
Metabolic Oscillation Phenomena in Clostridia Species—A Review

Department of Chemical and Biochemical Engineering, Western University, London, ON TEB 459, Canada
*
Author to whom correspondence should be addressed.
Fermentation 2024, 10(3), 156; https://doi.org/10.3390/fermentation10030156
Submission received: 1 February 2024
/
Revised: 5 March 2024
/
Accepted: 5 March 2024
/
Published: 9 March 2024
(This article belongs to the Special Issue Feature Review Papers in Microbial Metabolism, Physiology & Genetics 2023)
Abstract
:Clostridia are interesting candidates for biotechnological applications due to their diverse and unique metabolic abilities. Particularly in continuous fermentation processes, productivity-decreasing metabolic oscillations have been reported in many species. The resulting process instability and reduced productivity can be a serious hurdle for the development of industrially feasible processes. This review highlights the current state of knowledge about oscillatory metabolic phenomena in Clostridia, including the mechanisms, assumed and proven, behind those oscillations and methods to mitigate the phenomena if applicable. The nature of observed metabolic oscillations in Clostridia is diverse, including a wide range of periods of oscillation and different parameters in which the oscillation is observed. Some phenomena remain to be investigated further, while others are already well understood. However, knowledge of mechanisms is a very valuable asset in overcoming the metabolic oscillation to create a stable process.
1. Introduction
Oscillation phenomena are common in biological systems [1], with the most prominent example being the circadian rhythm of physical, mental and behavioral changes over a 24 h period that is present in many organisms established through molecular oscillators [2,3]. The circadian rhythm appears to be an evolutionary response to light and dark cycles, with easily observable effects present in macro-organisms. To be able to observe metabolic oscillation in the commonly measured parameters of a microbial culture, the oscillation must be synchronized. All (or close to all) cells in a given cultivation system need to be in the same state at the same time, which is unlikely to be the case just through internal processes alone. If there is not a completely external trigger, a mechanism has to be behind it that involves some form of interaction between the cells or between cells and the environment. Nevertheless, observations of oscillation in continuous cultivations of microbes are not uncommon [4]. Metabolic oscillations in continuous culture are reported for many bacteria, including Escherichia coli [5,6,7], Zymomonas mobilis [8] and Klebsiella pneumoniae [9,10] as well as the yeast Saccharomyces cerevisiae [11,12,13,14,15,16].
Any given system requires a negative feedback loop and a response delay in order to establish damped oscillatory behaviour. Undamped oscillation can occur for feedback systems with three or more steps (with a similar timescale and strong feedback required), while biological oscillators often occur if positive and negative feedback loops are coupled, resulting in Hopf bifurcation. For a detailed review of molecular system dynamics and bifurcation, refer to the specific literature (e.g., [17,18,19]). Continuous cultivation of yeasts or bacteria in bioreactors is typically described via unstructured models based on the respective concentrations of biomass (X [g/L]), substrate(s) (Si [g/L]) and product(s) (Pi [g/L]), where biomass or cells are seen as unstructured units. Clostridia are known for solvent production and their dynamic behavior in continuous culture focusing on a single product can be described as follows:
where D is the dilution rate [h−1] and Yn/m are the respective yield coefficients [g/g]. The specific growth rate µ [h−1] is a function of S and P (with negative feedback as often inhibitory at elevated concentrations), providing the required feedback loops. Depending on the dependency of S and P, bifurcation can occur and be described mathematically (e.g., [20]); however, understanding the metabolic reasons behind the oscillation requires an understanding of the underlying metabolic network [21].
Clostridia are a group of anaerobic gram-positive bacteria that have gained an interest due to their diverse metabolic abilities [22,23], for example, the fixation of inorganic carbon in the form of CO or CO2 [24], making them interesting candidates for biotechnological applications. Historically Clostridia, more specifically Clostridium acetobutylicum, were used during a period of multiple decades in the industrial-scale production of acetone and butanol (ABE fermentation), until petrochemical production processes took over [25]. Currently, several species of Clostridia are being investigated, especially for the production of solvents and biofuels like 1,3-propanediol, butanol, acetone and ethanol from alternative sustainable resources [22,23], as alternative production processes become increasingly relevant with the depletion of fossil resources and environmental concerns on the rise. Clostridia allow for the utilization of a diverse range of renewable feedstocks, including waste streams such as (pre-treated) lignocellulosic biomass and by-products such as the glycerol generated during biodiesel production, allowing for feedstock cost reduction and increased sustainability [26]. Another interesting sustainable feedstock that can be generated through gasification of renewable resources or biomass-containing waste is syngas, a mixture of mainly CO, H2 and CO2, which can be utilized by some species with carbon-fixing abilities [26,27].
Bioprocesses in the pharmaceutical industry are predominately conducted in batch mode [28]. However, processes targeting fuels or commodity chemicals typically operate under a different economic backdrop, where the main contributors to the product’s cost are the costs of raw materials and of bioreactor operation [29,30]. The requirements of product purity are subrationally lower, and the total product volume is high. Such low-value and high-volume processes benefit from continuous processing under anaerobic conditions, allowing constant steady-state operation of large-scale industrial bioreactors [31]. For the use of continuous fermentation in industrial applications, a true steady state, where the production rate is constant, is desired. However, for several Clostridia species metabolic oscillation phenomena in continuous culture have been observed. The reports differ in the affected fermentation parameters, but oscillation can commonly be seen, for example, in substrate and product concentrations or biomass. Oscillation is often detected when online data are collected at high measurement frequencies, such as in redox potential or off-gas measurements. Periods of oscillation from less than 1 h to multiple days have been reported (Figure 1). Such oscillation forms a challenge in the establishment of a true steady state, hindering more efficient production, and needs to be understood to be mitigated. Figure 1 compares the different timescales of the oscillatory behaviours of two different Clostridia species, both reported in 2020 [32,33]. The oscillation in gas consumption by Clostridium autoethanogenum follows a 60–100 h period [33], while the gas production of Clostridium pasteurianum [32] oscillates every 2 h. In both cases, the metabolites and cell concentrations are also affected by the oscillation. In the case of C. pasteurianum [32], this could only be shown through carefully timed sampling at the peaks and valleys of the online signals obtained from off-gas and redox probes (glycerol and butanol data shown in Figure 1). The oscillation would have likely evaded detection based on off-line analysis only, and butanol and glycerol data would have simply appeared as “noisy”, had the sampling time not been synchronized with the oscillation rate. The slow oscillation of C. autoethanogenum [33] is easy to detect in the biomass signal, while it is entirely possible that other fast-oscillating systems have remained unnoticed, potentially wasting metabolic potential. In the following sections, reports of metabolic oscillation in different Clostridia species are summarized and compared.
2. Metabolic Oscillation in Clostridia
Various forms of metabolic oscillations have been reported for a wide range of clostridia species as summarized in Table 1. A detailed discussion for selected species follows in the subsections below.
