
Nattokinase: Insights into Biological Activity, Therapeutic Applications, and the Influence of Microbial Fermentation
1
Beijing Key Laboratory of Bioprocess, College of Life Science and Technology, Beijing University of Chemical Technology, Beijing 100029, China
2
Steel Research Engineering Design Co., Ltd., Beijing 100081, China
3
National Energy R&D Center for Biorefinery, Beijing 100029, China
*
Authors to whom correspondence should be addressed.
Fermentation 2023, 9(11), 950; https://doi.org/10.3390/fermentation9110950
Submission received: 5 September 2023
/
Revised: 8 October 2023
/
Accepted: 23 October 2023
/
Published: 2 November 2023
(This article belongs to the Special Issue Research on Microbial Protein Synthesis)
Abstract
:Nattokinase, a serine protease that originates from the traditional food natto, has garnered widespread attention due to its pharmacological functions and therapeutic potential. This review aims to delve into the major advancements of nattokinase across various domains, particularly its emerging roles in Alzheimer’s disease prevention and the treatment of retinal diseases, thereby seeking to usher in a newfound hope in the fields of neurology and ophthalmology. However, the production and preservation of nattokinase present a multitude of challenges, including issues of unstable yield and enzyme activity loss. To address these challenges, we explore potential solutions such as the heterologous expression of the nattokinase gene, the optimization of microbial fermentation strategies, and innovative purification methods. Furthermore, we focus on enhancing the stability and protection of nattokinase through encapsulation and immobilization techniques, thus ensuring its sustainability across a wide array of applications. This review provides readers with the opportunity to gain an in-depth understanding of the diverse prospects for nattokinase applications. Future research directions will encompass a deeper exploration of its biological mechanisms, the development of novel nattokinase derivatives, and the extension of its applications into a broader spectrum of disease treatments and health maintenance.
1. Introduction
During the process of natto fermentation, Bacillus subtilis produces a diverse array of bioactive compounds (Figure 1A). These encompass pyrazines [1], which enhance food flavor; rennet, which is utilized in cheese production [2]; menaquinone-7 (vitamin K2) for the prevention of osteoporosis and cardiovascular diseases [3,4]; fructose polymers [5], poly-γ-glutamic acid (γ-PGA) [6]; and nattokinase. These substances find applications in various industrial and health-related contexts. Nattokinase (EC 3.4. 21.62), functioning as a fibrinolytic enzyme (Figure 1B), is believed to have potential benefits in preventing and treating cardiovascular diseases [7,8]. It not only directly hydrolyzes fibrin and fibrinolytic substrates to dissolve blood clots, but it also converts endogenous prourokinase into urokinase, degrades plasminogen activator inhibitor-1 (PAI-1), and stimulates endothelial cells to produce the tissue-type plasminogen activator (t-PA), thus enhancing thrombolysis [9]. Compared to traditional thrombolytic drugs like urokinase and streptokinase, nattokinase offers advantages in safety, suitability for long-term consumption, and an ease in oral administration [8]. Its products are widely available in tablet and capsule forms (Figure 1C), albeit with noticeable price variations based on their sources. As purification techniques continue to advance, there is a potential for reduced nattokinase production costs, thus rendering it an economically viable therapeutic option.
Nattokinase, a protease enzyme, is primarily produced through fermentation by Bacillus subtilis, but it can also be sourced from other microorganisms. These microorganisms encompass different species of Bacillus sp., such as B. subtilis [10], B. amyloliquefaciens [11], B. mojavensis [12], B. velezensis [13], and B. licheniformis [14], as well as a Pseudomonas sp. like P. aeruginosa [15]. Initially, it was isolated and identified from the traditional Japanese food, natto. Similar fibrinolytic-producing strains have been found in Chinese douchi [16] and soybean paste [17], as well as in Korean Chungkook-Jang [18], Doen-Jang [19], and pickle [13]. Nattokinase-producing strains can also be isolated from milk [15], oceans [20], rust [21], and soil [22]. Therefore, the sources of nattokinase production are remarkably diverse. This diversity offers extensive possibilities for the application of nattokinase in the food industry and biotechnology. It also underscores the importance and potential of microbial resources. Consequently, further research into nattokinase holds the promise of advancing the development of more beneficial applications while enriching our understanding of the role of microorganisms in production and the food industry.
