Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods

 and
and
Abstract
:1. Introduction
- (1)
- Fermented foods have a longer shelf life than the original foods.
- (2)
- The enhancement of organoleptic properties; for example, cheese has more enhanced organoleptic properties in terms of taste than its raw substrate viz. milk.
- (3)
- The removal of harmful/unwanted ingredients from raw materials—for example, during garri preparation, there is a reduction in the poisonous cyanide content of cassava, and the flatulence factors in soybeans are removed by fermentation.
- (4)
- The enhancement of nutritional properties due to the presence of fermenting microorganisms. For example, yeast in bread and yeast and lactic acid bacteria in garri add to its nutritive quality.
- (5)
- The fermentation process reduces the cooking time of food. For example, West African food, i.e., Ogi (prepared from fermented maize), and soybean products.
- (6)
- The fermented products consist of higher in vitro antioxidant capacity. For example, fermented milk and yogurt consist of higher antioxidant properties compared to milk, as there is a release of biopeptides that follow the proteolysis of milk proteins, particularly α-casein, α-lactalbumin, and β-lactoglobulin.
2. Enhancement of Nutritional Quality in Fermented Foods by Microorganisms
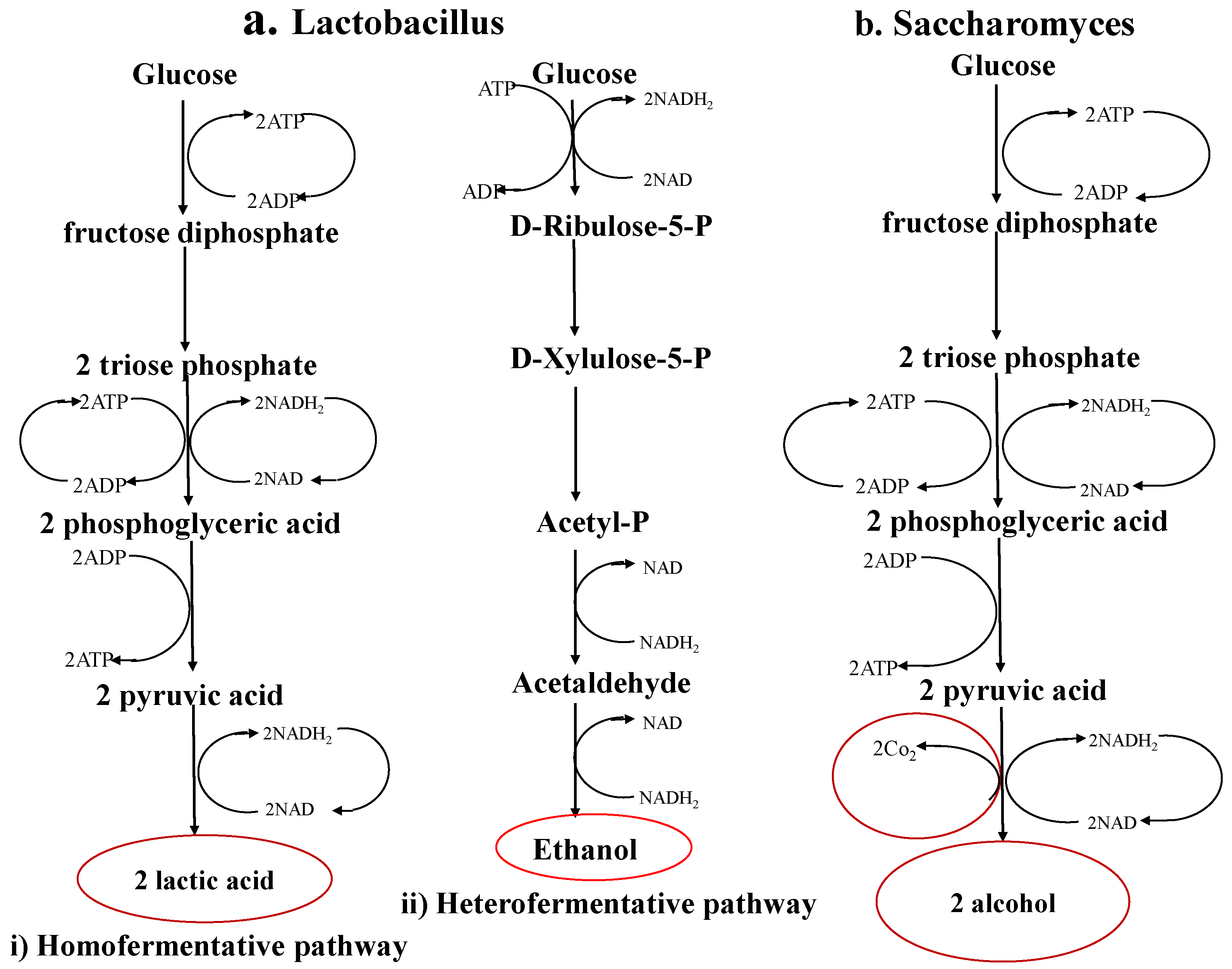
3. Effects of Lactic Acid Fermentation on the Nutritional Aspects of Food
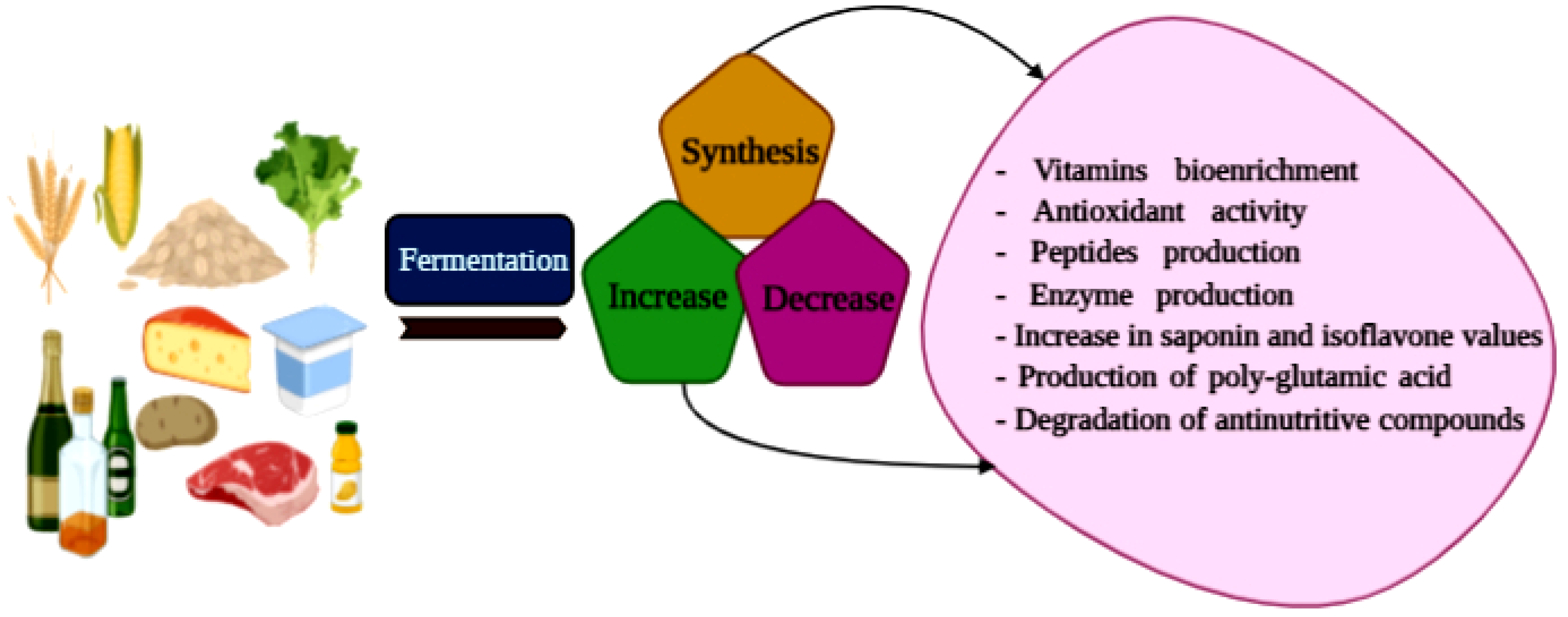
4. Enrichment and Changes of Biological Components in Fermented Foods
4.1. Vitamins Bio-Enrichment
4.2. Antioxidant Activity
4.3. Peptides Production
4.4. Enzymes Production through Microorganisms
4.5. Increase in Saponin and Isoflavone Values and Poly-Glutamic Acid Production
4.6. Anti-Nutritive Compounds Degradation
4.7. Biochemical Changes during Cereal Fermentation
4.8. Presence of Biogenic Amines in Juices and Vegetables Fermented with Lactic Acid Bacteria
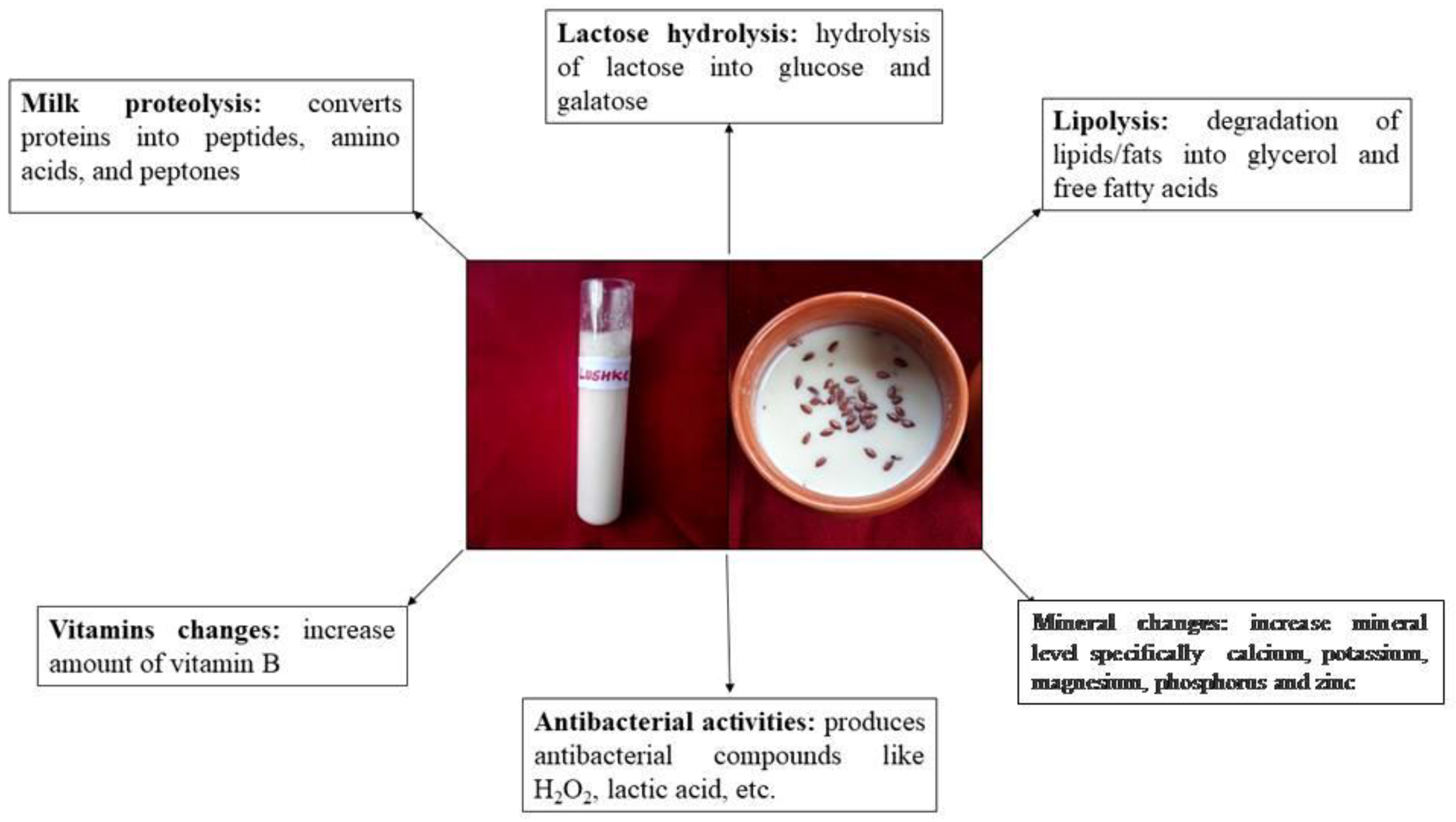
5. Nutritional Value of Fermented Dairy Products
- (a)
- Milk proteolysis
- (b)
- Lactose hydrolysis
- (c)
- Lipolysis
- (d)
- Vitamins changes
- (e)
- Antibacterial activity
- (f)
- Mineral changes
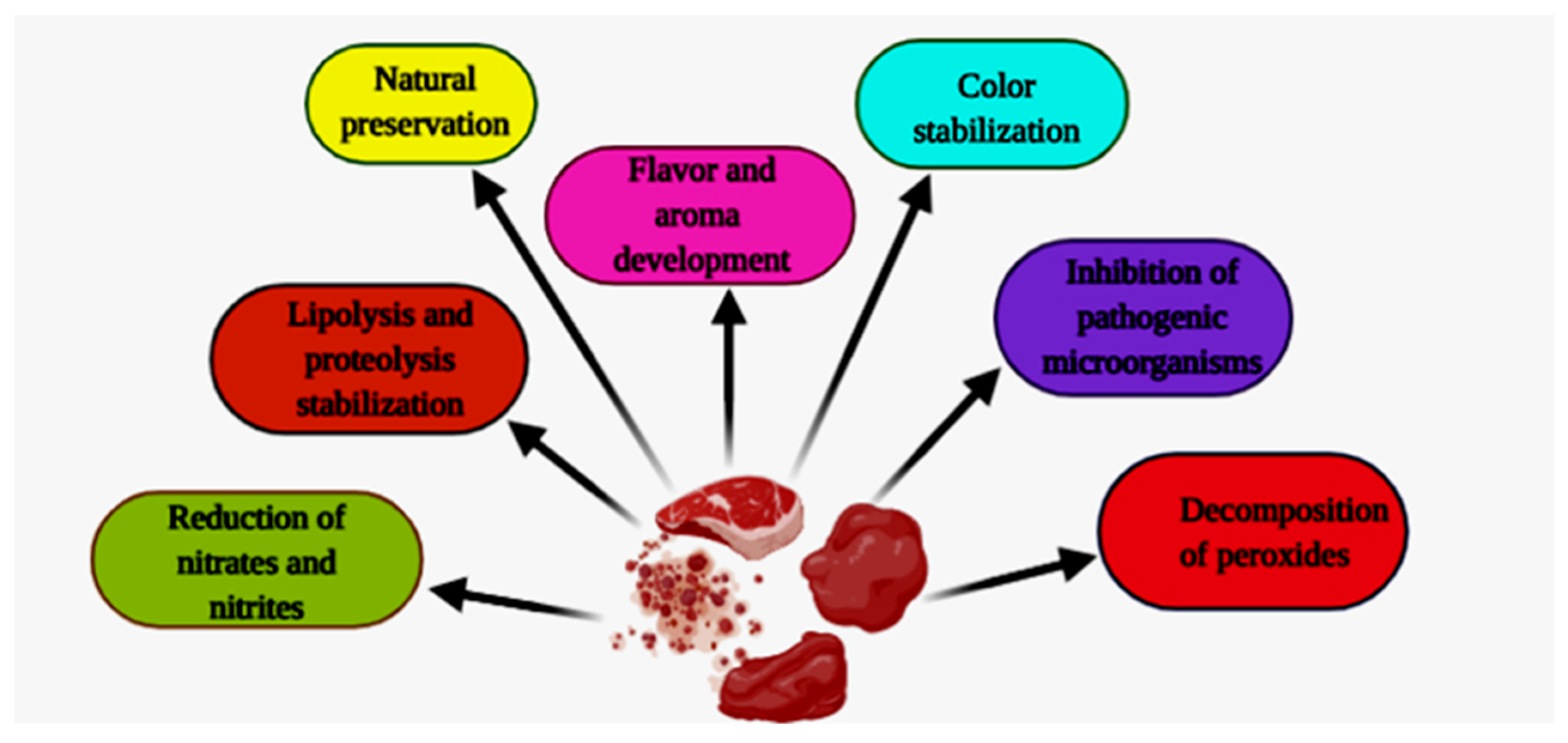
6. Biochemical Changes in Meat-Based Fermented Food Products
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, H.; Sun-Waterhouse, D.; Waterhouse, G.I.; Cui, C.; Ruan, Z. Fermentation-enabled wellness foods: A fresh perspective. Food Sci. Hum. Well. 2019, 8, 203–243. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V.; Luziatelli, F.; Ficca, A.G.; Ruzzi, M. Health-promoting components in fermented foods: An up-to-date systematic review. Nutrients 2019, 11, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanlier, N.; Gokcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Okafor, N. Fermented foods and their processing. In Biotechnology-Volume VIII: Fundamentals in Biotechnology; Eolss Publishers: Oxford, UK, 2009; Volume 8, p. 19. [Google Scholar]
- Mokoena, M.P. Lactic acid bacteria and their bacteriocins: Classification, biosynthesis, and applications against uropathogens: A mini-review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef]
- Anukam, K.C.; Reid, G. African traditional fermented foods and probiotics. J. Med. Food 2009, 12, 1177–1184. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Vagelas, I.; Gougoulias, N.; Nedesca, E.D.; Liviu, G. Bread contamination with fungus. Carpath. J. Food Sci. Technol. 2011, 3, 1–6. [Google Scholar]
- Doyle, M.P.; Meng, J. Bacteria in food and beverage production. In Prokaryotes; Springer: New York, NY, USA, 2006; pp. 797–811. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: Lights and shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, T.; Morley, A. An evolutionary perspective on the Crabtree effect. Front. Mol. Biosci. 2014, 1, 17. [Google Scholar] [CrossRef]
- Hasan, M.N.; Sultan, M.Z.; Mar-E-Um, M. Significance of fermented food in nutrition and food science. J. Sci. Res. 2014, 6, 373–386. [Google Scholar] [CrossRef]
- Kennedy, D.O. B vitamins and the brain: Mechanisms, dose, and efficacy—A review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, N.N.; Hotchkiss, J.H. Food Science, 5th ed.; C.B.S. Publishers and Distributors: New Delhi, India, 2006; pp. 264–277. [Google Scholar]