2.1. Metabolic Oscillation in Clostridium pasteurianum
C. pasteurianum was the first organism for which fixation of free nitrogen was shown [34]. Biotechnologically, it gained interest for its unique ability to transform glycerol into butanol [35,36], particularly as it was shown to tolerate the waste-stream glycerol (crude glycerol) from biodiesel plants [37].
Signs of the oscillating metabolism of C. pasteurianum were published by Johnson and Rehmann in 2016 in a publication about batch fermentations of C. pasteurianum on glycerol as a substrate under different circumstances [38]. For the lowest tested pH of 4.7, there was a synchronized oscillation visible in the CO2 production as well as the redox potential after 20 h of fermentation time. These observations were only mentioned briefly and not further explored. However, it was assumed that it is a “true metabolic response” (not an artifact of the equipment, etc.).
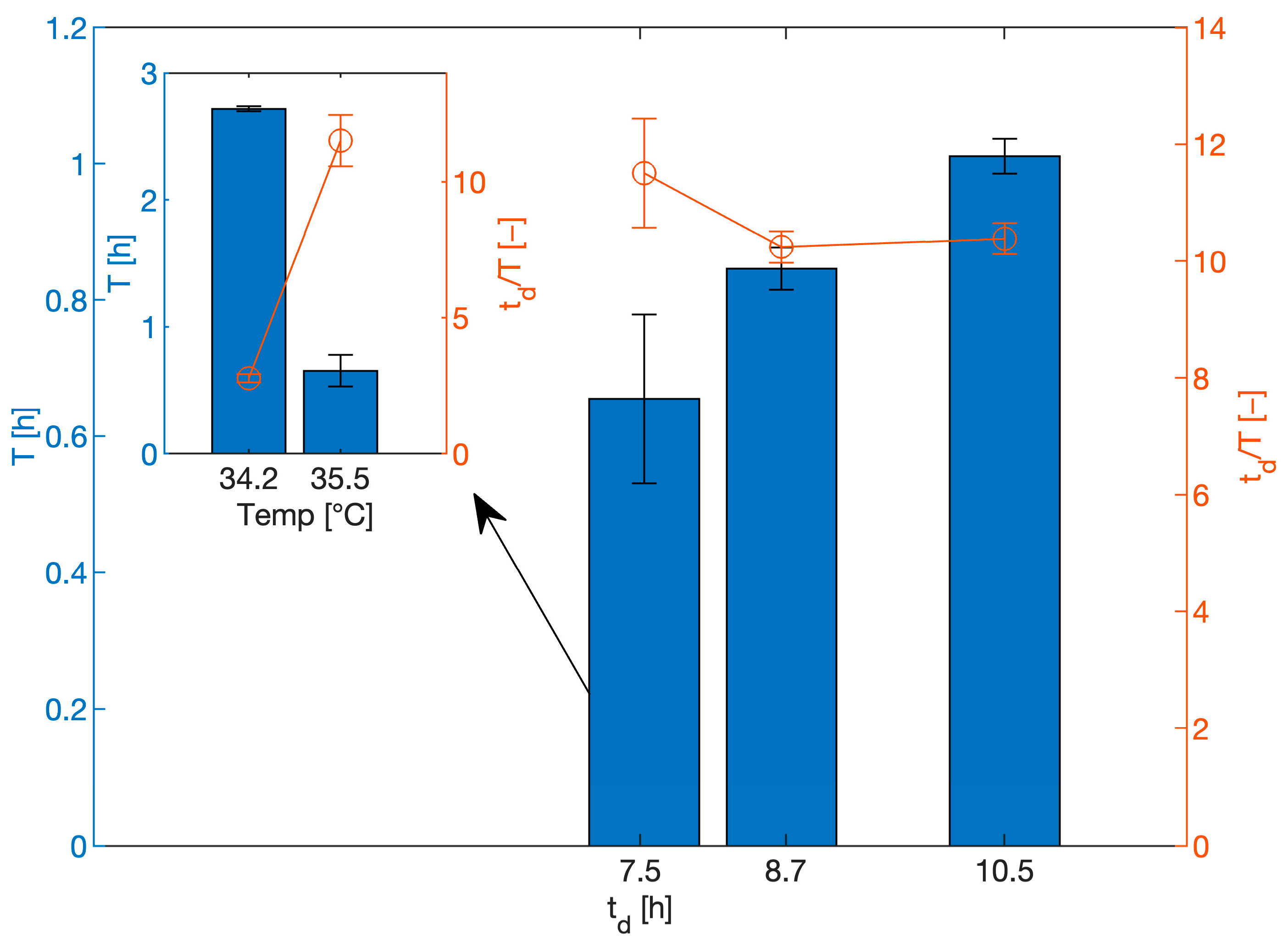
In 2020, Johnson and Rehmann dedicated a publication to the oscillating metabolic behaviour of C. pasteurianum in continuous fermentation with glycerol as the substrate [32]. Different fermentation conditions were examined to investigate their influence on metabolic oscillation. It was shown that the dilution rate D, and hence, the specific growth rate µ, directly correlated with the oscillation frequency ν under otherwise constant conditions. The number of oscillations per doubling time of the cells appeared constant under otherwise identical operating conditions (Figure 2). Under the given conditions, approximately 10 oscillations occurred per cell division, highlighting the high frequency of the oscillations, which were likely caused by a highly synchronized switch in the metabolism. However, the oscillation frequency at a given specific growth rate is affected by pH, the substrate concentration in the feed and the reactor temperature [32]. The oscillation frequency decreases with temperature (Figure 2), and a small change from 35.5 °C to 34.3 °C resulted in approximately a 4-fold decrease in the frequency. The oscillations could be fully eliminated at 31 °C and reinstated upon temperature increase [32]. The substrate concentration in the feed was also shown to influence the frequency of the oscillations, while at lower concentrations, no oscillation was observed. The frequency of oscillation also increases with increasing pH. In general, the period of the oscillation is within the order of a few hours. The mechanism behind the oscillation is not understood yet. However, it is postulated that it is based in the regulation of enzymes involved in the oxidative glycerol metabolism by intermediates or reductive equivalents. Temperature and pH appear to affect specific reactions differently; hence, some conditions result in an imbalanced reaction network, leading to the observed oscillation. The negative feedback loop leading to the oscillation was not caused by inhibition through the final product (butanol), as the authors showed by artificially increasing butanol levels in the reactor. The oscillation might be similar to oscillations observed in the glycolysis pathway in yeast or bacteria mediated by redox coenzyme couples (ATP/ADP and NADH/NAD) and via both intra- and intercellular acetaldehyde [39,40].
Gallazzi et al. observed oscillating behaviour in continuous fermentation of C. pasteurianum in the consumption of glycerol and the production of butanol and butyric acid, as well as cell viability and density [41]. In contrast to the work by Johnson and Rehmann, in Gallazzi et al.’s work, the period of oscillation was in the magnitude of multiple days. Gallazzi et al. speculated that the oscillation was due to butanol toxicity. In their observed oscillation, Johnson and Rehmann showed that butanol did not affect it, and hence, butanol toxicity did not play a role in the mechanism of fast oscillation [32]. The residual substrate concentration leaving the reactor varied between 2.5 g/L and 4 g/L during oscillation, hence implying that high substrate conversion is achievable in the absence of oscillation.