This review distinguishes itself with a focus on the latest nattokinase research, thereby encompassing its applications in various diseases, enzyme activity detection methods, and strategies to enhance production yield and stability. Unlike previous reviews, it explores recent applications—such as Alzheimer’s prevention, retinal disease, as well as tumor and inflammation treatment—and is supported by current evidence. Additionally, it delves into nattokinase’s stability, drawing insights from recent studies. This comprehensive approach sets this review apart, thereby providing readers with the latest insights into nattokinase’s evolving role and its potential impact on health and industry. Future investigations will strive to gain deeper insights into the molecular mechanisms and pathways of nattokinase, and they will employ innovative scientific methodologies and technological approaches to facilitate the industrial development of nattokinase. These efforts aim to contribute further wisdom and innovation to human health.
2. Physicochemical Properties and the Formation Process of Nattokinase
Nattokinase is an alkaline serine protease, and its enzymatic activity and stability are subject to the influence of factors such as temperature, pH, and metal ions. Research findings indicate that nattokinase exhibits stability within the temperature range of 40 °C to 65 °C [23]. Even after undergoing five freeze–thaw cycles, its activity can still be maintained at over 95% [24]. Nattokinase demonstrates relative stability in neutral and slightly alkaline environments. However, its stability is notably compromised under acidic conditions, particularly when the pH falls below 5, thus resulting in a complete loss of enzymatic activity [25]. Metal ions also exert varying effects on nattokinase activity. Co2+, Mg2+, and Ba2+ have been found to significantly enhance its activity, whereas Zn2+, Al3+, Fe3+, Cu2+, and Hg2+ can inhibit its activity to differing degrees [26].
The encoding gene of nattokinase is known as aprN, and it starts with a GTG initiation codon [27]. This gene encodes a total of 381 amino acids, which includes a signal peptide sequence (29 amino acids) that is responsible for guiding nattokinase to be secreted into the extracellular space, a propeptide sequence (77 amino acids) that aids in proper folding, and the mature nattokinase peptide sequence (275 amino acids) [27]. The mature nattokinase peptide primarily consists of 9 β-folds and 9 α-helices [28]. Upon correct folding, the mature peptide becomes a biologically active protein with a relative molecular mass of approximately 27.7 kDa and an isoelectric point of 8.6 ± 0.3 [29]. The catalytic activity center of nattokinase is a conserved catalytic triad consisting of Asp32, His64, and Ser221, while the substrate-binding site is composed of three conserved amino acids, namely Ser125, Leu126, and Gly127 [30]. These structural characteristics of nattokinase form the basis for its specific functions and activities.
Through pathway analysis, Unrean and Nguyen [31] revealed that, under glycerol substrate conditions, B. subtilis possesses 2650 metabolic pathways, with 1272 of them capable of producing nattokinase. The most efficient nattokinase synthesis pathway involves the conversion of glycerol into ATP and NADPH through glycolysis, the pentose phosphate pathway (PPP), and an incomplete tricarboxylic acid cycle, whereby nattokinase and CO2 are ultimately yielded. The production of nattokinase necessitates the simultaneous utilization of both the oxidative and non-oxidative branches of the PPP to generate amino acid precursors. These findings underscore the intricacy and efficiency of the nattokinase synthesis pathway in B. subtilis when grown on glycerol as a substrate, which holds significant relevance for understanding cellular adaptation and metabolic pathway design under varying environmental conditions.
3. The Health Benefits of Nattokinase
Currently, research on the functionalities of nattokinase focuses on its effects such as the antithrombotic [32], anticoagulant [33], antiplatelet [34], and antihypertensive activities [35]. In recent years, the additional functions of nattokinase have emerged, including its potential in preventing Alzheimer’s disease, treating retinal disorders, combating hyperlipidemia, and addressing inflammatory bowel diseases (Figure 2).
