Biological Demalication and Deacetification of Musts and Wines: Can Wine Yeasts Make the Wine Taste Better?


Department of Biology and Environment, Enology Building, School of Life Sciences and Environment, Chemistry Research Centre of Vila Real (CQ-VR), University of Trás-os-Montes and Alto Douro (UTAD), 5000-801 Vila Real, Portugal
Fermentation 2017, 3(4), 51; https://doi.org/10.3390/fermentation3040051
Submission received: 11 September 2017
/
Revised: 23 September 2017
/
Accepted: 27 September 2017
/
Published: 2 October 2017
(This article belongs to the Special Issue Microbiota of Fermented Beverages)
Abstract
:Grape musts sometimes reveal excess acidity. An excessive amount of organic acids negatively affect wine yeasts and yeast fermentation, and the obtained wines are characterized by an inappropriate balance between sweetness, acidity or sourness, and flavor/aroma components. An appropriate acidity, pleasant to the palate is more difficult to achieve in wines that have high acidity due to an excess of malic acid, because the Saccharomyces species in general, cannot effectively degrade malic acid during alcoholic fermentation. One approach to solving this problem is biological deacidification by lactic acid bacteria or non-Saccharomyces yeasts, like Schizosaccharomyces pombe that show the ability to degrade L-malic acid. Excessive volatile acidity in wine is also a problem in the wine industry. The use of free or immobilized Saccharomyces cells has been studied to solve both these problems since these yeasts are wine yeasts that show a good balance between taste/flavor and aromatic compounds during alcoholic fermentation. The aim of this review is to give some insights into the use of Saccharomyces cerevisiae strains to perform biological demalication (malic acid degradation) and deacetification (reduction of volatile acidity) of wine in an attempt to better understand their biochemistry and enological features.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. General Introduction
A balanced wine in terms of flavor should be the goal of every winemaker. Much is known about how taste components, such as acidity, sweetness, savoriness (″umami″) and alcohol can work together in coordination or discord on the palate [1,2,3]. In terms of organic acid content in grapes, malic acid and tartaric acid account for up to 90% of all the acids present [4,5] The organic acid composition of the grape can be influenced by environmental factors such as light, irrigation, humidity, and temperature. However, the cultivar and growing region are also relevant for acid concentration in grapes [6]. In some cases, grape must acidity may need to be corrected. An acidity adjustment involves either acid addition or acidity reduction. In some places, grapes are harvested with a total acidity under 10 g/L or higher.
Acetic acid is the prime component of the volatile acidity of grape musts and wines. Among other causes, acetic acid can be formed as a byproduct of alcoholic fermentation or as a byproduct of the metabolism of acetic and lactic acid bacteria, which can metabolize residual sugars to increase volatile acidity. This acid has a negative impact on yeast fermentation ability and affects the quality of wines when it is present above a given concentration [7].
There are three general methods that winemakers can use to lower high acidity in dry red or white wine production: physical methods (blending and amelioration), chemical methods (bicarbonates), and biological methods (yeast and bacteria). Biological deacidification only affects the malic acid in the total acidity of the wine, and does not reduce tartaric acid. Malolactic fermentation (MLF) is the most common method of biological deacidification or demalication. Lactic acid bacteria (LAB) have the ability to consume the malic acid and convert it to lactic acid [8]. Yeast also has the capability to consume malic acid by converting it into ethanol through malo-ethanolic deacidification. This microbiological process can cause a small increase in the wine’s alcohol content; but, sometimes this is preferred over the aroma and flavor of lactic acid produced by lactic acid bacteria. It has long been known that yeasts like Schizosaccharomyces pombe are especially efficient at converting malic acid [9,10].
Benito et al. [11], used the co-fermentation of two non-Saccharomyces yeast strains in the red wine making process. S. pombe totally consumed the malic acid that was present, while Lachancea thermotolerans produced lactic acid. This process avoided the use of LAB strains to perform malolactic fermentation, thus achieving microbiological stabilization of the wine. The lactic acid produced by L. thermotolerans allowed the wine to achieve its potential acidity and tastiness. The final wines presented more fruity notes and contained less acetic acid and biogenic amines than the controls that had endured malolactic fermentation [11]. However, these yeasts (non-Saccharomyces yeasts) have poor alcohol tolerance, so, in many cases they must be used in conjunction with Saccharomyces yeasts in order to complete wine fermentation. Moreover, according to van Rooyena and Tracel [9], if fermentation and deacidification using Schizosaccharomyces yeasts is planned, it can be expected that at the high fermentation temperatures required for complete deacidification (demalication), the wine quality will be adversely affected in comparison to the use of Saccharomyces cerevisiae together with induced MLF by selected strains of Oenococcus oeni or Lactobacillus plantarum [12].
Regarding excessive volatile acidity in wines, the available solutions (blending the filter-sterilized acidic wine with other wines of lower volatile acidity or using reverse osmosis [13]) are unsatisfactory due to the detrimental aroma/flavor of the final wine. We have studied an empirical biological deacidification process, previously discussed by Ribéreau-Gayon et al. [14], that aims to lower the volatile acidity of wines. This process consists in refermenting the acidic wines by mixing them with marc from a finished wine fermentation, freshly crushed grapes or musts. During the refermentation process acetic acid is consumed by yeasts [15]. We found that indigenous, as well as commercial wine yeasts, were able to metabolize acetic acid during alcoholic fermentation [15] and that deacidification could also be performed directly in acidic wines, without the addition of sugar [16].
Some genomic and hybridization approaches, including global transcription machinery engineering [17], to induce genetic variation have been employed to construct genetically modified yeast able to consume/degrade malic acid and acetic acid. Aiming to select new yeast strains for the industry, several strategies have been applied (Figure 1): (A) Exploring the diversity of natural yeasts from yeast collections, and by studying genotype–phenotype interactions and selecting the most interesting variants; (B) Combining the genomes of multiple strains, thus generating diversity; the new strains obtained are considered non-genetically modified yeasts, so they can be used without restrictions in the industry; and (C) Genetically engineering a new strain by recombining a specific gene (piece of DNA) that is known to confer a specific relevant phenotype for the industry [17]. However, this technique genetically modifies yeasts. Due to the controversy regarding the use of genetically modified organisms, and because of consumer concerns, strains generated using this technique are unlikely to be commercially viable in the near future [18].
Wild yeast strains and their hybrids can be effectively used in acidic grape must fermentation. Another approach to performing demalication consists in improving L-malic acid decomposition on a molecular level. The bacterial mleA genes from Oenococcus oeni and the mae1 genes (malate permease) from Schizosaccharomyces pombe yeast were cloned and co-expressed in S. cerevisiae [19]. The co-expression of the Candida utilis malic enzyme gene (cme1) and S. pombe malate transporter gene (mae1) in S. cerevisiae resulted in complete malate consumption [20]. However, only the work of Husnik et al. [19] was productive; the authors were able to construct a stable genetically engineered wine S. cerevisiae yeast, carrying the mae1 and mleA genes. The recombinant yeast received GRAS status (Generally Recognized as Safe) and is currently commercialized in Moldova and the USA [21].
In 2012, Kunicka-Styczynska and Rajkowska [22] examined the characterization of six commercial wine yeasts expressing high demalication activity. To promote L-malic acid fermentation and alcoholic fermentation, simultaneously, the wine yeasts S. cerevisiae and S. bayanus var. uvarum were crossed, and the natural hybrids were obtained. The hybrids were able to decompose up to 77% of malic acid present in musts [23]. In another study, Redzepovic et al. [24] worked with Saccharomyces paradoxus yeast strain isolated from Croatian grapes, as with S. bayanus and S. cerevisiae (Lalvin and Lallemand-France).
As mentioned, several molecular methods have been developed for bio-deacidification in order to achieve wines with a fine equilibrium between sugar and acid, however, they are limited to the metabolism of malic acid [19,25,26,27,28,29]. Nonetheless, genetically modified strains that substantially decrease acetate yields have been obtained [30,31,32,33].
2. Uptake of Malic Acid into the Yeast Cell
Carboxylic acids are organic compounds that participate in many cellular processes. Previous research has been carried out on yeasts in an effort to characterize the mechanisms of transport of these acids across the yeast cell membrane. Carboxylic acids are weak acids, they partially dissociate in aqueous systems. There is an equilibrium between undissociated, uncharged molecules and their anionic form(s), according to their pKa and to the pH of the medium. This property is important as it influences cell behavior and the mechanisms by which the acids can cross the yeast membrane [34].
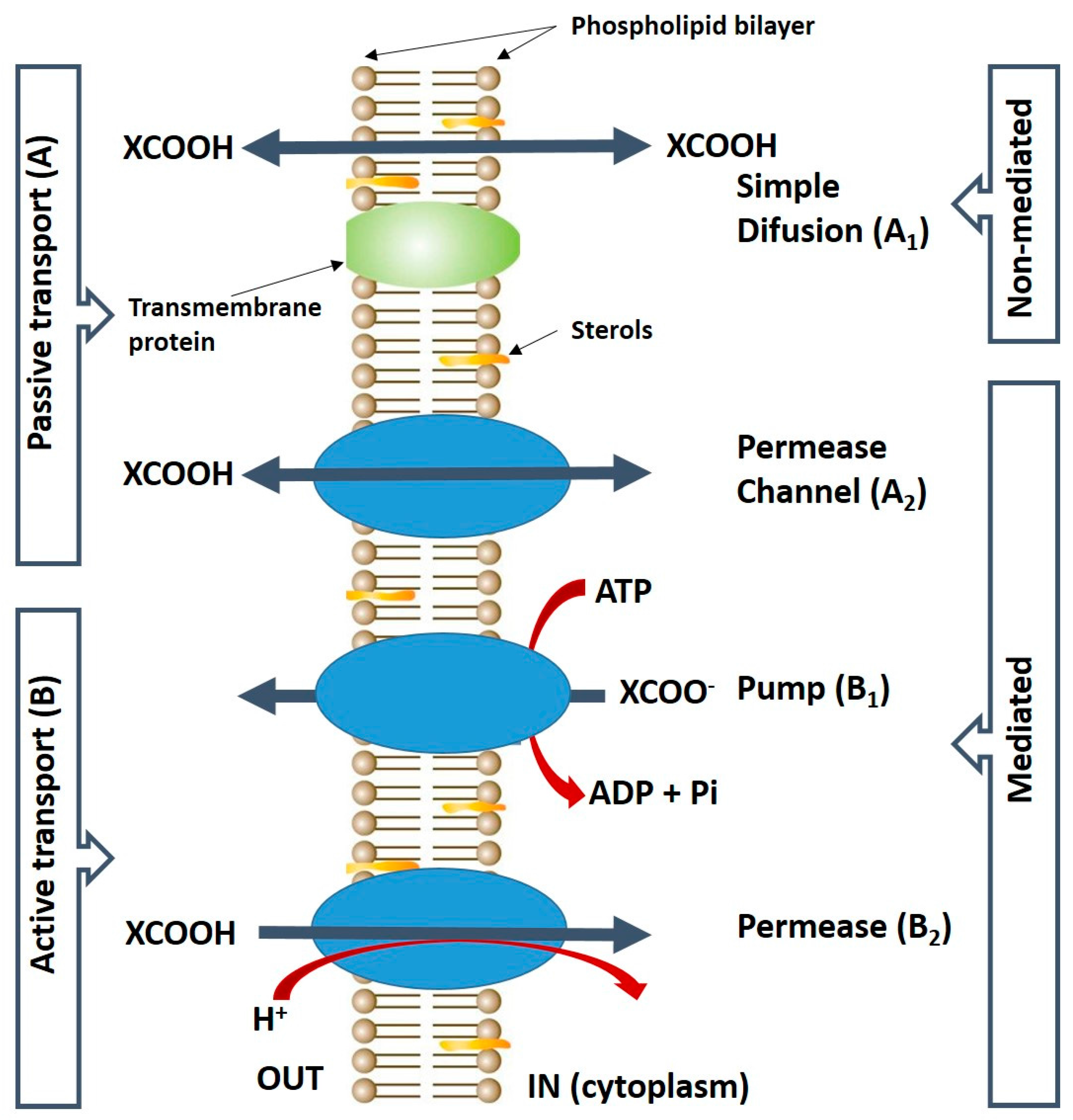
The transport mechanisms of carboxylic acids can be mediated or non-mediated; passive (Figure 2A) or active (Figure 2B), energy-independent or dependent (Figure 2). In (Figure 2A), simple diffusion (Figure 2A1) occurs, and/or facilitated diffusion mediated by a channel or permease protein transport (Figure 2A2), and this involves the transport of the uncharged/undissociated form of the acid. At low pH, the undissociated form of the acid is favored and it may cross the cell membrane by simple diffusion because it is lipid-soluble (Figure 2). Inside the cell cytoplasm, the neutral pH leads to the dissociation of the acid and the acid anion accumulates [34].