2.2. Metabolic Oscillation in Clostridium acetobutylicum
Clostridium acetobutylicum is the most prominent biotechnologically used member of the genus Clostridium, as it was utilized in ABE (acetone–butanol–ethanol) fermentation that was developed in the early 20th century and used for industrial production for decades [42].
Clarke et al. described oscillating behaviour in continuous fermentation of C. acetobutylicum, with a period length of about 30 h [43]. C. acetobutylicum is known to display biphasic behaviour in batch fermentation, where a phase of acid production is followed by a phase of solvent production [25,44]. In Clarke et al.’s experiment, the oscillation was visible in the growth rate synchronized with the alternating production of acids and solvents, where acids are produced when the growth rate is high and solvents are produced when it is low [43].
Mulchandani and Volesky described oscillation with a period length of about 24 h with several fermentation parameters in continuous ABE fermentation using C. acetobutylicum [45]. They used a spin-filter perfusion bioreactor to retain cells and only remove cell-free spent medium. For feed glucose concentrations over 49 g/L, no steady state could be achieved, and undamped oscillation was observed that ceased when, additionally to cell-free medium, a small amount of cell-containing medium was removed as well. The authors postulate that the oscillation happens due to product inhibition by butanol, as production and cell growth stops at a butanol concentration of about 10 g/L and restarts at about 6 g/L. At the higher glucose concentrations, a higher cell density is achieved, leading to higher butanol production. The butanol production rate is higher than the rate with which is it removed, so that it accumulates until it reaches toxic levels. The cells will stop growing and producing until enough butanol is washed out. Subsequently, they will start growing and producing when enough butanol is removed. Continuous removal of cells from the process leads to a lower overall cell concentration and therefore a lower rate of butanol production. Toxic levels will not be reached, and hence, there is no oscillation.
Metabolic oscillation in continuous fermentation of C. acetobutylicum was observed by Barbeau et al. as well [46]. For low dilution rates, they observed oscillation with a period length of several days in gas production and cell growth as well as solvent and acid production. The proposed reason for the metabolic oscillation is product inhibition through toxic effects of accumulated butanol, similar to the reasoning of Mulchandani and Volesky [45]. After ceasing butanol production at high butanol concentrations, butanol gets washed out, resulting in the lifting of the inhibition. The results of Mulchandani and Volesk are challenging to interpret, as the apparent oscillation only occurred during reactor operation with full cell retention. Under such conditions, a true steady state is not achievable. However, the dynamic butanol concentration varied between ~10 g/L and ~4.5 g/L with a ~22 h period length.
Guerrero et al. examined a degenerated strain of C. acetobutylicum that lacked the pSOL1 plasmid and hence was unable to sporulate or produce solvents for its potential in hydrogen production [47]. In continuous fermentation with a high dilution rate, for the first approximately 400 h, no steady state could be reached, and oscillation with a period of approximately 50 h was most prevalently visible in glucose consumption and cell growth but was also present in acid and hydrogen production. No possible explanation for the oscillation was given. An inherent instability of continuous fermentation with C. acetobutylicum, especially at high dilution rates, is mentioned in the literature [48]; however, it is thought to be caused by the different vegetative states the cells go through in the process of sporulation, which is mitigated in the examined strain.
Based on the available data, it appears that oscillation during the cultivation of C. acetobutylicum is either caused by product inhibition or by changes in the relative distribution of cells involved in either of the two metabolic stages of C. acetobutylicum (acidogenesis and solventogenesis).
2.3. Metabolic Oscillation in Clostridium saccharoperbutylacetonicum
Clostridium saccharoperbutylacetonicum has been shown to be able to convert butyrate into butanol when co-fed with glucose [49,50].
Richter et al. damped metabolic oscillation in the solventogenic activity during the conversion of n-butyrate to n-butanol by C. saccharoperbutylacetonicum in a continuous culture by introducing a second stage of continuous culture and establishing a two-stage system [51]. The first stage, which is only 1/10 of the volume of the second stage, is dedicated to biomass production, whereas the actual conversion of n-Butyrate to n-Butanol takes place in the second stage. The system also involved in situ product removal via gas stripping. Without the first stage dedicated to cell production (“continuous inoculum”), “dramatic” oscillations in n-butanol production were visible within a period of 4–6 days.
2.4. Metabolic Oscillation in Clostridium thermobutyricum
Clostridium thermobutyricum produces butyric acid under moderately thermophilic conditions [52].
Canganella and Wiegel performed continuous cultivation of Clostridium thermobutyricum in a rotary fermenter system to investigate growth behaviour and butyric acid production [53]. They observed a metabolic shift between the production of butyrate and acetate in an oscillatory fashion. The organism produces H2 when metabolizing glucose to generate energy. At lower H2 concentrations in the environment, the production of acetate, which releases more H2, is thermodynamically favorable over the formation of butyrate. Hence, as a mechanism for oscillation, they suggest that at a lower H2 concentration, the organism shifts to producing acetate, and then when a certain concentration is reached, shifts back to producing butyrate, which releases less H2. The concentration will decline until it shifts back to acetate production. Hence, the governing mechanism is similar to product inhibition, as is possibly the case caused by butanol for C. acetobutylicum.
2.5. Metabolic Oscillation in Clostridium beijerinckii
Sandoval-Espinola et al. showed mixotrophic carbon-capture by Clostridium beijerinckii [54]. A partial re-assimilation of CO2 and H2 was detected as synchronous oscillation of CO2 and H2 in fed-batch fermentation off-gasses for a brief period in the late log phase and with the onset of solventogenesis. They theorized that the oscillation was caused by feedback-loop controls as a response to a metabolic pathway change, which was supported by a coinciding decrease in specific growth rate as CO2 and H2 started to reaccumulate, which is indicative of diauxic growth. A possible connection to the redox state of the cell was also mentioned, as C1 assimilation can serve to balance redox potential, aiding in substrate uptake.
In contrast, a different kind of oscillation that is rooted in the nature of ABE fermentation was also observed in C. beijerinckii. During ABE fermentation, bacterial cells go through different cellular stages in which they grow and produce acid, do not grow and produce solvents, sporulate or die, which causes a general instability in ABE fermentation [55]. In continuous culture, an alternating pattern of domination between growing acetogenic cells vs. non-growing solventogenic cells can occur, leading to production of acids and solvents in an oscillatory fashion. This was observed in C. beijerinckii NCIMB-8052 by Paredes et al. [56] and was visible in cell and glucose concentrations as well as acid production, with a period of oscillation of approximately 50 h, and in C. beijerinckii BA101 by Ezeji et al. [55], being visible in acid and solvent production, with a period of oscillation between 20 and 100 h. These observations of alternating solvent and acid production are similar to observations for C. acetobutylicum (see “Metabolic oscillation in Clostridium acetobutylicum” Section 2.2).