3.1. Anti-Alzheimer’s Disease Effect of Nattokinase
Alzheimer’s disease (AD) is a neurodegenerative disorder that is characterized by its etiology, which involves multifaceted processes that include protein abnormalities, inflammation, and oxidative stress [36,37,38]. Within the cerebral milieu of afflicted individuals, anomalous proteins such as β-amyloid (Aβ) and Tau proteins aggregate, thereby forming plaques and neurofibrillary tangles that disrupt normal neuronal functionality [39,40,41]. Nattokinase, functioning as a serine protease, holds the capacity not only to dissolve thrombi, but to also effectively degrade Aβ, thereby demonstrating a certain therapeutic efficacy in the prevention of AD [42]. In a model induced by Aβ42, mice exhibited indications of depression, anxiety, and memory impairments. Following an extended administration of nattokinase, a normalization of neurosignaling components within the hippocampus of mice was observed, and this was concomitant with a relief of AD symptoms [38]. Ni et al. [43] further demonstrated that, upon entering the systemic circulation, nattokinase potentially clears Aβ42/Aβ40 deposits from various body compartments, thereby disrupting the dynamic equilibrium of Aβ42/Aβ40 between the brain and plasma. This phenomenon alleviates the burden of cerebral Aβ aggregation. Aβ42 within the brain can be transported to the peripheral system through various pathways, and elevated peripheral Aβ levels also impact its clearance within the brain. Therefore, it is imperative and pressing to establish the targeted binding and degradation of various Aβ42 aggregates by nattokinase, as well as the influence of nattokinase on the levels of Aβ in the bloodstream and its physiological significance. This could potentially represent a novel strategy and avenue for the prevention and treatment of AD.
3.2. Antiretinopathy Effect of Nattokinase
Retinopathy can arise from various etiologies, including diabetic retinopathy (DR) due to hyperglycemia, retinal vascular disorders that stem from hypertension or arteriosclerosis, and retinal vein occlusion [44,45]. Pathological neovascularization in the retina is a prevalent complication. Nuclear factor-erythroid 2-related factor-2 (Nrf2) and its downstream target heme oxygenase-1 (HO-1) play a role in promoting reparative vascular regeneration while regulating inflammation and oxidative responses [46]. In a murine model of oxygen-induced retinal pathology, nattokinase attenuates neuroinflammation and inhibits pathological neovascularization by activating the Nrf2/HO-1 signaling pathway and modulating neuroglial activation [45]. For diabetic macular edema, commonly employed treatments involve injections of anti-vascular endothelial growth factor agents or corticosteroids, but there are still side effects such as drug resistance, macular ischemia, and disease recurrence [44,47,48]. Maintaining the integrity of the blood–retinal barrier (BRB) is fundamental in treating DR. Huang et al. [47] demonstrated that nattokinase interferes with downstream signaling by inhibiting the expression of high-mobility group protein 1, thereby enhancing BRB function. This heralds the potential of nattokinase as a prospective alternative medication for treating diabetic retinal pathologies. However, selecting the appropriate delivery route, maintaining nattokinase activity, and determining the optimal dosage remain unresolved issues.
3.3. Antitumor Effect of Nattokinase
Tumor formation involves complex molecular mechanisms, including genetic mutations [49], aberrations in tumor suppressor genes and oncogenes [50], chromosomal anomalies [51], and environmental exposures [52]. The tumor physical microenvironment (TPME) influences the growth, metabolism, metastasis, and therapeutic efficacy of aberrant cells. The intratumoral injection of nattokinase in solid tumors has been demonstrated to modulate TPME, thereby enhancing intratumoral oxygen saturation and drug accumulation [53]. Furthermore, Zhang et al. [53] established a mouse–human breast tumor model, showing that pretreatment with nattokinase before chimeric antigen receptor–T cell therapy effectively suppressed solid tumor progression. FOXM1 and MMP2 are transcription factors, which promote cancer stem cell proliferation and significant targets in tumor regulation, respectively [54,55]. Zhang et al. [56] discovered that nattokinase has the capacity to prevent breast cancer occurrence and can suppress the expression of FOXM1 and MMP2 in tumor tissue, thus providing novel insights for future breast cancer treatment strategies. Yan et al. [57] treated a mouse model of hepatocellular carcinoma (HCC) with a nattokinase crude extract, thereby resulting in the inhibited expression of cancer-associated factors, as well as leading to an enhanced survival rate in HCC-afflicted mice. Despite the potential anti-tumor properties of nattokinase, there is still a need for further exploration due to the lack of clinical evidence, insufficient specificity and targeting, as well as the challenges regarding bioavailability and dosing.