- Mokoena, M.P.; Chelule, P.K.; Gqaleni, N. Reduction of fumonisin B1 and zearalenone by lactic acid bacteria in fermented maize meal. J. Food Prot. 2005, 68, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Rollan, G.C.; Gerez, C.L.; LeBlanc, J.G. Lactic fermentation as a strategy to improve the nutritional and functional values of pseudocereals. Front. Nutr. 2019, 6, 98. [Google Scholar] [CrossRef] [Green Version]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 1–14. [Google Scholar] [CrossRef]
- Hill, D.; Sugrue, I.; Arendt, E.; Hill, C.; Stanton, C.; Ross, R.P. Recent advances in microbial fermentation for dairy and health. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gangoliya, S.S.; Singh, N.K. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 2015, 52, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Jesch, E.D.; Carr, T.P. Food ingredients that inhibit cholesterol absorption. Prev. Nutr. Food Sci. 2017, 22, 67. [Google Scholar] [CrossRef]
- Minh, N.G. Investigation of pickled water spinach (Ipomoea aquatic) fermentation by Lactobacillus sp. Int. J. Multidiscip. Res. Dev. 2014, 1, 71–80. [Google Scholar]
- Chadare, F.J.; Idohou, R.; Nago, E.; Affonfere, M.; Agossadou, J.; Fassinou, T.K.; Kenou, C.; Honfo, S.; Azokpota, P.; Linnemann, A.R.; et al. Conventional and food-to-food fortification: An appraisal of past practices and lessons learned. Food Sci. Nutr. 2019, 7, 2781–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, K.D.; Gupta, M. Vitamin B1 thiamine deficiency (Beriberi). In StatPearls [Internet]; Stat Pearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Barennes, H.; Sengkhamyong, K.; Rene, J.P.; Phimmasane, M. Beriberi (thiamine deficiency) and high infant mortality in northern Laos. PLoS Negl. Trop. Dis. 2015, 9, e0003581. [Google Scholar] [CrossRef] [Green Version]
- Law, S.V.; Abu Bakar, F.; Mat Hashim, D.; Abdul Hamid, A. Popular fermented foods and beverages in Southeast Asia. Int. Food Res. J. 2011, 18, 475–484. [Google Scholar]
- Nout, M.J.R.; Kiers, J.L. Tempe fermentation, innovation, and functionality: Update into the third millenium. J. Appl. Microbiol. 2005, 98, 789–805. [Google Scholar] [CrossRef]
- Ghosh, D.; Chattopadhyay, P. Preparation of idli batter, its properties, and nutritional improvement during fermentation. J. Food Sci. Technol. 2011, 48, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalante, A.; Lopez Soto, D.R.; Velazquez Gutierrez, J.E.; Giles-Gomez, M.; Bolivar, F.; Lopez-Munguia, A. Pulque, a traditional Mexican alcoholic fermented beverage: Historical, microbiological, and technical aspects. Front. Microbiol. 2016, 7, 1026. [Google Scholar] [CrossRef] [Green Version]
- Escalante, A.; Giles-Gomez, M.; Flores, G.E.; Acuña, V.M.; Moreno-Terrazas, R.; Lopez-Munguia, A.; Lappe-Oliveras, P. Pulque fermentation. In Handbook of Plant-Based Fermented Food and Beverage Technology; CRC Press: Boca Raton, FL, USA, 2012; pp. 691–706. [Google Scholar]
- Tamang, J.P.; Cotter, P.D.; Endo, A.; Han, N.S.; Kort, R.; Liu, S.Q.; Mayo, B.; Westerik, N.; Hutkins, R. Fermented foods in a global age: East meets West. Compr. Rev. Food Sci. Food Saf. 2020, 19, 184–217. [Google Scholar] [CrossRef] [Green Version]
- Aka, S.; Konan, G.; Fokou, G.; Dje, K.M.; Bonfoh, B. Review of African traditional cereal beverages. Am. J. Res. Commun. 2014, 2, 103–153. [Google Scholar]
- Amoa-Awua, W.K.; Sampson, E.; Tano-Debrah, K. Growth of yeasts, lactic, and acetic acid bacteria in palm wine during tapping and fermentation from felled oil palm (Elaeis guineensis) in Ghana. J. Appl. Microbiol. 2007, 102, 599–606. [Google Scholar] [CrossRef]
- Karamoko, D.; Moroh, J.L.A.; Bouatenin, K.M.J.P.; Dje, K.M. Biochemical and microbial properties of palm wine: Effect of tapping length and varietal differences. Food Nutr. Sci. 2016, 7, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Anal, A.K. Quality ingredients and safety concerns for traditional fermented foods and beverages from Asia: A review. Fermentation 2019, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.F.; Pan, T.M. In vitro effects of lactic acid bacteria on cancer cell viability and antioxidant activity. J. Food Drug Anal. 2010, 18, 77–86. [Google Scholar]