Additionally, oscillation might occur during concurrent substrate saccharification and fermentation, as shown when C. beijerinckii 6A1 is cultivated on complex carbohydrates with alcohol stillage as a substrate [57], also falling loosely into the category of a slow switch from one metabolic stage (saccharification) to another (fermentation) that is governed by the respective concentration profile in the reactor.
2.6. Metabolic Oscillation in Clostridium autoethanogenum
C. autoethanogenum is capable of capturing inorganic carbon in the form of CO or CO2, which makes it an interesting candidate for capturing carbon from waste gases using gas fermentation [24].
Mahamkali et al. observed metabolic oscillation in continuous culture of C. autoethanogenum in gas uptake rate, extracellular by-products (acetate, ethanol, 2,3-butanediol, pyruvate) and biomass levels [33] during gas fermentation using syngas (CO, H2, CO2). The oscillation was characterized by changes in substrate consumption, with oscillations in the CO and H2 uptake rates. Initially, the organism grows on CO and then switches to co-utilizing CO and H2 (see Figure 1). It lasts until the thermodynamic driving force for the production of ethanol through the oxidoreductase (AOR) is exhausted due to ethanol accumulation, and the culture crashes (the growth rate falls under the dilution rate). H2 uptake as well as growth decline, and an increase in CO concentration causes a shift to growth on CO, which cannot support the culture by itself, and the cells keep declining until they hit the point for which the provided CO is supportive. The oscillation was not linked to translational control, but a coordination with the cell-internal redox potential was shown. The observed period of oscillation was about 6 days.
2.7. Metabolic Oscillation in Clostridium butyricum
Clostridium butyricum has been recognized for its ability to produce butyric acid but is also capable of converting glycerol into 1,3-propanediol [58].
Zhou et al. observed metabolic oscillation in a microbial consortium containing 94.64% Clostridiaceae during continuous fermentation for the production of 1,3-propanediol from crude glycerol under glycerol-limited conditions [59]. The period of oscillation was approximately 49 h. The make-up of the consortium fluctuated between 99.23% and 68.74% Clostridiaceae during oscillation. The consortium dominating C. butyricum was shown to oscillate by itself. Hence, the oscillations observed with the consortium are most likely caused by this organism and not a consequence of interaction between different species.
Subsequently, Zhou et al. examined metabolic oscillation in Clostridium butyricum during continuous fermentation for 1,3-propanediol production from glycerol [60]. The oscillation occurred under glycerol-limited conditions at low dilution rates and had a period of about 51 h. It was observable in biomass production, the concentration of glycerol and products like 1,3-propanediol, butyrate, acetate, lactate, formate, H2 and CO2, as well as periodic changes in cell morphology. Analysis of extracellular redox potential and the intracellular NAD+/NADH ratio showed a link between the oscillation and the redox state of the cells. Oscillation only happened under substrate-limited conditions and ceased for increased dilution rates (higher dilution rates require higher specific growth rates and hence higher residual concentrations of the limiting substrate). However, if the substrate availability is too low, no sustained oscillation can be achieved either, likely due to an insufficiently strong positive feedback loop. Toxic effects from known fermentation end products as a cause for oscillation were able to be ruled out, as with a sufficient glycerol supply, similar concentrations of end products but no oscillation were observed. However, Zhou et al. suggested that an accumulation of the toxic intermediate acetaldehyde could be one possible explanation. Additionally, an abnormal production pattern of lactate, formate and H2 could point towards a disorder of the pyruvate metabolism as a cause for the oscillation. The maximal production of lactate, formate and H2 was lagging behind those of other products like 1,3-propanediol, and two peaks occurred during one periodic cycle, pointing towards a blockage of the main pyruvate degradation route and a use of alternative pathways for its breakdown.

{kind=link}
{kind=link}
Table 1.
Overview of reported cases of observed oscillation in cultures of Clostridia.
| Organism | Process/Application | Oscillating Parameters | Oscillation Conditions | Period of Oscillation | Mechanism of Oscillation | Strategies to Mitigate | Source |
|---|---|---|---|---|---|---|---|
| C. pasteurianum | Butanol production | CO2 in off-gas; redox potential | Batch fermentations on glycerol, pH of 4.7, after 20 h of fermentation time | A few hours | “true metabolic response” | [38] | |
| Butanol production | CO2 in off-gas; redox potential | Continuous fermentation with glycerol as substrate | A few hours | Regulation of enzymes involved in the oxidative glycerol metabolism by intermediates or reductive equivalents | Oscillation ceases for lower fermentation temperatures | [32] | |
| Butanol production | Concentrations of glycerol, butanol, butyric acid; cell viability and density | Continuous fermentation | Multiple days | Product inhibition (butanol) | n/a | [41] | |
| C. acetobutylicum | ABE fermentation | Growth rate; concentrations of acids and solvents | Continuous fermentation | ~30 h | Biphasic behaviour; acidogenic vs. solventogenic phase | n/a | [43] |
| ABE fermentation | Biomass; concentrations of glucose, butanol, butyric acid, acetone, acetic acid, ethanol | Continuous fermentation; spin-filter perfusion bioreactor to retain cells and only remove cell-free spent medium; feed glucose concentrations over 49 g/L | ~24 h | Product inhibition (butanol) | Removal of cell-containing medium instead of only cell-free medium | [45] | |
| ABE fermentation | Gas production; cell growth; concentrations of acids and solvents | Continuous fermentation, low dilution rates | Several days | Product inhibition (butanol) | n/a | [46] | |
| H2 production | Cell growth; concentrations of glucose and acids; H2 outflow | Degenerated strain (no sporulation, no solventogenesis), continuous fermentation with high dilution rate | ~50 h | n/a | Eventually self-stabilized after 400 h of fermentation | [47] | |
| C. saccharoperbutylacetonicum | Conversion of n-Butyrate to n-Butanol | Concentration of butanol | Continuous culture, in situ product removal via gas stripping | 4–6 days | n/a | Establishing a two stage-system; first stage: “continuous inoculum”; second stage: actual conversion | [51] |
| C. thermobutyricum | Butyric acid production | Concentrations of butyrate and acetate | Continuous cultivation in rotary fermenter system, glucose as substrate | Several days | Product inhibition (H2) | n/a | [53] |
| C. beijerinckii | Butanol from gas fermentation | CO2 and H2 in fermentation off-gasses | Fed-batch fermentation | A few hours | Partial re-assimilation of CO2 and H2, mixotrophic carbon capture | n/a | [54] |
| ABE fermentation | Cell and glucose concentrations as well as acid production | Continuous culture | ~50 h | Biphasic behaviour; acidogenic vs. solventogenic phase | High dilution rates favour acid-producing cells, eventually self-stabilized after 60 h | [56] | |
| ABE fermentation | Acid and solvent production | Continuous culture | Between 20 and 100 h | Biphasic behaviour; acidogenic vs. solventogenic phase | n/a | [55] | |
| H2 production | Concentrations of glucose and ethanol | Batch culture of C. beijerinckii 6A1 with alcohol stillage supplemented with glucose as a substrate | ~70 h | Switch between saccharification and fermentation | n/a | [57] | |
| C. autoethanogenum | Gas fermentation using syngas | Gas uptake rate; concentrations of extracellular by-products (acetate, ethanol, 2,3-butanediol, pyruvate); biomass levels | Continuous culture | 6 days | Alternating growth on CO and CO and H2, coordinated with cell-internal redox potential | n/a | [33] |
| C. butyricum | 1,3-propandiol production from glycerol | Cell morphology; biomass; concentrations of glycerol, 1,3-propandiol, butyrate, acetate, formate, lactate; redox-potential; CO2 and H2 in off-gas | Continuous fermentation, glycerol-limited conditions at low dilution rates | 51 h (~2 days) | Connected to redox state of the cells | n/a | [60] |
| Clostridia-dominated Consortium | 1,3-Propandiol production form crude glycerol | Make-up of the consortium | Continuous fermentation, glycerol-limited conditions | 49 h (~2 days) | C. butyricum causes oscillation, not interaction within consortium | n/a | [59] |
3. Discussion
The described oscillation phenomena are diverse and not all caused by the same mechanisms. Unifying characters throughout are shifts in product profiles and growth rates.