3.4. Anti-Hyperlipidemic and Anti-Atherosclerotic Effects of Nattokinase
Hyperlipidemia is a disorder associated with disrupted lipid metabolism, and it is characterized by elevated levels of total cholesterol (TC) and triglycerides (TG), or reduced levels of high-density lipoprotein cholesterol (HDL-C) in the blood [58,59]. Several studies have indicated that the consumption of nattokinase or its derivatives in animal models significantly reduces TC and TG levels while increasing HDL-C levels, thus directly demonstrating its notable hypolipidemic effects [60,61,62]. Nattokinase not only holds benefits in lipid-lowering, but it also exhibits a potential against atherosclerosis (AS). Suzuki et al. [63] highlighted that the supplementation of nattokinase extract to mice with endothelial damage in the femoral artery significantly reduces intimal thickening, thus suggesting at an anti-atherosclerotic effect from nattokinase. Furthermore, nattokinase improves lipid metabolism in mice by inhibiting the oxidation of low-density lipoprotein cholesterol, thereby preventing the onset of AS [60]. Clinical research has translated nattokinase application into practice. A study involving 1062 volunteers demonstrated that a daily intake of 10,800 FU of nattokinase significantly reduced carotid intima-media thickness (CCA-IMT) and plaque size, with AS and hyperlipidemia improvements ranging from 66.5 to 95.4% [64]. Ren et al. [65] compared nattokinase to simvastatin, revealing that, after 26 weeks of treatment, the nattokinase-treated group exhibited more significant reductions in CCA-IMT and carotid plaque size. Nattokinase, as a potential alternative therapy for hyperlipidemia, holds promising prospects. It not only demonstrates significant lipid-lowering effects in animal models, but it may also play a positive role in combating atherosclerosis. Clinical research results support this notion, thereby showing that nattokinase has a significant impact on improving carotid intima-media thickness and plaque size. These findings provide new avenues for seeking more effective treatments for hyperlipidemia patients, especially those who cannot tolerate traditional drug therapies, which is where nattokinase could emerge as a crucial option. However, further research and long-term monitoring are necessary to comprehensively assess its safety and long-term effectiveness.
3.5. Other Effects of Nattokinase
In addition to its common antithrombotic, antihypertensive, and antiplatelet functions, the role of nattokinase in the treatment of colitis and post-stroke cognitive recovery has also been discovered in recent years. Escherichia coli Nissle 1917 is commonly utilized for the treatment of colitis [66]. Liang et al. [67] constructed an engineered Escherichia coli Nissle 1917 strain with nattokinase expression capacity, which was able to further enhance the anti-inflammatory effect, promote the proliferation and differentiation of epithelial stem cells, as well as aid in the repair of the damaged barriers in mice with colitis. In a murine model of inflammatory bowel disease (IBD) that was induced by sodium polysaccharide sulfate, nattokinase addresses chronic colitis by modulating the composition of intestinal microbiota, thus reducing the production of pro-inflammatory cytokines and inhibiting the mucosal barrier damage caused by cell apoptosis [68].
Post-stroke cognitive impairment (PSCI) after ischemic stroke (IS) represents a potential incapacitating effect, and it stands as one of the most prevalent complications of IS [69]. Irisin functions as a crucial modulator that enhances the expression of the brain-derived neurotrophic factor in the hippocampus [70]. Wu et al. [71] demonstrated that nattokinase treatment significantly elevates irisin levels, consequently stimulating hippocampal neurogenesis, regulating learning and memory processes, and ameliorating PSCI. However, the mechanisms underlying nattokinase’s absorption and its circulation pathways within the body remain to be elucidated.
4. Enzyme Activity Assay of Nattokinase
The activity of nattokinase can reflect its therapeutic efficacy as a pharmaceutical or health product. Selecting appropriate detection methods during production can not only provide accurate and reproducible measurement results, but it can also enhance the safety and compliance of nattokinase products. Methods for assessing the activity of nattokinase can be categorized into two main types: one that exploits its fibrinolytic properties, including techniques like the clot lysis time (CLT) method, fibrin plate method, and fibrin degradation method; and the other type, which is based on nattokinase’s hydrolytic properties and involves methods such as the TAME method, Folin phenol method, and tetrapeptide substrate method. However, there is currently no globally standardized detection method, which may lead to inconsistent results between different laboratories. To further advance the research and applications of nattokinase, international standardization organizations and experts in the field need to collaborate to establish globally recognized detection standards and methods. This will help ensure the quality and efficacy of nattokinase, as well as provide better medical options for patients. Table 1 provides a summary of the various activity determination methods.
