Bioconversion of Kaempferol and Quercetin Glucosides from Plant Sources Using Rhizopus spp.
by
 ,
,
Nguyen Thai Huynh
1,2,3, 
Guy Smagghe
2,
Gerard Bryan Gonzales
1,2,3,
John Van Camp
3 and
Katleen Raes
1,*
1
Department of Food Technology, Safety and Health, Faculty of Bioscience Engineering, Ghent University—Campus Kortrijk, Graaf Karel de Goedelaan 5, 8500 Kortrijk, Belgium
2
Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000 Ghent, Belgium
3
Department of Food Technology, Safety and Health, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000 Ghent, Belgium
*
Author to whom correspondence should be addressed.
Fermentation 2018, 4(4), 102; https://doi.org/10.3390/fermentation4040102
Submission received: 30 October 2018
/
Revised: 29 November 2018
/
Accepted: 3 December 2018
/
Published: 7 December 2018
(This article belongs to the Special Issue Fermentation and Bioactive Metabolites)
Abstract
:Fermentation with filamentous fungi is known for the ability to convert bioactive compounds. The aim of this research was to investigate the metabolism of glycosidic derivatives of kaempferol and quercetin during fungal fermentation of extracts from cauliflower outer leaves and onion by Rhizopus oryzae and R. azygosporus. The highest release of kaempferol and quercetin was observed after 2 days and 1 day of fermentation with R. oryzae, respectively. It was proposed that glycosidic compounds were initially deglycosylated to form kaempferol-3-glucoside and quercetin-3-glucoside and then further metabolized into their aglycones. Clear differences in conversion efficiency towards the aglycones were observed between the two Rhizopus strains. Although both flavonoids only differ in one hydroxyl group, the metabolism of the glycosides towards their respective aglycones, kaempferol or quercetin, was different. It is concluded that the fermentation with R. oryzae and R. azygosporus could be considered as a way to produce kaempferol and quercetin aglycone from their glycosidic derivatives.
1. Introduction
Kaempferol and quercetin are the dominant flavonoids found generally in vegetables and fruits, linked to the cell wall matrix. While kaempferol derivatives are commonly found in Brassica vegetables [1], onions (Allium cepa) have been reported as one of the major sources of quercetin [2,3]. Quercetin in the form of glucosides is dominantly present in both red and white onion [2,3]. Besides their presence in the edible part of the fruits and vegetables (onion, berries, apples, brassica) [2,3,4,5], these flavonoids are also found in large amounts in waste and by-products i.e., non-edible fractions (e.g., outer leaves, stems, florets), which are inherently linked to fruit and vegetable production. The presence of kaempferol and quercetin in these by-products could be a possible way to valorize these residues as flavonoids are known as antioxidants, and can thus be a protective factor against cardiovascular diseases (hypertension, obesity, arteriosclerosis, smoking, diabetes, and aging) [6,7]. Fukumoto et al. [8] stated that glycosylated phenolic compounds resulted in a lower antioxidant activity compared to their aglycones. The more sugar moieties attached to the aglycone, the more their antioxidant activity is reduced.
Microbial fermentation has been widely used for the biotransformation of phenolic compounds. In particular, the degradation of the cell wall matrix, as well as deglycosylation of phenolic compounds into their corresponding aglycones could be performed by microorganisms, which are known to produce a range of enzymatic activities, e.g., cellulase, pectinase, β-glucosidase [9,10,11], naringinase [12], α-rhamnosidase and hesperidinase [13]. Among these, β-glucosidase is well known for its ability to remove glucose moieties attached to flavonoids at the C3 and C7 positions [14], while α-L-rhamnosidase is responsible for the cleavage of terminal α-L-rhamnose in many natural glucosides, e.g., naringin, rutin, quercitrin, hesperidin, diosgene, terpenyl glycosides [15]. Incubation of soybeans (Glycine max.) with Bacillus pumilus, Bacillus subtilis, or Aspergillus oryzae resulted in the metabolism of daidzin, glycitin, and genistin into their isoflavone aglycones (daidzein, glycitein, and genistein) [16,17,18]. Fermentation of kaempferol-3-glucoside with Bifidobacterium pseudocatenulatum or quercetin-3-glucoside with Aspergillus awamori resulted in the formation of their aglycones [13,19]. Although the main structural difference between quercetin and kaempferol is limited, i.e., a hydroxyl group added to 3′-position on ring B, it is demonstrated that this results in a different bacterial metabolism [20,21,22].
In our previous study [23], lower glycosylated forms of kaempferol were found when the matrix (cauliflower outer leaves) was fermented with different filamentous fungi, but the formation of kaempferol as aglycone was not observed. Therefore, the objective of this study was to evaluate whether the cell wall matrix is a barrier to the formation of the flavonoid aglycone from their parent compounds. The effect of the structure of flavonoids on the extent of deglycosylation was also studied. For this purpose, the fermentation of glycosidic kaempferol and quercetin derivatives extracted from their matrices, i.e., cauliflower outer leaves and onion respectively, with two different filamentous fungi, i.e, Rhizopus oryzae and Rhizopus azygosporus, was performed.