- Abubakar, M.A.S.; Hassan, Z.; Imdakim, M.M.A.; Sharifah, N.R.S.A. Antioxidant activity of lactic acid bacteria (L.A.B.) fermented skim milk as determined by 1,1-diphenyl-2-picrylhydrazyl (D.P.P.H.) and ferrous chelating activity (F.C.A.). Afr. J. Microbiol. Res. 2012, 6, 6358–6364. [Google Scholar] [CrossRef]
- Chettri, R.; Tamang, J.P. Functional properties of Tungrymbai and Bekang, naturally fermented soybean foods of North East India. Int. J. Fermented Foods 2014, 3, 87–103. [Google Scholar] [CrossRef]
- Moktan, B.; Saha, J.; Sarkar, P.K. Antioxidant activities of soybean as affected by Bacillus-fermentation to kinema. Food Res. Int. 2008, 41, 586–593. [Google Scholar] [CrossRef]
- Tamang, J.P. Naturally fermented ethnic soybean foods of India. J. Ethn. Foods 2015, 2, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Astuti, M.N.; Suparmo, M.; Soesatyo, H.N.E. The role of black soybean tempe in increasing antioxidant enzyme activity and human lymphocyte proliferation in vivo. Int. J. Curr. Microbiol. Appl. Sci. 2013, 2, 316–327. [Google Scholar]
- Shon, M.Y.; Lee, J.; Choi, J.H.; Choi, S.Y.; Nam, S.H.; Seo, K.I.; Lee, S.W.; Sung, N.J.; Park, S.K. Antioxidant and free radical scavenging activity of methanol extract of chungkukjang. J. Food Compost. Anal. 2007, 20, 113–118. [Google Scholar] [CrossRef]
- Shin, D.; Jeong, D. Korean traditional fermented soybean products: Jang. J. Ethn. Foods 2015, 2, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Dajanta, K.; Janpum, P.; Leksing, W. Antioxidant capacities, total phenolics, and flavonoids in black and yellow soybeans fermented by Bacillus subtilis: A comparative study of Thai fermented soybeans (thuanao). Int. Food Res. J. 2013, 20, 3125. [Google Scholar] [CrossRef] [Green Version]
- Ping, P.; Shih, C.; Rong, T.; King, Q. Effect of isoflavone aglycone content and antioxidation activity in natto by various cultures of Bacillus subtilis during the fermentation period. J. Nutr. Food Sci. 2012, 2, 153. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.J.; Li, D.; Zou, L.; Dong Chen, X.; Cheng, Y.Q.; Yamaki, K.; Li, L.T. Antioxidative activity of douchi (a Chinese traditional salt-fermented soybean food) extracts during its processing. Int. J. Food Prop. 2007, 10, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Farvin, K.S.; Baron, C.P.; Nielsen, N.S.; Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 1-in vitro assays and evaluation in ω-3 enriched milk. Food Chem. 2010, 123, 1081–1089. [Google Scholar] [CrossRef]
- Park, J.M.; Shin, J.H.; Gu, J.G.; Yoon, S.J.; Song, J.C.; Jeon, W.M.; Suh, H.J.; Chang, U.J.; Yang, C.Y.; Kim, J.M. Effect of antioxidant activity in kimchi during a short-term and over-ripening fermentation period. J. Biosci. Bioeng. 2011, 112, 356–359. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, P.; Lou, L. Antioxidant activities of lactic acid bacteria for quality improvement of fermented sausage: Antioxidant activities of lactic acid bacteria. J. Food Sci. 2017, 82, 2960–2967. [Google Scholar] [CrossRef]
- De Mejia, E.G.; Dia, V.P. The role of nutraceutical proteins and peptides in apoptosis, angiogenesis, and metastasis of cancer cells. Cancer Metast. Rev. 2010, 29, 511–528. [Google Scholar] [CrossRef]
- Nagai, T.; Tamang, J.P. Fermented Soybeans Non-soybeans legumes foods. In Fermented Foods and Beverages of the World; Tamang, J.P., Kailasapathy, K., Eds.; CRC Press: New York, NY, USA, 2010; pp. 191–224. [Google Scholar]
- Phelan, M.; Kerins, D. The potential role of milk-derived peptides in cardiovascular disease. Food Funct. 2011, 2, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.C.; Liu, F.; Ren, D.X.; Han, W.W.; Du, Y.O. Effect of culturing conditions on the expression of key enzymes in the proteolytic system of Lactobacillus bulgaricus. J. Zhejiang Univ. Sci. 2015, 16, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Alcantara, C.; Bauerl, C.; Revilla-Guarinos, A.; Perez-Martinez, G.; Monedero, V.; Zuniga, M. Peptide and amino acid metabolism is controlled by an OmpR-family response regulator in Lactobacillus casei. Mol. Microbiol. 2016, 100, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Vukotic, G.; Strahinic, I.; Begovic, J.; Lukic, J.; Kojic, M.; Fira, D. Survey on proteolytic activity and diversity of proteinase genes in mesophilic lactobacilli. Microbiology 2016, 85, 33–41. [Google Scholar] [CrossRef]
