sORFs, like miPEPs, are embedded in lncRNAs, encoding functional micropeptides. These can be predicted using several bioinformatics tools, such as FuncPEP, SmProt, PsORF, and many more [62], and their existence can be confirmed through ribosome profiling and mass spectrometry methods [2]. Notably, micropeptides such as DEVIL1 [63], ROTUNDIFOLIA [64], and POLARIS [65] have been identified in Arabidopsis, which play an important role in organ development, leaf morphogenesis, and root growth, respectively. Additionally, Lin et al. [66] identified a subset of micropeptides involved in metabolite, energy, and defense-related processes in soybean.
Presently, single-cell RNA sequencing has been extensively used to identify cell-type-specific lncRNAs and lncRNA-associated gene regulatory networks [67]. Recently, He et al. [68] identified cell-type-specific lncRNA signatures from Arabidopsis seedlings and deposited all the data at the single-cell-based plant lncRNA atlas database (scPLAD), which can be used for functional dissection of lncRNAs in plants.
2.5. Circular RNAs (circRNAs)
Over the past decade, a multitude of circular RNAs (circRNAs) have been identified, demonstrating their ubiquity and abundance across all eukaryotes [14,15,69,70]. circRNAs are single-stranded, endogenous RNAs characterized by a distinct closed-loop structure. They are produced using a non-canonical process known as “back-splicing,” in which the 5′ and 3′ ends are linked by a covalent bond. Due to their closed-loop structure, circRNAs exhibit greater stability compared to linear RNAs and are less susceptible to degradation by Ribonuclease R (RNase R). Although back-splicing is generally less efficient than linear splicing and their expression levels are relatively low, circRNAs can accumulate in specific cell types or tissues in a temporally regulated manner owing to their high stability [71]. Usually, rRNA-depleted total RNA-seq is widely used in early genome-wide profiling studies of circRNAs. However, due to low expression levels of circRNAs, an additional protocol that uses linear RNase R for library preparation has become the preferred method for greatly enriching circRNAs [72,73].
The precise biogenesis of circRNAs remains unclear, but two conserved models have been proposed to explain their formation. In the first model, circRNAs typically originate from canonical splicing sites, facilitated by base pairing between inverted repeat elements located in upstream and downstream introns or through the dimerization of RNA-binding proteins (RBPs) binding to specific motifs in the flanking introns [15]. In the second model, circRNAs are produced from lariat precursors during exon-skipping or from intron lariats that escape debranching [74].
circRNAs can arise from a wide range of genomic positions and combinations, including exons (exonic circRNAs), introns (intronic circRNAs), intergenic regions (intergenic circRNAs), or UTR circRNAs [75]. circRNAs are involved in the regulation of gene expression at transcriptional and post-transcriptional levels. They are reported to function as miRNA sponges [76] or RBP sponges [77] and are considered ideal biomarkers [78].
2.6. piwi-Interacting RNAs (piRNAs)
piRNA, or piwi-interacting RNA, was first discovered in Drosophila melanogaster, derived from single-stranded RNA and exclusively present in animals. Mature piRNAs are around 21–34 nucleotides in length, with a preference for uridine at the 5′ end and 2′ O-methylation at the 3′ end [79]. piRNAs bind and guide PIWI proteins to specific mRNA targets through sequence complementarity between the target RNA and piRNAs. They play important roles in gene regulation, transposon element repression, and antiviral defense [80]. Since piRNAs are not found in plants, they will not be the focus of this review article.
3. Role of ncRNAs in Plant Growth and Development
Plant ncRNAs have emerged as important regulators of chromatin dynamics, exerting influence over transcriptional programs and contributing to various developmental processes such as root, leaf, flower, endosperm, and seed development. These ncRNAs are pivotal in shaping the epigenetic landscape and coordinating gene expression to achieve specific developmental outcomes in plants (Table 1).
3.1. Root Growth and Development
Roots are indispensable plant organs lying beneath the soil surface that not only provide essential structural support to the aerial portion of the plant but also play a vital role in acquiring nutrients and water for growth. In Arabidopsis, the root system architecture comprises primary roots (PRs) and individual lateral roots (LRs) [81]. PRs develop from stem cells surrounding the quiescent center through several rounds of asymmetrical cell divisions and expansions, eventually forming the root meristem [82]. Conversely, LRs originate from the initial specification of pericycle cells, a heterogeneous tissue composed of quiescent cells at the phloem poles and specified cells designated to form lateral root primordia opposite each xylem pole [83].
Several classes of ncRNAs, including siRNAs [84,85], miRNAs [86,87,88], lncRNAs [89,90,91,92,93,94], and circRNAs [95], have been identified as important regulators of root development in many plant species (Table 1). Among those studies, nuclear speckle RNA-binding proteins have been implicated in auxin-regulated LR formation and function as regulators of alternative splicing (AS) [93]. These proteins interact not only with some of their alternatively spliced mRNA targets but also engage with Alternative Splicing Competitor RNA (ASCO), a structured lncRNA comprising ENOD40 and lnc351 [90]. This interaction allows them to influence gene expression by hijacking AS regulators [92].
Root hairs (RHs) are specialized single-cell projections that originate from epidermal trichoblast cells [96]. They play essential roles in absorbing water-soluble nutrients, interacting with soil microorganisms, and anchoring the plant. Recently, the AUXIN-REGULATED PROMOTER LOOP (APOLO) lncRNA has been shown to recognize multiple spatially independent genes by sequence complementarity and DNA–RNA duplex formation, known as R-loops [89]. APOLO integrates both endogenous (PINOID) and exogenous signals, working in conjunction with a complex network of epigenetic factors such as the polycomb repressive complex 1 (PRC1) component LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), which maintains a repressive chromatin status [89]. APOLO lncRNA controls the master regulator of RHs, ROOT HAIR DEFECTIVE 6 (RHD6) at low temperature (10 °C) by decoying LHP1 away from chromatin through interaction with WRKY42 [94]. This modulates WRKY42 binding to the RHD6 promoter, leading to the activation of downstream target genes ROOT HAIR DEFECTIVE SIX-LIKE 2 (RSL2) and ROOT HAIR DEFECTIVE SIX-LIKE 4 (RSL4), which promote RH growth and development [91].
The formation and development of adventitious roots (ARs) are crucial for the vegetative propagation of woody species, particularly in poplar [97]. ARs originated from a cell layer reminiscent of the root pericycle, sharing histological and developmental characteristics with LRs [98]. Plant-specific WUSCHEL-related homeobox (WOX) transcription factors (TFs), especially WOX11, LATERAL ORGAN BOUNDARIES DOMAIN 16, and SMALL AUXIN-UP RNA 36 (SAUR36), are essential regulators of AR formation via the auxin pathway [99]. Numerous lncRNAs that regulate the expression of WOX11 have been identified [97,100]. Among them, lncWOX11a is located 54,293 bp upstream of PeWOX11a on chromosome 13 and is mainly expressed in 1-week-old roots. Overexpression of lncWOX11a inhibits AR development, while CRISPR/Cas9 knockout demonstrated that lncWOX11a acts as a negative regulator of adventitious rooting by downregulating PeWOX11a [97].
3.2. Leaf Development
Leaf development is an intricate and sequential process that plays a critical role in plant growth. Leaves, as lateral organs, go through various key developmental stages, including leaf primordium initiation (arising from the shoot apical meristem), leaf polarity establishment, development phase transition, leaf morphology modulation, and eventually leaf senescence [101,102].
In the realm of leaf development, multiple siRNAs [103,104,105], miRNAs [47,106,107,108,109], lncRNAs [110,111], circRNAs [112], and miRNA-transcription factor regulatory modules [113,114,115,116] have also been identified, each playing pivotal roles (Table 1). For example, miR164 negatively regulates the CUP-SHAPED COTYLEDON (CUC) genes, CUC1 and CUC2, which encode a paralogous pair of NAC transcription factor and are implicated in leaf serration [115]. Similarly, miR165/166 targets the class III homeodomain leucine zipper transcription factor (HD-ZIPIII), crucial for regulating processes such as meristem initiation, lateral organ polarity, and vascular development [117,118,119]. In Arabidopsis, the miR165/166 gene family comprises nine members, including two miR165 (miR165a and miR165b) and seven miR166 (miR166a to miR166g) [120]. Despite a single nucleotide difference between the mature microRNAs of these two families, their target sites within the HD-ZIPIII are highly conserved across all land plants. Notably, the expression patterns are complementary, with HD-ZIPIII transcription factor expressed in the meristem and on the adaxial side of developing leaves, while miR166 accumulates on the abaxial surface of young leaf primordia in Arabidopsis [118].
The evolutionarily conserved miR319 regulates TCP, a plant-specific transcription factor named after TEOSINTE BRANCHED, CYCLOIDEA, and PCF1/PCF2 [109]. miR319 affects TCP2, TCP3, TCP4, TCP10, and TCP24, as well as their homologs in different plant species [106]. Elevated levels of miR319 lead to the downregulation of these TCPs, resulting in dramatic alteration in Arabidopsis leaf morphogenesis and the development of crinkled leaves [121,122,123]. miR396 targets seven genes (GRF1-4 and GRF7-9) from the growth-regulating factor (GRF) [124,125]. Mutations in various GRFs or overexpression of miR396 result in reduced leaf size, indicating partial functional redundancy [114]. Conversely, overexpressing GRFs or mutating the miR396-binding site of GRF2, GRF7, or GRF9 results in slightly larger leaves [125,126].
As plants reach the maturation stage, leaves undergo organ-level senescence, which is a critical process for optimal offspring production and plant survival. A recent study by Kim et al. [111] conducted comprehensive RNA-seq analyses on Arabidopsis leaves, representing all developmental stages. This led to the identification of approximately 770 age-related (AR) lnRNAs. Among them, AT1G33415 and AT2G14878 were shown to be coexpressed with Autophagy 9 and ADENINE PHOSPHORIBOSYL TRANSFERASE 1, regulating leaf senescence through decreased cytokinin metabolism [127]. Furthermore, three competitive endogenous RNAs (ceRNAs) were identified, which involved lncRNA-miRNA-mRNA interactions. The AR-lncRNAs AT4G36648, AT5G23410, and AT1G26208 interact with miR164, miR169, and miR156 to target ORESARA1, NUCLEAR FACTOR Y (NF-Y), and SQUAMOSA PROMOTER BINDING-LIKE (SPL), respectively [111]. This regulation is crucial for age-dependent developmental transitions, specifically in leaf senescence, as well as responses to abiotic and biotic stresses [128]. Overall, these studies shed light on the intricate miRNA-TF regulatory network governing leaf development and offer potential avenues for manipulating leaf-related traits in plant species.
3.3. Seed, Endosperm, and Seed Nutrient Development
Grain development constitutes a pivotal biological process intricately connected to both grain yield and quality in plants. The complexity of grain yield as a trait is determined by several factors, including spike/panicle number per plant, grain number per spike/panicle, and one thousand kernel weights. Notably, these factors are predominantly influenced by the architecture of the spike/panicle [129].
Several ncRNAs [130,131,132,133,134,135,136,137] have been identified that regulate seed-related features, as summarized in Table 1. Notably, a comprehensive study by Zhang et al. [136] discovered nearly 65 lncRNAs associated with rice reproduction using the strand-specific RNA sequencing method. Among them, three lncRNAs (XLOC_0063639, XLOC_007072, and XLOC_057324) were highly expressed during the reproductive stage. Another study by He et al. [138] identified a quantitative trait locus linked to improved rice yield, comprising several tandemly arranged intronless leucine-rich repeat receptor kinase (LRK) genes. Subsequently, Wang et al. [134] identified lncRNA transcribed from the antisense strand of the neighboring gene LRK cluster, named LRK Antisense Intergenic RNA (LAIR). Silencing LAIR in transgenic lines resulted in inhibited growth and reduced expression levels for all LRK genes. Conversely, lines overexpressing LAIR exhibited significant increases in total grain yield, coupled with enhanced expression of some members of the LRK gene cluster.
In wheat, Cao et al. [137] also identified 2753 differentially expressed lncRNAs, highlighting their significant roles in spike development. However, this finding was limited to a single wheat genotype, leaving the genetic variations and evolutionary characteristics of wheat lncRNAs largely unexplored. Yang et al. [139] addressed this gap by conducting a comprehensive study involving 93 wheat genotypes to unveil the lncRNA landscape associated with wheat spikes. The analysis yielded a total of 35,913 lncRNAs, along with 1619 lncRNA-mRNA pairs. Among these pairs, lncRNA.127690.1-TraesCS2A02G518500.1 and lncRNA.104854.1-TraesCS6A02G050300 were identified as regulators of heading date and spike length, acting through pectin methylesterase inhibitor protein and autophagy protein (AtATG5), respectively.
Oil is a valuable component in seeds that plays an important role in human health [140]. Most oilseed crops produce and accumulate oil during seed development and maturation, involving morphogenesis, cell enlargement/reserve accumulation, and desiccation or developmental arrest [141]. Oil biosynthesis in seeds occurs in two subcellular compartments and involves four key steps: de novo fatty acid synthesis, acyl editing, triacylglycerol assembly, and oil drop formation [141]. Recently, lncRNAs have been implicated in regulating seed oil biosynthesis [142,143,144,145]. For instance, in the developing seeds of Brassica napus, the expression patterns of 13 lncRNAs were significantly correlated with lipid-related genes, suggesting potential roles for these lncRNAs in oil synthesis [142]. Subsequently, Li et al. [145] identified 8094 expressed lncRNAs across three developing stages in two accessions of Brassica napus, ZS11 (high oil content) and WH5557 (low oil content). Notably, lncRNAs MSTRG.22563 and MSTRG.86004 were predicted to be associated with seed oil accumulation. Overexpression of MSTRG.22563 reduced seed oil content by 3.1–3.9% and altered the fatty acid composition of seeds. In contrast, the lncRNA MSTRG.86004 boosted oil content by 2.0% while delaying seed development.