The active transport system (Figure 2B), is provided by two distinct processes: B1-pumps associated with yeast stress responses. The extrusion of the anion form of the acid accumulated in the cytoplasm occurs; and permeases B2-associated with metabolic routes of nonfermentable carbon sources [34]. All these transmembrane transporters are currently classified by the Transport Classification Database (http://www.tcdb.org).
The yeast S. cerevisiae has long been known as a poor metabolizer of extracellular malate, due to the lack of a mediated transport system for the acid [35]. The ability of a yeast strain to degrade extracellular l-malate is dependent, among other reasons, for the efficient transport of the dicarboxylic acid into the cell. S. cerevisiae lacks an active transport system for l-malate and extracellular l-malate enters the cells by means of simple diffusion (Figure 2A1); Moreover, the malic enzyme of S. cerevisiae has a considerably lower substrate affinity for l-malate (Km = 50 mM) [21]; and the S. cerevisiae malic enzyme is mitochondrial, whereas the S. pombe malic enzyme is cytosolic [36].
It is also known that strains of S. pombe and Zygosaccharomyces bailii can metabolize l-malate in the presence of glucose or another assimilable carbon source [37]. In contrast, Hansenula anomala, Candida sphaerica, Pichia stipitis and Pachysolen tannophilus are yeasts species able to utilize malic acid as a sole carbon source, although this skill is repressed in the presence of glucose [38]. Low efficiencies on malic acid metabolism have been observed for S. cerevisiae, Candida colliculosa, Candida stellata, and Hanseniaspora uvarum/Kloeckera apiculata [21].
As mentioned above, the ability of a yeast strain to degrade extracellular l-malate is dependent on the efficient transport of the dicarboxylic acid into the cell. The yeasts C. utilis, C. sphaerica, H. anomala and K. marxianus are able to transport malate by a proton symport/induced and glucose repressed system [21,39,40,41], while the transport of this acid is carrier-mediated in Kluyveromyces lactis (active transport/strain dependent and glucose repressed [42]). Z. bailii is also able to transport l-malate by facilitated diffusion for non-dissociated form and the transport is induced by glucose and repressed by fructose [43]. Schizosaccharomyces pombe possesses a proton symport/constitutive transport system [21].
3. The Demalication Activity of Saccharomyces Strains
The ability of Saccharomyces strains to degrade malic acid is strain dependent [24,44,45]. The rather low malate consumption can be explained by the absence of an active l-malate carrier, low substrate affinity and the mitochondrial location of the malic enzyme [46,47,48]. The malic enzyme, under fermentation conditions is regulated by the fermentative glucose metabolism that causes mitochondrial deterioration [20].
Redzepovic et al. [24] found that the indigenous Saccharomyces paradoxus strain RO88 degraded 38% of the malic acid in chardonnay must. The wine produced was of good quality; while S. pombe strain removed a much higher value of malic acid (90%), the wine produced was of poor quality. In the same study, the authors verified that S. cerevisiae Lalvin strain 71B, commercially promoted as a “malic-acid-degrading wine strain”, only degraded 18% of the malic acid. However, expression of the malic enzyme gene from S. paradoxus RO88 and S. cerevisiae 71B increased towards the end of fermentation, after glucose depletion.
3.1. Dicarboxylate Carrier Dic1p (YLR348c) in Yeasts
Lancar-Benba et al. [49] in 1996 isolated the dicarboxylates carrier protein Dic1p, from the mitochondria of yeast. The Dic1p carrier protein transports malate, succinate or malonate. Other molecules such as sulphate and thiosulphate are also exchanged by Dic1p for malate or phosphate. Additionally, this carrier protein is inhibited by the substrate equivalents butylmalonate, benzylmalonate and phenylsuccinate. The Km (transport affinity) of Dic1p for phosphate is 1.65 mM and for malate it is 0.56 mM [50].
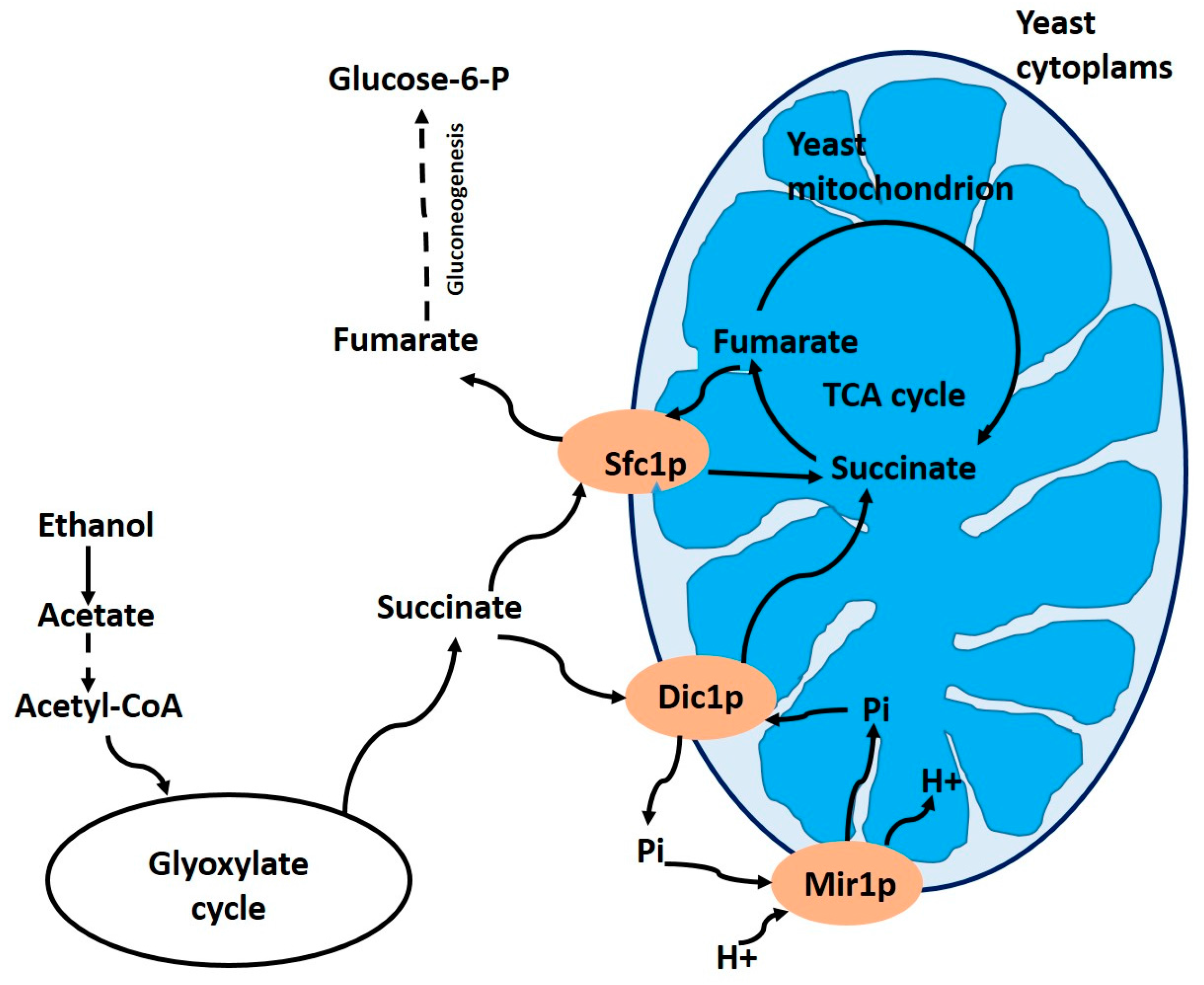
To understand the physiological role of the Dic1p carrier Palmieri et al. [51] constructed a yeast mutant with its depleted gene. They found that there is no other mitochondrial transport system capable of replacing the dicarboxylate carrier studied by Palmieri et al. [52]. This indicates that the main function of the Dic1p carrier is to transport cytoplasmic dicarboxylates into the mitochondrial matrix, working as an essential anaplerotic transporter for the Krebs cycle. The Dic1p carrier catalyzes the import of succinate into mitochondria in exchange for internal phosphate, in wild-type S. cerevisiae growing on ethanol or acetate (Figure 3).
Since phosphate is reprocessed into mitochondria by the Mir1p phosphate carrier, the combined activity of the Dic1p and Mir1p carriers is responsible for the uptake of succinate. Succinate is converted to fumarate, and, subsequently, to oxaloacetate. These reactions that occur within the mitochondria allow the oxidation of acetyl-CoA formally produced from acetate or ethanol (Figure 3).