- Griffiths, M.W.; Tellez, A.M. Lactobacillus helveticus: The proteolytic system. Front. Microbiol. 2013, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, M.; Ueno, H.M.; Watanabe, M.; Tatsuma, Y.; Seto, Y.; Miyamoto, T. Distinctive proteolytic activity of cell envelope proteinase of Lactobacillus helveticus isolated from airag, a traditional Mongolian fermented mare’s milk. Int. J. Food Microbiol. 2015, 197, 65–71. [Google Scholar] [CrossRef]
- Stefanovic, E.; Fitzgerald, G.; McAuliffe, O. Advances in the genomics and metabolomics of dairy lactobacilli: A review. Food Microbiol. 2017, 61, 33–49. [Google Scholar] [CrossRef]
- Singh, T.A.; Devi, K.R.; Ahmed, G.; Jeyaram, K. Microbial and endogenous origin of fibrinolytic activity in traditional fermented foods of Northeast India. Food Res. Int. 2014, 55, 356–362. [Google Scholar] [CrossRef]
- Qian, B.; Xing, M.; Cui, L.; Deng, Y.; Xu, Y.; Huang, M.; Zhang, S. Antioxidant, antihypertensive, and immunomodulatory activities of peptide fractions from fermented skim milk with Lactobacillus delbrueckii subsp. bulgaricus LB340. J. Dairy Res. 2011, 78, 72. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Z.; Chen, X.; Liu, Y.; Zhang, H.; Sun, T. Identification of angiotensin I-converting enzyme inhibitory peptides from koumiss, a traditional fermented mare’s milk. J. Dairy Sci. 2010, 93, 884–892. [Google Scholar] [CrossRef] [Green Version]
- Quiros, A.; Hernández-Ledesma, B.; Ramos, M.; Amigo, L.; Recio, I. Angiotensin-converting enzyme inhibitory activity of peptides derived from caprine kefir. J. Dairy Sci. 2005, 88, 3480–3487. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou, K.; Alegria, A.; Bron, P.A.; De Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Res. 2016, 80, 837–890. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.; Butikofer, U.; Walther, B.; Wechsler, D.; Sieber, R. Hot topic: Changes in angiotensin-converting enzyme inhibition and concentrations of the tripeptides Val-Pro-Pro and Ile-Pro-Pro during ripening of different Swiss cheese varieties. J. Dairy Sci. 2009, 92, 826–836. [Google Scholar] [CrossRef] [Green Version]
- Moslehishad, M.; Ehsani, M.R.; Salami, M.; Mirdamadi, S.; Ezzatpanah, H.; Naslaji, A.N.; Moosavi-Movahedi, A.A. The comparative assessment of ACE-inhibitory and antioxidant activities of peptide fractions obtained from fermented camel and bovine milk by Lactobacillus rhamnosus PTCC 1637. Int. Dairy J. 2013, 29, 82–87. [Google Scholar] [CrossRef]
- Ichimura, T.; Hu, J.; Aita, D.Q.; Maruyama, S. Angiotensin I-converting enzyme inhibitory activity and insulin secretion stimulative activity of fermented fish sauce. J. Biosci. Bioeng. 2003, 96, 496–499. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Nout, M.J.R.; Aidoo, K.E. Asian fungal fermented food. In Industrial Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 23–47. [Google Scholar] [CrossRef]
- Suganuma, T.; Fujita, K.; Kitahara, K. Some distinguishable properties between acid-stable and neutral types of α-amylases from acid-producing koji. J. Biosci. Bioeng. 2007, 104, 353–362. [Google Scholar] [CrossRef]
- Tamang, J.P.; Dewan, S.; Tamang, B.; Rai, A.; Schillinger, U.; Holzapfel, W.H. Lactic acid bacteria in hamei and marcha of North East India. Indian J. Microbiol. 2007, 47, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Tsuyoshi, N.; Fudou, R.; Yamanaka, S.; Kozaki, M.; Tamang, N.; Thapa, S.; Tamang, J.P. Identification of yeast strains isolated from marcha in Sikkim, a microbial starter for amylolytic fermentation. Int. J. Food Microbiol. 2005, 99, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Wong, A.H.K.; Jiang, B. Fibrinolytic enzymes in Asian traditional fermented foods. Food Res. Int. 2005, 38, 243–250. [Google Scholar] [CrossRef]
- Kotb, E. Fibrinolytic bacterial enzymes with thrombolytic activity. In Fibrinolytic Bacterial Enzymes with Thrombolytic Activity; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–74. [Google Scholar] [CrossRef]
- Chang, C.T.; Wang, P.M.; Hung, Y.F.; Chung, Y.C. Purification and biochemical properties of a fibrinolytic enzyme from Bacillus subtilis-fermented red bean. Food Chem. 2012, 133, 1611–1617. [Google Scholar] [CrossRef]