2. Results
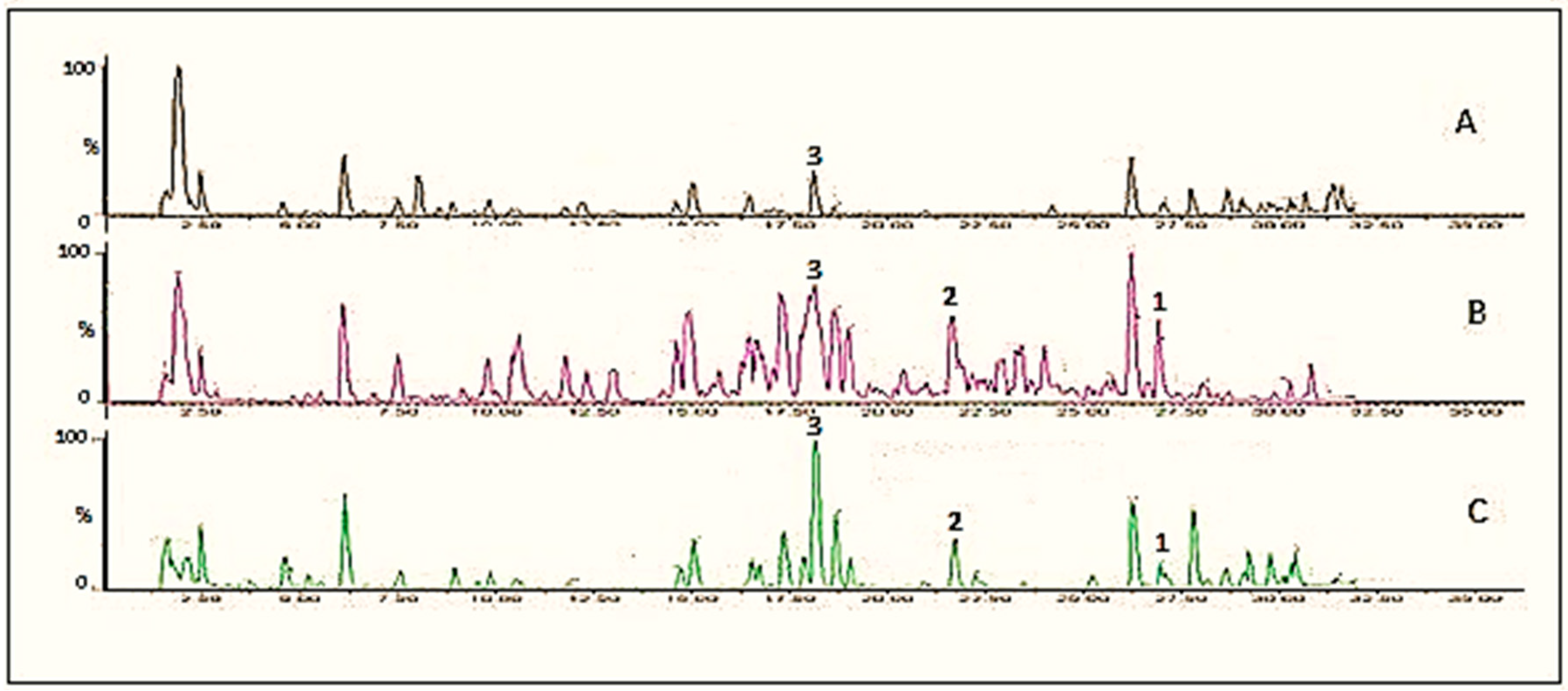
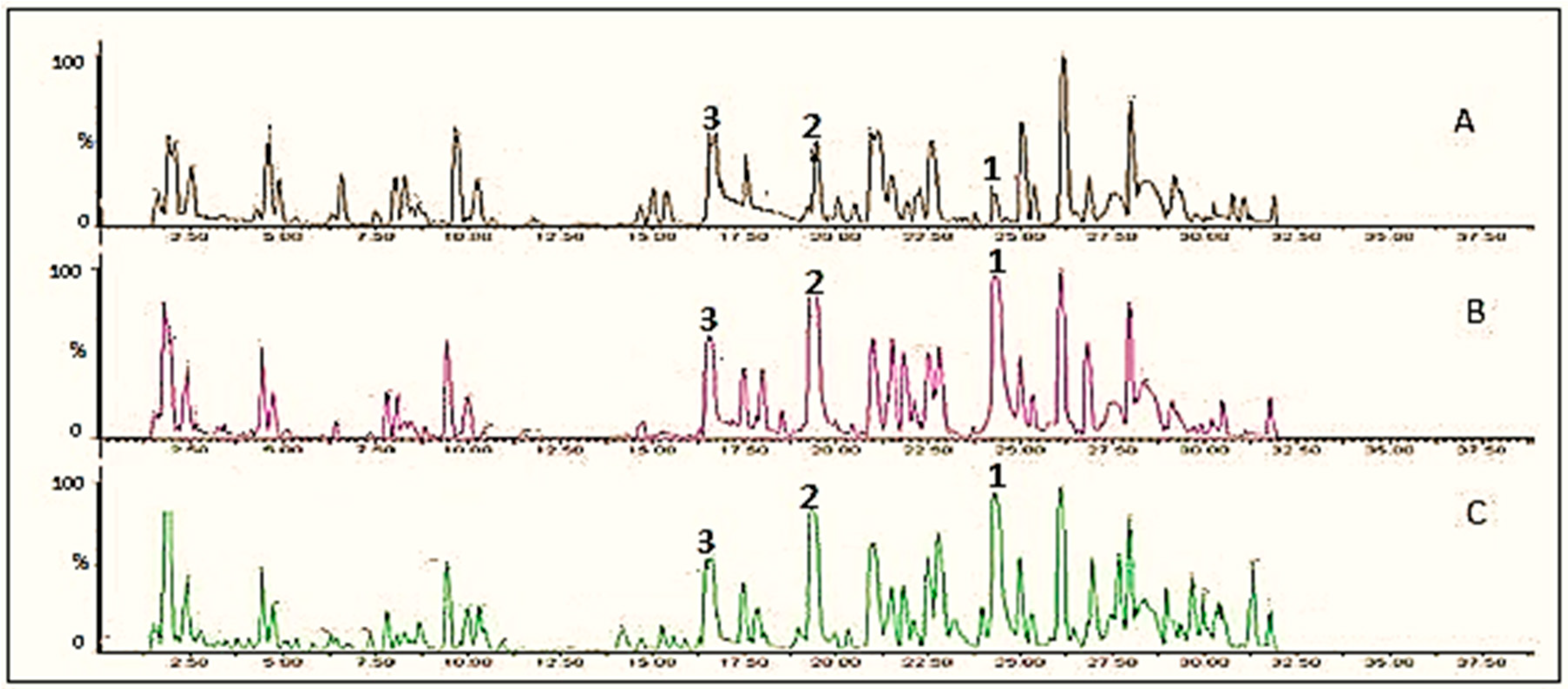
The profile of phenolic compounds of the extracts from cauliflower outer leaves and onion powder and their metabolites was analyzed at different time points of fungal incubations (Figure 1 and Figure 2 respectively). The identification of phenolic compounds (kaempferol, kaempferol-3-O-glucoside, kaempferol-3-O-diglucoside; quercetin, quercetin-3-O-glucoside, quercetin-3-O-glucoside-7-O-glucoside) was carried out through LC-MS/MS based on their retention time, exact mass, and fragments, the results of which are summarized in Table 1. The change in the main constituents during the fermentation was determined based on their peak area (mAU). Figure 3 and Figure 4 show the accumulation of some major kaempferol and quercetin metabolites during 7 days fermentation of extracts of cauliflower outer leaves and onion.
As shown in Figure 3, both fermentation treatments with R. oryzae and R. azygosporus increased the amount of kaempferol-3-O-glucoside released, with the highest level obtained on the 2nd day of incubation for R. oryzae and on 4th day for R. azygosporus, i.e., a 4.8 fold and 8.6 fold increase in the peak area compared to its content in the control sample, respectively. While the increase in the peak area of kaempferol-3-O-diglucoside was only observed on the 1st day of incubation by R. oryzae and R. azygosporus, with an increase of 50% and 12%, respectively, compared to the control samples (t = 0 h).