3.2. The Aspartate/Glutamate Carrier Agc1p (YPR021c) in Yeasts
The product of the yeast gene YPR021c was identified by the “similarity approach” as the yeast orthologue of human AGCs (mitochondrial aspartate/glutamate carriers) and was named Agc1p [51]. It has no N-terminal EF-hand motifs, but the C-terminal domain of Agc1p is 44% identical to AGC1 and 43% identical to AGC2 corresponding domains. Agc1p (C-terminal part) transport properties are indicative that Agc1p is an aspartate/glutamate carrier [53]. AGC performs a key role in the malate/aspartate NADH shuttle in mammalian mitochondria. It promotes the transference of reducing equivalents from the cytosol to the mitochondrial matrix, for electron transport. In yeasts, the enzymes required to complete this shuttle function are also present, but there are some questions about the role of the NADH shuttle [51].
Cavero et al. [53] characterized the physiological role of the Agc1p carrier, and deleted the AGC1 gene from two S. cerevisiae strains (W303 and YPH499). The resultant mutants were not able to grow in minimal synthetic medium supplemented with acetate or oleic acid. Surprisingly they showed viability and grow in a medium supplemented with other non-fermentable carbon sources like ethanol [53]. The hypothesis formulated was that the Agc1p carrier plays a role in the malate/aspartate shuttle, and this role is critical for growth on acetate and fatty acids (Figure 4).
4. Acetic Acid Metabolism in S. cerevisiae
The aroma threshold for acetic acid in wine, depends on the wine variety and style; a concentration of 0.8–0.9 g/L produces a sour aftertaste in wine [54]. According to current legislation [55], the maximum acceptable limit for volatile acidity in red and white table wines is 1.2 g/L of acetic acid [55].
S. cerevisiae is able to metabolize acetic acid during a refermentation process [14,56]. According to the authors and the work done by Vilela-Moura et al. [7,15,16,56] and Vasserot et al. [57], excessive volatile acidity can be removed by re-inoculating (performing a refermentation) with an appropriate S. cerevisiae wine yeast. Under aerobic conditions, acetate can be used as a carbon and energy source for the generation of energy and cellular biomass [58]. In S. cerevisiae, acetate transport and metabolism are subject to glucose repression. However, in chemostat cultures of S. cerevisiae grown in mixtures of acetic acid and glucose, if the glucose concentration is extremely low, the cells will no longer be repressed and will metabolize acetate concomitantly with glucose [59]. In a review by Vilela-Moura et al. [7] it is considered that the cellular uptake/transport of acetate into the yeast cell is an important step for its catabolism. In glucose-repressed yeast cells, at low pH, acetic acid enters mainly by simple diffusion [60]. Ethanol will enhance the passive influx of acetic acid, following the first-order kinetics with a constant rate, which increases exponentially with ethanol concentration [61], see Figure 5.
In 1999, Casal et al. [62] found that the transport carrier Jen1p is required for the uptake of lactate in S. cerevisiae. This transporter is also able to transport acetate. Later, it was found that the protein Ady2p was vital for acetate transport in acetic acid-grown cells [63]. Being the only available energy and carbon source, acetate is metabolized to acetyl coenzyme A (acetyl-CoA) by one of the two ACS proteins: Acs1p (peroxisomal) or Acs2p (cytosolic). Acetyl-CoA enters the mitochondria and is then oxidized in the TCA cycle. Acetyl-CoA, by entering the glyoxylate cycle, is also used to produce succinate, refilling the cell with biosynthetic precursors (Figure 5). This action involves the enzymes Icl1p (isocitrate lyase) in the cytosol, and Mls1p (malate synthase) located in the peroxisome [7]. Moreover, acetyl-CoA is used for the synthesis of macromolecules, requiring active gluconeogenesis [64].
VA Bio-Reduction by S. cerevisiae Yeasts Strains and its Limitations
Consumption of glucose and acetic acid is a characteristic of indigenous and commercial wine yeasts [15,65]. In these works [15,65] the characterization of indigenous and commercial S. cerevisiae strains regarding volatile acidity bio-reduction was performed. Among the strains studied, some exhibited a simultaneous consumption pattern of glucose and acetic acid in refermentation assays of acidic white wines with a synthetic culture medium simulating refermentation with a must (conditions of high glucose/low ethanol concentrations) and refermentation with a marc (conditions of low glucose/high ethanol concentrations). Under conditions where oxygen is limited, all the S. cerevisiae strains studied showed acetic acid degradation efficiencies like those of Z. bailii ISA 1307. Curiously, the commercial strain S26 also revealed the capacity to decrease the volatile acidity of wines in aerobic and limited aerobic conditions by 92% and 61%, respectively [15].
Upon inoculation of grape must or wine, a controlled oxygen environment is important for yeast acetic acid consumption [15]. Moreover, yeast cells need to adapt to the hostile fermentation environment, which gradually changes over time. In fermentation or refermentation, multiple stress conditions are imposed on the yeast cell such as high osmolarity; low pH; sulfur dioxide (SO2); ethanol toxicity; temperature variations; increasing nitrogen limitation; and elevated acetic acid concentrations. All of these stress conditions may lead to reduced cellular growth, cellular death and, consequently, stuck fermentations [14,66,67,68].
A controlled wine oxygenation can be accomplished through micro-oxygenation (MO) where oxygen is added by a stainless steel sparger that produces small bubbles, promoting oxygen dissolution in the must/wine. The influence of different combinations of the initial concentration of acetic acid, ethanol, SO2 and MO on acetic acid removal from wines with an excessive volatile acidity, by two commercial yeasts strains (S26 and S29) was evaluated [16,56]. Both strains (S. cerevisiae S26 and S29) reduced by 78% and 48%, respectively, the volatile acidity of an acidic wine with an initial concentration of 1.0 g/L in acetic acid. Sulfur dioxide in the concentration range of 95–170 mg/L inhibited the capacity of both strains to reduce the volatile acidity under the experimental conditions used. Deacetification of wines with the better performing strain, S26, was related to changes in the concentration of wine volatile compounds. The most pronounced increase was observed for compounds with a fruity flavor: isoamyl acetate and ethyl hexanoate with banana-like flavor and apple-pineapple-like flavor, with an 18- and 25-fold increment, respectively; both values above the detection threshold. Furthermore, deacetification led to increased fatty acids concentration, but with apparently no negative impact on the wine’s sensorial properties [16]. However, in the work of Vasserot et al. [57], the results were not as promising. In their work, a commercial S. cerevisiae strain, used in champagne winemaking, was tested for its ability to metabolize acetic acid during fermentation. The amount of acetic acid metabolized by the yeast strain increased with increasing initial acetic acid concentration. The consumption of acetic acid occurred during the second part of the exponential growth phase. However, when the initial acetic acid concentration was higher than 1.0 g/L, independently of the yeast strain used, the final acetic acid concentration could not be reduced to a legally acceptable level [55]. Acetic acid consumption modified yeast metabolism once more acetaldehyde (with a tart flavor reminiscent of green apples) and less glycerol (reduced wine softness) were produced. Considering the reduction of the NADPH/NADP+ (Nicotinamide Adenine Dinucleotide Phosphate reduced/Nicotinamide Adenine Dinucleotide Phosphate oxidized) ratio, resulting in acetic acid consumption, Vasserot et al. [57] proposed that acetic acid modifies yeast metabolism by reducing the Ald6p activity (NADP+ dependent aldehyde dehydrogenase).
In 2010, Vilela-Moura et al. [56], based on other previous studies of indigenous S. cerevisiae strains [15], tested the applicability of the S. cerevisiae strains (S26 and S29) for the deacidification of a white wine (from V. vinifera cv. Viosinho) and a red wine (from V. vinifera cv. Touriga Nacional), in a pilot study. The initial volatile acidity of the musts-wines was between 1.12–1.15 g/L acetic acid [56]. Moreover, the effect of the refermentation process on the red wine, and of MO on acetic acid removal efficiency and aromatic composition of the wine was also studied.
The results showed that the commercial strains S26 and S29 efficiently reduced acetic acid (43 and 47%, respectively) and sugar (100%) after 264 h of refermentation of the acidic white wine that had grape must added. For red wine deacidification, by using grape must independently of MO, it was observed that 60%–66% of the acetic acid was removed. Wines refermented with the must had significantly lower acetic acid concentration and a higher total SO2 in comparison to the wines deacidified with the addition of grape marcs. The composition of the volatile aroma compounds of the resulting red wines was dependent on the deacidification process used, rather than on MO. Conversely, when residual marc from fermentation and strain S26 was used, the aromatic profile of wine changed and the concentration of aromatic esters increased. Esters are well known for their positive contribution to the wine bouquet with strong fruity aromatic notes. Independently of MO, esters such as ethyl isovalerate (fruity-flavor [69]), ethyl isobutyrate (strawberry, ethereal, buttery, ripe-flavor [69], ethyl 2-methylbutyrate (sweet, floral, fruity, apple flavor [69]) and ethyl propionate (rum-like flavor [69]), were found in higher concentrations [56]. Contrary to what was found by Vasserot et al. [57], the final concentration of acetaldehyde was rather low. The initial concentration in acidic red wine/must and red wine/marc mixtures was 14.3 and 20.0 mg/L, respectively, and increased during deacidification to values between 21–86 mg/L, below the detection limit, that is 100 mg/L [70].