Endosperm development is a crucial process in cereal grains that directly impacts human nutrition. It is an important tissue for studying the molecular mechanisms that drive its development as well as metabolic processes contributing to the formation of its storage constituents. The development sequence initiates with an initial syncytial phase, followed by cellularization, in which the endosperm nuclei undergo multiple rounds of mitosis without cytokinesis, resulting in a multinucleate cell [146]. The transition from syncytium to cellularization is pivotal for determining final seed size, making it a useful model for studying the cell cycle and cytokinesis. The helicase family protein (HeFP) plays a vital role in endosperm cellularization [147]. Hara et al. [148] identified the rice SNF2 family helicase ENDOSPERMLESS 1, which regulates syncytial endosperm development. In a subsequent study, Zhou et al. [149] uncovered a mutant with a T-DNA insertion in a lncRNA, MIS-SHAPEN ENDOSPERM (MISSEN; XLOC_057324), maternally expressed and repressed by histone H3 lysine 27 trimethylation (H3K27me3) post-pollination. MISSEN suppressed endosperm development by negatively regulating HeFP, leading to a prominent dent and bulge in the seed. Both studies revealed the potential applications of the HeFP pathway in breeding for improved grain yield in rice.
The endosperm is primarily made up of starch, which forms a complex glucose polymer with a linear chain of amylose and highly branched amylopectin [150]. The dietary consumption of cereals, particularly those with high glycemic content, is often associated with diet-related diseases such as obesity, type-2 diabetes, and various cardiovascular diseases. Elevating amylose content contributes to type 2 resistant starch (RS), as it undergoes slow digestion in the intestine, resulting in a low glycemic index [151]. In a recent study, Madhawan et al. [152] uncovered a lncRNA–miRNA–mRNA regulatory network where TCONS_00130663 interacts with miR1128 to regulate lysophospholipase gene expression. Concurrently, this lncRNA, which has a negative correlation with starch branching enzyme IIb (SBEIIb) in a high RS mutant line, may have a role in regulating the RS via an unknown mechanism. In another study, Zhang et al. [153] also identified several lncRNAs, which coexpressed with genes involved in starch metabolism such as Brittle 1 (TraesLNC6D200.1; TaBT1-6D), cell wall invertase genes (TaCwi-5D; TraesLNC5D5400.1 and TaCwi-4A; TraesLNC4A35700.1), sucrose transporter gene (TaSUT-1A; TraesLNC1A1200.1), sucrose non-fermenting 1-related protein kinase 2 (TaSnRK2.10-4A; TraesLNC4A37800.1), and granule bound starch synthase II (TaGBSSII; TraesLNC2B15700.1). These lncRNAs not only regulate starch metabolism but also serve as potential candidates for functional studies on wheat grain size and weight regulation.
Storage protein (SP), which includes glutelin and gliadin, represents another important component in the endosperm that not only determines the baking quality but also contains several important amino acids such as proline, glutamine, asparagine, and arginine [154]. Several lncRNAs have been found to coexpress with SP in wheat [153,155]. For instance, the antisense lncRNA TraesLNC1D26001.1, which is transcribed from the promoter of the high molecular weight glutelin subunit (HMW-GS), coexpressed strongly with TaGLU-1Dy. Functional analysis of TraesLNC1D26001.1 revealed no significant changes in protein level, suggesting that lncRNA does not regulate the transcription or translation of its adjacent gene. However, the study identified ABSCISIC ACID-INSENSITIVE 5 (AIB5), a putatively trans-regulated gene involved in seed maturation, germination, and abscisic acid (ABA) response, showing higher expression in lncRNA-overexpressed constructs [155]. The AIB5 further exhibited reduced germination compared to the wild type, especially with increased ABA construction, indicating that TraesLNC1D26001.1 overexpression enhanced ABA sensitivity during seed germination [155]. The delay of seed germination was also associated with a seed vigor-related lncRNA (SVR; lncRNA MSTRG.182510.6) that interacts with members of the SAUR gene family. CRISPR/Cas9-engineered mutations in SVR delayed rice germination through the auxin-activated signaling pathway [156]. Overall, this research provides valuable insights into the regulatory role of lncRNAs in seed-related biological processes, offering potential applications in crop improvement.
4. Role of ncRNAs in Various Abiotic Stress Resilience
Plants, as sessile organisms, are exposed to a variety of abiotic stresses, including heat, drought, salinity, flooding, low temperatures, and others. These stresses can lead to a substantial reduction in plant growth and overall productivity, with documented impacts ranging from 50 to 70% worldwide [157,158,159]. Abiotic stresses, especially those linked to global warming, have a substantial and pervasive deleterious impact on plants on a broad geographic scale. The escalating prevalence of these stresses, exacerbated by factors including climate change, poses significant challenges to global agriculture and plant ecosystems. To effectively handle this scenario, dedicated research, the development of stress-tolerant crop varieties, and the implementation of sustainable agricultural practices are essential. These approaches aim to reduce the negative impacts of abiotic stresses on plant productivity while increasing their resilience in the face of environmental challenges.
4.1. Drought Stress
Drought stress poses a major agricultural risk, prompting urgent global efforts to enhance drought-tolerant crops. Drought stress has a deleterious impact on plants, including osmotic imbalance, membrane system degradation, reduced respiration, and photosynthetic rates. These responses not only impede plant growth and metabolism throughout its life cycle but also diminish crop yield and quality.
A list of several drought-responsive ncRNAs has been summarized by Gelaw et al. [160], which are listed in Table 1 [161,162,163,164,165,166,167,168,169,170,171,172,173]. Most of them target genes involved in ethylene and ABA synthesis or signaling, calcium signaling, starch and sucrose synthesis, and a variety of other metabolic processes. For instance, the lncRNAs present in rice, NAT Os02g0250700-01 and NAT Os02g0180800-01, target the late embryogenesis abundant protein and cinnamoyl CoA reductase genes, respectively [168]. Numerous lncRNAs in maize were discovered from three distinct tissues (ear, tassel, and leaf) across four developmental stages (V12, V14, V18, and R1) under both well-watered and drought conditions [169,170]. Among the total 3,488 high-confidence identified lncRNAs, MSTRG.2834.1 responded to drought specifically in the ear, with the greatest expression changes at the R1 stage and negligible changes at the V12, V14, and V18 stages upon drought stress. However, drought stress barely influenced the expression of lncRNA MSTRG.43642.1 in the ear at the V12, V14, and V18 stages, but it was significantly downregulated at the R1 stage. They also identified 1177 lncRNA-mRNA interactions, with lncRNA MSTRG.6838.1 targeting vpp4 (GRMZM2G028432), encoding a vacuolar (H+)-pumping ATPase subunit gene. The expressions of vpp4 and MSTRG.6838.1 were significantly correlated in various tissues and development stages, and they were both down-regulated under drought stress, indicating that lncRNA MSTRG.6838.1 and vpp4 may be promising cis-acting pairs that hold potential for co-regulation.
The discovery of drought-responsive lncRNA (DRIR) demonstrates its crucial role in regulating plant tolerance to drought and salt stresses [171,172]. Zhang et al. [173] recently conducted large-scale profiling and identified 2174 and 1354 high-confidence circRNAs in maize and Arabidopsis, respectively. Specifically, they found two circRNAs associated with the GRMZM2G032852 and GRMZM5G817173 genes, which encode a calcium-dependent protein kinase (CPK) and cytokinin oxidase/dehydrogenase (CKX), respectively. These circRNAs, named circCPK and circCPK, were formed through back-splicing between exons, resulting in the insertion and deletion of alleles in the maize panel. The expression levels of circCPK and the survival rate of maize plants with the insertion alleles were notably higher than those with the deletion alleles under both normal and drought conditions. This observation suggests a potential association between circRNA expression and plant drought tolerance.
Furthermore, in another study, the researchers investigated the role of circGORK, which is hosted by the GORK (Guard cell outward-rectifying K+-channel) gene [174]. GORK encodes a potassium output channel that is critical for the regulation of ABA signaling and water stress responses [175]. Overexpression of circGORK in transgenic Arabidopsis plants had a negative impact on the ABA response. Specifically, the germination rate of transgenic seeds was considerably lower on ABA plates than that of control seeds, suggesting that circGORK may modulate ABA signaling pathways and influence water stress responses.
4.2. Salt Stress
Salt stress poses a substantial barrier to plant growth and development, affecting approximately 33% of arable land. It increases both ionic and intracellular osmotic pressures in plants, resulting in increased toxicity, oxidative damage, and osmotic stress [176]. Rice is vulnerable to salt stress, with most cultivated varieties having an electrical conductivity threshold of 3 dSm−1 [177].
Limited evidence is available on ncRNAs associated with the regulation of salt stress in plants [177,178,179,180,181,182,183,184,185,186,187,188,189,190], as listed in Table 1. Nearly 75 conserved miRNAs were identified from the control and NaCl-treated salt-tolerant rice varieties [180]. Among them, osa-miR12477, which targets L-ascorbate oxidase, showed an accumulation of oxidative stress in the plant upon salt treatment. Similarly, Jain et al. [191] identified 1500 and 2100 putative lncRNAs from the root and leaf tissue of salt-tolerant (Horkuch) and salt-sensitive (IR-29) rice genotypes, respectively. The study further identified two differentially expressed (DE)- lncRNAs, such as TCONS_00008914 and TCONS_00008749, as potential target mimics of known rice miRNAs. In another study, using another salt-tolerant rice (FL478), Mansuri et al. [177,187] identified DE-lncRNA.2-FL, that showed multiple interaction with osa-miR2926 miRNA. This interaction regulates the expression of various salt-stress-responsive genes, such as chloride channel protein, potassium transporter, and some genes involved in sensing and signaling, such as cysteine-rich receptor-like protein kinase 8 precursors and serine/threonine protein kinase. The lncRNA also identified pentatricopeptide repeat protein as a cis-target and multiple trans-targets that mediate regulatory role in a salt-tolerant rice variety.
lncRNA DRIR was identified as a positive regulator of the plant response to salt stress, in addition to its role in drought stress [171]. Under normal conditions, DRIR expression was modest, but it was significantly induced by drought, salt stress, and ABA treatment. A genome-wide transcriptome analysis was conducted to investigate the impact of DRIR on gene expression in response to stress. The results indicated changes in the expression of genes involved in ABA signaling, water transport, and other stress-relief processes in both the drir mutant and overexpressing plants. The study provides valuable insights into the regulatory role of DRIR in the complex network of molecular responses that plants undergo to cope with drought and salt stress.
4.3. Flooding/Waterlogging/Hypoxia Stress
Flooding is the temporary submersion of land by water, while waterlogging happens when the soil is either fully saturated or near saturation most of the time. Both processes hinder various physiological processes in plants, such as photosynthesis, disturb cellular energy production, and disrupt ionic balance [192]. These effects worsen when plants encounter lower oxygen content below optimal conditions, a phenomenon known as hypoxia, which frequently happens in the natural environment of plants. The restricted availability of oxygen in the root zone adversely impacts the overall metabolism of the plant, impairing its growth and development [193].
Several ncRNAs have been detected in different plant species exposed to waterlogging or hypoxia conditions, such as Arabidopsis [194], maize [195,196], rice [197], Populus [198], Medicago [199], tomato [200], and cucumber [201,202], and some of them are shown in Table 1. For instance, in rice, nearly 291 and 58 miRNAs responsive to deepwater stress were discovered in the internode and node, respectively [197]. Further, degradome analysis predicted nearly 1586 and 744 targets cleaved by 230 and 53 miRNAs from the internode and nodes, respectively. Notably, miR156 targeted the SQUAMOSA-promoter binding protein (SBP)-box gene (cleaved at 1163 nts), and NF-Y transcription factor (cleaved at 1729 nts) was targeted by miR169. The study further identified miRNA-lncRNA-mRNA regulatory networks, such as nov-miR145-TCONS_00011544 and miR159e-TCONS_00059798, targeting ZFD (Os11g36430) and MYB (Os01g36460) genes, respectively.
Flooding is also associated with reduced energy metabolism, as detected by a sucrose non-fermenting-related protein kinase (SnRK1) [203]. Overexpression of miR156 has previously been shown to improve abiotic stress tolerance in various plants via regulating SQUAMOSA-promoter binding protein-like (SPL) transcription factors [204,205]. Later, Feyissa et al. [199] studied the role and putative mode of action of the miR156/SPL module in alfalfa flooding tolerance. They utilized many genetic constructs, including miR156 overexpression (miR156OE), SPL6RNAi, SPL13_RNAi, wild-type, and empty vector plants, along with flood-tolerant (AAC-Trueman) and flood-sensitive (AC-Caribou) alfalfa cultivars. The study revealed that the miR156/SPL module mediates physiological responses in alfalfa under flooding conditions. Specifically, in miR156OE alfalfa plants, SnRK1 expression increased in an ABA-dependent manner. SnRK1 is a key regulator of energy and stress signals in plants. The findings suggest that the miR156/SPL module contributes to flooding tolerance by regulating physiological processes and SnRK1 expression, highlighting the intricate regulatory mechanisms involved in plant stress responses [199].
4.4. Heat and Cold Stress
Both high and low temperatures (chilling at 0–15 °C and freezing at <0 °C) impact plant growth, development, and various physiological functions. Numerous temperature-responsive regulatory genes, such as heat shock factors and C-repeat/dehydration-responsive element binding factors (CBF/DREB1s), bind to the promoters of heat stress response genes and cold-regulated genes (COR), thereby promoting plant adaptability to heat and cold stress, respectively [206].
An array of heat and cold stress-responsive ncRNAs have been identified, comprising several siRNAs [207,208,209], circRNAs [210,211,212], and miRNA-targets [213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229]. Notable examples of miRNAs and their targets that are linked to either heat and cold stress include but are not limited to: miR156-SPL [213,214,215], miR159-GAMYB [216], miR160-ARF [217], miR164-NAC [218], miR165/166-homeodomain leucine zipper class-III transcription factor mainly PHBULOSA and ATHB-8 [219], miR167-ARF6 [220], miR168-AGO1 [221], miR171-GRAS gene family [222], miR172-APETALA2 [223], miR319-TCP [224,225,226], miR393-auxin receptors with F-box domains; TIR1/AFB2 [227], miR396-GRF [228], miR397-laccase gene; lignin-degrading enzyme [222], miR402-DEMETER-LIKE PROTEIN3; a 5-methylcytosine DNA glycosidase [229]. Genome-wide transcriptomic analyses have further identified conserved miRNA (miR166) and unique miRNAs (miR168, miR444, and miR528) in the maize leaves during the V3 stage under heat stress [230]. In sweet potato seedlings, specific miRNAs such as IbmiR162, IbmiR164, IbmiR171, and IbmiR397 have been implicated in physiological changes, particularly oxidative stress during chilling and heat stress [231].