- Zeng, W.; Li, W.; Shu, L.; Yi, J.; Chen, G.; Liang, Z. Non-sterilized fermentative co-production of poly (γ-glutamic acid) and fibrinolytic enzyme by a thermophilic Bacillus subtilis GXA-28. Bioresour. Technol. 2013, 142, 697–700. [Google Scholar] [CrossRef]
- Montriwong, A.; Rodtong, S.; Yongsawatdigul, J. Detergent-stable salt-activated proteinases from Virgibacillus halodenitrificans SK1-3-7 isolated from fish sauce fermentation. Appl. Biochem. Biotechnol. 2015, 176, 505–517. [Google Scholar] [CrossRef]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect, and bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Chiou, R.Y.Y.; Cheng, S.L. Isoflavone transformation during soybean koji preparation and subsequent miso fermentation supplemented with ethanol and NaCl. J. Agric. Food Chem. 2001, 49, 3656–3660. [Google Scholar] [CrossRef]
- Wang, L.J.; Yin, L.J.; Li, D.; Zou, L.; Saito, M.; Tatsumi, E. Influences of processing and NaCl supplementation on isoflavone contents and composition during douchi manufacturing. Food Chem. 2007, 101, 1247–1253. [Google Scholar] [CrossRef]
- Lee, Y.W.; Kim, J.D.; Zheng, J.; Row, K.H. Comparisons of isoflavones from Korean and Chinese soybean and processed products. Biochem. Eng. J. 2007, 36, 49–53. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, W.; Shan, Y.; Zhi-Qiang, E.; Wang, L.Q. Study on the inhibition of fermented soybean to cancer cells. J. Northeast. Agric. Univ. (Engl. Ed.) 2009, 16, 25–28. [Google Scholar]
- Nakajima, N.; Nozaki, N.; Ishihara, K.; Ishikawa, A.; Tsuji, H. Analysis of isoflavone content in tempeh, a fermented soybean, and preparation of new isoflavone-enriched tempeh. J. Biosci. Bioeng. 2005, 100, 685–687. [Google Scholar] [CrossRef]
- Kwak, C.S.; Park, S.C.; Song, K.Y. Doenjang, a fermented soybean paste, decreased visceral fat accumulation and adipocyte size in rats fed with a high-fat diet more effectively than non-fermented soybeans. J. Med. Food 2012, 15, 1–9. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Amaya-Farfan, J.; Berhow, M.A.; Mandarino, J.M.G.; de Mejia, E.G.; Chang, Y.K. A high-protein soybean cultivar contains lower isoflavones and saponins but higher minerals and bioactive peptides than a low-protein cultivar. Food Chem. 2010, 120, 15–21. [Google Scholar] [CrossRef]
- Ellington, A.A.; Berhow, M.A.; Singletary, K.W. Inhibition of Akt signaling and enhanced ERK1/2 activity are involved in the induction of macroautophagy by triterpenoid B-group soyasaponins in colon cancer cells. Carcinogenesis 2006, 27, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Tanizawa, H. Effects of soyasaponins on lipid peroxidation through the secretion of thyroid hormones. Biol. Pharm. Bull. 2006, 29, 1759–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Hodai, T.; Yamamoto, H.; Matsumori, N.; Oishi, T. Membrane interaction of soyasaponins in association with their antioxidation effect-Analysis of biomembrane interaction. Soy Prot. Res. 2006, 9, 82–86. [Google Scholar]
- Yanagisawa, Y.; Sumi, H. Natto Bacillus contains a large amount of water-soluble vitamin K (menaquinone-7). J. Food Biochem. 2005, 29, 267–277. [Google Scholar] [CrossRef]
- Omizu, Y.; Tsukaoto, C.; Chettri, R.; Tamang, J.P. Determination of saponin contents in raw soybean and fermented soybean foods of India. J. Sci. Ind. Res. India 2011, 70, 533–538. [Google Scholar]
- Meerak, J.; Iida, H.; Watanabe, Y.; Miyashita, M.; Sato, H.; Nakagawa, Y.; Tahara, Y. Phylogeny of γ-polyglutamic acid-producing Bacillus strains isolated from fermented soybean foods manufactured in Asian countries. J. Gen. Appl. Microbiol. 2007, 53, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Nishito, Y.; Osana, Y.; Hachiya, T.; Popendorf, K.; Toyoda, A.; Fujiyama, A.; Itaya, M.; Sakakibara, Y. A whole-genome assembly of a natto production strain Bacillus subtilis from very short read data. BMC Genom. 2010, 11, 243. [Google Scholar] [CrossRef] [Green Version]
- Stanley-Wall, N.; Lazazzera, B.A. Defining the genetic differences between wild and domestic strains of Bacillus subtilis that affect poly--dl-glutamic acid production and biofilm formation. Mol. Microbiol. 2005, 57, 1143–1158. [Google Scholar] [CrossRef]
- Oppermann-Sanio, F.B.; Steinbüchel, A. Occurrence, functions and biosynthesis of polyamides in microorganisms and biotechnological production. Naturwissenschaften 2002, 89, 11–22. [Google Scholar] [CrossRef]