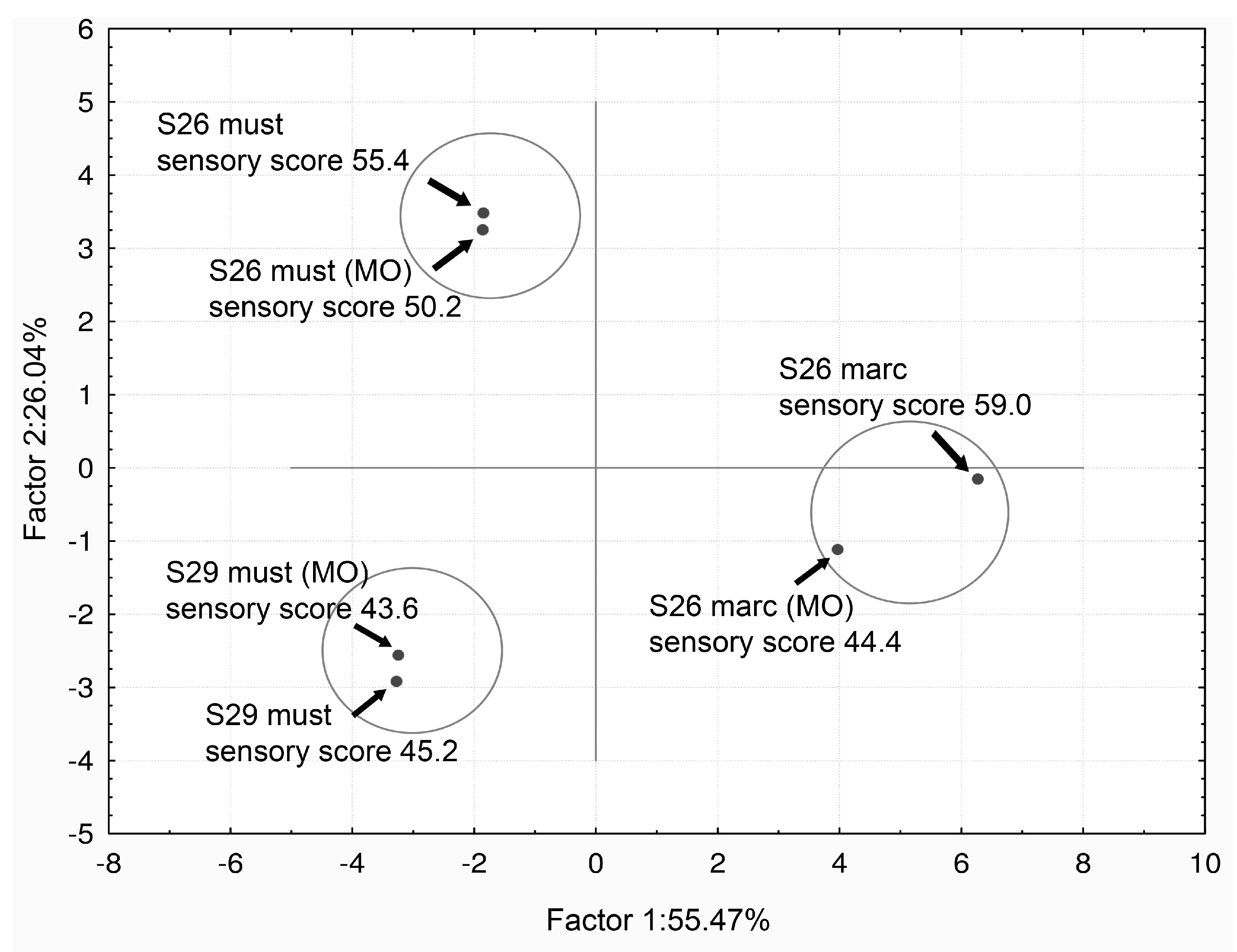
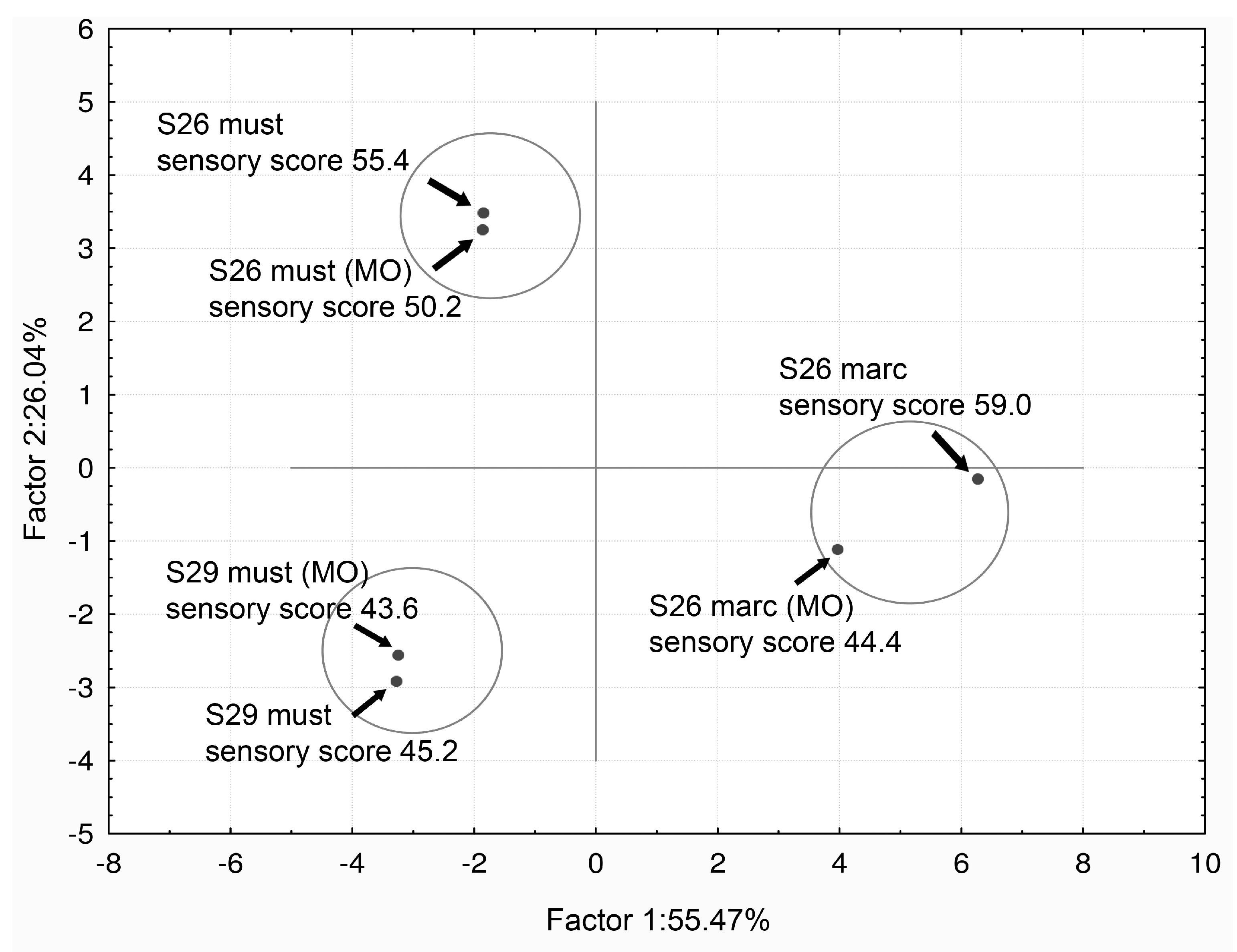
The best sensory classification was gained by the marc-deacidified wine obtained by the use of strain S26 and without MO. Oral perception and aroma (orthonasal sensations) were the sensorial qualities that best distinguished the wines. When data from all analytical and sensorial evaluations were combined, a correlation made by PCA (Principal Component analysis based on correlation matrix) separated the wines into three groups. The separation was according to the strain (S26 or S29) and the refermentation process (must or marc) independent of the oxygen addition (MO), Figure 6 [56].
As mentioned before, yeast cells need to adapt to a hostile environment during fermentation or refermentation, so, cell immobilization by entrapment in beads is a technique that offers some advantages like continuous cell utilization; protection of immobilized cells against inhibitory substances in the fermentation medium; increased fermentation rates; stimulation of production, and protection from shear forces [71]. In 2013 Vilela et al. [72]. studied the efficiency of acetic acid consumption by the previously characterized commercial S26 strain, but the cells were immobilized in a matrix of alginate-chitosan. Several important parameters were analyzed such as initial pH; number/composition of the immobilization matrix; cell concentration and deacetification process efficiency. Immobilized cells were able to reduce 21.6% of the initial volatile acidity 72 h after inoculation. The initial acidic wine had a concentration of 1.1 g/L of acetic acid; ethanol 12.5% (v/v) and a pH of 3.5 h. However, deacetification did not change after 168 h and was accompanied by a slight decrease in ethanol concentration and cell leakage. Initial wine pH adjustment to 3.12 and duplication of initial cell concentration led to an increase in volatile acidity removal up to 61.8%, and no cell leakage from the beads was observed [72].
5. Final Remarks
Production of premium wines is dependent on the optimal adjustment of wine acidity in relation to the other wine components, thus obtaining a balanced wine with an optimum flavor, color and texture profile. The conversion of L-malic acid to L-lactic acid during malolactic fermentation by strains of Lactic Acid Bacteria (LAB) is the traditional method used by winemakers to deacidify wines. Nonetheless, due to intrinsic problems associated with malolactic fermentation (production of acetic acid, for instance) and its unsuitability in some fruity floral wines, other biological methods for the deacidification of wines will be of great benefit to the wine industry.
The use of wine yeast strains with the ability to consume malic acid will reduce the use of LAB and other cellar operations conducted to reduce acidity. Genetically engineered yeasts are considered efficient in malate metabolism but, they cannot always be used because they lack GRAS status. However, the hybrids produced by the natural hybridization of wine yeast are safe but sometimes, and this is a major concern, it is impossible to maintain their genetic stability which can mainly be improved in the process of adaptive evolution. Additionally, many wine yeasts already considered commercial strains, are interspecific hybrids between yeasts of the Saccharomyces sensu stricto complex.
The metabolic influence of yeast on the volatile acidity of wine is undisputable and the right selection of yeast strains determines the performance of the fermentation or refermentation process, leading to higher quality wine. To meet winemakers′ demands, there is an endless search for new wine yeast strains to be used in highly acidic musts and wines. Deacetification of wine with a S. cerevisiae yeast strain may eliminate the use of physical methods like reverse-osmosis or blending wines to decrease volatile acidity, which in turn will reduce the economic losses endured by winemakers.
Acknowledgments
This work is supported by the Chemistry Research Centre of Vila Real (CQ-VR). Additional thanks to the Project NORTE-01-0145-FEDER-000038 (I&D INNOVINE&WINE–Innovation Platform of Vine & Wine).
Conflicts of Interest
The author declare no conflict of interest.