Several lncRNAs were also identified for both heat and cold stress using chip-based microarray and strand-specific sequencing approaches, with high specificity in tissue types and developmental stages [232,233,234,235,236,237]. For instance, lnc-173 targets the SUCROSE SYNTHASE4 gene, and under heat stress, only the target gene was expressed, not the lncRNA itself [237]. Heat shock transcription factors (HSFs) play a vital role in the response and acclimation of eukaryotes under heat stress. In Arabidopsis, asHSFB2a (natural antisense transcript of HSFB2a) was found to be induced by heat stress and negatively regulate HSFB2a expression [238]. Recently, a Heat-induced long intergenic noncoding RNA 1 (HILinc1) was identified in pear [239]. The target of this lncRNA binds with HSF to enhance its transcriptional activity, leading to the upregulation of multiprotein bridging factor 1c during heat response.
During cold stress, plants undergo a process known as vernalization, which prevents flowering during vegetative growth in winter and promotes flowering during the reproductive phase under favorable conditions. This regulatory mechanism of vernalization is controlled by FLC. Several lncRNAs, such as COLDAIR [58], COLDWRAP [59], and COOLAIR [60], suppress the expression of FLC through chromatin modification [240]. Furthermore, lncRNAs SVALKA [56], cryptic antisense CBF1 (asCBF1), and CIR (CBF intergenic lncRNA) regulate freezing tolerance through transcription of the CBF1 gene [241]. The COLD INDUCED lncRNA1 (CIL) participates in low-temperature stress response by affecting the reactive oxygen pathway or osmotic adjustment substance [61], while the cis-acting NAT lncRNA_2962 activates transcription of cold-responsive MADS AFFECTING FLOWERING 4 [242]. These lncRNAs affect plant responses to cold by regulating the expression of multiple stress-related genes during the developmental stage.
4.5. Combined Stresses
In the face of unexpected climate conditions, plants often undergo simultaneous exposure to multiple stress conditions, which significantly impact their growth, development, and overall yield. Several miRNAs have been identified that target not only transcription factors but also hormone-signaling genes under combined stress conditions. For instance, miR156 has been found to enhance heat and cold stress tolerance in Arabidopsis and rice plants, respectively [204,215]. While in alfalfa, miR156 increased plant tolerance to salt, heat, and drought stress by downregulating the SPL gene [214,243,244]. Another notable example is miR393, which targets the auxin receptors with F-box domains, TIR1 and AFBs, thus forming the miR393-TIR1/AFBs module. This module is highly conserved in many plant species and links auxin response with transcriptional regulation of plants under salt [245] and drought stress conditions [246]. miR394 targets the LEAF CURLING RESPONSIVENESS GENE, an F-box protein (SKP1-Cullin/CDC53-F-box), which regulates drought and salt stress responses through ABA-mediated and CBF-dependent cold acclimation pathways [247,248]. Additionally, miR408 targets both the blue copper protein and laccase genes and ultimately enhances plant tolerance to salt, cold, and oxidative stress [164].
In the context of hormone-mediated responses to combined stress, Qin et al. [171] identified a novel lncRNA, DRIR, that regulates drought and salt stress responses by mediating the expression of ABA signaling genes, stomatal closure, and proline accumulation. Additionally, lncRNA77580 in soybeans modulates the expression of a number of transcription factors under salt and drought stress conditions [249]. Furthermore, using high-throughput sequencing technology, approximately 467 circRNAs were identified in tomatoes in response to drought, heat, and combined stress conditions [250]. Among them, 156 circRNAs were shared among all stress conditions, with circRNA197 and circRNA29 showing significant expression levels in three stress conditions and being predicted to play an important role in photosynthesis and other biological processes.
Nowadays, deep learning-based methods, such as the PLncPRO (Plant Long Non-Coding RNA Prediction by Random fOrest) tool, are widely used to predict lncRNAs. Singh et al. [22], utilizing this tool, predicted and identified many lncRNAs that were differentially expressed in drought- and salt-tolerant chickpea genotypes at the early and late reproductive stages, respectively.
5. Conclusions and Future Perspectives
Advances in high-throughput sequencing technologies, such as single-cell RNA sequencing, coupled with bioinformatics tools and validatory methods such as northern blot, quantitative real-time PCR, mass spectrometry, and CRISPR/Cas9, have greatly aided the identification of ncRNAs that play a critical role in a variety of biological processes. ncRNAs are dynamic entities with a shared origin and conserved mode of action across different plant species. They exhibit discrete expression patterns across tissues, which can influence growth and development. At the molecular level, multiple regulatory mechanisms, including epigenetic, transcriptional, and post-transcriptional processes, all have an impact on plant performance in response to various abiotic stress conditions. In contemporary research, deep learning methods are frequently employed to predict tissue-specific and environmental-specific novel ncRNAs in many plant species. The lab-based validation of these ncRNAs not only enhances our understanding of their biological function but also equips researchers with tools to harness these molecules for developing plants with improved resilience, yields, and quality in the face of changing climatic conditions.

{kind=link}
Table 1.
List of studies that identified ncRNAs (siRNAs, miRNAs and their targets, lncRNAs, and circRNAs) for growth, development, and stress resilience in various plant species.
Table 1.
List of studies that identified ncRNAs (siRNAs, miRNAs and their targets, lncRNAs, and circRNAs) for growth, development, and stress resilience in various plant species.
| Trait | siRNAs | Reference |
|---|---|---|
| Root development | TAS3-siRNA (ARFs) | Marin et al. [84] |
| TAS3-siRNA (leafbladeless1) | Gautam et al. [85] | |
| Leaf development | TAS3-siRNA (ARF3) | Chitwoodet al. [103] |
| TAS3-siRNA | Schwab et al. [104] | |
| TAS3-siRNA (leafbladeless1) | Dotto et al. [105] | |
| Seed, endosperm, nutrient development | maternal p4-siRNAs | Lu et al. [130] Yuan et al. [131] |
| Drought stress | Dehydration induced siRNAs | Jung et al. [161] |
| Yao et al. [162] | ||
| Salt stress | OsSIDP301 (stress-induced DUF1644 protein) | Ge et al. [182] |
| Response to ABA and Salt 1 | Ren et al. [183] | |
| OsSKL2 and OsASR | Jiang et al. [184] | |
| NATs of SRO5 and P5CDH | Borsani et al. [185] | |
| Flooding/hypoxia | hypoxia-induced tasiRNA | Moldovan et al. [194] |
| Heat stress | HEAT-INDUCED TAS1 TARGET1 | Li et al. [207] |
| Phased siRNA | Pokhrel et al. [208] | |
| Cold stress | cold-induced tasiRNA | Kume et al. [209] |
| miRNAs and corresponding targets | ||
| Root development | miR165/166-RLD1/2 | Gautam et al. [85] |
| miR156-SPL10 | Barrera-Rojas et al. [86] | |
| miR165-SHR | Carlsbecker et al. [87] | |
| Leaf development | miR319-TCP | Bresso et al. [106] |
| miR390-TAS3 | Husbands et al. [108] | |
| miR396-GRF | Omidbakhshfard et al. [124] | |
| Seed, endosperm, nutrient development | miR156, miR166, miR172, miR319, miR396 | Khemka et al. [132] |
| miR156-SPL14/SPL16 | Miao et al. [133] | |
| Drought stress | miR169-NFYA5 | Li et al. [128] |
| miR408a-Laccase | Jiao et al. [163] | |
| miRNA-mediated drought-responsive regulatory network | Balyan et al. [165] | |
| miR169-NFYA3 | Ni et al. [167] | |
| Salt stress | osa-miR12477-LAO | Parmar et al. [180] |
| Flooding/hypoxia | miR156/SPL | Feyissa et al. [199] |
| Heat stress | miR164-NAC | Tsai et al. [218] |
| Cold stress | miR319-TCP | Thiebaut et al. [225] |
| miR319-OsPCF6 and OsTCP21 | Wang et al. [226] | |
| lncRNAs | ||
| Root development | APOLO | Ariel et al. [89] Moison et al. [91] |
| lncWOX11a | Ran et al. [97] | |
| lncWOX5 | Qi et al. [100] | |
| Leaf development | TWISTED LEAF | Liu et al. [110] |
| age-related lncRNAs | Kim et al. [111] | |
| Seed, endosperm, nutrient development | LAIR | Wang et al. [134] |
| Drought stress | DRIR | Qin et al. [171] Dong et al. [172] |
| Salt stress | Salt-responsive DE-lncRNAs | Mansuri et al. [177] Mansuri et al. [187] |
| Genome-wide lncRNAs in Medicago truncatula | Wang et al. [188] | |
| Flooding/hypoxia | DE-lncRNAs in response to waterlogging stress | Yu et al. [196] Kęska et al. [202] |
| Heat stress | TCONS_00048391 and TCONS_00010856 | Wang et al. [235] |
| Cold stress | CIL1 | Liu et al. [61] |
| circRNAs | ||
| Root development | DE-circRNAs | Xu et al. [95] |
| Leaf development | circ-AT1G29965, circ-AT4G08300, circ-AT5G18590 | Liu et al. [112] |
| Seed, endosperm, nutrient development | DE-circRNAs participate in fatty acid metabolism | Zhou et al. [135] |
| Drought stress | Circular RNA profiling | Zhang et al. [173] |
| circGORK | Ache et al. [174] Becker et al. [175] | |
| Salt stress | circ_000260, circ_001362, and circ_001730 circRNA Chr10:20,345,844|20,346,873 | Liu et al. [189] Yin et al. [190] |
| Heat stress | 1583 heat-specific | Pan et al. [212] |
| Cold stress | 163 chilling responsive 1830 low temperatures induced | Zuo et al. [211] Yang et al. [210] |
Author Contributions
Conceptualization, A.Y., J.M., A.K.D. and A.S.; writing the original draft A.Y., A.K.D. and A.S.; revising and editing the manuscript, A.Y., J.M., A.K.D. and A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
APOLO: AUXIN-REGULATED PROMOTER LOOP, ARF: AUXIN RESPONSE FACTORS, ASR1: Chaperone-like protein, CIL1: COLD INDUCED lncRNA1, circRNAs: Circular RNAs, DE-circRNAs: Differentially expressed-circRNAs, DE-lncRNAs: Differentially expressed-lncRNAs, DRIR: DROUGHT-INDUCED INTERGENIC lncRNA, GORK: Guard cell outward-rectifying K+ -channel, GRF: GROWTH REGULATING FACTOR, LAIR: LRK Antisense Intergenic RNA, LAO: L-ascorbate oxidase, LBL1: leafbladeless1, miRNAs: MicroRNAs, NAC: NAM, ATAF1/2, and CUC2, NATs: Natural cis-Antisense transcripts, NFYA: Nuclear factor YA, P5CDH: Δ1-pyrroline-5-carboxylate dehydrogenase, PCF: Proliferating cell factors, RLD1/2: ROLLED1 and ROLLED2, SHR: SHORT ROOT, siRNAs: Small interfering RNAs, SKL2: shikimate kinase-like 2, SPL: SQUAMOSA PROMOTER BINDING PROTEIN-LIKE, TAS3: TRANS-ACTING SMALL INTERFERING RNA3, TCP: TEOSINTE BRANCHED/CYCLOIDEA/PROLIFERATING CELL FACTOR.