Regarding the production of kaempferol aglycone from the extract of cauliflower outer leaves, kaempferol was found from day 1 until day 7 of fermentation with R. oryzae, reaching its highest peak area at day 2 and decreased on the 4th day of incubation, while only a very low amount was detected in the sample fermented for 4 days with R. azygosporus prior to disappearing on day 7.
To obtain more information on the possible metabolism of different glycosidic flavonoids, extracts from onion powder, which has been reported to contain mainly quercetin glucosides, was also incubated for 7 days with R. oryzae and R. azygosporus. The change in the peak areas of the three main flavonoid compounds identified, i.e., quercetin, quercetin-3-O-glucoside, and quercetin-3-O-glucoside-7-O-glucoside, during the incubation period is presented in Figure 4. Both treatments with R. oryzae as well as with R. azygosporus after 1 day of incubation resulted in a more than 10-fold higher peak area of quercetin. However, a longer fermentation up to 4 days led to a reduction in the peak area of quercetin, as observed for the incubations with R. azygosporus. A similar pattern of time of incubation was observed with quercetin-3-O-glucoside. This compound increased significantly after 1 day of fermentation with R. oryzae and R. azygosporus, to 190% and 50% of its concentration in the control sample, respectively, and decreased gradually with the time of incubation.
In contrast to the changes observed for the released amounts of quercetin and quercetin-3-O-glucoside, these amounts of quercetin-3-O-glucoside-7-O-glucoside decreased continuously during the fermentation by both R. oryzae and R. azygosporus, with a 5-fold lower peak area after 4 days incubation compared to the control (Figure 4).