A few of the oscillations in C. acetobutylicum and C. pasteurianum are united by the fact that they are likely caused by product inhibition through the toxic effect of butanol [41,43,46]. In a continuous fermentation, when the organism produces butanol at a higher rate than it is removed by the outflow, it will accumulate and lead to a toxic effect on the cells. The culture declines and stops producing butanol until the outflow has decreased the butanol concentration below toxic levels. Product inhibition effectively limits the maximum effective substrate concentration that can be fed to a continuous reactor, assuming a constant product yield coefficient. Typically, product inhibition is modelled by adding a product-dependent additional term to the Monod equation describing the specific growth rate as a function of the substrate concentration. Such a model would simply predict a steady state that results in a growth rate equal to the reactor’s dilution rate at a given substrate and product concentration. The observed oscillation, hence, could be a result of latency in the system, resulting in the cells’ control mechanism not being able to reach a steady state and hence oscillating around it. The time scale and proposed mechanism behind this type of oscillation do not require complete synchronization of the cell population in a given bioreactor.
Zhou et al. suggest a similar mechanism through accumulation of acetaldehyde for the oscillations observed in C. butyricum [60]; however, in this case, the acetaldehyde is not an end product of the fermentation but an intermediate of the metabolism. Nevertheless, its conversion could be the rate-limiting step of the organism’s metabolism. Other than in the above-discussed scenario of end-product inhibition, the inhibition caused by a non-secreted intermediate would not be removed through washout but would require the inhibition of an enzyme upstream in the pathway while still allowing for the further conversion of the inhibiting intermediate. The observed timescale of the oscillation in the order of days implies a lack of cell synchronization and a dynamic distribution of cells that are either in the inhibited or non-inhibited metabolic state.
Also caused by the accumulation of a product, a high concentration of the by-product H2 causes a shift in the product profile of C. thermobutyricum [53], which makes it produce less H2 until the H2 level is decreased again. However, this effect is not caused by toxic effects of H2 but by thermodynamic reasons that make the production of H2 when there is already a high presence of H2 unfavorable. The metabolic diversity of Clostridia might therefore be used to make adjustments based on environmental conditions. The time needed to activate alternate pathways might vary from strain to strain and pathway to pathway, possibly explaining the observed oscillations as well as their vastly different timescales.
Supporting this hypothesis are the observations for C. acetobutylicum [43] and C. beijerinckii [55,56], where oscillation is reportedly linked to the biphasic nature that is also observable in batch fermentation with these organisms [25,44]. A phase of acid production increases the amount of acid, and a high amount of acid causes a shift to solvent production, which has similarity to product inhibition in that the accumulation of the product will eventually cause a shift in the production profile.
Utilization of different substrates can also lead to the observation of oscillation. In the case of gas fermentation with C. autoethanogenum, a switch between utilization of CO only at high CO concentrations and utilization of H2 and CO when CO is low can be observed [33], which in turn leads to an increase in CO. For C. beijerinckii [54], a shift from organotrophic to mixotrophic metabolism causes a short period of oscillation that is visible in the CO2 and H2 concentrations in the off-gas as those gases get partially re-uptaken. The gradual breakdown of complex substrates might also lead to observable oscillation in fermentation parameters, as observed in C. beijerinckii [57], clearly placing the oscillation in a different category from what was discussed above.
The period of oscillation varies between a few hours and several days. This might have to do with the different mechanisms behind the oscillation that make it follow different timelines. For the outstanding shortest reported periods of oscillation which were reported in C. pasteurianum [32,38], the mechanism for oscillation is not yet fully understood, but a likely link to the cell-internal redox state is assumed. For other organisms, a link to the cell’s redox potential is reported [33,60] or assumed [54] as well. An oscillation frequency of ν = 1.1.53 h−1 has been reported for C. pasteurianum while growing at a specific growth rate of µ = 0.092 h−1, translating into over 10 oscillations per doubling time [32]. A high degree of synchronization is required to explain such rapid changes being observable at the reactor level.
For the oscillations observed in C. saccharoperbutylacetonicum, no mechanism for oscillation has been proposed [51]; however in batch culture, it is most comparable to C. beijerinckii. The coverage of strategies to mitigate oscillation varies between different reports. For C. saccharoperbutylacetonicum, dampening the oscillation by introducing a two-stage system including a continuous inoculum likely physically separates the acidogenic and the solventogenic stage, removing the need to oscillate between the two of them [51]. For the continuous cultivation of C. acetobutylicum in a spin-filter perfusion bioreactor, which can retain the cells, the oscillation ceases when, additionally to cell-free medium, a small amount of cell-containing medium is removed as well [45]. The removal of cells helps overcome product inhibition by butanol, as a lower amount of cells leads to a lower butanol production rate and toxic levels will not be hit.
Other reports mention that oscillation only happens under certain conditions or that certain conditions for substrate concentration, dilution rate in continuous culture, temperature or pH can reduce or prevent oscillation [32,38,45,59,60]. However, adjusting those fermentation parameters to meet these conditions could be unfavorable when optimizing other aspects of the process.
4. Conclusions
The diverse reported metabolic oscillations are a challenge that needs to be overcome in the commercialization of fermentation processes with Clostridia. As the reasons for oscillation are diverse, deductions from reported oscillations need to be made carefully, and an easy transfer of knowledge about the oscillation process between species or even within the same species is not always possible. The reported oscillations fall into three main categories:
- (A)
- Strains that follow a biphasic growth cycle in batch cultivation (e.g., acid formation followed by solvent formation) can switch between the two growth stages in continuous cultivation, resulting in observed oscillation. Such processes, similar to the production of secondary metabolites, are not well suited for single-reactor continuous processing and benefit from reactor cascades.
