
Co-Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii Improves Flavour Compound Profiles of Salt-Free Fermented Wheat Gluten
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Reagents and Microorganisms
2.2. Solid-State Fermentation (SSF)
2.3. Liquid-State Fermentation (LSF)
2.4. Microbial Enumeration
2.5. Non-Volatile Compound Analysis
2.6. Volatile Compound Analysis
2.7. Statistical Analysis
3. Results
3.1. Changes in Microbial Populations
3.2. Changes in Sugar and Organic Acids
3.3. Changes in Free Amino Acids during Liquid State
3.4. Changes in Volatile Compounds during Liquid-State Fermentation
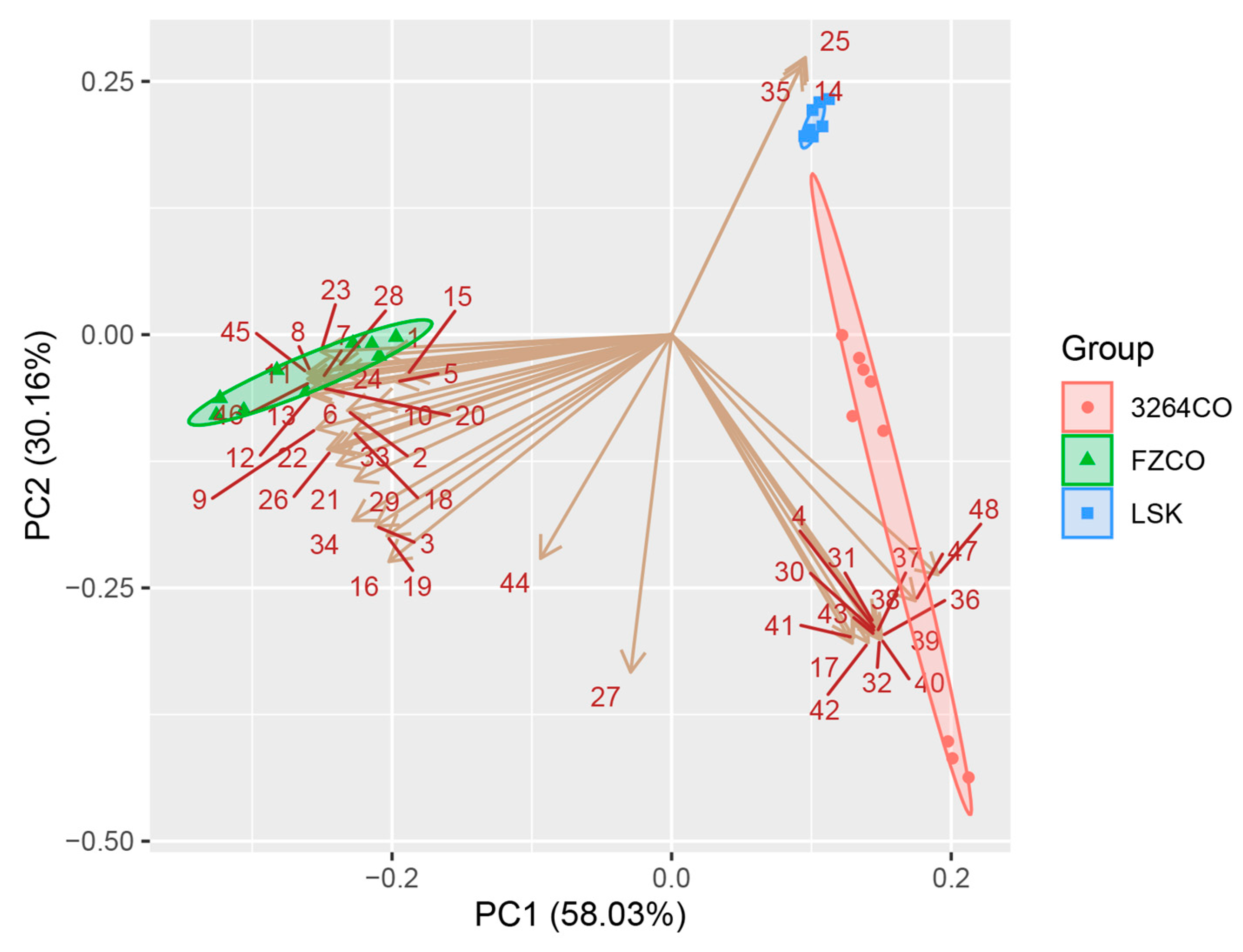
3.5. Principal Component Analysis of Volatile Compounds in Fermented Wheat Gluten Sauce
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nuraida, L. A review: Health promoting lactic acid bacteria in traditional Indonesian fermented foods. Food Sci. Hum. Wellness 2015, 4, 47–55. [Google Scholar] [CrossRef]
- De Castilhos, M.B.M.; Del Bianchi, V.L.; Gómez-Alonso, S.; García-Romero, E.; Hermosín-Gutiérrez, I. Sensory descriptive and comprehensive GC-MS as suitable tools to characterize the effects of alternative winemaking procedures on wine aroma. Part I: BRS Carmem and BRS Violeta. Food Chem. 2019, 272, 462–470. [Google Scholar] [CrossRef]
- Ferreira, I.M.; Guido, L.F. Impact of wort amino acids on beer flavour: A review. Fermentation 2018, 4, 23. [Google Scholar] [CrossRef]
- Holt, S.; Miks, M.H.; De Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, X.; Hu, F.; Fu, J.; Zhang, Z.; Liu, Z.; Wang, B.; He, R.; Ma, H.; Ho, C.T. The latest advances on soy sauce research in the past decade: Emphasis on the advances in China. Food Res. Int. 2023, 173, 113407. [Google Scholar] [CrossRef]
- Gao, X.; Liu, E.; Zhang, J.; Yang, L.; Huang, Q.; Chen, S.; Ma, H.; Ho, C.T.; Liao, L. Accelerating aroma formation of raw soy sauce using low intensity sonication. Food Chem. 2020, 329, 127118. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; Van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef]
- Vanderhaegen, B.; Neven, H.; Coghe, S.; Verstrepen, K.J.; Verachtert, H.; Derdelinckx, G. Evolution of chemical and sensory properties during aging of top-fermented beer. J. Agric. Food Chem. 2003, 51, 6782–6790. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Sakumoto, N.; Kaneko, Y.; Harashima, S. Mga2p is a putative sensor for low temperature and oxygen to induce OLE1 transcription in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2002, 291, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, D.; Yoshimoto, H.; Sone, H.; Harashima, S.; Tamai, Y. Transcriptional co-regulation of Saccharomyces cerevisiae alcohol acetyltransferase gene, ATF1 and Δ-9 fatty acid desaturase gene, OLE1 by unsaturated fatty acids. Yeast 1998, 14, 711–721. [Google Scholar] [CrossRef]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2008, 15, 1–8. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Technol. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, L.; Xing, G.; Xu, X.; Tu, C.; Dong, M. Effect of co-fermentation with lactic acid bacteria and K. marxianus on physicochemical and sensory properties of goat milk. Foods 2020, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, J.; Lassabliere, B.; Yu, B.; Liu, S.Q. Coffee flavour modification through controlled fermentation of green coffee beans by Saccharomyces cerevisiae and Pichia kluyveri: Part II. Mixed cultures with or without lactic acid bacteria. Food Res. Int. 2020, 136, 109452. [Google Scholar] [CrossRef] [PubMed]
- Devanthi, P.V.P.; Linforth, R.; Onyeaka, H.; Gkatzionis, K. Effects of co-inoculation and sequential inoculation of Tetragenococcus halophilus and Zygosaccharomyces rouxii on soy sauce fermentation. Food Chem. 2018, 240, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Singracha, P.; Niamsiri, N.; Visessanguan, W.; Lertsiri, S.; Assavanig, A. Application of lactic acid bacteria and yeasts as starter cultures for reduced-salt soy sauce (moromi) fermentation. LWT-Food Sci. Technol. 2017, 78, 181–188. [Google Scholar] [CrossRef]
- Cui, R.Y.; Zheng, J.; Wu, C.D.; Zhou, R.Q. Effect of different halophilic microbial fermentation patterns on the volatile compound profiles and sensory properties of soy sauce moromi. Eur. Food Res. Technol. 2014, 239, 321–331. [Google Scholar] [CrossRef]
- Wah, T.T.; Walaisri, S.; Assavanig, A.; Niamsiri, N.; Lertsiri, S. Co-culturing of Pichia guilliermondii enhanced volatile flavor compound formation by Zygosaccharomyces rouxii in the model system of Thai soy sauce fermentation. Int. J. Food Microbiol. 2013, 160, 282–289. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, F.; Ananta, E.; Muller, J.A.; Liang, Y.; Lee, Y.K.; Liu, S.-Q. Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii improves flavor compound profiles of salt-free fermented wheat-gluten: Effects from single strain inoculation. Curr. Res. Nutr. Food Sci. 2023, 6, 100492. [Google Scholar] [CrossRef]
- Baensch, J.; Gaier, W.; Lim, B.G.; Lai, H.L.; Khoo, H.G.N. Hydrolyzate Seasoning Preparation in the Presence of Lactic Acid Bacteria. United States Patent US 5,965,178 A, 12 October 1999. Available online: https://patft.uspto.gov/netacgi/nph-Parser?Sect1=PTO1&Sect2=HITOFF&d=PALL&p=1&u=%2Fnetahtml%2FPTO%2Fsrchnum.htm&r=1&f=G&l=50&s1=5965178.PN.&OS=PN/5965178&RS=PN/5965178 (accessed on 25 July 2022).