5. Optimization Strategies for Nattokinase Production
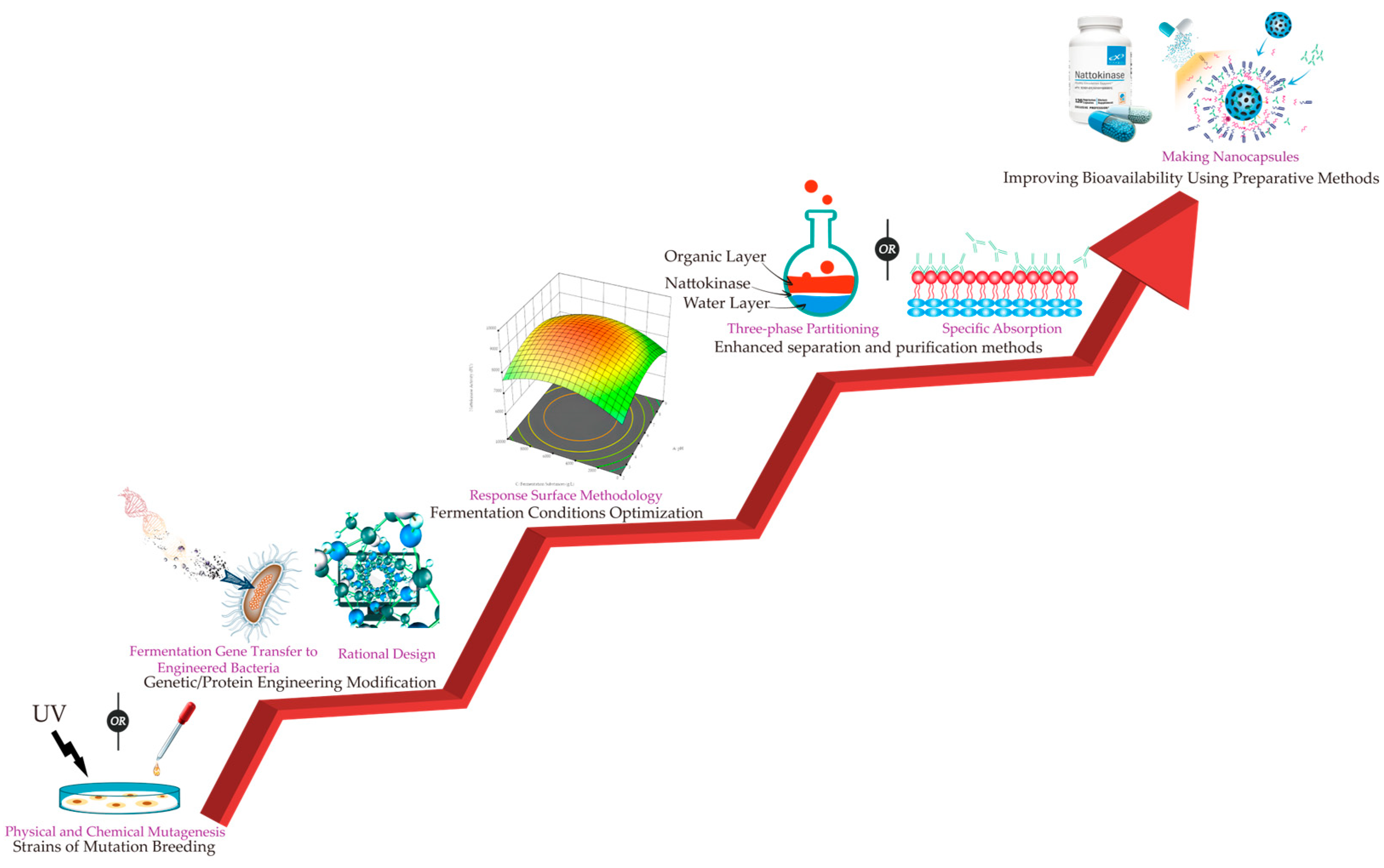
In the pursuit of an industrial-scale production of nattokinase, the enhancement of nattokinase yield emerges as a paramount consideration. The strategies frequently employed encompass the screening and mutagenesis of wild-type microorganisms, thereby utilizing genetic engineering techniques to engineer the nattokinase gene, as well as the optimization of fermentation media and cultivation parameters. Furthermore, ensuring the stability and efficacy of nattokinase during its purification and preparation process is also of paramount importance (Figure 3).