References
- Vilela, A.; Inês, A.; Cosme, F. Is wine savory? Umami taste in wine. SDRP J. Food Sci. Technol. 2016, 1, 1–6. [Google Scholar]
- Vilela, A.; Jordão, A.M.; Cosme, F. Wine phenolics: Looking for a smooth mouthfeel. SDRP J. Food Sci. Technol. 2016, 1, 1–8. [Google Scholar] [CrossRef]
- Jordão, A.M.; Vilela, A.; Cosme, F. From Sugar of Grape to Alcohol of Wine: Sensorial Impact of Alcohol in Wine. Beverages 2015, 1, 292–310. [Google Scholar] [CrossRef]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Rozès, N.; Mas, A.; Guillamón, J.M. Effect of organic acids and nitrogen source on alcoholic fermentation: Study of their buffering capacity. J. Agric. Food Chem. 2003, 51, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.F.; Wu, B.H.; Fan, P.G.; Li, S.H.; Li, L.S. Sugar and acid concentrations in 98 grape cultivars analyzed by principal component analysis. J. Sci. Food Agric. 2006, 86, 1526–1536. [Google Scholar] [CrossRef]
- Lamikanra, O.; Inyang, I.; Leong, S. Distribution and effect of grape maturity on organic acid content of red Muscadine grapes. J. Agric. Food Chem. 1995, 43, 3026–3028. [Google Scholar] [CrossRef]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Silva, R.F.; Chaves, S.R.; Sousa, M.J.; Côrte-Real, M. The impact of acetate metabolism on yeast fermentative performance and wine quality: Reduction of volatile acidity of grape-musts and wines—Minireview. Appl. Microbiol. Biotechnol. 2011, 89, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Inês, A.; Tenreiro, T.; Tenreiro, R.; Mendes-Faia, A. Revisão: As bactérias do ácido láctico do vinho-Parte I. Ciênc. Téc. Vitiviníc. 2008, 23, 81–96. [Google Scholar]
- Van Rooyen, T.J.; Tracel, R.P. Biological Deacidification of Musts Induced by Yeasts or Malolactic Bacteria and the Effect on Wine Quality. S. Afr. J. Enol. Vitic. 1987, 8, 60–69. [Google Scholar] [CrossRef]
- Camarasa, C.; Bidard, F.; Bony, M.; Barre, P.; Dequin, S. Characterization of Schizosaccharomyces pombe Malate Permease by Expression in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2001, 67, 4144–4151. [Google Scholar] [CrossRef] [PubMed]
- Benito, A.; Calderón, F.; Palomero, F.; Benito, S. Combined use of selected Schizosaccharomyces pombe and Lachancea thermotolerans yeast strains as an alternative to the traditional malolactic fermentation in red wine production. Molecules 2015, 20, 9510–9523. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zoecklein, B.W.; Fugelsang, K.C.; Gump, B.H.; Nury, F.S. Volatile acidity. In Wine Analysis and Production, 1st ed.; Chapman & Hall: New York, NY, USA, 1995; pp. 192–198. ISBN 978-1-4757-6967-8. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. The microbiology of wine and vinifications. In Handbook of Enology, 1st ed.; John Wiley & Sons Ltd.: Chichester, UK, 2000; Volume 1, p. 454. ISBN 13:978-0-470-01034-1 (HB). [Google Scholar]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Reduction of volatile acidity of wines by selected yeast strains. Appl. Microbiol. Biotechnol. 2008, 80, 881–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela-Moura, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Effects of acetic acid, ethanol and SO2 on the removal of volatile acidity from acidic wines by two Saccharomyces cerevisiae commercial strains. Appl. Microbiol. Biotechnol. 2010, 87, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. Fems Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [PubMed]
- Schuller, D. Better yeast for better wine—Genetic improvement of Saccharomyces cerevisiae winemaking strains. In Progress in Mycology; Rai, M., Kövics, G., Eds.; Scientific Publishers: Jodhpur, India, 2010; pp. 1–51. ISBN 978-90-481-3713-8. [Google Scholar]
- Husnik, J.I.; Volschenk, H.; Bauer, J.; Colavizza, D.; Luo, Z.; van Vuuren, H.J. Metabolic engineering of malolactic wine yeast. Metab. Eng. 2006, 8, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Saayman, M.; van Zyl, W.H.; Viljoen-Bloom, M. Cloning, characterization, and heterologous expression of the Candida utilis malic enzyme gene. Curr. Genet. 2006, 49, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Saayman, M.; Viljoen-Bloom, M. The biochemistry of malic acid metabolism by wine yeasts—A review. S. Afr. J. Enol. Vitic. 2006, 27, 113–122. [Google Scholar] [CrossRef]
- Kunicka-Styczynska, A.; Rajkowska, K. Phenotypic and genotypic diversity of wine yeasts used for acidic musts. World J. Microbiol. Biotechnol. 2012, 28, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Kunicka-Styczynska, A.; Rajkowska, K. Physiological and genetic stability of hybrids of industrial wine yeasts Saccharomyces sensu stricto complex. J. Appl. Microbiol. 2011, 110, 1538–1549. [Google Scholar] [CrossRef] [PubMed]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- Bony, M.; Bidart, F.; Camarasa, C.; Ansanay, V.; Dulau, L.; Barre, P.; Dequin, S. Metabolic analysis of S. cerevisiae strains engineered for malolactic fermentation. FEBS Lett. 1997, 410, 452–456. [Google Scholar] [CrossRef]
- Husnik, J.I.; Delaquis, P.J.; Cliff, M.A.; van Vuuren, H.J.J. Functional analyses of the malolactic wine yeast ml01. Am. J. Enol. Vitic. 2007, 58, 42–52. [Google Scholar]
- Main, G.L.; Threlfall, R.T.; Morris, J.R. Reduction of malic acid in wine using natural and genetically enhanced microorganisms. Am. J. Enol. Vitic. 2007, 58, 341–345. [Google Scholar]
- Silva, S.; Ramon-Portugal, F.; Andrade, P.; Abreu, S.; Texeira, M.D.; Strehaiano, P. Malic acid consumption by dry immobilized cells of Schizosaccharomyces pombe. Am. J. Enol. Vitic. 2003, 54, 50–55. [Google Scholar]
- Sousa, M.J.; Teixeira, J.A.; Mota, M. Must deacidification with an induced flocculent yeast strain of Schizosaccharomyces pombe. Appl. Microbiol. Biotechnol. 1993, 39, 189–193. [Google Scholar] [CrossRef]
- Remize, F.; Andrieu, E.; Dequin, S. Engineering of the pyruvate dehydrogenase bypass in Saccharomyces cerevisiae: role of the cytosolic Mg2+ and mitochondrial K+ acetaldehyde dehydrogenases Ald6p and Ald4p in acetate formation during alcoholic fermentation. Appl. Environ. Microbiol. 2000, 66, 3151–3159. [Google Scholar] [CrossRef] [PubMed]
- Walkey, C.J.; Luo, Z.; Madilao, L.L.; van Vuuren, H.J.J. The Fermentation Stress Response Protein Aaf1p/Yml081Wp Regulates Acetate Production in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e51551. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Cordero-Bueso, G.; Pretorius, I.S.; Curtin, C.D. Novel wine yeast with mutations in YAP1 that produce less acetic acid during fermentation. FEMS Yeast Res. 2013, 13, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Walkey, C.J.; Madilao, L.L.; Measday, V.; Van Vuuren, H.J.J. Functional improvement of Saccharomyces cerevisiae to reduce volatile acidity in wine. FEMS Yeast Res. 2013, 13, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.; Paiva, S.; Queirós, O.; Soares-Silva, I. Transport of carboxylic acids in yeasts. FEMS Microbiol. Rev. 2008, 32, 974–994. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J.M. L-Malic acid permeation in resting cells of anaerobically grown Saccharomyces cerevisiae. Biochim. Biophys. Acta 1987, 901, 30–34. [Google Scholar] [CrossRef]
- Benito, Á.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.-Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe strains have characteristics that are beneficial for winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef] [PubMed]
- Rodriquez, S.B.; Thornton, R.J. A malic acid dependent mutant of Schizosaccharomyces malidevorans. Arch. Microbiol. 1989, 152, 564–566. [Google Scholar] [CrossRef]
- Gao, C.; Fleet, G.H. Degradation of malic and tartaric acids by high density cell suspensions of wine yeasts. Food Microbiol. 1995, 12, 65–71. [Google Scholar] [CrossRef]
- Côrte-Real, M.; Leão, C.; Van Uden, N. Transport of L(−)malic acid and other dicarboxylic acids in the yeast Candida sphaerica. Appl. Microbiol. Biotechnol. 1989, 31, 551–555. [Google Scholar] [CrossRef]
- Côrte-Real, M.; Leão, C. Transport of malic acid and other dicarboxylic acids in the yeast Hansenula anomala. Appl. Environ. Microbiol. 1990, 56, 1109–1113. [Google Scholar] [PubMed]
- Queirós, O.; Casal, M.; Althoff, S.; Moradas-Ferreira, P.; Leão, C. Isolation and characterization of Kluyveromyces marxianus mutants deficient in malate transport. Yeast 1998, 14, 401–407. [Google Scholar] [CrossRef]