- Yoon, S.H.; Do, J.H.; Lee, S.Y.; Chang, H.N. Production of poly-γ-glutamic acid by fed-batch culture of Bacillus licheniformis. Biotechnol. Lett. 2000, 22, 585–588. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Omolara, B.O. Cyanide content of commercial gari from different areas of Ekiti State, Nigeria. World J. Nutr. Health 2014, 2, 58–60. [Google Scholar] [CrossRef]
- Lambri, M.; Fumi, M.D.; Roda, A.; De Faveri, D.M. Improved processing methods to reduce the total cyanide content of cassava roots from Burundi. Afr. J. Biotechnol. 2013, 12. [Google Scholar] [CrossRef]
- Bamidele, O.P.; Fasogbon, M.B.; Oladiran, D.A.; Akande, E.O. Nutritional composition of fufu analog flour produced from Cassava root (Manihot esculenta) and Cocoyam (Colocasia esculenta) tuber. Food Sci. Nutr. 2015, 3, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, P.C. Philippine Fermented Foods: Principles and Technology; UP Press: Quezon City, Philippines, 2008. [Google Scholar]
- Laskowski, W.; Gorska-Warsewicz, H.; Rejman, K.; Czeczotko, M.; Zwolinska, J. How important are cereals and cereal products in the average polish diet? Nutrients 2019, 11, 679. [Google Scholar] [CrossRef] [Green Version]
- Verni, M.; Rizzello, C.G.; Coda, R. Fermentation biotechnology applied to cereal industry by-products: Nutritional and functional insights. Front. Nutr. 2019, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Erkmen, O.; Bozoglu, T.F. Fermented cereal and grain products. In Food Microbiology: Principles into Practice; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar] [CrossRef]
- Singh, A.K.; Rehal, J.; Kaur, A.; Jyot, G. Enhancement of attributes of cereals by germination and fermentation: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1575–1589. [Google Scholar] [CrossRef]
- Mariod, A.A.; Idris, Y.M.; Osman, N.M.; Mohamed, M.A.; Sukrab, A.M.; Farag, M.Y.; Matthaus, B. Three Sudanese sorghum-based fermented foods (kisra, hulu-mur, and abreh): Comparison of proximate, nutritional value, microbiological load, and acrylamide content. Ukr. J. Food Sci. 2016, 4, 216–228. [Google Scholar] [CrossRef]
- Walker, G.M.; Stewart, G.G. Saccharomyces cerevisiae in the production of fermented beverages. Beverages 2016, 2, 30. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Liptakova, D.; Matejcekova, Z.; Valik, L. Lactic acid bacteria and fermentation of cereals and pseudocereals. Fermen. Proc. 2017, 10, 65459. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, M.; Collins, M.D. Lactic acid bacteria and human clinical infection. J. Appl. Bacteriol. 1993, 75, 95–107. [Google Scholar] [CrossRef]
- Garneau, J.E.; Moineau, S. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb. Cell Factories 2011, 10, S20, BioMed Central. [Google Scholar] [CrossRef] [Green Version]
- Aragon, F.; Perdigon, G.; de LeBlanc, A.D.M. Modification in the diet can induce beneficial effects against breast cancer. World J. Clin. Oncol. 2014, 5, 455. [Google Scholar] [CrossRef]
- Hagan, N.D.; Tabe, L.M.; Molvig, L.; Higgins, T.J.V. Modifying the amino acid composition of grains using gene technology. In Plant Biotechnology 2002 and Beyond; Springer: Dordrecht, The Netherlands, 2003; pp. 305–308. [Google Scholar] [CrossRef]
- Biji, K.B.; Ravishankar, C.N.; Venkateswarlu, R.; Mohan, C.O.; Gopal, T.S. Biogenic amines in seafood: A review. Int. J. Food Sci. Technol. 2016, 53, 2210–2218. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Durak-Dados, A.; Michalski, M.; Osek, J. Histamine and other biogenic amines in food. J. Vet. Res. 2020, 64, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Penas, E.; Martinez-Villaluenga, C.; Frias, J. Sauerkraut: Production, composition, and health benefits. In Fermented Foods in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2017; pp. 557–576. [Google Scholar] [CrossRef]
- Kalac, P.; Spicka, J.; Krizek, M.; Pelikanova, T. The effects of lactic acid bacteria inoculants on biogenic amines formation in sauerkraut. Food Chem. 2000, 70, 355–359. [Google Scholar] [CrossRef]
- Karovicova, J.; Kohajdova, Z. Lactic acid fermented vegetable juices. HortSci (Pargue) 2003, 30, 152–158. [Google Scholar]