References
- Hajieghrari, B.; Farrokhi, N. Plant RNA-Mediated Gene Regulatory Network. Genomics 2022, 114, 409–442. [Google Scholar] [CrossRef] [PubMed]
- Giambruno, R.; Mihailovich, M.; Bonaldi, T. Mass Spectrometry-Based Proteomics to Unveil the Non-Coding RNA World. Front. Mol. Biosci. 2018, 5, 90. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.M.; Chen, Y.Q. Principles and Innovative Technologies for Decrypting Noncoding RNAs: From Discovery and Functional Prediction to Clinical Application. J. Hematol. Oncol. 2020, 13, 109. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Zhu, W.; Sun, M.; Shi, L. Bioinformatics Analysis of Long Non-Coding RNA and Related Diseases: An Overview. Front. Genet. 2021, 12, 813873. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, W.F. Is Junk DNA Bunk? A Critique of ENCODE. Proc. Natl. Acad. Sci. USA 2013, 110, 5294–5300. [Google Scholar] [CrossRef]
- Patra, G.K.; Gupta, D.; Rout, G.R.; Panda, S.K. Role of Long Non Coding RNA in Plants under Abiotic and Biotic Stresses. Plant Physiol. Biochem. 2023, 194, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and Their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Wajahat, M.; Bracken, C.P.; Orang, A. Emerging Functions for Snornas and Snorna-derived Fragments. Int. J. Mol. Sci. 2021, 22, 10193. [Google Scholar] [CrossRef]
- Alves, C.S.; Nogueira, F.T.S. Plant Small RNA World Growing Bigger: TRNA-Derived Fragments, Longstanding Players in Regulatory Processes. Front. Mol. Biosci. 2021, 8, 638911. [Google Scholar] [CrossRef]
- Waititu, J.K.; Zhang, C.; Liu, J.; Wang, H. Plant Non-Coding Rnas: Origin, Biogenesis, Mode of Action and Their Roles in Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 8401. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long Non-Coding RNAs: Definitions, Functions, Challenges and Recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Fu, X.D. Non-Coding RNA: A New Frontier in Regulatory Biology. Natl. Sci. Rev. 2014, 1, 190–204. [Google Scholar] [CrossRef]
- Domínguez-Rosas, E.; Hernández-Oñate, M.Á.; Fernandez-Valverde, S.-L.; Tiznado-Hernández, M.E. Plant Long Non-Coding RNAs: Identification and Analysis to Unveil Their Physiological Functions. Front. Plant Sci. 2023, 14, 1275399. [Google Scholar] [CrossRef]
- Zhao, W.; Chu, S.; Jiao, Y. Present Scenario of Circular RNAs (CircRNAs) in Plants. Front. Plant Sci. 2019, 10, 379. [Google Scholar] [CrossRef]
- Zhang, P.; Li, S.; Chen, M. Characterization and Function of Circular RNAs in Plants. Front. Mol. Biosci. 2020, 7, 91. [Google Scholar] [CrossRef]
- Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Posttranscriptional Gene Regulation by Long Noncoding RNA. J. Mol. Biol. 2013, 425, 3723–3730. [Google Scholar] [CrossRef]
- Sebastian-Delacruz, M.; Gonzalez-Moro, I.; Olazagoitia-Garmendia, A.; Castellanos-Rubio, A.; Santin, I. Non-Coding RNA The Role of LncRNAs in Gene Expression Regulation through MRNA Stabilization. Non-Coding RNA 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, Y.; Zhu, W.; Liu, J.; Cheng, F. Non-Coding RNAs Fine-Tune the Balance between Plant Growth and Abiotic Stress Tolerance. Front. Plant Sci. 2022, 13, 965745. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Sze, C.C.; Li, M.W.; Lam, H.M. Roles of Non-Coding RNAs in the Hormonal and Nutritional Regulation in Nodulation and Nitrogen Fixation. Front. Plant Sci. 2022, 13, 997037. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, W.; Zhao, M.; Li, Z.; Wang, G.L.; Liu, W.; Liang, C. Transcriptome Analysis Reveals a LncRNA-MiRNA-MRNA Regulatory Network in OsRpp30-Mediated Disease Resistance in Rice. BMC Genom. 2023, 24, 643. [Google Scholar] [CrossRef] [PubMed]
- Hüttenhofer, A.; Vogel, J. Experimental Approaches to Identify Non-Coding RNAs. Nucleic Acids Res. 2006, 34, 635–646. [Google Scholar] [CrossRef]
- Singh, U.; Khemka, N.; Rajkumar, M.S.; Garg, R.; Jain, M. PLncPRO for Prediction of Long Non-Coding RNAs (LncRNAs) in Plants and Its Application for Discovery of Abiotic Stress-Responsive LncRNAs in Rice and Chickpea. Nucleic Acids Res. 2017, 45, e183. [Google Scholar] [CrossRef] [PubMed]
- Bhogireddy, S.; Mangrauthia, S.K.; Kumar, R.; Pandey, A.K.; Singh, S.; Jain, A.; Budak, H.; Varshney, R.K.; Kudapa, H. Regulatory Non-Coding RNAs: A New Frontier in Regulation of Plant Biology. Funct. Integr. Genom. 2021, 21, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Wang, X.; Duan, J.; Ma, J. Rhizobial TRNA-Derived Small RNAs Are Signal Molecules Regulating Plant Nodulation. Science 2019, 365, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Choudury, S.G.; Slotkin, R.K. TRNA-Derived Small RNAs Target Transposable Element Transcripts. Nucleic Acids Res. 2017, 45, 5142–5152. [Google Scholar] [CrossRef]
- Voinnet, O. Origin, Biogenesis, and Activity of Plant MicroRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Brosnan, C.A.; Voinnet, O. Cell-to-Cell and Long-Distance SiRNA Movement in Plants: Mechanisms and Biological Implications. Curr. Opin. Plant Biol. 2011, 14, 580–587. [Google Scholar] [CrossRef]
- Patil, V.S.; Zhou, R.; Rana, T.M. Gene Regulation by Non-Coding RNAs. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 16–32. [Google Scholar] [CrossRef]
- Mallory, A.C.; Vaucheret, H. Functions of Micrornas and Related Small a Rnas in Plants. Nat. Genet. 2006, 38, S31–S36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xia, J.; Lii, Y.E.; Barrera-Figueroa, B.E.; Zhou, X.; Gao, S.; Lu, L.; Niu, D.; Chen, Z.; Leung, C.; et al. Genome-Wide Analysis of Plant Nat-SiRNAs Reveals Insights into Their Distribution, Biogenesis and Function. Genome Biol. 2012, 13, R20. [Google Scholar] [CrossRef]
- Sanan-Mishra, N.; Abdul Kader Jailani, A.; Mandal, B.; Mukherjee, S.K. Secondary SiRNAs in Plants: Biosynthesis, Various Functions, and Applications in Virology. Front. Plant Sci. 2021, 12, 610283. [Google Scholar] [CrossRef]
- Zhang, X.; Lii, Y.; Wu, Z.; Polishko, A.; Zhang, H.; Chinnusamy, V.; Lonardi, S.; Zhu, J.K.; Liu, R.; Jin, H. Mechanisms of Small RNA Generation from Cis-NATs in Response to Environmental and Developmental Cues. Mol. Plant 2013, 6, 704–715. [Google Scholar] [CrossRef]
- Liu, Y.; Teng, C.; Xia, R.; Meyers, B.C. PhasiRNAs in Plants: Their Biogenesis, Genic Sources, and Roles in Stress Responses, Development, and Reproduction. Plant Cell 2020, 32, 3059–3080. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Muhammad, S.; Cao, M.; Wu, L. Biogenesis and Regulatory Hierarchy of Phased Small Interfering RNAs in Plants. Plant Biotechnol. J. 2018, 16, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tao, X.; Li, M.; Gao, M.; Chen, J.; Zhou, N.; Mei, G.; Fang, L.; Ding, L.; Zhou, B.; et al. Role of PhasiRNAs from Two Distinct Phasing Frames of GhMYB2 Loci in Cis- Gene Regulation in the Cotton Genome. BMC Plant Biol. 2020, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Barber, W.T.; Zhang, W.; Win, H.; Varala, K.K.; Dorweiler, J.E.; Hudson, M.E.; Moose, S.P. Repeat Associated Small RNAs Vary among Parents and Following Hybridization in Maize. Proc. Natl. Acad. Sci. USA 2012, 109, 10444–10449. [Google Scholar] [CrossRef] [PubMed]
- Katiyar-Agarwal, S.; Gao, S.; Vivian-Smith, A.; Jin, H. A Novel Class of Bacteria-Induced Small RNAs in Arabidopsis. Genes Dev. 2007, 21, 3123–3134. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. MicroRNAs and Developmental Timing. Curr. Opin. Genet. Dev. 2011, 21, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of MiRNAs and SiRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Huntzinger, E.; Izaurralde, E. Getting to the Root of MiRNA-Mediated Gene Silencing. Cell 2008, 132, 9–14. [Google Scholar] [CrossRef]
- Yadav, A.; Sanyal, I.; Rai, S.P.; Lata, C. An Overview on MiRNA-Encoded Peptides in Plant Biology Research. Genomics 2021, 113, 2385–2391. [Google Scholar] [CrossRef]
- Feng, Y.Z.; Zhu, Q.F.; Xue, J.; Chen, P.; Yu, Y. Shining in the Dark: The Big World of Small Peptides in Plants. aBIOTECH 2023, 4, 238–256. [Google Scholar] [CrossRef]
- Lauressergues, D.; Couzigou, J.M.; San Clemente, H.; Martinez, Y.; Dunand, C.; Bécard, G.; Combier, J.P. Primary Transcripts of MicroRNAs Encode Regulatory Peptides. Nature 2015, 520, 90–93. [Google Scholar] [CrossRef]
- Chen, Q.J.; Deng, B.H.; Gao, J.; Zhao, Z.Y.; Chen, Z.L.; Song, S.R.; Wang, L.; Zhao, L.P.; Xu, W.P.; Zhang, C.X.; et al. A Mirna-Encoded Small Peptide, Vvi-MiPEP171d1, Regulates Adventitious Root Formation. Plant Physiol. 2020, 183, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Badola, P.K.; Bhatia, C.; Sharma, D.; Trivedi, P.K. Primary Transcript of MiR858 Encodes Regulatory Peptide and Controls Flavonoid Biosynthesis and Development in Arabidopsis. Nat. Plants 2020, 6, 1262–1274. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Stellwag, E.J. Identification of Soybean MicroRNAs and Their Targets. Planta 2008, 229, 161–182. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Gu, Y.; Zhang, Z.; Tang, G. The Interaction between MiR160 and MiR165/166 in the Control of Leaf Development and Drought Tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. MicroRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 824240. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2017; Volume 1008, pp. 133–154. [Google Scholar]
- Wierzbicki, A.T.; Blevins, T.; Swiezewski, S. Annual Review of Plant Biology Long Noncoding RNAs in Plants. Annu. Rev. Plant Biol. 2021, 72, 2021. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, Q.H.; Kaufmann, K. Long Non-Coding RNAs in Plants: Emerging Modulators of Gene Activity in Development and Stress Responses. Planta 2020, 252, 92. [Google Scholar] [CrossRef]
- Bouba, I.; Kang, Q.; Luan, Y.S.; Meng, J. Predicting MiRNA-LncRNA Interactions and Recognizing Their Regulatory Roles in Stress Response of Plants. Math. Biosci. 2019, 312, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Li, A.; Yu, B.; Li, S. Interplay between MiRNAs and LncRNAs: Mode of Action and Biological Roles in Plant Development and Stress Adaptation. Comput. Struct. Biotechnol. J. 2021, 19, 2567–2574. [Google Scholar] [CrossRef]
- Kouchi, H.; Hata, S. Isolation and Characterization of Novel Nodulin CDNAs Representing Genes Expressed at Early Stages of Soybean Nodule Development. Mol. Gen. Genet. 1993, 238, 106–119. [Google Scholar] [CrossRef]
- Waseem, M.; Liu, Y.; Xia, R. Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants. Int. J. Mol. Sci. 2021, 22, 86. [Google Scholar] [CrossRef] [PubMed]
- Kindgren, P.; Ard, R.; Ivanov, M.; Marquardt, S. Transcriptional Read-through of the Long Non-Coding RNA SVALKA Governs Plant Cold Acclimation. Nat. Commun. 2018, 9, 4561. [Google Scholar] [CrossRef] [PubMed]
- Fedak, H.; Palusinska, M.; Krzyczmonik, K.; Brzezniak, L.; Yatusevich, R.; Pietras, Z.; Kaczanowski, S.; Swiezewski, S. Control of Seed Dormancy in Arabidopsis by a Cis-Acting Noncoding Antisense Transcript. Proc. Natl. Acad. Sci. USA 2016, 113, E7846–E7855. [Google Scholar] [CrossRef]
- Heo, J.B.; Sung, S. Vernalization-Mediated Epigenetic Silencing by a Long Intronic Noncoding RNA. Science 2011, 1197349, 331. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, S. Vernalization-Triggered Intragenic Chromatin Loop Formation by Long Noncoding RNAs. Dev. Cell 2017, 40, 302–312.e4. [Google Scholar] [CrossRef]
- Hawkes, E.J.; Hennelly, S.P.; Novikova, I.V.; Irwin, J.A.; Dean, C.; Sanbonmatsu, K.Y. COOLAIR Antisense RNAs Form Evolutionarily Conserved Elaborate Secondary Structures. Cell Rep. 2016, 16, 3087–3096. [Google Scholar] [CrossRef]