- Lim, B.G.; Ng, T.L.; Heyland, S.; Dac, T.H. Production of a Hydrolysate. United States Patent US 6,383,532 B1, 7 May 2002. Available online: https://patents.justia.com/patent/6383532 (accessed on 19 January 2024).
- Lu, Y.; Chan, L.J.; Li, X.; Liu, S.Q. Effects of different inoculation strategies of Saccharomyces cerevisiae and Williopsis saturnus on chemical components of mango wine. LWT-Food Sci. Technol. 2018, 87, 85–92. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually stimulating interactions between lactic acid bacteria and Saccharomyces cerevisiae in sourdough fermentation. LWT-Food Sci. Technol. 2018, 90, 201–206. [Google Scholar] [CrossRef]
- Sieuwerts, S.; De Bok, F.A.M.; Hugenholtz, J.; Van Hylckama Vlieg, J.E.T. Unraveling microbial interactions in food fermentations: From classical to genomics approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.E.; Lee, S.M.; Choi, Y.H.; Hurh, B.S.; Kim, Y.S. Comparative volatile profiles in soy sauce according to inoculated microorganisms. Biosci. Biotechnol. Biochem. 2013, 77, 2192–2200. [Google Scholar] [CrossRef] [PubMed]
- Noda, F.; Hayashi, K.; Mizunuma, T. Antagonism between osmophilic lactic acid bacteria and yeasts in brine fermentation of soy sauce. Appl. Environ. Microbiol. 1980, 40, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Giannattasio, S.; Guaragnella, N.; Corte-Real, M.; Passarella, S.; Marra, E. Acid stress adaptation protects Saccharomyces cerevisiae from acetic acid-induced programmed cell death. Gene 2005, 354, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Ludovico, P.; Sousa, M.J.; Silva, M.T.; Leão, C.; Côrte-Real, M. Saccharomyces cerevisiae commits to a programmed cell death process in response to acetic acid. Microbiology 2001, 147, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Ludovico, P.; Sansonetty, F.; Silva, M.T.; Côrte-Real, M. Acetic acid induces a programmed cell death process in the food spoilage yeast Zygosaccharomyces bailii. FEMS Yeast Res. 2003, 3, 91–96. [Google Scholar] [CrossRef]
- Derrick, S.; Large, P.J. Activities of the enzymes of the Ehrlich pathway and formation of branched-chain alcohols in Saccharomyces cerevisiae and Candida utilis grown in continuous culture on valine or ammonium as sole nitrogen source. J. Gen. Microbiol. 1993, 139, 2783–2792. [Google Scholar] [CrossRef]
- Dickinson, J.R.; Norte, V. A study of branched-chain amino acid aminotransferase and isolation of mutations affecting the catabolism of branched-chain amino acids in Saccharomyces cerevisiae. FEBS Lett. 1993, 326, 29–32. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Bánsági, T.; Taylor, A.F. Ester hydrolysis: Conditions for acid autocatalysis and a kinetic switch. Tetrahedron 2017, 73, 5018–5022. [Google Scholar] [CrossRef]