- Kalac, P.; Spicka, J.; Krizek, M.; Pelikanova, T. Changes in biogenic amine concentrations during sauerkraut storage. Food Chem. 2000, 69, 309–314. [Google Scholar] [CrossRef]
- Kirschbaum, J.; Rebscher, K.; Brückner, H. Liquid chromatographic determination of biogenic amines in fermented foods after derivatization with 3, 5-dinitrobenzoyl chloride. J. Chromatogr. A 2000, 881, 517–530. [Google Scholar] [CrossRef]
- Hornero-Mendez, D.; Garrido-Fernandez, A. Rapid high-performance liquid chromatography analysis of biogenic amines in fermented vegetable brines. J. Food Prot. 1997, 60, 414–419. [Google Scholar] [CrossRef]
- Kolesarova, E. Vyskyt a vznik biogennych aminov v potravinach. Bull. PV 1995, 34, 109–122. [Google Scholar]
- Mehta, B.M. Chemical composition of milk and milk products. In Handbook of Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2015; pp. 511–553. [Google Scholar]
- Shiby, V.K.; Mishra, H.N. Fermented milk and milk products as functional foods—A review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef]
- Hao, W.; Tian, P.; Zheng, M.; Wang, H.; Xu, C. Characteristics of proteolytic microorganisms and their effects on proteolysis in total mixed ration silages of soybean curd residue. Asian Austral. J. Anim. 2020, 33, 100. [Google Scholar] [CrossRef] [Green Version]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Dutra Rosolen, M.; Gennari, A.; Volpato, G.; Volken de Souza, C.F. Lactose hydrolysis in milk and dairy whey using microbial β-galactosidases. Enzym. Res. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Ye, A.; Cui, J.; Dalgleish, D.; Singh, H. Effect of homogenization and heat treatment on the behavior of protein and fat globules during gastric digestion of milk. J. Dairy Sci. 2017, 100, 36–47. [Google Scholar] [CrossRef]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of dietary and microbial vitamin B family in the regulation of host immunity. Front. Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Burgos, M.; Moreno-Fernandez, J.; Alferez, M.J.; Diaz-Castro, J.; Lopez-Aliaga, I. New perspectives in fermented dairy products and their health relevance. J. Funct. Foods 2020, 72, 104059. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665. [Google Scholar] [CrossRef] [PubMed]
- Mastanjevic, K.; Kovacevic, D.; Frece, J.; Markov, K.; Pleadin, J. The effect of autochthonous starter culture, sugars, and temperature on the fermentation of Slavonian Kulen. Food Technol. Biotechnol. 2017, 55, 67–76. [Google Scholar] [CrossRef]
- Papagianni, M. Metabolic engineering of lactic acid bacteria for the production of industrially important compounds. Comput. Struct. Biotechnol. 2012, 3, e201210003. [Google Scholar] [CrossRef] [Green Version]
- Dalcanton, F.; Carrasco, E.; Perez-Rodríguez, F.; Posada-Izquierdo, G.D.; Falcao de Aragao, G.M.; Garcia-Gimeno, R.M. Modeling the combined effects of temperature, pH, and sodium chloride and sodium lactate concentrations on the growth rate of Lactobacillus plantarum ATCC 8014. J. Food Qual. 2018. [Google Scholar] [CrossRef] [Green Version]
- Juturu, V.; Wu, J.C. Microbial production of lactic acid: The latest development. Crit. Rev. Biotechnol. 2016, 36, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Fadda, S.; Vignolo, G.; Oliver, G. Meat protein degradation by tissue and lactic acid bacteria enzymes. In Food Microbiology Protocols; Humana Press: Totova, NJ, USA, 2001; pp. 147–162. [Google Scholar] [CrossRef]
- Toldrá, F. The role of muscle enzymes in dry-cured meat products with different drying conditions. Trends Food Sci. Technol. 2006, 17, 164–168. [Google Scholar] [CrossRef]
- Pasini, F.; Soglia, F.; Petracci, M.; Caboni, M.F.; Marziali, S.; Montanari, C.; Gardini, F.; Grazia, L.; Tabanelli, G. Effect of fermentation with different lactic acid bacteria starter cultures on biogenic amine content and ripening patterns in dry fermented sausages. Nutrients 2018, 10, 1497. [Google Scholar] [CrossRef] [Green Version]