- Liu, G.; Liu, F.; Wang, Y.; Liu, X. A Novel Long Noncoding RNA CIL1 Enhances Cold Stress Tolerance in Arabidopsis. Plant Sci. 2022, 323, 111370. [Google Scholar] [CrossRef]
- Cui, J. Editorial: LncRNA in Plants: Function, Mechanisms and Applications. Front. Plant Sci. 2023, 14, 1238185. [Google Scholar] [CrossRef]
- Wen, J.; Lease, K.A.; Walker, J.C. DVL, a Novel Class of Small Polypeptides: Overexpression Alters Arabidopsis Development. Plant J. 2004, 37, 668–677. [Google Scholar] [CrossRef]
- Narita, N.N.; Moore, S.; Horiguchi, G.; Kubo, M.; Demura, T.; Fukuda, H.; Goodrich, J.; Tsukaya, H. Overexpression of a Novel Small Peptide Rotundifolia4 Decreases Cell Proliferation and Alters Leaf Shape in Arabidopsis Thaliana. Plant J. 2004, 38, 699–713. [Google Scholar] [CrossRef]
- Chilley, P.M.; Casson, S.A.; Tarkowski, P.; Hawkins, N.; Wang, K.L.C.; Hussey, P.J.; Beale, M.; Ecker, J.R.; Sandberg, G.K.; Lindsey, K. The POLARIS Peptide of Arabidopsis Regulates Auxin Transport Root Growth via Effects on Ethylene Signaling. Plant Cell 2006, 18, 3058–3072. [Google Scholar] [CrossRef]
- Lin, X.; Lin, W.; Ku, Y.S.; Wong, F.L.; Li, M.W.; Lam, H.M.; Ngai, S.M.; Chan, T.F. Analysis of Soybean Long Non-Coding RNAs Reveals a Subset of Small Peptide-Coding Transcripts1[OPEN]. Plant Physiol. 2020, 182, 1359–1374. [Google Scholar] [CrossRef]
- Zhao, X.; Lan, Y.; Chen, D. Exploring Long Non-Coding RNA Networks from Single Cell Omics Data. Comput. Struct. Biotechnol. J. 2022, 20, 4381–4389. [Google Scholar] [CrossRef]
- He, Z.; Luo, Y.; Zhou, X.; Zhu, T.; Lan, Y.; Chen, D. ScPlantDB: A Comprehensive Database for Exploring Cell Types and Markers of Plant Cell Atlases. Nucleic Acids Res. 2023, 52, D1629–D1638. [Google Scholar] [CrossRef]
- Salzman, J. Circular RNA Expression: Its Potential Regulation and Function. Trends Genet. 2016, 32, 309–316. [Google Scholar] [CrossRef]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, Present, and Future of Circ RNA s. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Liu, S.; Guo, X.Y.; Shang, Q.J.; Gao, P. The Biogenesis, Biological Functions and Modification of Circular RNAs. Exp. Mol. Pathol. 2023, 131, 104861. [Google Scholar] [CrossRef]
- Feng, X.Y.; Zhu, S.X.; Pu, K.J.; Huang, H.J.; Chen, Y.Q.; Wang, W.T. New Insight into CircRNAs: Characterization, Strategies, and Biomedical Applications. Exp. Hematol. Oncol. 2023, 12, 91. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, Y.; Jia, E.; Liu, Z.; Pan, M.; Bai, Y.; Zhao, X.; Ge, Q. Comparative Analysis of Circular RNA Enrichment Methods. RNA Biol. 2022, 19, 55–67. [Google Scholar] [CrossRef]
- Long, F.; Lin, Z.; Li, L.; Ma, M.; Lu, Z.; Jing, L.; Li, X.; Lin, C. Comprehensive Landscape and Future Perspectives of Circular RNAs in Colorectal Cancer. Mol. Cancer 2021, 20, 26. [Google Scholar] [CrossRef]
- Ye, C.Y.; Zhang, X.; Chu, Q.; Liu, C.; Yu, Y.; Jiang, W.; Zhu, Q.H.; Fan, L.; Guo, L. Full-Length Sequence Assembly Reveals Circular RNAs with Diverse Non-GT/AG Splicing Signals in Rice. RNA Biol. 2017, 14, 1055–1063. [Google Scholar] [CrossRef]
- Olesen, M.T.J.; Kristensen, L.S. Circular RNAs as MicroRNA Sponges: Evidence and Controversies. Essays Biochem. 2021, 65, 685–696. [Google Scholar]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Lukiw, W.J. Circular RNA (CircRNA) in Alzheimer’s Disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, S.; Liu, K. Structural Insights into PiRNA Biogenesis. Biochim. Biophys. Acta Gene Regul. Mech. 2022, 1865, 194799. [Google Scholar] [CrossRef]
- Huang, S.; Yoshitake, K.; Asakawa, S. A Review of Discovery Profiling of Piwi-Interacting Rnas and Their Diverse Functions in Metazoans. Int. J. Mol. Sci. 2021, 22, 11166. [Google Scholar] [CrossRef]
- Tian, H.; Jia, Y.; Niu, T.; Yu, Q.; Ding, Z. The Key Players of the Primary Root Growth and Development Also Function in Lateral Roots in Arabidopsis. Plant Cell Rep. 2014, 33, 745–753. [Google Scholar] [CrossRef]
- Smet, W.; Blilou, I. A Blast from the Past: Understanding Stem Cell Specification in Plant Roots Using Laser Ablation. Quant. Plant Biol. 2023, 4, e14. [Google Scholar] [CrossRef] [PubMed]
- Jansen, L.; Roberts, I.; de Rycke, R.; Beeckman, T. Phloem-Associated Auxin Response Maxima Determine Radial Positioning of Lateral Roots in Maize. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1525–1533. [Google Scholar] [CrossRef]
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Gautam, V.; Singh, A.; Yadav, S.; Singh, S.; Kumar, P.; Das, S.S.; Sarkar, A.K. Conserved LBL1-Ta-SiRNA and MiR165/166-RLD1/2 Modules Regulate Root Development in Maize. Development 2021, 148, dev190033. [Google Scholar] [CrossRef]
- Barrera-Rojas, C.H.; Rocha, G.H.B.; Polverari, L.; Di Pinheiro Brito, A.; Di Batista, S.; Notini, M.M.; Da Cruz, A.C.F.; Morea, E.G.O.; Sabatini, S.; Otoni, W.C.; et al. MiR156-Targeted SPL10 Controls Arabidopsis Root Meristem Activity and Root-Derived de Novo Shoot Regeneration via Cytokinin Responses. J. Exp. Bot. 2020, 71, 934–950. [Google Scholar] [CrossRef]
- Carlsbecker, A.; Lee, J.Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vatén, A.; Thitamadee, S.; et al. Cell Signalling by MicroRNA165/6 Directs Gene Dose-Dependent Root Cell Fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Xie, Y.; Guo, F.; Han, N.; Ma, S.; Zeng, Z.; Wang, J.; Yang, Y.; Zhu, M. Distinctive Expression Patterns and Roles of the MiRNA393/TIR1 Homolog Module in Regulating Flag Leaf Inclination and Primary and Crown Root Growth in Rice (Oryza Sativa). New Phytol. 2012, 196, 149–161. [Google Scholar] [CrossRef]
- Ariel, F.; Lucero, L.; Christ, A.; Mammarella, M.F.; Jegu, T.; Veluchamy, A.; Mariappan, K.; Latrasse, D.; Blein, T.; Liu, C.; et al. R-Loop Mediated Trans Action of the APOLO Long Noncoding RNA. Mol. Cell 2020, 77, 1055–1065.e4. [Google Scholar] [CrossRef]
- Rigo, R.; Bazin, J.; Romero-Barrios, N.; Moison, M.; Lucero, L.; Christ, A.; Benhamed, M.; Blein, T.; Huguet, S.; Charon, C.; et al. The Arabidopsis Lnc RNA ASCO Modulates the Transcriptome through Interaction with Splicing Factors. EMBO Rep. 2020, 21, e48977. [Google Scholar] [CrossRef]
- Moison, M.; Pacheco, J.M.; Lucero, L.; Fonouni-Farde, C.; Rodríguez-Melo, J.; Mansilla, N.; Christ, A.; Bazin, J.; Benhamed, M.; Ibañez, F.; et al. The LncRNA APOLO Interacts with the Transcription Factor WRKY42 to Trigger Root Hair Cell Expansion in Response to Cold. Mol. Plant 2021, 14, 937–948. [Google Scholar] [CrossRef]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.S.; Crespi, M. Long Noncoding RNA Modulates Alternative Splicing Regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef]
- Bazin, J.; Romero, N.; Rigo, R.; Charon, C.; Blein, T.; Ariel, F.; Crespi, M. Nuclear Speckle Rna Binding Proteins Remodel Alternative Splicing and the Non-Coding Arabidopsis Transcriptome to Regulate a Cross-Talk between Auxin and Immune Responses. Front. Plant Sci. 2018, 9, 1209. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.M.; Mansilla, N.; Moison, M.; Lucero, L.; Gabarain, V.B.; Ariel, F.; Estevez, J.M. The LncRNA APOLO and the Transcription Factor WRKY42 Target Common Cell Wall EXTENSIN Encoding Genes to Trigger Root Hair Cell Elongation. Plant Signal. Behav. 2021, 16, 1920191. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, Y.; Lin, T.; Cui, D. Identification and Characterization of CircRNAs Involved in the Regulation of Wheat Root Length. Biol. Res. 2019, 52, 19. [Google Scholar] [CrossRef]
- Zhang, D.-J.; Yang, Y.-J.; Liu, C.-Y.; Zhang, F.; Wu, Q.-S. Root Hair Growth and Development in Response to Nutrients and Phytohormones. Root Biol. 2018, 52, 65–84. [Google Scholar]
- Ran, N.; Liu, S.; Qi, H.; Wang, J.; Shen, T.; Xu, W.; Xu, M. Long Non-Coding RNA LncWOX11a Suppresses Adventitious Root Formation of Poplar by Regulating the Expression of PeWOX11a. Int. J. Mol. Sci. 2023, 24, 5766. [Google Scholar] [CrossRef]
- Sundararajan, S.; Sivakumar, H.P.; Rajendran, V.; Kumariah, M.; Ramalingam, S. Adventitious Roots in Rice, the Model Cereal: Genetic Factors and the Influence of Environmental Cues—A Mini Review. Plant Cell Tissue Organ Cult. 2023, 154, 1–12. [Google Scholar] [CrossRef]
- Liu, R.; Wen, S.S.; Sun, T.T.; Wang, R.; Zuo, W.T.; Yang, T.; Wang, C.; Hu, J.J.; Lu, M.Z.; Wang, L.Q. PagWOX11/12a Positively Regulates the PagSAUR36 Gene That Enhances Adventitious Root Development in Poplar. J. Exp. Bot. 2022, 73, 7298–7311. [Google Scholar] [CrossRef]
- Qi, H.; Wu, L.; Shen, T.; Liu, S.; Cai, H.; Ran, N.; Wang, J.; Xu, M. Overexpression of the Long Non-Coding RNA LncWOX5 Negatively Regulates the Development of Adventitious Roots in Populus. Ind. Crops Prod. 2023, 192, 116054. [Google Scholar] [CrossRef]
- Paul Alvarez, J.; Furumizu, C.; Efroni, I.; Eshed, Y.; Bowman, J.L. Active Suppression of a Leaf Meristem Orchestrates Determinate Leaf Growth. eLife 2016, 5, e15023. [Google Scholar] [CrossRef]
- Yang, T.; Wang, Y.; Teotia, S.; Zhang, Z.; Tang, G. The Making of Leaves: How Small RNA Networks Modulate Leaf Development. Front. Plant Sci. 2018, 9, 824. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Nogueira, F.T.; Howell, M.D.; Montgomery, T.A.; Carrington, J.C.; Timmermans, M.C. Pattern formation via small RNA mobility. Genes Dev. 2009, 1, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Schwab, R.; Maizel, A.; Ruiz-Ferrer, V.; Garcia, D.; Bayer, M.; Crespi, M.; Voinnet, O.; Martienssen, R.A. Endogenous TasiRNAs Mediate Non-Cell Autonomous Effects on Gene Regulation in Arabidopsis thaliana. PLoS ONE 2009, 4, e5980. [Google Scholar] [CrossRef] [PubMed]
- Dotto, M.C.; Petsch, K.A.; Aukerman, M.J.; Beatty, M.; Hammell, M.; Timmermans, M.C.P. Genome-Wide Analysis of Leafbladeless1-Regulated and Phased Small RNAs Underscores the Importance of the TAS3 Ta-SiRNA Pathway to Maize Development. PLoS Genet. 2014, 10, e1004826. [Google Scholar] [CrossRef] [PubMed]
- Bresso, E.G.; Chorostecki, U.; Rodriguez, R.E.; Palatnik, J.F.; Schommer, C. Spatial Control of Gene Expression by MiR319-Regulated TCP Transcription Factors in Leaf Development. Plant Physiol. 2018, 176, 1694–1708. [Google Scholar] [CrossRef]
- Gupta, M.D.; Nathut, U. Divergence in Patterns of Leaf Growth Polarity Is Associated with the Expression Divergence of MiR396. Plant Cell 2015, 27, 2785–2799. [Google Scholar] [CrossRef]
- Husbands, A.Y.; Chitwood, D.H.; Plavskin, Y.; Timmermans, M.C.P. Signals and Prepatterns: New Insights into Organ Polarity in Plants. Genes Dev. 2009, 23, 1986–1997. [Google Scholar] [CrossRef]
- Schommer, C.; Debernardi, J.M.; Bresso, E.G.; Rodriguez, R.E.; Palatnik, J.F. Repression of Cell Proliferation by MiR319-Regulated TCP4. Mol. Plant 2014, 7, 1533–1544. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Zhang, D.; Yin, D.; Zhao, Y.; Ji, C.; Zhao, X.; Li, X.; He, Q.; Chen, R.; et al. A Novel Antisense Long Noncoding RNA, TWISTED LEAF, Maintains Leaf Blade Flattening by Regulating Its Associated Sense R2R3-MYB Gene in Rice. New Phytol. 2018, 218, 774–788. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, J.; Kang, M.H.; Trang, T.T.M.; Lee, J.; Lee, H.; Jeong, H.; Lim, P.O. Dynamic Landscape of Long Noncoding RNAs during Leaf Aging in Arabidopsis. Front. Plant Sci. 2022, 13, 1068163. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Chen, G.; Shi, T. Identifying and Characterizing the Circular RNAs during the Lifespan of Arabidopsis Leaves. Front. Plant Sci. 2017, 8, 1278. [Google Scholar] [CrossRef]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial Patterning of Arabidopsis Shoots by Class III HD-ZIP and KANADI Genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef]