- Amárita, F.; Fernández-Esplá, D.; Requena, T.; Pelaez, C. Conversion of methionine to methional by Lactococcus lactis. FEMS Microbiol. Lett. 2001, 204, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Perpète, P.; Duthoit, O.; De Maeyer, S.; Imray, L.; Lawton, A.I.; Stavropoulos, K.E.; Gitonga, V.W.; Hewlins, M.J.E.; Richard Dickinson, J. Methionine catabolism in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Yivon, M.; Rijnen, L. Cheese flavour formation by amino acid catabolism. Int. Dairy J. 2001, 11, 195–201. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Schwab, C. Exploitation of the metabolic potential of lactic acid bacteria for improved quality of gluten-free bread. In The Science of Gluten-Free Foods and Beverages; Arendt, E.K., Bello, F.B., Eds.; AACC International, Inc.: Cork, Ireland, 2009; pp. 99–111. [Google Scholar] [CrossRef]
- Damiani, P.; Gobbetti, M.; Cossignani, L.; Corsetti, A.; Simonetti, M.S.; Rossi, J. The sourdough microflora. Characterization of hetero- and homofermentative lactic acid bacteria, yeasts and their interactions on the basis of the volatile compounds produced. LWT-Food Sci. Technol. 1996, 29, 63–70. [Google Scholar] [CrossRef]

 changes in cell count of co-culture of L. sakei LTH673 and P. kluyveri FrootZen;
changes in cell count of co-culture of L. sakei LTH673 and P. kluyveri FrootZen;  changes in cell count of co-culture of L. sakei LTH673 and S. boulardii NCYC3264.
changes in cell count of co-culture of L. sakei LTH673 and P. kluyveri FrootZen; changes in cell count of co-culture of L. sakei LTH673 and S. boulardii NCYC3264.
changes in cell count of co-culture of L. sakei LTH673 and S. boulardii NCYC3264.
changes in cell count of co-culture of L. sakei LTH673 and P. kluyveri FrootZen; changes in cell count of co-culture of L. sakei LTH673 and S. boulardii NCYC3264.

 glucose,
glucose,  lactic acid,
lactic acid,  acetic acid,
acetic acid,  quinic acid,
quinic acid,  oxalic acid and
oxalic acid and  formic acid concentration in samples of FZCO (co-culture of L. sakei LTH673 and P. kluveri FrootZen) (a), 3264CO (co-culture of L. sakei LTH673 and S. boulardii NCYC3264) (b) during liquid-state fermentation of wheat gluten slurries.
glucose, lactic acid, acetic acid, quinic acid, oxalic acid and formic acid concentration in samples of FZCO (co-culture of L. sakei LTH673 and P. kluveri FrootZen) (a), 3264CO (co-culture of L. sakei LTH673 and S. boulardii NCYC3264) (b) during liquid-state fermentation of wheat gluten slurries.
formic acid concentration in samples of FZCO (co-culture of L. sakei LTH673 and P. kluveri FrootZen) (a), 3264CO (co-culture of L. sakei LTH673 and S. boulardii NCYC3264) (b) during liquid-state fermentation of wheat gluten slurries.