- Zmijewski, M.J.; Macquillan, A.M. Dual effects of glucose on dicarboxylic acid transport in Kluyveromyces lactis. Can. J. Microbiol. 1975, 21, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, K.; Radler, F. The glucose dependent transport of l-malate in Zygosaccharomyces bailii. Antonie Van Leeuwenhoek 1984, 50, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S.; Bauer, F.F. Meeting the consumer challenge through genetically customized wine-yeast strains. Trends Biotechnol. 2002, 20, 426–432. [Google Scholar] [CrossRef]
- Rodriguez, S.B.; Thornton, R.J. Factors influencing the utilization of l-malate by yeasts. FEMS Microbiol. Lett. 1990, 60, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Volschenk, H.; van Vuuren, H.J.J.; Viljoen-Bloom, M. Malo-ethanolic fermentation in Saccharomyces and Schizosaccharomyces. Curr. Genet. 2003, 43, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Osothsilp, C.; Subden, R.E. Isolation and characterization of Schizosaccharomyces pombe mutants with defective NAD-dependent malic enzyme. Can. J. Microbiol. 1986, 32, 481–486. [Google Scholar] [CrossRef]
- Volschenk, H.; Viljoen, M.; Grobler, J.; Petzold, B.; Bauer, F.; Subden, R.E.; Young, R.A.; Lonvaud, A.; Denayrolles, M.; van Vuuren, H.J.J. Engineering pathways for malate degradation in Saccharomyces cerevisiae. Nat. Biotechnol. 1997, 15, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Lancar-Benba, J.; Foucher, B.; Saint-Macary, M. Characterization, purification and properties of the yeast mitochondrial dicarboxylate carrier (Saccharomyces cerevisiae). Biochimie 1996, 78, 195–200. [Google Scholar] [CrossRef]
- Palmieri, L.; Palmieri, F.; Runswick, M.J.; Walker, J.E. Identification by bacterial expression and functional reconstitution of the yeast genomic sequence encoding the mitochondrial dicarboxylate carrier protein. FEBS Lett. 1996, 399, 299–302. [Google Scholar] [CrossRef]
- Palmieri, F.; Agrimi, G.; Blanco, E.; Castegna, A.; Di Noia, M.-A.; Iacobazzi, V.; Lasorsa, F.-M.; Marobbio, C.M.T.; Palmieri, L.; Scarcia, P.; et al. Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim. Biophys. Acta 2006, 1757, 1249–1262. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, L.; Vozza, A.; Honlinger, A.; Dietmeier, K.; Palmisano, A.; Zara, V.; Palmieri, F. The mitochondrial dicarboxylate carrier is essential for the growth of Saccharomyces cerevisiae on ethanol or acetate as the sole carbon source. Mol. Microbiol. 1999, 31, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Cavero, S.; Vozza, A.; del Arco, A.; Palmieri, L.; Villa, A.; Blanco, E.; Runswick, M.J.; Walker, J.E.; Cerdan, S.; Palmieri, F.; et al. Identification and metabolic role of the mitochondrial aspartate-glutamate transporter in Saccharomyces cerevisiae. Mol. Microbiol. 2003, 50, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Alcohols and other volatile compounds. The chemistry of wine stabilization and treatments. In Handbook of Enology, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006; Volume 2, pp. 51–64. [Google Scholar] [CrossRef]
- Office Internationale de la Vigne et du Vin. International Code of Oenological Practices; OIV: Paris, France, 2010. [Google Scholar]
- Vilela-Moura, A.; Schuller, D.; Falco, V.; Mendes-Faia, A.; Côrte-Real, M. Effect of refermentation conditions and micro-oxygenation on the reduction of volatile acidity by commercial S. cerevisiae strains and their impact on the aromatic profile of wines. Int. J. Food Microbiol. 2010, 141, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Vasserot, Y.; Mornet, F.; Jeandet, P. Acetic acid removal by Saccharomyces cerevisiae during fermentation in oenological conditions. Metabolic consequences. Food Chem. 2010, 119, 1220–1223. [Google Scholar] [CrossRef]
- Schüller, H.J. Transcriptional control of nonfermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 2003, 43, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.M.; Gombert, A.K.; Christensen, B.; Olsson, L.; Nielsen, J. Identification of in vivo enzyme activities in the cometabolism of glucose and acetate by Saccharomyces cerevisiae by using 13C-labeled substrates. Eukaryot. Cell 2003, 2, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.; Cardoso, H.; Leão, C. Mechanisms regulating the transport of acetic acid in Saccharomyces cerevisiae. Microbiology 1996, 142, 1385–1390. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.; Cardoso, H.; Leão, C. Effects of ethanol and other alkanols on transport of acetic acid in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1998, 64, 665–668. [Google Scholar] [PubMed]
- Casal, M.; Paiva, S.; Andrade, R.P.; Gancedo, C.; Leão, C. The lactate-proton symport of Saccharomyces cerevisiae is encoded by JEN1. J. Bacteriol. 1999, 181, 2620–2623. [Google Scholar] [PubMed]
- Paiva, S.; Devaux, F.; Barbosa, S.; Jacq, C.; Casal, M. Ady2p is essential for the acetate permease activity in the yeast Saccharomyces cerevisiae. Yeast 2004, 21, 201–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela-Moura, A. Isolation and Characterization of Yeasts: Application in Controlled Processes of Volatile Acidity Bio-Reduction in Wines. Ph.D. Thesis, University os Trás-os-Montes e Alto Douro (UTAD), Vila Real, Portugal, 2010. [Google Scholar]
- Vilela, A.; Amaral, C.; Shuller, D.; Mendes-Faia, A.; Corte-Real, M. Combined use of Wallerstein and Zygosaccharomyces bailii modified differential media to isolate yeasts for the controlled reduction of volatile acidity of grape musts and wines. J. Biotech. Res. 2015, 6, 43–53. [Google Scholar]
- Pizarro, F.; Vargas, F.A.; Agosin, E. A systems biology perspective of wine fermentations. Yeast 2007, 24, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Moreno, R.; Morales, P.; Gonzalez, R.; Mas, A.; Beltran, G. Biomass production and alcoholic fermentation performance of Saccharomyces cerevisiae as a function of nitrogen source. FEMS Yeast Res. 2012, 12, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Eglinton, J.M.; Henschke, P.A. Restarting incomplete fermentations: The effect of high concentrations of acetic acid. Aust. J. Grape Wine Res. 1999, 52, 71–78. [Google Scholar] [CrossRef]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Carlton, W.K.; Gump, B.; Fugelsang, K.; Hasson, A.S. Monitoring acetaldehyde concentrations during MO of red wine by headspace solid-phase micro extraction with on-fiber derivatization. J. Agric. Food Chem. 2007, 55, 5620–5625. [Google Scholar] [CrossRef] [PubMed]
- Genisheva, Z.; Teixeira, J.A.; Oliveira, J.M. Immobilized cell systems for batch and continuous winemaking. Trends Food Sci. Technol. 2014, 40, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Vilela, A.; Schuller, D.; Mendes-Faia, A.; Côrte-Real, M. Reduction of volatile acidity of acidic wines by immobilized Saccharomyces cerevisiae cells. Appl. Microbiol. Biotechnol. 2013, 97, 4991–5000. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of strategies to obtain superior industrial yeast strains: (A) Natural diversity; (B) Artificial diversity and (C) Genetic modification. Adapted from Steensels et al. [17].
Figure 1.
Overview of strategies to obtain superior industrial yeast strains: (A) Natural diversity; (B) Artificial diversity and (C) Genetic modification. Adapted from Steensels et al. [17].

Figure 2.
Schematic representation of the carboxylic acids transport systems in the yeast cell membrane: Carriers (transporters) are represented by the three blue-oval shapes; XCOOH, carboxylic acid. XCOO − , anion form of the acid. Adapted from Casal et al. [34].
Figure 2.
Schematic representation of the carboxylic acids transport systems in the yeast cell membrane: Carriers (transporters) are represented by the three blue-oval shapes; XCOOH, carboxylic acid. XCOO − , anion form of the acid. Adapted from Casal et al. [34].
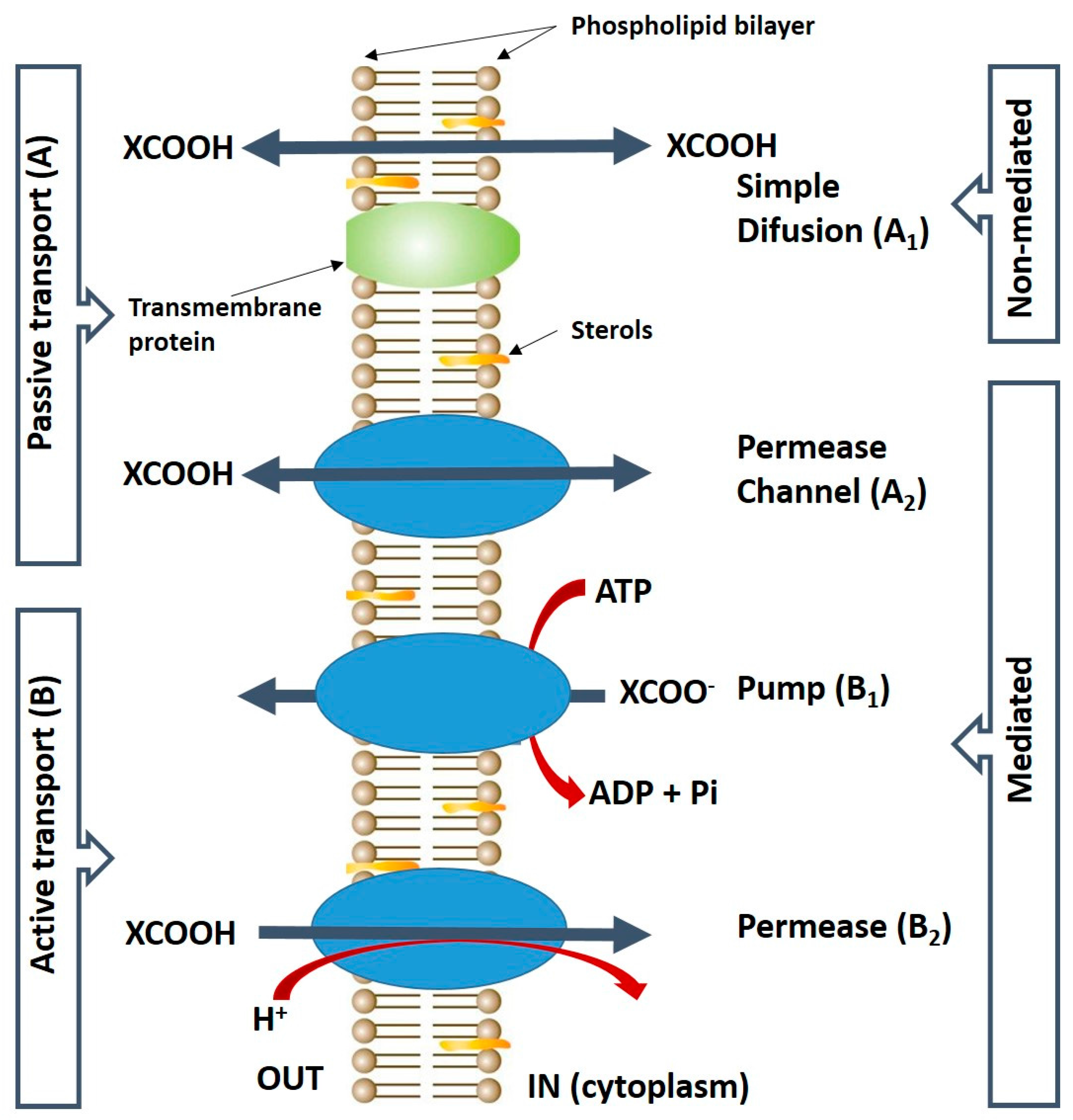
Figure 3.
Schematic representation of the actions of the mitochondrial Dic1p carrier and the Sfc1p mitochondrial succinate/fumarate carrier, when yeast cells are grown on acetate or ethanol as the sole carbon source. Abbreviations: TCA cycle, tricarboxylic acid cycle; Mir1p, mitochondrial phosphate carrier; Pi, phosphate. Adapted from Palmieri et al. [51].
Figure 3.
Schematic representation of the actions of the mitochondrial Dic1p carrier and the Sfc1p mitochondrial succinate/fumarate carrier, when yeast cells are grown on acetate or ethanol as the sole carbon source. Abbreviations: TCA cycle, tricarboxylic acid cycle; Mir1p, mitochondrial phosphate carrier; Pi, phosphate. Adapted from Palmieri et al. [51].