3. Discussion
The use of filamentous fungi in solid-state fermentations of agricultural by-products and industrial waste from food processing is widely performed, mostly focusing on enzyme production and release of bioactive compounds. Since agricultural by-products are rich in phenolic compounds, fungal solid-state fermentation can be a possible way to improve their valorization potential. However, the possible metabolism of phenolic compounds by these fungi is poorly characterized and should be considered when using this valorization strategy. In this study, two Rhizopus strains, i.e., R. oryzae and R. azygosporus were selected, based on the results obtained in a former study using cauliflower outer leaves [24]. In the former study [23], the major low molecular weight kaempferol metabolites that were formed on the cauliflower outer leaves were kaempferol-3-O-diglucoside and kaempferol-3-O-glucoside. No kaempferol as aglycone was observed in that study, probably because the whole cell wall matrix was incubated with the fungi. Using extracts from the plant material, as was done in this study, resulted in the production of the aglycones. Therefore, it could be suggested that the presence of the free form of the aglycone (kaempferol and quercetin) detected in the fermented medium indicated the presence of β-glucosidase in the medium. The β-glucosidase secreted by these fungi was possibly able to remove the glucose molecule both on the 3-O and 7-O position. From this study, and from our previous work [23], it becomes clear that the location or bounding of the flavonoids glucosides on the cell wall material is an important factor to consider when the aim is to produce aglycone flavonoids from plant material by microbial fermentations. More research is needed to understand the location and the linkage between flavonoids and cell wall structures. If the type of binding and location of linkage is known, one could select better the enzymes or microbes needed to degrade these linkages. Furthermore, a specific and detailed information of all phenolic structures, especially in terms of glycosides linkages would help to select those enzymes able to specifically degrade certain bounds.
As shown in previous studies, the strains used in this work have the ability to produce cell-wall degrading enzymes through fermentation, such as β-glucosidase produced by R. oryzae [25,26], as well as by R. azygosporus [27,28]. The action of this enzyme may result in the bioconversion of these compounds into their corresponding metabolites and could have been involved in the increase in the release of kaempferol-3-O-glucoside. It is hence proposed that the production of kaempferol-3-O-glucoside could originate from various kaempferol glucosides present in the extract, such as kaempferol-3-O-diglucoside-7-O-glucoside, kaempferol-3-O-diglucoside-7-O-diglucoside, kaempf-erol-3-O-triglucoside-7-O-diglucoside, kaempferol-3-O-triglucoside and kaempferol-3-O-diglucos-ide, as determined in earlier research [23]. Similarly, quercetin glucosides from onion extract, e.g., quercetin-diglucoside, rutin [29,30] may be converted into quercetin-3-O-glucoside under fermentation conditions, and thus lead to its increase during incubation with R. oryzae or on the first day with R. azygosporus. The amount of quercetin-3-O-glucoside increased as a possible explanation for the reduction of quercetin-3-O-glucoside-7-O-glucoside during fermentation.
The cleavage of the 3-O-glycosidic bond of the sugar moiety to the aglycone molecule is in agreement with the previous findings that the aglycone could come from their corresponding glycosidic form by the activity of β-glucosidase, such as the bioconversion of rutin into quercetin by A. awamori [13] or A. niger [31]; daidzin, glycitin, genistin into daidzein, glycitein, genistein by A. oryzae [18] or R. azygosporus, Rhizopus sp. [27].
Among the two fungal strains employed in this work, R. oryzae may exhibit a greater ability to produce β-glucosidase or to produce a more active enzyme than R. azygosporus. Indeed, Lee et al. [27] reported low β-glucosidase activity derived from R. azygosporus, while R. oryzae has been considered as a good producer of this enzyme [24,32]. This could cause the higher amount of kaempferol aglycone found in the fermentation medium prepared with R. oryzae than the one incubated with R. azygosporus. Another possible explanation for our results is that R. azygosporus probably produces phenolic-degrading enzymes with a higher activity, which immediately degrade the formed kaempferol produced into other (non-) phenolic metabolites during fermentation [22,33,34]. This results in the lack of or in the presence of a modest amount of kaempferol in fermented medium with R. azygosporus.
Although the biotransformation of glycosylated forms into their aglycones (kaempferol and quercetin) undergo the same pathway for the removal of sugar molecules attached at the 3- and/or 7-position, the formation of kaempferol was limited for R. azygosporus during fermentation except on the 4th day where a modest amount was detected. On the contrary, a higher accumulation of quercetin was found in the fermented medium, which may be attributed to the existence of an additional –OH group at the 3′-position on ring B of quercetin compared to the kaempferol structure. Oka et al. [20] and Lin et al. [21] indicated that a hydroxyl group on the aromatic ring A and B of both flavonoids resulted in a considerable decrease in maximum velocity and Michaelis constant of the phenolic-degrading enzymes. Therefore, quercetin, which possesses one extra –OH group at the 3′-position, was degraded at a lower rate compared to kaempferol during fungal degradation. This could also be a reason for the faster degradation of kaempferol by R. oryzae after 2 days when its accumulation in fermented medium seems to be sufficient for fungal detoxification as mediated by the fungal enzymes [35].
4. Materials and Methods
4.1. Material
4.1.1. Plant Material
Outer fresh leaves of cauliflowers were collected during the harvest of the cauliflower (July 2012) from a local non-organic farm in West-Flanders, Belgium. The leaves were stored at −18 °C until further analysis. Onion powder was kindly provided by Raps Benelux (Best, The Netherlands).