5.1. Mutation Breeding of a Nattokinase-Producing Strain
In microbial fermentation, obtaining high-quality products efficiently is crucial, and this is closely related to having a well-characterized and genetically stable strain. Strains selected from nature often fail to meet the production demands of industrial fermentation. Therefore, applying mutagenesis breeding to the modification of nattokinase-producing strains can enhance both enzyme production capacity and purity. Physical or chemical methods are commonly used for mutagenesis breeding. Yang [79] achieved a maximum nattokinase activity of 534.95 U/mL and an 11.84% improvement over the wild-type strain by subjecting the original Bacillus subtilis strain to UV mutagenesis. On the other hand, Wang et al. [80] used a combination of UV and 60Co-γ irradiation to mutate the wild-type strain Bacillus subtilis JNFE0126, thereby resulting in a mutant strain (Bacillus subtilis JNFE1126) with doubled nattokinase production. A comparative analysis of the whole genomes and transcriptomes of these two strains revealed 28 single nucleotide polymorphisms, 15 insertion–deletions, and significant differential expression (≥2-fold change) in 1425 genes, thus providing valuable genomic information for screening high-yield nattokinase genes. Wang et al. [81] also performed chemical mutagenesis using N-methyl-N’-nitroso-N-nitroguanidine (NTG) in addition to UV and γ-radiation on the original strain, thereby ultimately obtaining a stable and highly productive mutant strain—B. subtilis LD-8547.
The mutagenesis breeding technique holds significant importance in the production of nattokinase as it can optimize the enzyme production capacity of the original strain and can create novel strains that can aid in reducing production costs. However, mutagenesis breeding also entails certain risks, such as mutations that may cause the strain to lose its growth ability or have adverse effects on the environment. Therefore, strict control over the entire process of the mutagenesis strain is necessary.
5.2. Construction of Nattokinase Expression Systems and Molecular Modification
In 1992, Nakamura et al. [27] determined the nucleotide sequence of the nattokinase gene, thus enabling the possibility of using genetic engineering techniques to enhance nattokinase production. Subsequently, researchers have successfully amplified the coding gene of nattokinase from the genome of Bacillus subtilis, which secretes nattokinase. They constructed expression vectors for the nattokinase gene and achieved expression in bacteria, fungi, animal cells, and plants. The characteristics of the different expression systems are summarized in Table 2.
The use of protein engineering techniques such as site-directed mutagenesis, protein cleavage, and rational design allows for targeted modifications of the amino acid sequence of nattokinase, thereby improving its activity and stability. Weng et al. [76] constructed a single mutant variant I31L of nattokinase through site-directed mutagenesis, which exhibited twice the catalytic activity when compared to the wild-type enzyme. By applying a computer-aided rational design to the molecular modification of nattokinase, not only can the protein’s structure be precisely designed and modified, but it can also reduce the number of mutants and improve work efficiency [25]. Liu et al. [88] employed three rational design strategies, including surface charge engineering, sequence comparison, and literature-based design, to perform a site-directed mutagenesis of nattokinase, as well as to enhance its resistance. The obtained single-point mutants, Q59E, S78T, and N218D, exhibited improved specific activity, acid stability, and thermal stability, respectively.
Currently, there are limitations in using protein engineering to modify nattokinase. These include a limited understanding of the structure–function relationship of nattokinase, challenges in predicting the effects of mutations on enzyme activity and stability, and the possibility of introducing unpredictable changes in other characteristics. Therefore, a comprehensive evaluation and characterization are necessary before application to ensure the safety and efficacy of the modifications.
5.3. Optimizing Nattokinase Production through Variations in Fermentation Conditions
Traditional solid-state fermentation has been conventionally employed for nattokinase production. However, this approach leads to the generation of high-viscosity γ-PGA polymers, which pose challenges in terms of separation and potential allergenic reactions [17]. Additionally, this methodology is associated with extended production cycles, erratic yield patterns, and susceptibility to contamination, thereby rendering it inadequate for meeting the demands of industrial-scale production [25].
Liquid-state fermentation is widely adopted for nattokinase production. Many studies have focused on optimizing the nattokinase production environment through modifications in carbon and nitrogen sources in the culture medium, as well as through variations in factors such as temperature, pH, and fermentation duration. These efforts aim to create the most favorable conditions for nattokinase production, thereby enhancing yields. Investigations into the impact of process scale up on nattokinase production have also been conducted (Table 3).
In addition to optimizing production conditions during fermentation, reducing the cost of culture media is crucial. Previous studies have demonstrated the feasibility of utilizing waste materials to produce nattokinase, thereby promoting both the circular economy and low-cost production. Li et al. [89] investigated the use of tofu processing wastewater, which was rich in nitrogen sources, as a substitute for traditional culture media in nattokinase production. Through the optimization of medium composition and parameters, Bacillus subtilis 13932 achieved a nattokinase activity of 10,661.97 IU/mL in a 100 L bioreactor. Sahoo et al. [75] employed industrial waste cheese whey as a low-cost culture medium for nattokinase fermentation. By mixing cheese whey with 10 g/L of yeast extract, the nattokinase yield reached fermentation levels comparable to those achieved with glucose as a substrate, thus resulting in cost savings of 55–60%. These findings highlighted the pivotal role of selecting appropriate fermentation conditions and substrates in enhancing nattokinase production efficiency. However, further exploration is still required to address mass transfer and oxygen solubility issues in industrial-scale amplification processes.