glucose, lactic acid, acetic acid, quinic acid, oxalic acid and formic acid concentration in samples of FZCO (co-culture of L. sakei LTH673 and P. kluveri FrootZen) (a), 3264CO (co-culture of L. sakei LTH673 and S. boulardii NCYC3264) (b) during liquid-state fermentation of wheat gluten slurries.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Free Amino Acid | Day 0 | Day 1 | Day 2 | Day 3 | Day 4 |
|---|---|---|---|---|---|
| Concentration * (mM) (mean ± S.D) in FZCO | |||||
| Alanine | 7.13 ± 1.42 | 34.55 ± 9.78 | 31.57 ± 2.71 | 31.65 ± 0.75 | 33.41 ± 0.86 |
| Arginine | 2.25 ± 0.31 | 0.10 ± 0.03 | 0.12 ± 0.01 | 0.13 ± 0.02 | 0.12 ± 0.00 |
| Aspartic acid | 4.91 ± 0.42 | 7.18 ± 1.56 | 7.44 ± 1.54 | 7.95 ± 0.87 | 9.41 ± 0.54 |
| Cysteine | 0.99 ± 0.11 | 0.22 ± 0.13 | 0.38 ± 0.02 | 0.44 ± 0.12 | 0.59 ± 0.13 |
| Glutamic acid | 37.91 ± 6.00 | 119.13 ± 2.24 | 152.12 ± 6.17 | 165.07 ± 13.08 | 178.75 ± 10.98 |
| Glutamine | 3.20 ± 0.57 | 15.51 ± 3.50 | 8.18 ± 1.10 | 5.25 ± 1.33 | 3.61 ± 2.42 |
| Glycine | 6.36 ± 0.60 | 28.94 ± 8.12 | 26.38 ± 1.40 | 26.44 ± 1.04 | 27.88 ± 0.80 |
| Histidine | 1.81 ± 0.32 | 8.34 ± 1.77 | 7.20 ± 0.40 | 6.85 ± 0.15 | 7.14 ± 0.55 |
| Isoleucine | 7.49 ± 1.12 | 28.23 ± 7.65 | 25.94 ± 2.87 | 25.88 ± 1.59 | 28.36 ± 1.50 |
| Lysine | 3.94 ± 0.56 | 7.08 ± 1.79 | 6.40 ± 0.48 | 6.38 ± 0.20 | 6.99 ± 0.33 |
| Leucine | 11.56 ± 3.70 | 41.79 ± 10.09 | 37.91 ± 4.67 | 37.78 ± 2.45 | 40.78 ± 2.34 |
| Methionine | 2.14 ± 0.39 | 6.56 ± 1.52 | 6.08 ± 0.36 | 5.86 ± 0.20 | 6.36 ± 0.34 |
| Ornithine | 0.26 ± 0.07 | 15.45 ± 3.87 | 11.77 ± 1.51 | 10.88 ± 0.30 | 11.41 ± 1.37 |
| Phenylalanine | 6.52 ± 2.47 | 27.84 ± 7.11 | 23.71 ± 1.53 | 22.78 ± 1.58 | 24.43 ± 0.79 |
| Proline | 21.38 ± 5.56 | 60.66 ± 5.84 | 57.46 ± 4.65 | 53.64 ± 6.95 | 56.03 ± 5.05 |
| Serine | 8.80 ± 3.14 | 6.32 ± 0.83 | 5.04 ± 0.87 | 5.25 ± 2.23 | 7.12 ± 2.63 |
| Threonine | 4.31 ± 0.41 | 18.14 ± 3.44 | 21.05 ± 8.46 | 22.05 ± 3.95 | 22.99 ± 0.64 |
| Tryptophan | 0.85 ± 0.16 | 1.96 ± 1.43 | 3.10 ± 0.37 | 3.14 ± 0.28 | 3.23 ± 0.10 |
| Tyrosine | 5.59 ± 1.42 | 16.54 ± 4.34 | 12.30 ± 1.28 | 10.98 ± 0.62 | 11.12 ± 0.48 |
| Valine | 13.76 ± 3.72 | 28.29 ± 6.79 | 26.26 ± 2.05 | 26.24 ± 1.39 | 28.48 ± 1.01 |
| Concentration (mM) (mean ± S.D) in FZCO | |||||
| Alanine | 4.89 ± 0.66 | 25.00 ± 1.80 | 28.57 ± 1.52 | 33.69 ± 3.01 | 36.53 ± 6.52 |
| Arginine | 2.20 ± 0.54 | 1.83 ± 0.38 | 2.15 ± 0.45 | 1.64 ± 0.08 | 1.62 ± 0.87 |