- Liu, D.; Song, Y.; Chen, Z.; Yu, D. Ectopic Expression of MiR396 Suppresses GRF Target Gene Expression and Alters Leaf Growth in Arabidopsis. Physiol. Plant. 2009, 136, 223–236. [Google Scholar] [CrossRef]
- Nikovics, K.; Blein, T.; Peaucelle, A.; Ishida, T.; Morin, H.; Aida, M.; Laufs, P. The Balance between the MIR164A and CUC2 Genes Controls Leaf Margin Serration in Arabidopsis. Plant Cell 2006, 18, 2929–2945. [Google Scholar] [CrossRef]
- Sieber, P.; Wellmer, F.; Gheyselinck, J.; Riechmann, J.L.; Meyerowitz, E.M. Redundancy and Specialization among Plant MicroRNAs: Role of the MIR164 Family in Developmental Robustness. Development 2007, 134, 1051–1060. [Google Scholar] [CrossRef]
- Zhou, G.K.; Kubo, M.; Zhong, R.; Demura, T.; Ye, Z.H. Overexpression of MiR165 Affects Apical Meristem Formation, Organ Polarity Establishment and Vascular Development in Arabidopsis. Plant Cell Physiol. 2007, 48, 391–404. [Google Scholar] [CrossRef]
- Merelo, P.; Ram, H.; Caggiano, M.P.; Ohno, C.; Ott, F.; Straub, D.; Graeff, M.; Cho, S.K.; Yang, S.W.; Wenkel, S.; et al. Regulation of MIR165/166 by Class II and Class III Homeodomain Leucine Zipper Proteins Establishes Leaf Polarity. Proc. Natl. Acad. Sci. USA 2016, 113, 11973–11978. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X. Argonautes Compete for MiR165/166 to Regulate Shoot Apical Meristem Development. Curr. Opin. Plant Biol. 2012, 15, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, H. The Role of HD-ZIP III Transcription Factors and MiR165/166 in Vascular Development and Secondary Cell Wall Formation. Plant Signal. Behav. 2015, 10, e1078955. [Google Scholar] [CrossRef] [PubMed]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. Roles of MiR319 and TCP Transcription Factors in Leaf Development. Plant Physiol. 2017, 175, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of Leaf Morphogenesis by MicroRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Efroni, I.; Blum, E.; Goldshmidt, A.; Eshed, Y. A Protracted and Dynamic Maturation Schedule Underlies Arabidopsis Leaf Development. Plant Cell 2008, 20, 2293–2306. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A Small Transcription Factor Family with Important Functions in Plant Biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef]
- Wang, L.; Tian, Y.; Shi, W.; Yu, P.; Hu, Y.; Lv, J.; Fu, C.; Fan, M.; Bai, M.Y. The MiR396-GRFs Module Mediates the Prevention of Photo-Oxidative Damage by Brassinosteroids during Seedling de-Etiolation in Arabidopsis. Plant Cell 2020, 32, 2525–2542. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Mecchia, M.A.; Vercruyssen, L.; Smaczniak, C.; Kaufmann, K.; Inze, D.; Rodriguez, R.E.; Palatnik, J.F. Post-Transcriptional Control of GRF Transcription Factors by MicroRNA MiR396 and GIF Co-Activator Affects Leaf Size and Longevity. Plant J. 2014, 79, 413–426. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Lin, X.; Hong, X.; Zhu, Y.; Li, W.; He, W.; An, F.; Guo, H. Adenine Phosphoribosyl Transferase 1 Is a Key Enzyme Catalyzing Cytokinin Conversion from Nucleobases to Nucleotides in Arabidopsis. Mol. Plant 2013, 6, 1661–1672. [Google Scholar] [CrossRef]
- Li, W.X.; Oono, Y.; Zhu, J.; He, X.J.; Wu, J.M.; Iida, K.; Lu, X.Y.; Cui, X.; Jin, H.; Zhu, J.K. The Arabidopsis NFYA5 Transcription Factor Is Regulated Transcriptionally and Posttranscriptionally to Promote Drought Resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef]
- Li, G.; Zhang, H.; Li, J.; Zhang, Z.; Li, Z. Genetic Control of Panicle Architecture in Rice. Crop. J. 2021, 9, 590–597. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, C.; Baulcombe, D.C.; Chen, Z.J. Maternal SiRNAs as Regulators of Parental Genome Imbalance and Gene Expression in Endosperm of Arabidopsis Seeds. Proc. Natl. Acad. Sci. USA 2012, 109, 5529–5534. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, S.; Jiao, W.; Wang, L.; Wang, L.; Ye, W.; Lu, J.; Hong, D.; You, S.; Cheng, Z.; et al. Both Maternally and Paternally Imprinted Genes Regulate Seed Development in Rice. New Phytol. 2017, 216, 373–387. [Google Scholar] [CrossRef]
- Khemka, N.; Singh Rajkumar, M.; Garg, R.; Jain, M. Genome-Wide Profiling of MiRNAs during Seed Development Reveals Their Functional Relevance in Seed Size/Weight Determination in Chickpea. Plant Direct 2021, 5, e00299. [Google Scholar] [CrossRef] [PubMed]
- Miao, C.; Wang, Z.; Zhang, L.; Yao, J.; Hua, K.; Liu, X.; Shi, H.; Zhu, J.K. The Grain Yield Modulator MiR156 Regulates Seed Dormancy through the Gibberellin Pathway in Rice. Nat. Commun. 2019, 10, 3822. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, X.; Sun, F.; Hu, J.; Zha, X.; Su, W.; Yang, J. Overexpressing LncRNA LAIR Increases Grain Yield and Regulates Neighbouring Gene Cluster Expression in Rice. Nat. Commun. 2018, 9, 3516. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Liu, Y.; Wang, W.; Wu, L.; Ma, J.; Zhang, S.; Wang, J.; Feng, F.; Yuan, H.; Huang, X. Identification and Functional Prediction of CircRNAs of Developing Seeds in High Oleic Acid Sunflower (Helianthus annuus L.). Acta Physiol. Plant 2023, 45, 13. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Qu, L.H.; Shu, W.S.; Chen, Y.Q. Genome-Wide Screening and Functional Analysis Identify a Large Number of Long Noncoding RNAs Involved in the Sexual Reproduction of Rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef]
- Cao, P.; Fan, W.; Li, P.; Hu, Y. Genome-Wide Profiling of Long Noncoding RNAs Involved in Wheat Spike Development. BMC Genom. 2021, 22, 493. [Google Scholar] [CrossRef]
- He, G.; Luo, X.; Tian, F.; Li, K.; Zhu, Z.; Su, W.; Qian, X.; Fu, Y.; Wang, X.; Sun, C.; et al. Haplotype Variation in Structure and Expression of a Gene Cluster Associated with a Quantitative Trait Locus for Improved Yield in Rice. Genome Res. 2006, 16, 618–626. [Google Scholar] [CrossRef]
- Yang, G.; Deng, P.; Guo, Q.; Shi, T.; Pan, W.; Cui, L.; Liu, X.; Nie, X. Population Transcriptomic Analysis Identifies the Comprehensive LncRNAs Landscape of Spike in Wheat (Triticum aestivum L.). BMC Plant Biol. 2022, 22, 450. [Google Scholar] [CrossRef]
- Sagun, J.V.; Yadav, U.P.; Alonso, A.P. Progress in Understanding and Improving Oil Content and Quality in Seeds. Front. Plant Sci. 2023, 14, 1116894. [Google Scholar] [CrossRef]
- Bhati, K.K.; Riyazuddin, R.; Pathak, A.; Singh, A. The survey of genetic engineering approaches for oil/fatty acid content improvement in oilseed crops. In Genome Engineering for Crop Improvement; Upadhyay, S.K., Ed.; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar] [CrossRef]
- Shen, E.; Zhu, X.; Hua, S.; Chen, H.; Ye, C.; Zhou, L.; Liu, Q.; Zhu, Q.H.; Fan, L.; Chen, X. Genome-Wide Identification of Oil Biosynthesis-Related Long Non-Coding RNAs in Allopolyploid Brassica Napus 06 Biological Sciences 0604 Genetics 06 Biological Sciences 0607 Plant Biology. BMC Genom. 2018, 19, 745. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, A.; Zhao, Q.; Li, Z.; Lamboro, A.; He, H.; Li, Y.; Jiao, S.; Guan, S.; Liu, S.; et al. Genome-Wide Identification and Analysis of Long Non-Coding RNAs Involved in Fatty Acid Biosynthesis in Young Soybean Pods. Sci. Rep. 2021, 11, 7603. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, D.; Li, X.; Zhou, B.; Chang, T.; Hong, B.; Guan, C.; Guan, M. Integrated Analysis of LncRNA–MRNA Regulatory Networks Related to Lipid Metabolism in High-Oleic-Acid Rapeseed. Int. J. Mol. Sci. 2023, 24, 6277. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, Z.; Zeng, C.; Xiao, M.; Lin, S.; Yao, W.; Li, Q.; Guo, L.; Lu, S. Regulation of Seed Oil Accumulation by LncRNAs in Brassica Napus. Biotechnol. Biofuels Bioprod. 2023, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Mathan, J.; Yadav, A.; Goyal, A.K.; Chaudhury, A. Molecular and Transcriptional Regulation of Seed Development in Cereals: Present Status and Future Prospects; InTech Open: London, UK, 2021. [Google Scholar]
- Yoon, J.; Min, C.W.; Kim, J.; Baek, G.; Kim, D.; Jang, J.W.; Gupta, R.; Kim, S.T.; Cho, L.H. Quantitative Proteomic Analysis Deciphers the Molecular Mechanism for Endosperm Nuclear Division in Early Rice Seed Development. Plants 2023, 12, 3715. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Katoh, H.; Ogawa, D.; Kagaya, Y.; Sato, Y.; Kitano, H.; Nagato, Y.; Ishikawa, R.; Ono, A.; Kinoshita, T.; et al. Rice SNF2 Family Helicase ENL1 Is Essential for Syncytial Endosperm Development. Plant J. 2015, 81, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.F.; Zhang, Y.C.; Sun, Y.M.; Yu, Y.; Lei, M.Q.; Yang, Y.W.; Lian, J.P.; Feng, Y.Z.; Zhang, Z.; Yang, L.; et al. The Parent-of-Origin LncRNA MISSEN Regulates Rice Endosperm Development. Nat. Commun. 2021, 12, 6525. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Yadav, A.; Roy, J.K.; Bhati, K.K. Role of genome engineering for the development of resistant starch-rich, allergen-free and processing quality improved cereal crops. In Genome Engineering for Crop Improvement; Upadhyay, S.K., Ed.; Wiley: Hoboken, NJ, USA, 2021. [Google Scholar]
- Mishra, A.; Singh, A.; Sharma, M.; Kumar, P.; Roy, J. Development of EMS-Induced Mutation Population for Amylose and Resistant Starch Variation in Bread Wheat (Triticum Aestivum) and Identification of Candidate Genes Responsible for Amylose Variation. BMC Plant Biol. 2016, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Madhawan, A.; Sharma, A.; Bhandawat, A.; Rahim, M.S.; Kumar, P.; Mishra, A.; Parveen, A.; Sharma, H.; Verma, S.K.; Roy, J. Identification and Characterization of Long Non-Coding RNAs Regulating Resistant Starch Biosynthesis in Bread Wheat (Triticum aestivum L.). Genomics 2020, 112, 3065–3074. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R.; Meng, F.; Chen, Y.; Wang, W.; Yang, K.; Gao, Y.; Xin, M.; Du, J.; Hu, Z.; et al. A Comprehensive Atlas of Long Non-Coding RNAs Provides Insight into Grain Development in Wheat. Seed Biol. 2023, 2, 12. [Google Scholar] [CrossRef]
- Khalid, A.; Hameed, A.; Tahir, M.F. Wheat Quality: A Review on Chemical Composition, Nutritional Attributes, Grain Anatomy, Types, Classification, and Function of Seed Storage Proteins in Bread Making Quality. Front. Nutr. 2023, 10, 1053196. [Google Scholar] [CrossRef]
- Collin, A.; Daszkowska-Golec, A.; Szarejko, I. Updates on the Role of ABSCISIC ACID INSENSITIVE 5 (ABI5) and ABSCISIC ACID-RESPONSIVE ELEMENT BINDING FACTORS (ABFs) in ABA Signaling in Different Developmental Stages in Plants. Cells 2021, 10, 1996. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, J.; Weng, H.; Yuan, X.; Xiao, W.; Wang, H. A Long Noncoding RNA Derived from LncRNA–MRNA Networks Modulates Seed Vigor. Int. J. Mol. Sci. 2022, 23, 9472. [Google Scholar] [CrossRef]
- Hazra, S.; Moulick, D.; Mukherjee, A.; Sahib, S.; Chowardhara, B.; Majumdar, A.; Upadhyay, M.K.; Yadav, P.; Roy, P.; Santra, S.C.; et al. Evaluation of Efficacy of Non-Coding RNA in Abiotic Stress Management of Field Crops: Current Status and Future Prospective. Plant Physiol. Biochem. 2023, 203, 107940. [Google Scholar] [CrossRef]
- Yang, H.; Cui, Y.; Feng, Y.; Hu, Y.; Liu, L.; Duan, L. Long Non-Coding RNAs of Plants in Response to Abiotic Stresses and Their Regulating Roles in Promoting Environmental Adaption. Cells 2023, 12, 729. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhao, F.; Zhou, B. The Characters of Non-Coding RNAs and Their Biological Roles in Plant Development and Abiotic Stress Response. Int. J. Mol. Sci. 2022, 23, 4124. [Google Scholar] [CrossRef] [PubMed]