4.1.2. Chemicals and Reagents
HPLC-grade methanol and formic acid were purchased from VWR International (Leuven, Belgium), and gallic acid, hesperetin, and rutin from Sigma-Aldrich (Bornem, Belgium). Dox Czapek, malt extract, soy peptone, bacteriological agar, casein tryptone were purchased from Oxoid (Erembodegem, Belgium).
4.1.3. Fungal Strains and Preparation of the Inoculum
The cultures of two food-grade fungi, i.e., R. oryzae (CBS 1125.86, isolated from tempeh, Singapore, BSL-level 1), R. azygosporus (CBS 357.93, isolated from tempeh, Java, BSL-level 1) were used and kept at 4 °C on malt extract agar (MEA) slant. From these slants, few spores were brought on new MEA slants, and these slants were incubated for 7 days at 30 °C to obtain the subcultures. After 7 days incubation, 2 mL Tween 80 (0.05%) was added on the slant. This inoculum suspension was mixed with 5 g sterile glass beads in a sterile Erlenmeyer flask, and shaken for 1 min. Then the suspension was centrifuged (2000× g for 20 min) to remove the mycelia. After discarding the supernatants, the pellet was suspended with saline solution (0.85% NaCl) and centrifuged 2 times (2000× g for 20 min. The pellet obtained was used to prepare the spore suspension at an initial inoculum concentration of (106 spores/mL as determined by microscopic measurements) for the fermentation process.
4.2. Experimental Procedure
4.2.1. Preparation of Extract
The samples (cauliflower outer leaves, onion powder) were extracted by the method of Olsen et al. [36]. In brief, after weighing 5 g sample, 15 mL of methanol was added, followed by mixing for 40 s using an ultraturrax. The homogenate was incubated in ice water for 15 min. After centrifugation (13,000× g, 10 min and 4 °C), the supernatant was collected. To the pellet, 10 mL methanol/water (80/20; v/v) was added and re-extracted as described before. Then, both supernatants were pooled and filtered through filter paper (VWR, grade 413, VWR International, Leuven, Belgium). The total volume of the extract was made up to 25 mL final volume by adding methanol. Finally, methanol present in the extract was evaporated using a rotary evaporator to obtain a concentrated extract which was used in this experiment.
4.2.2. Fermentation Process
The inoculum (1 mL) for the different fungi prepared as described in the procedure was added to an Erlenmeyer flask containing 10mL of Dox Czapek medium, the flasks were then incubated for 3 days at 30 °C with shaking. After the third day, 1 mL of phenolic extracts (obtained from cauliflower outer leave or onion powder) was added to the Erlenmeyer flasks. The extracts were added after 3 days preincubation to avoid any antimicrobial effect of the phenolic compounds towards the fungi. The extracts were fermented with R. oryzae, R. azygosporus during 7 days, and samples were taken on day 0, 1, 2, 4 and 7. Control fermentations were done, consisting of the same medium composition, without inoculation with the fungi. All fermentations were done in three independent replicates.
4.3. Analytical Methods
After fermentation, 10 mL methanol (80%) was added to the Erlenmeyer flasks, and the mixture was homogenized for 40 s using an Ultraturrax. This sample extraction was based on the method of Olsen et al. [36], as described above. Further identification of the phenolic compounds in the extracts was done as described in our previous studies [23,37]. Briefly, 1 mL methanol extract (1 mL) was diluted to a final volume of 20 mL using water (0.1 % formic acid). Then the internal standard (100 µL of 100 µg/mL hesperitin) was added. Preconditioning of the SPE C18 column was performed using MeOH (0.1 % formic acid). The diluted methanol extract was then loaded onto a SPE C18 column (50 mg, 4 mL), followed by washing the column with 5 mL water (0.1 % formic acid), and then by adding 3 mL of MeOH (0.1 % formic acid). The eluent obtained was dried under nitrogen, and the residue was re-dissolved in 100 µL MeOH (0.1 % formic acid), followed by adding 900 µL water (0.1 % formic acid). The extracts obtained after SPE purification were analyzed through UPLC-DAD (Liquid Chromatography-Diode Array Detector) with a Waters Acquity UPLC system (Waters Corp., Milford, MA, USA), using a bride ethylene hybrid (BEH) C18 column (150 × 2.1 mm, 1.7 µm particle size). The temperature of the column was set at 40 °C, and of the autosampler at 4 °C.The mobile phase consisted of solvent A (0.1 % formic acid in water) and solvent B (0.1 % formic acid in methanol). The following elution gradient was used: 0 min, 10 %B; 0–6 min, 20% B linear; 6–12 min, 20% B isocratic; 12–13 min, 30% B linear; 13–23 min, 50% B linear, 23–30 min, 90% B linear; 30–35 min, 90% isocratic; 35–40 min, 10% B linear; and 40–45 min 10% isocratic. A flow-rate of 250 µL/min was used. DAD detection was carried out between 280 nm and 370 nm. A Synapt HDMS-TOF mass spectrometer (Micromass, Manchester, UK) equipped with an electrospray ionization (ESI) source was connected to an ACQUITY UPLC system. The settings for the MS were as follows: capillary voltage, 2 kV; sampling cone voltage, 40V; extraction cone voltage, 4 V; source temperature 150 °C; desolvation temperature, 350 °C; cone gas flow rate, 50 L/h; desolvation gas flow rate, 550 L/h. Collision energies were set at 45 V and 6 V for high and low energy, respectively. The spectra were recorded between 100 and 1500 Da using MassLynx software (Waters, Corp.).