5.4. Purification of Nattokinase
In recent years, the development of novel purification techniques has made it possible to obtain low-cost, readily available, and highly active nattokinase. The three-phase partitioning (TPP) technique—which is commonly used for protein purification due to its mild conditions, scalability, and cost-effectiveness—has received limited attention in the enrichment of nattokinase. Garg and Thorat [95] employed the TPP method to enrich crude nattokinase at the interface between the lower aqueous phase (ammonium sulfate) and the upper organic phase (tert-butanol), and this was achieved by investigating the effects of ammonium sulfate and the tert–butanol ratio, as well as the temperature and pH on purification efficiency. The purification fold of nattokinase reached 5.6 with a recovery rate of 129.5%. In addition to the three-phase partitioning technique, the ultrasound-assisted separation of nattokinase from a fermentation crude extract provided a new approach [96]. Hu et al. [97] utilized expanded bed adsorption to purify the nattokinase from the fermentation broth, thereby achieving a purification fold of 8.2 with a recovery rate of 95%. This method not only increased the recovery rate by 50%, but it also significantly shortened the purification time compared to traditional methods. The expanded bed adsorption method based on electrostatic adsorption principles achieved solid–liquid separation, product concentration, and preliminary purification in a single unit operation, thereby fully demonstrating the integrated advantages of the separation process. Compared to the other methods, magnetic microsphere affinity adsorption technology allows for the specific adsorption of proteins directly from the fermentation broth. This not only reduces protein loss, but it also shortens the pretreatment time of the fermentation broth. Yang et al. [98] successfully achieved the direct purification and separation of nattokinase from the fermentation broth using magnetic polymethylmethacrylate (PMMA) beads, whereby only 40 minutes were required to achieve purification fold and activity recovery rates of 8.7 and 85%, respectively. Liu et al. [99] effectively adsorbed nattokinase from the fermentation broth using arginine-modified magnetic nanoparticles, thereby achieving purification fold and maximum adsorption capacities of 8.0 and 3.79 U/mg nanoparticles, respectively (which was twice that of lysine-modified nanoparticles). It is evident that by using magnetic metals or metal oxide carriers with shell modifications and functional groups that bind to the target protein, the rapid separation and purification of nattokinase can be achieved under the action of an external magnetic field. The higher the purity and activity of nattokinase, the greater its potential in pharmaceutical and food health applications. Therefore, to achieve the large-scale industrial production of high-quality nattokinase, efficient, rapid, and cost-effective purification and separation methods need to be continually explored.
5.5. Protection of Nattokinase
Due to its potential benefits in preventing thrombosis and hypertension, nattokinase is considered a promising novel therapeutic agent for various human diseases. However, its susceptibility to degradation by proteases and acids in the gastrointestinal tract has led to relatively low oral bioavailability of nattokinase [10,17]. Additionally, the poor permeability of the intestinal mucosa, along with the enzyme’s instability and susceptibility to inactivation, further complicates its absorption in the gastrointestinal tract. Addressing this issue, current research focuses on the development of nattokinase microcapsules using various biodegradable materials such as lipids, polysaccharides, and biopolymers. These microcapsules aim to enhance the thermal stability, pH stability, and gastrointestinal absorption of nattokinase (Table 4) [100].
The existing delivery systems face certain limitations, including the inability of the porous structure of polysaccharides to effectively prevent gastric fluid permeation, the biotoxicity associated with surface-active emulsifiers, and the unclear mechanism of drug absorption by small intestinal epithelial cells [100]. Further research efforts should be directed towards the development of acid-resistant materials, the refinement of emulsion delivery systems, and exploration of the absorption mechanisms within the gastrointestinal tract.