- Gelaw, T.A.; Sanan-Mishra, N. Non-Coding RNAs in Response to Drought Stress. Int. J. Mol. Sci. 2021, 22, 12519. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.; Ahn, H.; Shin, S.J.; Kim, J.; Kwon, H.B.; Jung, W.; Kim, S. Clustering and evolutionary analysis of small RNAs identify regulatory siRNA clusters induced under drought stress in rice. BMC Syst. Biol. 2016, 23, 115. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ni, Z.; Peng, H.; Sun, F.; Xin, M.; Sunkar, R.; Zhu, J.K.; Sun, Q. Non-coding small RNAs responsive to abiotic stress in wheat (Triticum aestivum L.). Funct. Integr. Genom. 2010, 10, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Ma, R.; Wang, C.; Chen, N.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Integration of MRNA and MicroRNA Analysis Reveals the Molecular Mechanisms Underlying Drought Stress Tolerance in Maize (Zea mays L.). Front. Plant Sci. 2022, 13, 932667. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Burd, S.; Lers, A. MiR408 Is Involved in Abiotic Stress Responses in Arabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Balyan, S.; Kumar, M.; Mutum, R.D.; Raghuvanshi, U.; Agarwal, P.; Mathur, S.; Raghuvanshi, S. Identification of MiRNA-Mediated Drought Responsive Multi-Tiered Regulatory Network in Drought Tolerant Rice, Nagina. Sci. Rep. 2017, 7, 15446. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Liang, R.; Ge, L.; Li, W.; Xiao, H.; Lin, H.; Ruan, K.; Jin, Y. Identification of Drought-Induced MicroRNAs in Rice. Biochem. Biophys. Res. Commun. 2007, 354, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Hu, Z.; Jiang, Q.; Zhang, H. GmNFYA3, a Target Gene of MiR169, Is a Positive Regulator of Plant Tolerance to Drought Stress. Plant Mol. Biol. 2013, 82, 113–129. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome Profiling of Drought Responsive Noncoding RNAs and Their Target Genes in Rice. BMC Genom. 2016, 17, 563. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, S.R.; Danilevskaya, O.N.; Meng, X.; Beatty, M.; Zastrow-Hayes, G.; Harris, C.; Van Allen, B.; Habben, J.; Li, B. Genome-Wide Analysis of Alternative Splicing during Development and Drought Stress in Maize1[OPEN]. Plant Physiol. 2016, 170, 586–599. [Google Scholar] [CrossRef]
- Pang, J.; Zhang, X.; Ma, X.; Zhao, J. Spatio-Temporal Transcriptional Dynamics of Maize Long Non-Coding RNAs Responsive to Drought Stress. Genes 2019, 10, 138. [Google Scholar] [CrossRef]
- Qin, T.; Zhao, H.; Cui, P.; Albesher, N.; Xionga, L. A Nucleus-Localized Long Non-Coding Rna Enhances Drought and Salt Stress Tolerance. Plant Physiol. 2017, 175, 1321–1336. [Google Scholar] [CrossRef]
- Dong, S.-M.; Xiao, L.; Li, Z.-B.; Shen, J.; Yan, H.-B.; Li, S.-X.; Liao, W.-B.; Peng, M. A Novel Long Non-Coding RNA, DIR, Increases Drought Tolerance in Cassava by Modifying Stress-Related Gene Expression. J. Integr. Agric. 2022, 21, 2588–2602. [Google Scholar] [CrossRef]
- Zhang, P.; Fan, Y.; Sun, X.; Chen, L.; Terzaghi, W.; Bucher, E.; Li, L.; Dai, M. A Large-Scale Circular RNA Profiling Reveals Universal Molecular Mechanisms Responsive to Drought Stress in Maize and Arabidopsis. Plant J. 2019, 98, 697–713. [Google Scholar] [CrossRef]
- Ache, P.; Becker, D.; Ivashikina, N.; Dietrich, P.; Roelfsema, M.R.G.; Hedrich, R. GORK, a Delayed Outward Rectifier Expressed in Guard Cells of Arabidopsis Thaliana, Is a K+-Selective, K+-Sensing Ion Channel. FEBS Lett. 2000, 486, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Hoth, S.; Ache, P.; Wenkel, S.; Roelfsema, M.R.G.; Meyerhoff, O.; Hartung, W.; Hedrich, R. Regulation of the ABA-Sensitive Arabidopsis Potassium Channel Gene GORK in Response to Water Stress. FEBS Lett. 2003, 554, 119–126. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Plant Responses and Tolerance to Salt Stress: Physiological and Molecular Interventions. Int. J. Mol. Sci. 2022, 23, 15740. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Shobbar, Z.S.; Babaeian Jelodar, N.; Ghaffari, M.R.; Nematzadeh, G.A.; Asari, S. Dissecting Molecular Mechanisms Underlying Salt Tolerance in Rice: A Comparative Transcriptional Profiling of the Contrasting Genotypes. Rice 2019, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Wu, J.; Luo, Q.; Zhang, D.; Zhuang, W.; Xiao, G.; Deng, Q.; Bai, B. Effects of Salinity Stress at Reproductive Growth Stage on Rice (Oryza sativa L.) Composition, Starch Structure, and Physicochemical Properties. Front. Nutr. 2022, 9, 926217. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Agarwala, N.; Gill, S.S.; Varshney, R.K. Emerging Role of Plant Long Non Coding RNAs (LncRNAs) in Salinity Stress Response. Plant Stress 2023, 10, 100265. [Google Scholar] [CrossRef]
- Parmar, S.; Gharat, S.A.; Tagirasa, R.; Chandra, T.; Behera, L.; Dash, S.K.; Shaw, B.P. Identification and Expression Analysis of MiRNAs and Elucidation of Their Role in Salt Tolerance in Rice Varieties Susceptible and Tolerant to Salinity. PLoS ONE 2020, 15, e0230958. [Google Scholar] [CrossRef]
- Wang, Z.; Li, N.; Yu, Q.; Wang, H. Genome-Wide Characterization of Salt-Responsive Mirnas, Circrnas and Associated Cerna Networks in Tomatoes. Int. J. Mol. Sci. 2021, 22, 2238. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Guo, H.; Li, X.; Tang, M.; Guo, C.; Bao, H.; Huang, L.; Yi, Y.; Cui, Y.; Chen, L. OsSIDP301, a Member of the DUF1644 Family, Negatively Regulates Salt Stress and Grain Size in Rice. Front. Plant Sci. 2022, 13, 863233. [Google Scholar] [CrossRef]
- Ren, Z.; Zheng, Z.; Chinnusamy, V.; Zhu, J.; Cui, X.; Iida, K.; Zhu, J.K. RAS1, a Quantitative Trait Locus for Salt Tolerance and ABA Sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 5669–5674. [Google Scholar] [CrossRef]
- Jiang, Y.; Peng, X.; Zhang, Q.; Liu, Y.; Li, A.; Cheng, B.; Wu, J. Regulation of Drought and Salt Tolerance by OsSKL2 and OsASR1 in Rice. Rice 2022, 15, 46. [Google Scholar] [CrossRef]
- Borsani, O.; Zhu, J.; Verslues, P.E.; Sunkar, R.; Zhu, J.K. Endogenous SiRNAs Derived from a Pair of Natural Cis-Antisense Transcripts Regulate Salt Tolerance in Arabidopsis. Cell 2005, 123, 1279–1291. [Google Scholar] [CrossRef]
- Wan, J.; Meng, S.; Wang, Q.; Zhao, J.; Qiu, X.; Wang, L.; Li, J.; Lin, Y.; Mu, L.; Dang, K.; et al. Suppression of MicroRNA168 Enhances Salt Tolerance in Rice (Oryza sativa L.). BMC Plant Biol. 2022, 22, 563. [Google Scholar] [CrossRef]
- Mirdar Mansuri, R.; Azizi, A.H.; Sadri, A.H.; Shobbar, Z.S. Long Non-Coding RNAs as the Regulatory Hubs in Rice Response to Salt Stress. Sci. Rep. 2022, 12, 21696. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Z.; Liu, M.; Zhao, M.G.; Chen, R.; Zhang, W.H. Identification and Characterization of Long Non-Coding RNAs Involved in Osmotic and Salt Stress in Medicago Truncatula Using Genome-Wide High-Throughput Sequencing. BMC Plant Biol. 2015, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, Y.; Liu, H.; Liang, Z.; Zhang, M.; Zou, C.; Yuan, G.; Gao, S.; Pan, G.; Shen, Y.; et al. A Combination of a Genome-Wide Association Study and a Transcriptome Analysis Reveals CircRNAs as New Regulators Involved in the Response to Salt Stress in Maize. Int. J. Mol. Sci. 2022, 23, 9755. [Google Scholar] [CrossRef]
- Yin, J.; Liu, Y.; Lu, L.; Zhang, J.; Chen, S.; Wang, B. Comparison of Tolerant and Susceptible Cultivars Revealed the Roles of Circular RNAs in Rice Responding to Salt Stress. Plant Growth. Regul. 2022, 96, 243–254. [Google Scholar] [CrossRef]
- Jain, P.; Hussian, S.; Nishad, J.; Dubey, H.; Bisht, D.S.; Sharma, T.R.; Mondal, T.K. Identification and Functional Prediction of Long Non-Coding RNAs of Rice (Oryza sativa L.) at Reproductive Stage under Salinity Stress. Mol. Biol. Rep. 2021, 48, 2261–2271. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and Waterlogging Stress in Plants: A Review Highlighting Research Opportunities and Understudied Aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Aslam, A.; Mahmood, A.; Ur-Rehman, H.; Li, C.; Liang, X.; Shao, J.; Negm, S.; Moustafa, M.; Aamer, M.; Hassan, M.U. Plant Adaptation to Flooding Stress under Changing Climate Conditions: Ongoing Breakthroughs and Future Challenges. Plants 2023, 12, 3824. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, D.; Spriggs, A.; Yang, J.; Pogson, B.J.; Dennis, E.S.; Wilson, I.W. Hypoxia-Responsive MicroRNAs and Trans-Acting Small Interfering RNAs in Arabidopsis. J. Exp. Bot. 2010, 61, 165–177. [Google Scholar] [CrossRef]
- Zhai, L.; Liu, Z.; Zou, X.; Jiang, Y.; Qiu, F.; Zheng, Y.; Zhang, Z. Genome-Wide Identification and Analysis of MicroRNA Responding to Long-Term Waterlogging in Crown Roots of Maize Seedlings. Physiol. Plant. 2013, 147, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Tan, Z.; Fang, T.; Tang, K.; Liang, K.; Qiu, F. A Comprehensive Transcriptomics Analysis Reveals Long Non-Coding RNA to Be Involved in the Key Metabolic Pathway in Response to Waterlogging Stress in Maize. Genes 2020, 11, 267. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.K.; Rawal, H.C.; Jain, P.; Mishra, V.; Nishad, J.; Chowrasia, S.; Sarkar, A.K.; Sen, P.; Naik, S.K.; Mondal, T.K. Identification and Analysis of MiRNAs-LncRNAs-MRNAs Modules Involved in Stem-Elongation of Deepwater Rice (Oryza sativa L.). Physiol. Plant. 2022, 174, e13736. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Chen, L.; Zhang, Y.; Kang, X.; Zhang, Z.; Wang, Y. Identification of Novel and Conserved Populus Tomentosa MicroRNA as Components of a Response to Water Stress. Funct. Integr. Genom. 2012, 12, 327–339. [Google Scholar] [CrossRef]
- Feyissa, B.A.; Amyot, L.; Nasrollahi, V.; Papadopoulos, Y.; Kohalmi, S.E.; Hannoufa, A. Involvement of the MiR156/SPL Module in Flooding Response in Medicago sativa. Sci. Rep. 2021, 11, 3243. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Jiang, F.; Zheng, X.; Wu, Z. Identification and Analysis of Oxygen Responsive MicroRNAs in the Root of Wild Tomato (S. habrochaites). BMC Plant Biol. 2019, 19, 100. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, K.; Pan, J.; Chen, X. Small RNA Sequencing Identifies Cucumber MiRNA Roles in Waterlogging-Triggered Adventitious Root Primordia Formation. Mol. Biol. Rep. 2019, 46, 6381–6389. [Google Scholar] [CrossRef] [PubMed]
- Kęska, K.; Szcześniak, M.W.; Adamus, A.; Czernicka, M. Waterlogging-Stress-Responsive LncRNAs, Their Regulatory Relationships with MiRNAs and Target Genes in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2021, 22, 8197. [Google Scholar] [CrossRef]
- Cho, Y.H.; Hong, J.W.; Kim, E.C.; Yoo, S.D. Regulatory Functions of SnRK1 in Stress-Responsive Gene Expression and in Plant Growth and Development. Plant Physiol. 2012, 158, 1955–1964. [Google Scholar] [CrossRef]
- Stief, A.; Altmann, S.; Hoffmann, K.; Pant, B.D.; Scheible, W.R.; Bäurle, I. Arabidopsis MiR156 Regulates Tolerance to Recurring Environmental Stress through SPL Transcription Factors. Plant Cell 2014, 26, 1792–1807. [Google Scholar] [CrossRef]
- Jeyakumar, J.M.J.; Ali, A.; Wang, W.M.; Thiruvengadam, M. Characterizing the Role of the MiR156-SPL Network in Plant Development and Stress Response. Plants 2020, 9, 1206. [Google Scholar] [CrossRef]
- Wang, H.; Lu, S.; Guan, X.; Jiang, Y.; Wang, B.; Hua, J.; Zou, B. Dehydration-Responsive Element Binding Protein 1C, 1E, and 1G Promote Stress Tolerance to Chilling, Heat, Drought, and Salt in Rice. Front. Plant Sci. 2022, 13, 851731. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, J.; Liu, Z.; Li, X.; Wu, F.; He, Y. HEAT-INDUCED TAS1 TARGET1 Mediates Thermotolerance via Heat Stress Transcription Factor A1a-Directed Pathways in Arabidopsis. Plant Cell 2014, 26, 1764–1780. [Google Scholar] [CrossRef]
- Pokhrel, S.; Meyers, B.C. Heat-Responsive MicroRNAs and Phased Small Interfering RNAs in Reproductive Development of Flax. Plant Direct 2022, 6, e385. [Google Scholar] [CrossRef] [PubMed]