5. Conclusions
This study evaluated the bioconversion of phenolic compounds supported by food-degrading microorganisms. The fermentation with filamentous fungi used in this study (R. oryzae and R. azygosporus) revealed that glucoside derivatives of kaempferol and quercetin extracted from cauliflower outer leaves and onion can be deglycosylated to form their precursor products (kaempferol-3-O-glucoside and quercetin-3-O-glucoside), which were further metabolized into their corresponding aglycones (kaempferol and quercetin).it As both kaempferol and quercetin aglycones were formed using phenolic-rich extracts, it can be concluded that cell wall material can be a barrier to produce flavonoid aglycones. Further researches need to be performed on fungal metabolism of various flavonoids with different substitutions in hydroxyl groups to get a better insight into the impact of flavonoids structure on the interaction between the enzyme and the phenolic compound.
Author Contributions
Methodology: N.T.H. & K.R.; Writing Original Draft Preparation: N.T.H.; Formal Analysis: N.T.H. & G.B.G.; Writing-Review & Editing: J.V.C., G.S., and K.R.; Supervision: J.V.C., G.S. & K.R.
Funding
Huynh T. Nguyen is grateful for the PhD-grant financed by MOET-Vietnam. The authors thank the Special Research Fund (BOF) of the Ghent University (BOF05V02512) and the Hercules foundation (AUGE028 and AUGE014) for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollman, P.C.H.; Katan, M.B. Dietary flavonoids: Intake, health effects and bioavailability. Food Chem. Toxicol. 1999, 37, 937–942. [Google Scholar] [CrossRef]
- Pérez-Gregorio, R.M.; García-Falcón, M.S.; Simal-Gándara, J.; Rodrigues, A.S.; Almeida, D.P.F. Identification and quantification of flavonoids in traditional cultivars of red and white onions at harvest. J. Food Compos. Anal. 2010, 23, 592–598. [Google Scholar] [CrossRef]
- Hakkinen, S.H.; Karenlamp, S.O.; Heinonen, I.M.; Mykkanen, H.M.; Torronen, A.R. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J. Agric. Food Chem. 1999, 47, 2774–2779. [Google Scholar] [CrossRef]
- Erlun, I. Review of the flavonoids quercetin, hesperetin and naringenin. Dietary sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 2004, 24, 851–874. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Mateos, A.; Heiss, C.; Borges, G.; Crozier, A. Berry (poly)phenols and cardiovascular health. J. Agric. Food Chem. 2014, 62, 3842–3851. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Zheng, Z.; Shetty, K. Solid-state bioconversion of phenolics from cranberry pomace and role of Lentinus edodes β-glucosidase. J. Agric. Food Chem. 2000, 48, 895–900. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Huang, D. Food grade fungal stress on germinating peanut seeds induced phytoalexins and enhanced polyphenolic antioxidants. J. Agric. Food Chem. 2011, 59, 5993–6003. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Yu, H.S.; Feng, B.; Kang, L.P.; Pang, X.; Xiong, C.Q.; Zhao, Y.; Li, C.M.; Zhang, Y.; Ma, B.P. Selective hydrolysis of flavonoid glycosides by Curvularia lunata. Chin. J. Nat. Med. 2013, 11, 684–689. [Google Scholar] [CrossRef]
- Lin, S.; Zhu, Q.; Wen, L.; Yang, B.; Jiang, G.; Gao, H.; Chen, F.; Jiang, Y. Production of quercetin, kaempferol and their glycosidic derivatives from the aqueous-organic extracted residue of litchi pericarp with Aspergillus awamori. Food Chem. 2014, 145, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Rho, H.S.; Kim, D.H.; Chang, I.S. Enzymatic preparation of kaempferol from green tea seed and Its antioxidant activity. J. Agric. Food Chem. 2006, 54, 2951–2956. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Yadav, P.K.; Yadav, S.; Yadav, K.D.S. α-l-Rhamnosidase: A review. Process Biochem. 2010, 45, 1226–1235. [Google Scholar] [CrossRef]
- Cho, K.M.; Hong, S.Y.; Math, R.K.; Lee, J.H.; Kambiranda, D.M.; Kim, J.M.; Islam, S.M.A.; Yun, M.G.; Cho, J.J.; Lim, W.J.; et al. Biotransformation of phenolics (isoflavones, flavanols and phenolic acids) during the fermentation of cheonggukjang by Bacillus pumilus HY1. Food Chem. 2009, 114, 413–419. [Google Scholar] [CrossRef]
- Cho, K.M.; Lee, J.H.; Yun, H.D.; Ahn, B.Y.; Kim, H.; Seo, W.T. Changes of phytochemical constituents (isoflavones, flavanols, and phenolic acids) during cheonggukjang soybeans fermentation using potential probiotics Bacillus subtilis CS90. J. Food Compos. Anal. 2011, 24, 402–410. [Google Scholar] [CrossRef]
- Lee, S.H.; Seo, M.H.; Oh, D.K. Deglycosylation of isoflavones in isoflavone-rich soy germ flour by Aspergillus oryzae KACC 40247. J. Agric. Food Chem. 2013, 61, 12101–12110. [Google Scholar] [CrossRef]