- (B)
- Processes using multiple or complex substrates and rates depending on the respective concentrations of these substrates cannot achieve a steady state in response to dynamic change in the substrate concentrations caused by cell growth. Finely tuned process control might be used to provide substrates at suitable ratios to avoid oscillations between them.
- (C)
- In terms of inhibition caused by either a product or an intermediate, the oscillations attributed to this mechanism can be challenging to mitigate and are the least understood, as inhibition is typically expected to simply result in a different steady state. Different degrees of cell synchronization appear during these oscillations, resulting in vastly different timescales. Such systems offer excellent opportunities to investigate the dynamics of complex metabolic networks.
5. Future Directions
With the different states of knowledge on what causes the metabolic oscillations for some of the reported oscillations, more extensive research on the mechanisms of oscillation is necessary, as they are not well understood yet. However, even in cases where a theory of a mechanism is established, further research is necessary to understand the molecular interactions causing it and prove the theoretical suggestions. Single-cell metabolite measurements have been established for E. coli [40], and similar techniques could be used for clostridia. Autofluorescence of NADH was used to monitor glycolytic oscillations at the single-cell level in HeLa cervical cancer cells [61], a technique that might be suitable for investigating glycolytic oscillation in C. pasteurianum. The data obtained through such systematic studies could be used to establish dynamic models of metabolic networks, which could then be tested and verified by designing dedicated experiments to study the system’s response (e.g., pulsing or step changes). A better understanding of what causes the oscillations will help to mitigate them and create stable and efficient processes. A molecular understanding of oscillation mechanisms might enable the genetic engineering of Clostridia strains that do not display metabolic oscillation.
Author Contributions
Conceptualization, L.R. and A.T.; investigation, A.T.; writing—original draft preparation, A.T.; writing—review and editing, L.R.; visualization, L.R.; supervision, L.R.; project administration, L.R.; funding acquisition, L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded the Natural Science and Engineering Research Council of Canada (NSERC), [grant number: RGPIN-2020-06060], Genome Canada [grant number: GAAP RD 16-OGI-190] and the Canada Foundation for Innovation (CFI) [grant number: 27300].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Cao, Y.; Lopatkin, A.; You, L. Elements of Biological Oscillations in Time and Space. Nat. Struct. Mol. Biol. 2016, 23, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular Mechanisms and Physiological Importance of Circadian Rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular Architecture of the Mammalian Circadian Clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.E.F.; Topiwala, H.H. Transient and Oscillatory States of Continuous Culture. In Advances in Biochemical Engineering, Volume 3; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 1974; Volume 3, pp. 167–219. ISBN 978-3-540-06546-3. [Google Scholar]
- Weber, J.; Kayser, A.; Rinas, U. Metabolic Flux Analysis of Escherichia coli in Glucose-Limited Continuous Culture. II. Dynamic Response to Famine and Feast, Activation of the Methylglyoxal Pathway and Oscillatory Behaviour. Microbiology 2005, 151, 707–716. [Google Scholar] [CrossRef]
- Ofiţeru, I.D.; Ferdeş, M.; Knapp, C.W.; Graham, D.W.; Lavric, V. Conditional Confined Oscillatory Dynamics of Escherichia coli Strain K12-MG1655 in Chemostat Systems. Appl. Microbiol. Biotechnol. 2012, 94, 185–192. [Google Scholar] [CrossRef]
- Robert, M.; Murray, D.; Honma, M.; Nakahigashi, K.; Soga, T.; Tomita, M. Extracellular Metabolite Dynamics and Temporal Organization of Metabolic Function in E. coli. In Proceedings of the 2012 ICME International Conference on Complex Medical Engineering (CME), Kobe, Japan, 1–4 July 2012; pp. 197–202. [Google Scholar]
- Jöbses, I.M.L.; Egberts, G.T.C.; Luyben, K.C.A.M.; Roels, J.A. Fermentation Kinetics of Zymomonas mobilis at High Ethanol Concentrations: Oscillations in Continuous Cultures. Biotechnol. Bioeng. 1986, 28, 868–877. [Google Scholar] [CrossRef]
- Menzel, K.; Zeng, A.-P.; Biebl, H.; Deckwer, W.-D. Kinetic, Dynamic, and Pathway Studies of Glycerol Metabolism by Klebsiella pneumoniae in Anaerobic Continuous Culture: I. The Phenomena and Characterization of Oscillation and Hysteresis. Biotechnol. Bioeng. 1996, 52, 549–560. [Google Scholar] [CrossRef]
- Sun, L.-H.; Song, Z.-Y.; Sun, Y.-Q.; Xiu, Z.-L. Dynamic Behavior of Glycerol–Glucose Co-Fermentation for 1,3-Propanediol Production by Klebsiella pneumoniae DSM 2026 under Micro-Aerobic Conditions. World J. Microbiol. Biotechnol. 2010, 26, 1401–1407. [Google Scholar] [CrossRef]
- Richard, P. The Rhythm of Yeast. FEMS Microbiol. Rev. 2003, 27, 547–557. [Google Scholar] [CrossRef]
- Tu, B.P.; McKnight, S.L. The Yeast Metabolic Cycle: Insights into the Life of a Eukaryotic Cell. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 339–343. [Google Scholar] [CrossRef]
- Meyenburg, H.K.V. Energetics of the Budding Cycle of Saccharomyces cerevisiae during Glucose Limited Aerobic Growth. Arch. Mikrobiol. 1969, 66, 289–303. [Google Scholar] [CrossRef]
- Murray, D.B.; Beckmann, M.; Kitano, H. Regulation of Yeast Oscillatory Dynamics. Proc. Natl. Acad. Sci. USA 2007, 104, 2241–2246. [Google Scholar] [CrossRef]
- Satroutdinov, A.D.; Kuriyama, H.; Kobayashi, H. Oscillatory Metabolism of Saccharomyces cerevisiae in Continuous Culture. FEMS Microbiol. Lett. 1992, 98, 261–267. [Google Scholar] [CrossRef]
- Xu, Z.; Tsurugi, K. A Potential Mechanism of Energy-metabolism Oscillation in an Aerobic Chemostat Culture of the Yeast Saccharomyces cerevisiae. FEBS J. 2006, 273, 1696–1709. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Barik, D. Pulsatile Signaling of Bistable Switches Reveal the Distinct Nature of Pulse Processing by Mutual Activation and Mutual Inhibition Loop. J. Theor. Biol. 2022, 540, 111075. [Google Scholar] [CrossRef]