| Aspartic acid | 4.06 ± 0.30 | 3.48 ± 0.15 | 4.87 ± 0.90 | 5.51 ± 1.26 | 6.47 ± 1.50 |
| Cysteine | 0.47 ± 0.10 | 0.09 ± 0.01 | 0.20 ± 0.03 | 0.42 ± 0.04 | 0.55 ± 0.11 |
| Glutamic acid | 26.50 ± 1.61 | 117.64 ± 6.85 | 124.16 ± 10.08 | 143.32 ± 9.82 | 149.87 ± 9.27 |
| Glutamine | 2.42 ± 0.39 | 13.29 ± 0.31 | 12.13 ± 0.14 | 11.63 ± 0.62 | 11.89 ± 2.07 |
| Glycine | 5.39 ± 1.01 | 17.99 ± 0.41 | 20.62 ± 0.76 | 24.81 ± 2.27 | 29.19 ± 5.77 |
| Histidine | 1.07 ± 0.09 | 5.85 ± 0.17 | 6.31 ± 0.46 | 7.20 ± 0.59 | 7.62 ± 1.23 |
| Isoleucine | 4.23 ± 0.61 | 14.04 ± 0.95 | 16.87 ± 1.21 | 19.91 ± 1.95 | 21.12 ± 3.72 |
| Lysine | 3.37 ± 0.16 | 4.75 ± 0.08 | 5.40 ± 0.31 | 6.38 ± 0.68 | 7.32 ± 1.29 |
| Leucine | 5.76 ± 0.82 | 19.88 ± 1.43 | 23.57 ± 1.64 | 27.78 ± 2.45 | 30.17 ± 6.05 |
| Methionine | 1.36 ± 0.37 | 4.28 ± 0.30 | 4.98 ± 0.30 | 5.59 ± 0.49 | 5.98 ± 0.98 |
| Ornithine | 0.35 ± 0.44 | 8.18 ± 1.22 | 9.92 ± 0.68 | 11.05 ± 0.98 | 11.29 ± 1.72 |
| Phenylalanine | 3.39 ± 0.27 | 18.73 ± 0.59 | 20.23 ± 0.64 | 22.38 ± 1.63 | 23.57 ± 3.17 |
| Proline | 7.83 ± 1.39 | 63.85 ± 5.33 | 59.96 ± 4.44 | 62.57 ± 7.67 | 66.24 ± 4.46 |
| Serine | 4.08 ± 0.30 | N.D | N.D | 1.24 ± 1.06 | 2.47 ± 1.19 |
| Threonine | 4.24 ± 0.49 | 14.40 ± 0.21 | 14.39 ± 0.14 | 14.66 ± 0.21 | 14.61 ± 0.26 |
| Tryptophan | 0.48 ± 0.04 | 2.70 ± 0.53 | 3.32 ± 0.20 | 3.42 ± 0.34 | 3.59 ± 0.47 |
| Tyrosine | 3.40 ± 0.16 | 12.27 ± 0.63 | 10.97 ± 0.89 | 11.17 ± 0.68 | 12.20 ± 1.59 |
| Valine | 7.83 ± 0.97 | 16.60 ± 1.17 | 18.95 ± 1.30 | 22.35 ± 2.20 | 23.42 ± 3.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Zhang, F.; Ananta, E.; Muller, J.A.; Liang, Y.; Lee, Y.K.; Liu, S.-Q. Co-Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii Improves Flavour Compound Profiles of Salt-Free Fermented Wheat Gluten. Fermentation 2024, 10, 75. https://doi.org/10.3390/fermentation10020075
Chen S, Zhang F, Ananta E, Muller JA, Liang Y, Lee YK, Liu S-Q. Co-Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii Improves Flavour Compound Profiles of Salt-Free Fermented Wheat Gluten. Fermentation. 2024; 10(2):75. https://doi.org/10.3390/fermentation10020075
Chicago/Turabian StyleChen, Shuoyu, Fanxin Zhang, Edwin Ananta, Jeroen André Muller, Youyun Liang, Yuan Kun Lee, and Shao-Quan Liu. 2024. "Co-Inoculation of Latilactobacillus sakei with Pichia kluyveri or Saccharomyces boulardii Improves Flavour Compound Profiles of Salt-Free Fermented Wheat Gluten" Fermentation 10, no. 2: 75. https://doi.org/10.3390/fermentation10020075