- Di Gioia, D.; Strahsburger, E.; Lopez de Lacey, A.M.; Bregola, V.; Marotti, I.; Aloisio, I.; Biavati, B.; Dinelli, G. Flavonoid bioconversion in Bifidobacterium pseudocatenulatum B7003: A potential probiotic strain for functional food development. J. Funct. Foods 2014, 7, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Oka, T.; Simpson, F.J.; Krishnamurty, H.G. Degradation of rutin by Aspergillus flavus. Studies on specificity, inhibition, and possible reaction mechanism of quercetinase. Can. J. Microbiol. 1972, 18, 493–508. [Google Scholar] [CrossRef]
- Lin, Y.T.; Hsiu, S.L.; Hou, Y.C.; Chen, H.Y.; Chao, P.D.L. Degradation of flavonoid aglycones by rabbit, rat and human fecal flora. Biol. Pharm. Bull. 2003, 26, 747–751. [Google Scholar] [CrossRef]
- Serra, A.; Macià, A.; Romero, M.P.; Reguant, J.; Ortega, N.; Motilva, M.J. Metabolic pathways of the colonic metabolism of flavonoids (flavonols, flavones and flavanones) and phenolic acids. Food Chem. 2012, 130, 383–393. [Google Scholar] [CrossRef]
- Huynh, T.N.; Smagghe, G.; Gonzales, G.B.; Van Camp, J.; Raes, K. Extraction and bioconversion of kaempferol metabolites from cauliflower outer leaves through fungal fermentation. Biochem. Eng. J. 2016, 116, 27–33. [Google Scholar] [CrossRef]
- Irimat, R. Fungal Bioconversion of Flavonoid Compounds in Cauliflower Waste as a Source of Bioactive Extracts. Master’s Thesis, Ghent University, Gent, Belgium, 2014. [Google Scholar]
- Taki, Y.; Ikeda, K.; Sato, C.; Yano, M.; Sato, T.; Konno, H. Production and characterization of β-glucosidase from Rhizopus oryzae MIBA348. J. Biol. Macromol. 2005, 5, 11–16. [Google Scholar]
- Yadav, G.; Singh, A.; Bhattacharya, P.; Yuvraj, J.; Banerjee, R. Comparative analysis of solid-state bioprocessing and enzymatic treatment of finger millet for mobilization of bound phenolics. Bioprocess. Biosyst. Eng. 2013, 36, 1563–1569. [Google Scholar] [CrossRef]
- Lee, I.H.; Chou, C.C. Distribution profiles of isoflavone isomers in black bean kojis prepared with various filamentous fungi. J. Agric. Food Chem. 2006, 54, 1309–1314. [Google Scholar] [CrossRef]
- Lee, I.H.; Hung, Y.H.; Chou, C.C. Solid-state fermentation with fungi to enhance the antioxidative activity, total phenolic and anthocyanin contents of black bean. Int. J. Food Microbiol. 2008, 121, 150–156. [Google Scholar] [CrossRef]
- Marotti, M.; Piccaglia, R. Characterization of flavonoids in different cultivars of onion (Allium cepa L.). J. Food Sci. 2002, 67, 1229–1232. [Google Scholar] [CrossRef]
- Rodríguez Galdón, B.; Rodríguez Rodríguez, E.M.; Díaz Romero, C. Flavonoids in onion cultivars (Allium cepa L.). J. Food Sci. 2008, 73, 599–605. [Google Scholar] [CrossRef]
- You, H.J.; Ahn, H.J.; Ji, G.E. Transformation of rutin to antiproliferative quercetin-3-glucoside by Aspergillus niger. J. Agric. Food Chem. 2010, 58, 10886–10892. [Google Scholar] [CrossRef]
- Battaglia, E.; Benoit, I.; van den Brink, J.; Wiebenga, A.; Coutinho, P.; Henrissat, B.; de Vries, R. Carbohydrate-active enzymes from the zygomycete fungus Rhizopus oryzae: A highly specialized approach to carbohydrate degradation depicted at genome level. BMC Genom. 2011, 12. [Google Scholar] [CrossRef]
- Gesell, M.; Hammer, E.; Mikolasch, A.; Schauer, F. Oxidation and ring cleavage of dibenzofuran by the filamentous fungus Paecilomyces lilacinus. Arch. Microbiol. 2004, 182, 51–59. [Google Scholar] [CrossRef]
- Schlueter, R.; Röder, A.; Czekalski, N.; Gliesche, D.; Mikolasch, A.; Schauer, F. Novel mechanisms of biotransformation of p-tert-amylphenol by bacteria and fungi with special degradation abilities and simultaneous detoxification of the disinfectant. Appl. Microbiol. Biotechnol. 2013, 98, 373–384. [Google Scholar] [CrossRef]
- Slana, M.; Žigon, D.; Makovec, T.; Lenasi, H. The response of filamentous fungus Rhizopus nigricans to flavonoids. J. Basic Microbiol. 2011, 51, 433–441. [Google Scholar] [CrossRef]
- Olsen, H.; Aaby, K.; Borge, G.I.A. Characterisation and quantificiaton of flavonoids and hydroxycinnamic acids in curly kale (Brassica oleracea L. Convar. acephala Var; sabellica) by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2009, 57, 2816–2825. [Google Scholar] [CrossRef]
- Gonzales, G.B.; Raes, K.; Coelus, S.; Struijs, K.; Smagghe, G.; Van Camp, J. Ultra(high)-pressure liquid chromatography–electrospray ionization-time-of-flight-ion mobility-high definition mass spectrometry for the rapid identification and structural characterization of flavonoid glycosides from cauliflower waste. J. Chromatogr. 2014, 1323, 39–48. [Google Scholar] [CrossRef]
Figure 1.