- Tyson, J.J.; Novak, B. A Dynamical Paradigm for Molecular Cell Biology. Trends Cell Biol. 2020, 30, 504–515. [Google Scholar] [CrossRef]
- Tong, C.S.; Xu, X.J.; Wu, M. Periodicity, Mixed-Mode Oscillations, and Multiple Timescales in a Phosphoinositide-Rho GTPase Network. Cell Rep. 2023, 42, 112857. [Google Scholar] [CrossRef]
- Jarzębski, A.B. Modelling of Oscillatory Behaviour in Continuous Ethanol Fermentation. Biotechnol. Lett. 1992, 14, 137–142. [Google Scholar] [CrossRef]
- Kardynska, M.; Kogut, D.; Pacholczyk, M.; Smieja, J. Mathematical Modeling of Regulatory Networks of Intracellular Processes—Aims and Selected Methods. Comput. Struct. Biotechnol. J. 2023, 21, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Yang, S.-T. Editorial: Development and Application of Clostridia as Microbial Cell-Factories for Biofuels and Biochemicals Production. Front. Bioeng. Biotechnol. 2022, 9, 831135. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Bao, T.; Yang, S.-T. Engineering Clostridium for Improved Solvent Production: Recent Progress and Perspective. Appl. Microbiol. Biotechnol. 2019, 103, 5549–5566. [Google Scholar] [CrossRef] [PubMed]
- Heffernan, J.K.; Valgepea, K.; De Souza Pinto Lemgruber, R.; Casini, I.; Plan, M.; Tappel, R.; Simpson, S.D.; Köpke, M.; Nielsen, L.K.; Marcellin, E. Enhancing CO2-Valorization Using Clostridium Autoethanogenum for Sustainable Fuel and Chemicals Production. Front. Bioeng. Biotechnol. 2020, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Woods, D.R. Acetone-Butanol Fermentation Revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef]
- Liberato, V.; Benevenuti, C.; Coelho, F.; Botelho, A.; Amaral, P.; Pereira, N.; Ferreira, T. Clostridium sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context. Catalysts 2019, 9, 962. [Google Scholar] [CrossRef]
- Oliveira, L.; Rückel, A.; Nordgauer, L.; Schlumprecht, P.; Hutter, E.; Weuster-Botz, D. Comparison of Syngas-Fermenting Clostridia in Stirred-Tank Bioreactors and the Effects of Varying Syngas Impurities. Microorganisms 2022, 10, 681. [Google Scholar] [CrossRef] [PubMed]
- Kasemiire, A.; Avohou, H.T.; De Bleye, C.; Sacre, P.-Y.; Dumont, E.; Hubert, P.; Ziemons, E. Design of Experiments and Design Space Approaches in the Pharmaceutical Bioprocess Optimization. Eur. J. Pharm. Biopharm. 2021, 166, 144–154. [Google Scholar] [CrossRef]
- Llano, T.; Arce, C.; Gallart, L.E.; Perales, A.; Coz, A. Techno-Economic Analysis of Macroalgae Biorefineries: A Comparison between Ethanol and Butanol Facilities. Fermentation 2023, 9, 340. [Google Scholar] [CrossRef]
- Meramo-Hurtado, S.I.; González-Delgado, Á.; Rehmann, L.; Quinones-Bolanos, E.; Mehvar, M. Comparative Analysis of Biorefinery Designs Based on Acetone-Butanol-Ethanol Fermentation under Exergetic, Techno-Economic, and Sensitivity Analyses towards a Sustainability Perspective. J. Clean. Prod. 2021, 298, 126761. [Google Scholar] [CrossRef]
- Xie, D. Continuous Biomanufacturing with Microbes—Upstream Progresses and Challenges. Curr. Opin. Biotechnol. 2022, 78, 102793. [Google Scholar] [CrossRef]
- Johnson, E.E.; Rehmann, L. Self-Synchronized Oscillatory Metabolism of Clostridium pasteurianum in Continuous Culture. Processes 2020, 8, 137. [Google Scholar] [CrossRef]
- Mahamkali, V.; Valgepea, K.; De Souza Pinto Lemgruber, R.; Plan, M.; Tappel, R.; Köpke, M.; Simpson, S.D.; Nielsen, L.K.; Marcellin, E. Redox Controls Metabolic Robustness in the Gas-Fermenting Acetogen Clostridium autoethanogenum. Proc. Natl. Acad. Sci. USA 2020, 117, 13168–13175. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, M.; Gutnick, D. Sergei Winogradsky: A Founder of Modern Microbiology and the First Microbial Ecologist. FEMS Microbiol. Rev. 2012, 36, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Heyndrickx, M.; Vos, P.D.; Vancanneyt, M.; Ley, J.D. The Fermentation of Glycerol by Clostridium butyricum LMG 1212t2 and 1213tl and C. pasteurianum LMG 3285. Appl. Microbiol. Biotechnol. 1991, 34, 637–642. [Google Scholar] [CrossRef]
- Biebl, H. Fermentation of Glycerol by Clostridium pasteurianum—Batch and Continuous Culture Studies. J. Ind. Microbiol. Biotechnol. 2001, 27, 18–26. [Google Scholar] [CrossRef]
- Jensen, T.Ø.; Kvist, T.; Mikkelsen, M.J.; Christensen, P.V.; Westermann, P. Fermentation of Crude Glycerol from Biodiesel Production by Clostridium pasteurianum. J. Ind. Microbiol. Biotechnol. 2012, 39, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.E.; Rehmann, L. The Role of 1,3-Propanediol Production in Fermentation of Glycerol by Clostridium pasteurianum. Bioresour. Technol. 2016, 209, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Passarge, J.; Somsen, O.J.G.; Snoep, J.L.; Heinrich, R.; Westerhoff, H.V. Transduction of Intracellular and Intercellular Dynamics in Yeast Glycolytic Oscillations. Biophys. J. 2000, 78, 1145–1153. [Google Scholar] [CrossRef]
- Bi, S.; Kargeti, M.; Colin, R.; Farke, N.; Link, H.; Sourjik, V. Dynamic Fluctuations in a Bacterial Metabolic Network. Nat. Commun. 2023, 14, 2173. [Google Scholar] [CrossRef]
- Gallazzi, A.; Branska, B.; Marinelli, F.; Patakova, P. Continuous Production of N-Butanol by Clostridium pasteurianum DSM 525 Using Suspended and Surface-Immobilized Cells. J. Biotechnol. 2015, 216, 29–35. [Google Scholar] [CrossRef]
- Dürre, P. Fermentative Butanol Production: Bulk Chemical and Biofuel. Ann. N. Y. Acad. Sci. 2008, 1125, 353–362. [Google Scholar] [CrossRef]
- Clarke, K.G.; Hansford, G.S.; Jones, D.T. Nature and Significance of Oscillatory Behavior during Solvent Production by Clostridium acetobutylicum in Continuous Culture. Biotechnol. Bioeng. 1988, 32, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, L.; Ke, C.; Pang, Z.; Liu, L. Pathway Dissection, Regulation, Engineering and Application: Lessons Learned from Biobutanol Production by Solventogenic Clostridia. Biotechnol. Biofuels 2020, 13, 39. [Google Scholar] [CrossRef] [PubMed]