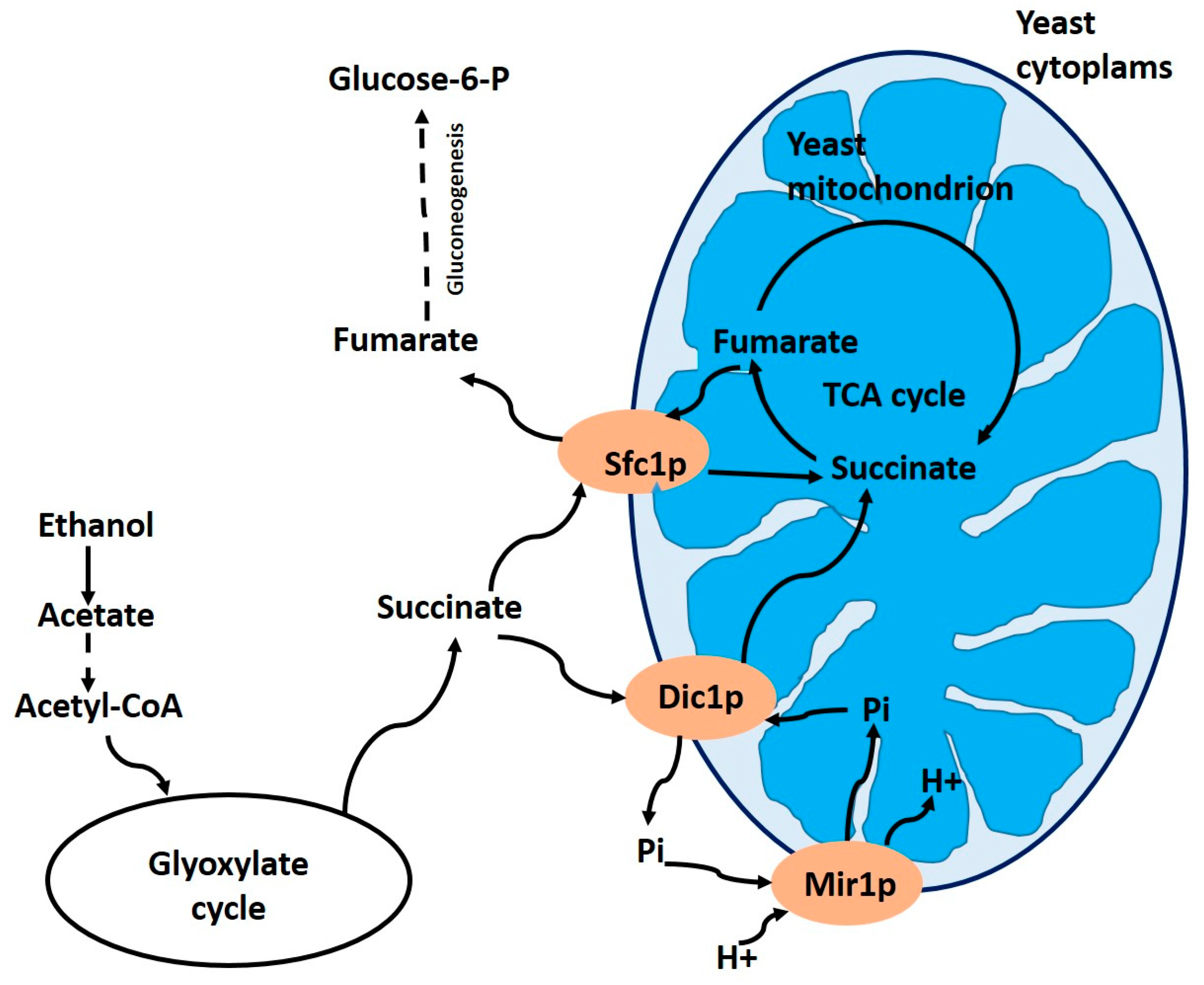
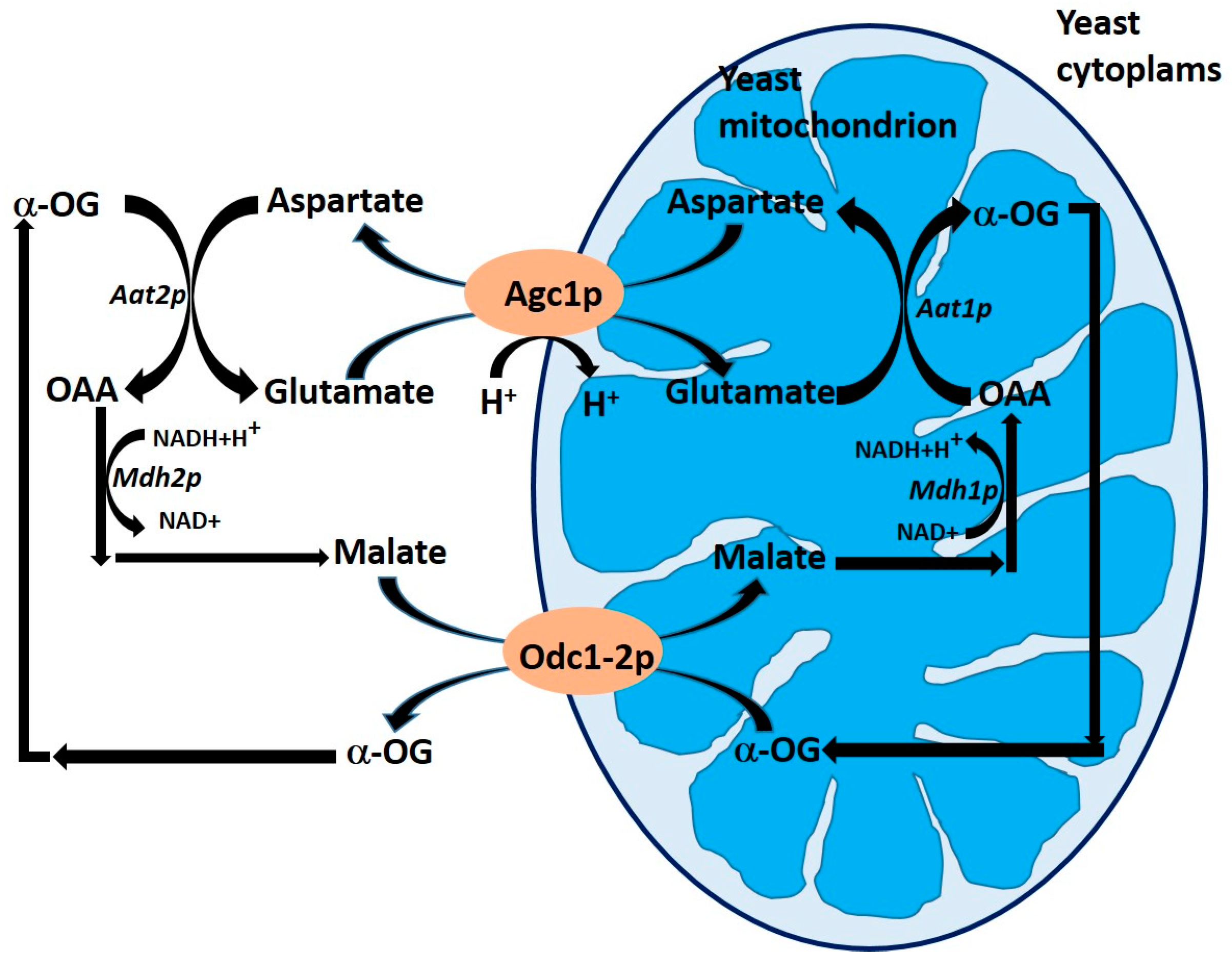
Figure 4.
Schematic representation of the role of oxodicarboxylate Odc1p and Odc2p and mitochondrial aspartate/glutamate Agc1p carriers in the malate/aspartate transport system. Abbreviations: Aat1p, mitochondrial aspartate aminotransferase; Aat2p, cytosolic aspartate aminotransferase; Mdh1p, mitochondrial malate dehydrogenase; Mdh2p, cytosolic malate dehydrogenase; α-OG, 2-oxoglutarate; OAA, oxaloacetate. Adapted from Palmieri et al. [51].
Figure 4.
Schematic representation of the role of oxodicarboxylate Odc1p and Odc2p and mitochondrial aspartate/glutamate Agc1p carriers in the malate/aspartate transport system. Abbreviations: Aat1p, mitochondrial aspartate aminotransferase; Aat2p, cytosolic aspartate aminotransferase; Mdh1p, mitochondrial malate dehydrogenase; Mdh2p, cytosolic malate dehydrogenase; α-OG, 2-oxoglutarate; OAA, oxaloacetate. Adapted from Palmieri et al. [51].