- Kume, K.; Tsutsumi, K.I.; Saitoh, Y. TAS1 Trans-Acting SiRNA Targets Are Differentially Regulated at Low Temperature, and TAS1 Trans-Acting SiRNA Mediates Temperature-Controlled At1g51670 Expression. Biosci. Biotechnol. Biochem. 2010, 74, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Y.; Zhang, H.; Wang, J.; Zinta, G.; Xie, S.; Zhu, W.; Nie, W.F. Genome-Wide Identification of Circular RNAs in Response to Low-Temperature Stress in Tomato Leaves. Front. Genet. 2020, 11, 591806. [Google Scholar] [CrossRef]
- Zuo, J.; Wang, Q.; Zhu, B.; Luo, Y.; Gao, L. Deciphering the Roles of CircRNAs on Chilling Injury in Tomato. Biochem. Biophys. Res. Commun. 2016, 479, 132–138. [Google Scholar] [CrossRef]
- Pan, T.; Sun, X.; Liu, Y.; Li, H.; Deng, G.; Lin, H.; Wang, S. Heat Stress Alters Genome-Wide Profiles of Circular RNAs in Arabidopsis. Plant Mol. Biol. 2018, 96, 217–229. [Google Scholar] [CrossRef]
- Mao, H.D.; Yu, L.J.; Li, Z.J.; Yan, Y.; Han, R.; Liu, H.; Ma, M. Genome-Wide Analysis of the SPL Family Transcription Factors and Their Responses to Abiotic Stresses in Maize. Plant Gene 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Arshad, M.; Hannoufa, A. Alfalfa Transcriptome Profiling Provides Insight into MiR156-Mediated Molecular Mechanisms of Heat Stress Tolerance. Genome 2022, 65, 315–330. [Google Scholar] [CrossRef]
- Zhou, M.; Tang, W. MicroRNA156 Amplifies Transcription Factor-Associated Cold Stress Tolerance in Plant Cells. Mol. Genet. Genom. 2019, 294, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, F.; Cao, H.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. TamiR159 Directed Wheat TaGAMYB Cleavage and Its Involvement in Anther Development and Heat Response. PLoS ONE 2012, 7, e48445. [Google Scholar] [CrossRef]
- Lin, J.S.; Kuo, C.C.; Yang, I.C.; Tsai, W.A.; Shen, Y.H.; Lin, C.C.; Liang, Y.C.; Li, Y.C.; Kuo, Y.W.; King, Y.C.; et al. MicroRNA160 Modulates Plant Development and Heat Shock Protein Gene Expression to Mediate Heat Tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.A.; Sung, P.H.; Kuo, Y.W.; Chen, M.C.; Jeng, S.T.; Lin, J.S. Involvement of MicroRNA164 in Responses to Heat Stress in Arabidopsis. Plant Sci. 2023, 329, 111598. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Zhang, J.; Zhu, C.; Tang, G.; Yan, J. The MiR165/166–PHABULOSA Module Promotes Thermotolerance by Transcriptionally and Posttranslationally Regulating HSFA1. Plant Cell 2023, 35, 2952–2971. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, D.; Li, H.; Chen, Q.; Zhang, Z.; Liu, M.; Liu, J.; Song, Y.; He, J.; Xu, W.; et al. Characterization and Identification of Grapevine Heat Stress-Responsive MicroRNAs Revealed the Positive Regulated Function of Vvi-MiR167 in Thermostability. Plant Sci. 2023, 329, 111623. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, C.; Cheng, X.; Zhao, X.; Li, T.; Jiang, J. MiR168 Targets Argonaute1A Mediated MiRNAs Regulation Pathways in Response to Potassium Deficiency Stress in Tomato. BMC Plant Biol. 2020, 20, 477. [Google Scholar] [CrossRef]
- Mahale, B.M.; Fakrudin, B.; Ghosh, S.; Krishnaraj, P.U. LNA Mediated in Situ Hybridization of MiR171 and MiR397a in Leaf and Ambient Root Tissues Revealed Expressional Homogeneity in Response to Shoot Heat Shock in Arabidopsis thaliana. J. Plant Biochem. Biotechnol. 2014, 23, 93–103. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, P.J.; Ahn, J.H.; Park, C.M. Arabidopsis RNA-Binding Protein FCA Regulates MicroRNA172 Processing in Thermosensory Flowering. J. Biol. Chem. 2012, 287, 16007–16016. [Google Scholar] [CrossRef]
- Fang, Y.; Zheng, Y.; Lu, W.; Li, J.; Duan, Y.; Zhang, S.; Wang, Y. Roles of MiR319-Regulated TCPs in Plant Development and Response to Abiotic Stress. Crop. J. 2021, 9, 17–28. [Google Scholar] [CrossRef]
- Thiebaut, F.; Rojas, C.A.; Almeida, K.L.; Grativol, C.; Domiciano, G.C.; Lamb, C.R.C.; De Almeida Engler, J.; Hemerly, A.S.; Ferreira, P.C.G. Regulation of MiR319 during Cold Stress in Sugarcane. Plant Cell Environ. 2012, 35, 502–512. [Google Scholar] [CrossRef]
- Wang, S.T.; Sun, X.L.; Hoshino, Y.; Yu, Y.; Jia, B.; Sun, Z.W.; Sun, M.Z.; Duan, X.B.; Zhu, Y.M. MicroRNA319 Positively Regulates Cold Tolerance by Targeting OsPCF6 and OsTCP21 in Rice (Oryza sativa L.). PLoS ONE 2014, 9, e91357. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, S.; Zhou, M.; Yuan, N.; Li, Z.; Hu, Q.; Bethea, F.G.; Liu, H.; Li, S.; Luo, H. Transgenic Creeping Bentgrass Overexpressing Osa-MiR393a Exhibits Altered Plant Development and Improved Multiple Stress Tolerance. Plant Biotechnol. J. 2019, 17, 233–251. [Google Scholar] [CrossRef]
- Ding, B.; Yue, Y.; Chen, X.; Long, X.; Zhou, Z. Identification and Expression Analysis of MiR396 and Its Target Genes in Jerusalem Artichoke under Temperature Stress. Gene 2024, 893, 147908. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kwak, K.J.; Jung, H.J.; Lee, H.J.; Kang, H. MicroRNA402 Affects Seed Germination of Arabidopsis Thaliana under Stress Conditions via Targeting DEMETER-LIKE Protein3 MRNA. Plant Cell Physiol. 2010, 51, 1079–1083. [Google Scholar] [CrossRef]
- He, J.; Jiang, Z.; Gao, L.; You, C.; Ma, X.; Wang, X.; Xu, X.; Mo, B.; Chen, X.; Liu, L. Genome-Wide Transcript and Small RNA Profiling Reveals Transcriptomic Responses to Heat Stress. Plant Physiol. 2019, 181, 609–629. [Google Scholar] [CrossRef]
- Yu, J.; Su, D.; Yang, D.; Dong, T.; Tang, Z.; Li, H.; Han, Y.; Li, Z.; Zhang, B. Chilling and Heat Stress-Induced Physiological Changes and MicroRNA-Related Mechanism in Sweetpotato (Ipomoea batatas L.). Front. Plant Sci. 2020, 11, 687. [Google Scholar] [CrossRef]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and Characterization of Wheat Long Non-Protein Coding RNAs Responsive to Powdery Mildew Infection and Heat Stress by Using Microarray Analysis and SBS Sequencing. BMC Plant Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Dai, L.; Ai, J.; Wang, Y.; Ren, F. Identification and Functional Prediction of Cold-Related Long Non-Coding RNA (LncRNA) in Grapevine. Sci. Rep. 2019, 9, 6638. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Yang, X.; Aslam, M.M.; Li, M.; Zhu, L.; Chen, S.; Zhu, G.; Li, Y.; Liu, P. Genome-Wide Identification of Long Non-Coding RNAs in Two Contrasting Rapeseed (Brassica Napus L.) Genotypes Subjected to Cold Stress. Environ. Exp. Bot. 2022, 201, 104969. [Google Scholar] [CrossRef]
- Wang, A.; Hu, J.; Gao, C.; Chen, G.; Wang, B.; Lin, C.; Song, L.; Ding, Y.; Zhou, G. Genome-Wide Analysis of Long Non-Coding RNAs Unveils the Regulatory Roles in the Heat Tolerance of Chinese Cabbage (Brassica Rapa Ssp.Chinensis). Sci. Rep. 2019, 9, 5002. [Google Scholar] [CrossRef]
- Song, Y.; Chen, P.; Liu, P.; Bu, C.; Zhang, D. High-Temperature-Responsive Poplar LncRNAs Modulate Target Gene Expression via RNA Interference and Act as RNA Scaffolds to Enhance Heat Tolerance. Int. J. Mol. Sci. 2020, 21, 808. [Google Scholar] [CrossRef]
- Di, C.; Yuan, J.; Wu, Y.; Li, J.; Lin, H.; Hu, L.; Zhang, T.; Qi, Y.; Gerstein, M.B.; Guo, Y.; et al. Characterization of Stress-Responsive LncRNAs in Arabidopsis Thaliana by Integrating Expression, Epigenetic and Structural Features. Plant J. 2014, 80, 848–861. [Google Scholar] [CrossRef]
- Wunderlich, M.; Groß-Hardt, R.; Schöffl, F. Heat Shock Factor HSFB2a Involved in Gametophyte Development of Arabidopsis Thaliana and Its Expression Is Controlled by a Heat-Inducible Long Non-Coding Antisense RNA. Plant Mol. Biol. 2014, 85, 541–550. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Li, W.; Wang, S.; Hao, L.; Xu, C.; Yu, Y.; Xiang, L.; Li, T.; Jiang, F. A Long Noncoding RNA HILinc1 Enhances Pear Thermotolerance by Stabilizing PbHILT1 Transcripts through Complementary Base Pairing. Commun. Biol. 2022, 5, 1134. [Google Scholar] [CrossRef] [PubMed]
- Jampala, P.; Garhewal, A.; Lodha, M. Functions of Long Non-Coding RNA in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1925440. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Nayyar, H.; Roychowdhury, R.; Prasad, P.V.V.; Parida, S.K.; Siddique, K.H.M. Non-Coding RNAs (NcRNAs) in Plant: Master Regulators for Adapting to Extreme Temperature Conditions. Plant Physiol. Biochem. 2023, 205, 108164. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Lian, B.; Gu, H.; Li, Y.; Qi, Y. Global Identification of Arabidopsis LncRNAs Reveals the Regulation of MAF4 by a Natural Antisense RNA. Nat. Commun. 2018, 9, 5056. [Google Scholar] [CrossRef]
- Arshad, M.; Feyissa, B.A.; Amyot, L.; Aung, B.; Hannoufa, A. MicroRNA156 Improves Drought Stress Tolerance in Alfalfa (Medicago Sativa) by Silencing SPL13. Plant Sci. 2017, 258, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An Insight into MicroRNA156 Role in Salinity Stress Responses of Alfalfa. Front. Plant Sci. 2017, 8, 356. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Terrile, M.C.; Windels, D.; Lombardo, M.C.; Bartoli, C.G.; Vazquez, F.; Estelle, M.; Casalongué, C.A. MiR393 Regulation of Auxin Signaling and Redox-Related Components during Acclimation to Salinity in Arabidopsis. PLoS ONE 2014, 9, e107678. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Suo, J.; Shi, B.; Zhou, C.; Bai, B.; Bian, H.; Zhu, M.; Han, N. The Barley MiR393 Has Multiple Roles in Regulation of Seedling Growth, Stomatal Density, and Drought Stress Tolerance. Plant Physiol. Biochem. 2019, 142, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Bo Song, J.; Gao, S.; Sun, D.; Li, H.; Xia Shu, X.; Min Yang, Z. MiR394 and LCR Are Involved in Arabidopsis Salt and Drought Stress Responses in an Abscisic Acid-Dependent Manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Song, J.B.; Gao, S.; Wang, Y.; Li, B.W.; Zhang, Y.L.; Yang, Z.M. MiR394 and Its Target Gene LCR Are Involved in Cold Stress Response in Arabidopsis. Plant Gene 2016, 5, 56–64. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, X.; Niu, F.; Sun, X.; Hu, Z.; Gao, F.; Zhang, H.; Jiang, Q. Overexpression of LncRNA77580 Regulates Drought and Salinity Stress Responses in Soybean. Plants 2023, 12, 181. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Zhao, T. High Throughput Sequencing of CircRNAs in Tomato Leaves Responding to Multiple Stresses of Drought and Heat. Hortic. Plant J. 2020, 6, 34–38. [Google Scholar] [CrossRef]
Figure 1.
Classification of non-coding RNAs (ncRNAs). Constitutive ncRNAs include transfer, ribosomal, small nuclear, small nucleolar, and small Cajal body-specific RNAs, while regulatory ncRNAs comprise small ncRNAs (small interfering, piwi, and microRNAs), circular ncRNAs, and long ncRNAs.
Figure 1.
Classification of non-coding RNAs (ncRNAs). Constitutive ncRNAs include transfer, ribosomal, small nuclear, small nucleolar, and small Cajal body-specific RNAs, while regulatory ncRNAs comprise small ncRNAs (small interfering, piwi, and microRNAs), circular ncRNAs, and long ncRNAs.
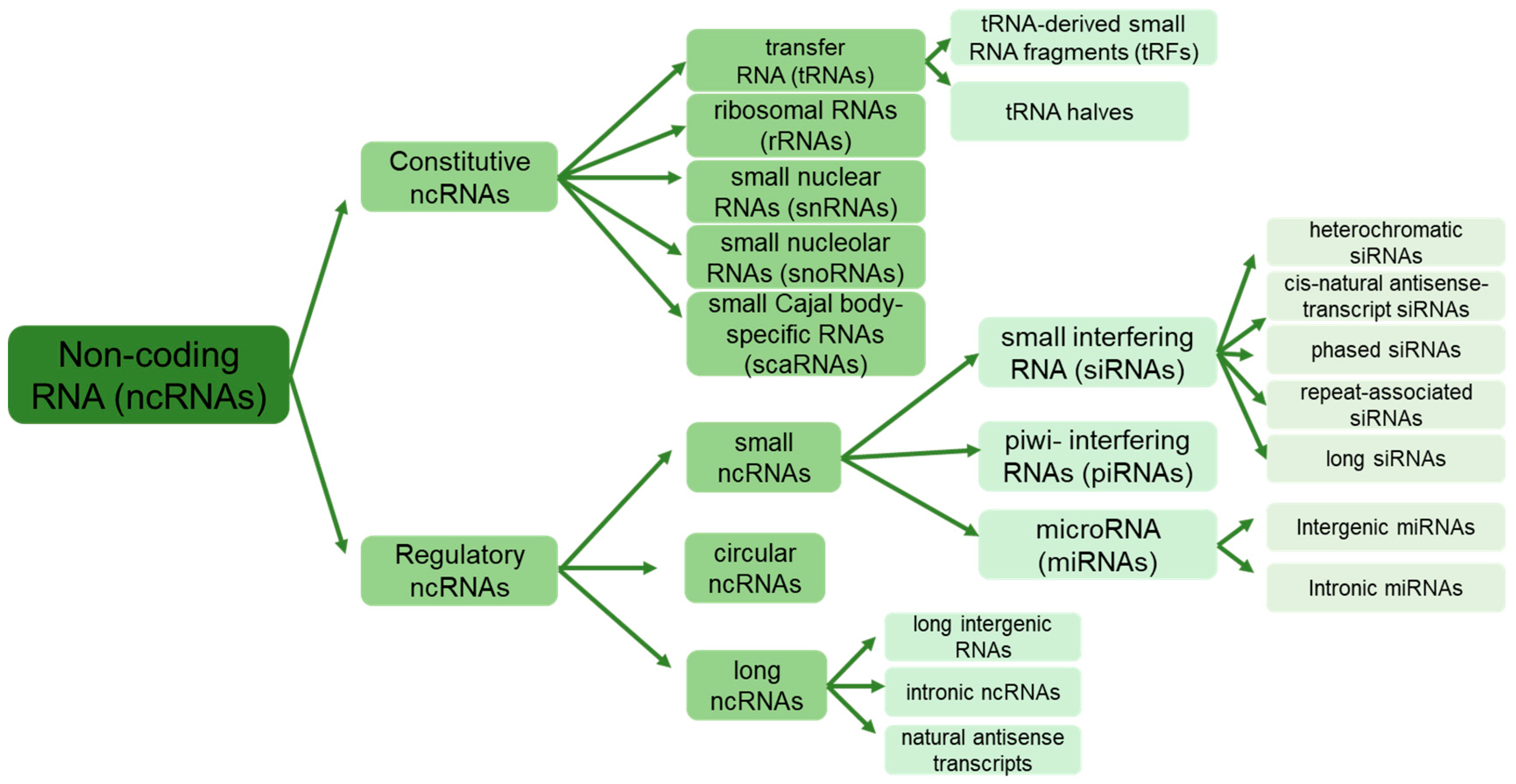
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yadav, A.; Mathan, J.; Dubey, A.K.; Singh, A. The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling. Non-Coding RNA 2024, 10, 13. https://doi.org/10.3390/ncrna10010013
AMA Style
Yadav A, Mathan J, Dubey AK, Singh A. The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling. Non-Coding RNA. 2024; 10(1):13. https://doi.org/10.3390/ncrna10010013
Chicago/Turabian StyleYadav, Amit, Jyotirmaya Mathan, Arvind Kumar Dubey, and Anuradha Singh. 2024. "The Emerging Role of Non-Coding RNAs (ncRNAs) in Plant Growth, Development, and Stress Response Signaling" Non-Coding RNA 10, no. 1: 13. https://doi.org/10.3390/ncrna10010013
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.