- Mulchandani, A.; Volesky, B. Production of Acetone-Butanol-Ethanol by Clostridium acetobutylicum Using a Spin Filter Perfusion Bioreactor. J. Biotechnol. 1994, 34, 51–60. [Google Scholar] [CrossRef]
- Barbeau, J.Y.; Marchal, R.; Vandecasteele, J.P. Conditions Promoting Stability of Solventogenesis or Culture Degeneration in Continuous Fermentations of Clostridium acetobutylicum. Appl. Microbiol. Biotechnol. 1988, 29, 447–455. [Google Scholar] [CrossRef]
- Guerrero, K.; Gallardo, R.; Paredes, I.; Quintero, J.; Mau, S.; Conejeros, R.; Gentina, J.C.; Aroca, G. Continuous Biohydrogen Production by a Degenerated Strain of Clostridium acetobutylicum ATCC 824. Int. J. Hydrogen Energy 2021, 46, 5100–5111. [Google Scholar] [CrossRef]
- Janssen, H.; Wang, Y.; Blaschek, H.P. CLOSTRIDIUM|Clostridium acetobutylicum. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 449–457. ISBN 978-0-12-384733-1. [Google Scholar]
- Tashiro, Y.; Takeda, K.; Kobayashi, G.; Sonomoto, K.; Ishizaki, A.; Yoshino, S. High Butanol Production by Clostridium saccharoperbutylacetonicum N1-4 in Fed-Batch Culture with pH-Stat Continuous Butyric Acid and Glucose Feeding Method. J. Biosci. Bioeng. 2004, 98, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Al-Shorgani, N.K.N.; Ali, E.; Kalil, M.S.; Yusoff, W.M.W. Bioconversion of Butyric Acid to Butanol by Clostridium saccharoperbutylacetonicum N1-4 (ATCC 13564) in a Limited Nutrient Medium. BioEnergy Res. 2012, 5, 287–293. [Google Scholar] [CrossRef]
- Richter, H.; Qureshi, N.; Heger, S.; Dien, B.; Cotta, M.A.; Angenent, L.T. Prolonged Conversion of n-butyrate to n-butanol with Clostridium Saccharoperbutylacetonicum in a Two-stage Continuous Culture with In-situ Product Removal. Biotechnol. Bioeng. 2012, 109, 913–921. [Google Scholar] [CrossRef]
- Wiegel, J.; Kuk, S.-U.; Kohring, G.W. Clostridium thermobutyricum sp. nov., a Moderate Thermophile Isolated from a Cellulolytic Culture, That Produces Butyrate as the Major Product. Int. J. Syst. Bacteriol. 1989, 39, 199–204. [Google Scholar] [CrossRef]
- Canganella, F.; Wiegel, J. Continuous Cultivation of Clostridium thermobutyricum in a Rotary Fermentor System. J. Ind. Microbiol. Biotechnol. 2000, 24, 7–13. [Google Scholar] [CrossRef]
- Sandoval-Espinola, W.J.; Chinn, M.S.; Thon, M.R.; Bruno-Bárcena, J.M. Evidence of Mixotrophic Carbon-Capture by n-Butanol-Producer Clostridium beijerinckii. Sci. Rep. 2017, 7, 12759. [Google Scholar] [CrossRef] [PubMed]
- Ezeji, T.; Qureshi, N.; Blaschek, H.P. Production of Acetone–Butanol–Ethanol (ABE) in a Continuous Flow Bioreactor Using Degermed Corn and Clostridium beijerinckii. Process Biochem. 2007, 42, 34–39. [Google Scholar] [CrossRef]
- Paredes, I.; Quintero, J.; Guerrero, K.; Gallardo, R.; Mau, S.; Conejeros, R.; Gentina, J.C.; Aroca, G. Kinetics of ABE Fermentation Considering the Different Phenotypes Present in a Batch Culture of Clostridium beijerinckii NCIMB-8052. Electron. J. Biotechnol. 2022, 56, 12–21. [Google Scholar] [CrossRef]
- Beschkov, V.; Parvanova-Mancheva, T.; Vasileva, E. Experimental Study of Bio-Hydrogen Production by Clostridium beijerinckii from Different Substrates. Energies 2023, 16, 2747. [Google Scholar] [CrossRef]
- Szymanowska-Powałowska, D.; Orczyk, D.; Leja, K. Biotechnological Potential of Clostridium butyricum Bacteria. Braz. J. Microbiol. 2014, 45, 892–901. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Shen, J.-T.; Wang, X.-L.; Sun, Y.-Q.; Xiu, Z.-L. Stability and Oscillatory Behavior of Microbial Consortium in Continuous Conversion of Crude Glycerol to 1,3-Propanediol. Appl. Microbiol. Biotechnol. 2018, 102, 8291–8305. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Shen, J.-T.; Wang, X.-L.; Sun, Y.-Q.; Xiu, Z.-L. Metabolism, Morphology and Transcriptome Analysis of Oscillatory Behavior of Clostridium butyricum during Long-Term Continuous Fermentation for 1,3-Propanediol Production. Biotechnol. Biofuels 2020, 13, 191. [Google Scholar] [CrossRef]
- Amemiya, T.; Shibata, K.; Yamaguchi, T. Metabolic Oscillations and Glycolytic Phenotypes of Cancer Cells. Int. J. Mol. Sci. 2023, 24, 1914. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Reported examples of oscillation in continuous culture of C. pasteurianum (data extracted from Johnson and Rehmann, 2020 [32]) and C. autoethanogenum (data extracted from Mahamkali et al. [33]). The insert shows the C. pasteurianum data at a different timescale.

Figure 2.
Period length (T) of oscillation as a function of doubling time td (controlled through the dilution rate) in continuous culture of C. pasteurianum (data extracted from Johnson and Rehmann, 2020 [32]), temperature = 35.5 °C, pH 5.0, feed glycerol = 31.1 ± 0.05 g·L−1 and D = 0.066, 0.08 and 0.092 h−1, respectively. The red line shows the number of oscillations per doubling time. The insert shows effect of temperature at a doubling time of 7.5 h in otherwise identical conditions; no oscillation was observed at a temperature of 31 °C.
Figure 2.
Period length (T) of oscillation as a function of doubling time td (controlled through the dilution rate) in continuous culture of C. pasteurianum (data extracted from Johnson and Rehmann, 2020 [32]), temperature = 35.5 °C, pH 5.0, feed glycerol = 31.1 ± 0.05 g·L−1 and D = 0.066, 0.08 and 0.092 h−1, respectively. The red line shows the number of oscillations per doubling time. The insert shows effect of temperature at a doubling time of 7.5 h in otherwise identical conditions; no oscillation was observed at a temperature of 31 °C.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tyszak, A.; Rehmann, L. Metabolic Oscillation Phenomena in Clostridia Species—A Review. Fermentation 2024, 10, 156. https://doi.org/10.3390/fermentation10030156
AMA Style
Tyszak A, Rehmann L. Metabolic Oscillation Phenomena in Clostridia Species—A Review. Fermentation. 2024; 10(3):156. https://doi.org/10.3390/fermentation10030156
Chicago/Turabian StyleTyszak, Annika, and Lars Rehmann. 2024. "Metabolic Oscillation Phenomena in Clostridia Species—A Review" Fermentation 10, no. 3: 156. https://doi.org/10.3390/fermentation10030156
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.