UPLC chromatograms of extract of cauliflower outer leaves, A, control; B, fermented with Rhizopus oryzae for 2 days; C, fermented with Rhizopus azygosporus for 4 days. The numbers indicate the following compounds: 1 = kaempferol; 2 = kaempferol-3-O-glucoside, 3 = kaempferol-3-O-diglucoside.
Figure 1.
UPLC chromatograms of extract of cauliflower outer leaves, A, control; B, fermented with Rhizopus oryzae for 2 days; C, fermented with Rhizopus azygosporus for 4 days. The numbers indicate the following compounds: 1 = kaempferol; 2 = kaempferol-3-O-glucoside, 3 = kaempferol-3-O-diglucoside.

Figure 2.
UPLC chromatograms of onion extract, A, control; B, fermented with Rhizopus oryzae for 1 day; C, fermented with Rhizopus azygosporus for 1 day. The numbers indicate the following compounds: 1 = quercetin; 2 = quercetin-3-O-glucoside, 3 = quercetin-3-O-glucoside-7-O-glucoside.
Figure 2.
UPLC chromatograms of onion extract, A, control; B, fermented with Rhizopus oryzae for 1 day; C, fermented with Rhizopus azygosporus for 1 day. The numbers indicate the following compounds: 1 = quercetin; 2 = quercetin-3-O-glucoside, 3 = quercetin-3-O-glucoside-7-O-glucoside.

Figure 3.
Accumulation of kaempferol, kaempferol-3-O-glucoside, kaempferol-3-diglucoside during 7 days of fermentation of cauliflower outer leave extracts (n = 3).
Figure 3.
Accumulation of kaempferol, kaempferol-3-O-glucoside, kaempferol-3-diglucoside during 7 days of fermentation of cauliflower outer leave extracts (n = 3).

Figure 4.
Accumulation of quercetin, and quercetin-3-O-glucoside, and quercetin-3-O-glucoside-7-O-glucoside during 7 days of fermentation of onion powder extracts (n = 3).
Figure 4.
Accumulation of quercetin, and quercetin-3-O-glucoside, and quercetin-3-O-glucoside-7-O-glucoside during 7 days of fermentation of onion powder extracts (n = 3).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification of flavonoid compounds selected from cauliflower outer leaves and onion extract.
Table 1.
Identification of flavonoid compounds selected from cauliflower outer leaves and onion extract.
| Extract | Rt (min) | m/z Values | Identity | |
|---|---|---|---|---|
| Exact Mass | Fragments | |||
| Cauliflower Outer Leaves | 18.15 | 609 | 609,284 | Kaempferol-3-O-diglucoside |
| 21.58 | 447 | 447,284 | Kaempferol-3-O-glucoside | |
| 26.91 | 285 | 285 | Kaempferol | |
| Onion | 16.57 | 625 | 625,463,300 | Quercetin-3-O-glucoside-7-O-glucoside |
| 19.42 | 463 | 463,300 | Quercetin-3-O-glucoside | |
| 24.29 | 301 | 301 | Quercetin | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huynh, N.T.; Smagghe, G.; Gonzales, G.B.; Van Camp, J.; Raes, K. Bioconversion of Kaempferol and Quercetin Glucosides from Plant Sources Using Rhizopus spp. Fermentation 2018, 4, 102. https://doi.org/10.3390/fermentation4040102
AMA Style
Huynh NT, Smagghe G, Gonzales GB, Van Camp J, Raes K. Bioconversion of Kaempferol and Quercetin Glucosides from Plant Sources Using Rhizopus spp. Fermentation. 2018; 4(4):102. https://doi.org/10.3390/fermentation4040102
Chicago/Turabian StyleHuynh, Nguyen Thai, Guy Smagghe, Gerard Bryan Gonzales, John Van Camp, and Katleen Raes. 2018. "Bioconversion of Kaempferol and Quercetin Glucosides from Plant Sources Using Rhizopus spp." Fermentation 4, no. 4: 102. https://doi.org/10.3390/fermentation4040102
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.