- Karslioglu, B.; Cicek, U.; Kolsarici, N.; Candogan, K. Lipolytic changes in fermented sausages produced with Turkey meat: Effects of starter culture and heat treatment. Korean J. Food Sci. Anim. Resour. 2014, 34, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Gianelli, M.P.; Salazar, V.; Mojica, L.; Friz, M. Volatile compounds present in traditional meat products (charqui and longaiza sausage) in Chile. Braz. Arch. Biol. Technol. 2012, 55, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Kosowska, M.; Majcher, M.A.; Fortuna, T. Volatile compounds in meat and meat products. Food Sci. Technol. 2017, 37, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Erginkaya, Z. Determination of organic acids in fermented sausage. Gida 1993, 18, 373–376. [Google Scholar]
- Kasankala, L.M.; Xiong, Y.L.; Chen, J. Enzymatic activity and flavor compound production in fermented silver carp fish paste inoculated with a douchi starter culture. J. Agric. Food Chem. 2012, 60, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, D.; Raemaekers, M.; Rizzo, A.; Holck, A.; De Smedt, A.; Ten Brink, B.; Hagen, B.; Montel, C.; Zanardi, E.; Murbrekk, E.; et al. Control of bioflavour and safety in fermented sausages: First results of a European project. Food Res. Int. 2000, 33, 171–180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermented Foods/Beverages | Substrates Used | Microorganisms Involved in Fermentation |
|---|---|---|
| Dairy products | Milk and milk casein | Lactobacillus bulgaricus, Lactococcus lactis, L. acidophilus, L. cremoris, L. casei, L. paracasei, L. thermophilus, L. kefiri, L. caucasicus, Penicillium camemberti, P. roqueforti, Acetobacter lovaniensis, Kluyveromyces lactis, Saccharomyces cerevisiae |
| Curd, Yogurt, Cheese, Yakult, Kefir | ||
| Vegetable products | Soybean, cabbage, ginger, cucumber, broccoli, radish | Leuconostoc mesenteroides, Aspergillus sp., Rhizopus oligosporus, R. oryzae, L. sakei, L. plantarum, Thermotoga sp., L. hokkaidonensis, L. rhamnosus, Rhodotorula rubra, Leuconostoc carnosum, Bifidobacterium dentium, Enterococcus faecalis, Weissella confusa, Candida sake |
| Kimchi, Tempeh, Natto, Miso, Sauerkraut | ||
| Cereals | Wheat, maize, sorghum, millet, rice | L. pantheris, L. plantarum, Penicillium sp., S. cerevisiae, L. mesenteroides, E. faecalis, Trichosporon pullulans, Pediococcus acidilactici, P. cerevisiae, Delbrueckii hansenii, Deb. tamari |
| Bahtura, Ambali, Chilra, Dosa, Kunu-Zaki, Marchu | ||
| Beverages | Grapes, rice, cereals | Aspergillus oryzae, Zygosaccharomyces bailii, S. cerevisiae, Acetobacter pasteurianus, Gluconacetobacter, Acetobacter xylinus, Komagataeibacter xylinus |
| Wine, Beer, Kombucha, Sake | ||
| Meat Products | Meat | L. sakei, L. curvatus, L. plantarum, Leuconostoc carnosum, Leuconostoc gelidium, B. licheniformis, E. faecalis, E. hirae, E. durans, Bacillus subtilis, L. divergens, L. carnis, E. cecorum, B. lentus |
| Sucuk, Salami, Arjia, Jama, Nham |
| Substrates | Enzymes | Microbial Source | Enzymatic Action/Process |
|---|---|---|---|
| Dairy | Protease | A. niger, A. orzyae and B. subtilis | Cheese production Removing H2O2 Lactose-free milk |
| Catalase | S. boydii and Bacillus sp. | ||
| Lactase | B. subtilis | ||
| Cereals | Amylase | B. licheniformis and B. subtilis | Malting, mashing, liquefaction, and production of flavor esters |
| Protease | A. niger | ||
| Pentosanase | Trichoderma sp. | ||
| Glucose oxidase | P. notatum | ||
| Phytase | A. niger | ||
| Pullulanase | B. acidopullulyticus | ||
| Xylanase | A. oryzae and B. subtilis | ||
| Lipases | Aspergillus niger | ||
| Β-glucanase | B. subtilis, A. niger and P. funiculosum | ||
| A-acetolactate-decarboxylase | B. subtilis | ||
| Amyloglucosidase | A. niger and A. flavus | ||
| Cellulase | T. longibrachiatum | ||
| Pectinase | A. niger | ||
| Beverages | Glucose oxidase | P. notatum | Clarification of juices Removing O2 Hydrolysis of esters |
| Tannase | A. niger | ||
| Meat | Papain | S. aureus | Tenderization of meat |
| Protease | T. longibrachiatum, A. niger, A. oryzae and B. subtilis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation 2020, 6, 106. https://doi.org/10.3390/fermentation6040106
Sharma R, Garg P, Kumar P, Bhatia SK, Kulshrestha S. Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods. Fermentation. 2020; 6(4):106. https://doi.org/10.3390/fermentation6040106
Chicago/Turabian StyleSharma, Ranjana, Prakrati Garg, Pradeep Kumar, Shashi Kant Bhatia, and Saurabh Kulshrestha. 2020. "Microbial Fermentation and Its Role in Quality Improvement of Fermented Foods" Fermentation 6, no. 4: 106. https://doi.org/10.3390/fermentation6040106