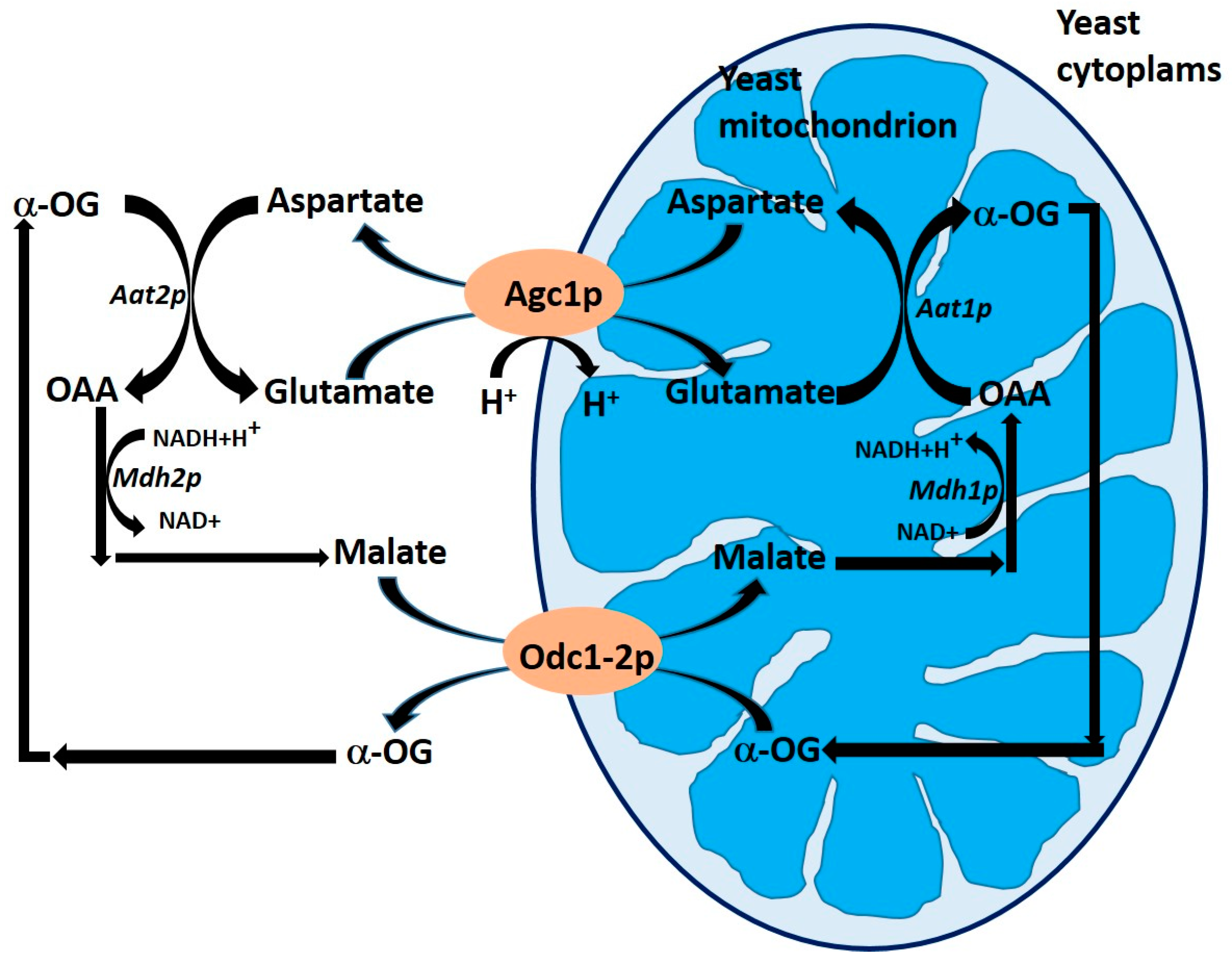
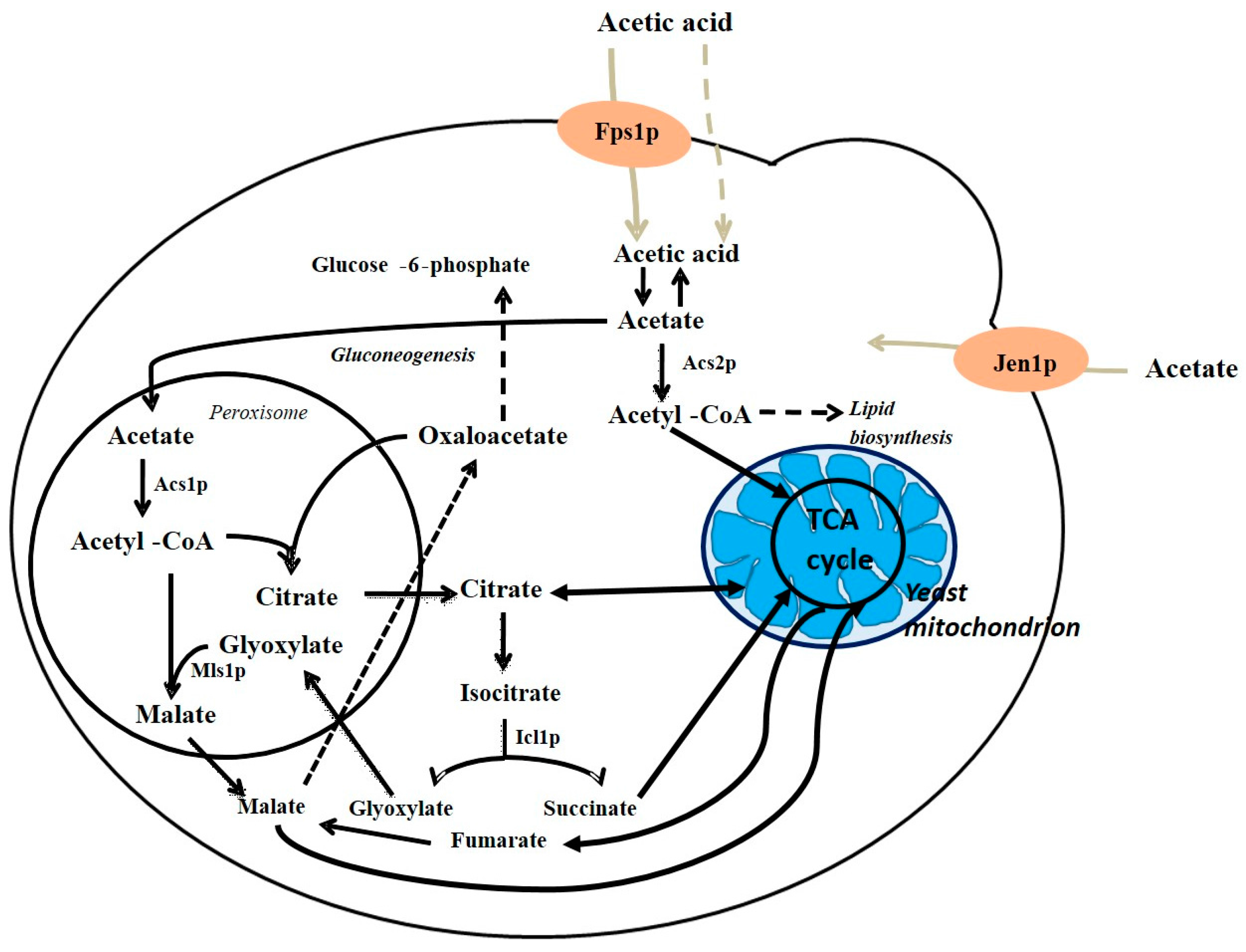
Figure 5.
Cell compartmentation of acetate metabolism in yeast: Black arrows show metabolic pathways and light brown arrows represent acetic acid or acetate transport systems; Fps1p, aquaglyceroporin that mediates the transport of the undissociated form of acetic acid across yeast membranes (solid line); Acetic acid can also enter de cell by simple diffusion (dotted line); Jen1p, S. cerevisiae lactate transporter that is also able to transport acetate. Adapted from Vilela-Moura [64].
Figure 5.
Cell compartmentation of acetate metabolism in yeast: Black arrows show metabolic pathways and light brown arrows represent acetic acid or acetate transport systems; Fps1p, aquaglyceroporin that mediates the transport of the undissociated form of acetic acid across yeast membranes (solid line); Acetic acid can also enter de cell by simple diffusion (dotted line); Jen1p, S. cerevisiae lactate transporter that is also able to transport acetate. Adapted from Vilela-Moura [64].
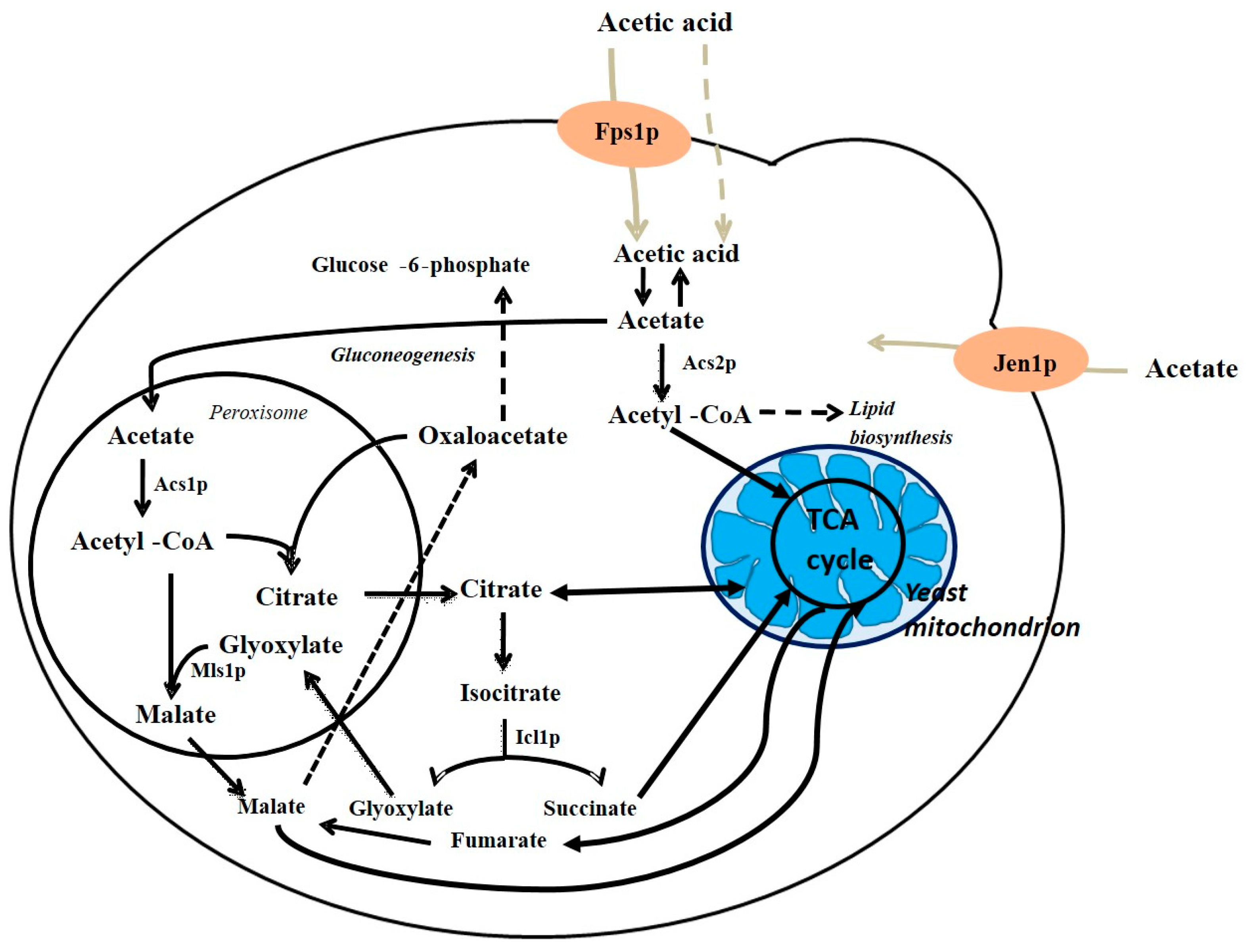
Figure 6.
Bi-dimensional plot of principal component analysis (PCA) values after combining data from chemical and sensorial analysis. Projection of the data according to the strains and the deacidification process. Adapted from Vilela-Moura et al. [56].
Figure 6.
Bi-dimensional plot of principal component analysis (PCA) values after combining data from chemical and sensorial analysis. Projection of the data according to the strains and the deacidification process. Adapted from Vilela-Moura et al. [56].
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vilela, A. Biological Demalication and Deacetification of Musts and Wines: Can Wine Yeasts Make the Wine Taste Better? Fermentation 2017, 3, 51. https://doi.org/10.3390/fermentation3040051
AMA Style
Vilela A. Biological Demalication and Deacetification of Musts and Wines: Can Wine Yeasts Make the Wine Taste Better? Fermentation. 2017; 3(4):51. https://doi.org/10.3390/fermentation3040051
Chicago/Turabian StyleVilela, Alice. 2017. "Biological Demalication and Deacetification of Musts and Wines: Can Wine Yeasts Make the Wine Taste Better?" Fermentation 3, no. 4: 51. https://doi.org/10.3390/fermentation3040051
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.