6. Conclusions
Nattokinase is a third-generation fibrinolytic enzyme that is extracted and purified from fermented soy-based food in Japan. This review describes the functions of nattokinase beyond the treatment of cardiovascular diseases, including the prevention of Alzheimer’s disease and retinopathy, as well as resistance to hyperlipidemia and atherosclerosis. These research findings provide references for the further development of nattokinase as pharmaceuticals or functional health products. Additionally, the activity and quality of nattokinase significantly impact its efficacy as a product, thereby necessitating the selection of appropriate activity detection methods in different experimental and production environments. Various techniques have been employed to facilitate the industrial production of nattokinase. The mutagenesis of nattokinase-producing strains, the construction of expression systems, and molecular modification have been used to improve the stability of nattokinase and reduce the difficulty of purification. Optimization of fermentation conditions, the enhancement of isolation and purification methods, and the use of encapsulation and immobilization techniques have been used to improve and maintain the activity of nattokinase. To improve the bioavailability of nattokinase, the continuous exploration of its biological effects and the discovery of more convenient, precise, and versatile detection methods are necessary. Furthermore, research on strategies to enhance nattokinase production and activity is needed to provide new insights for further development and production.
Author Contributions
M.F.: conceptualization; original draft preparation; data curation; and writing—review and editing; visualization. B.Y.: original draft preparation; data curation; and project administration. M.W.: writing—review and editing; visualization; supervision; and project administration. J.L.: original draft preparation; data curation; and conceptualization. Z.W.: writing—review and editing; supervision; and project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Key Research and Development Program of China (No. 2018YFA0900201).
Conflicts of Interest
Author Yuan Beichen was employed by the company Steel Research Engineering Design Co., Ltd at the time of the study. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Kłosowski, G.; Mikulski, D.; Pielech-Przybylska, K. Pyrazines Biosynthesis by Bacillus Strains Isolated from Natto Fermented Soybean. Biomolecules 2021, 11, 1736. [Google Scholar] [CrossRef] [PubMed]
- Shieh, C.-J.; Phan Thi, L.-A.; Shih, I.-L. Milk-clotting Enzymes Produced by Culture of Bacillus subtilis Natto. Biochem. Eng. J. 2009, 43, 85–91. [Google Scholar] [CrossRef]
- Bus, K.; Szterk, A. Relationship between Structure and Biological Activity of Various Vitamin K Forms. Foods 2021, 10, 3136. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, X.; Wang, L.; Wu, H.; Zhao, G.; Liu, H.; Wang, P.; Zheng, Z. Coproduction of Menaquinone-7 and Nattokinase by Bacillus subtilis Using Soybean Curd Residue as a Renewable Substrate Combined with a Dissolved Oxygen Control Strategy. Ann. Microbiol. 2018, 68, 655–665. [Google Scholar] [CrossRef]
- Vieira, A.M.; Zahed, F.; Crispim, A.C.; de Souza Bento, E.; França, R.d.F.O.; Pinheiro, I.O.; Pardo, L.A.; Carvalho, B.M. Production of Levan from Bacillus subtilis var. natto and Apoptotic Effect on SH-SY5Y Neuroblastoma Cells. Carbohydr. Polym. 2021, 273, 118613. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Li, S.; Tian, Z.; Ma, X. Study on the Mechanism of Production of γ-PGA and Nattokinase in Bacillus subtilis Natto Based on RNA-seq Analysis. Microb. Cell Factories 2021, 20, 83. [Google Scholar] [CrossRef]
- Dabbagh, F.; Negahdaripour, M.; Berenjian, A.; Behfar, A.; Mohammadi, F.; Zamani, M.; Irajie, C.; Ghasemi, Y. Nattokinase: Production and Application. Appl. Microbiol. Biotechnol. 2014, 98, 9199–9206. [Google Scholar] [CrossRef]
- Wu, H.; Wang, H.; Xu, F.; Chen, J.; Duan, L.; Zhang, F. Acute Toxicity and Genotoxicity Evaluations of Nattokinase, a Promising Agent for Cardiovascular Diseases Prevention. Regul. Toxicol. Pharmacol. 2019, 103, 205–209. [Google Scholar] [CrossRef]
- Weng, Y.; Yao, J.; Sparks, S.; Wang, K.Y. Nattokinase: An Oral Antithrombotic Agent for the Prevention of Cardiovascular Disease. Int. J. Mol. Sci. 2017, 18, 523. [Google Scholar] [CrossRef]
- Zhang, X.; Tong, Y.; Wang, J.; Lyu, X.; Yang, R. Screening of a Bacillus subtilis Strain Producing both Nattokinase and Milk-clotting Enzyme and Its Application in Fermented Milk with Thrombolytic Activity. J. Dairy. Sci. 2021, 104, 9437–9449. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, Q.; Zhang, R.-h.; Zhang, Y.-z. Purification and Characterization of a Fibrinolytic Enzyme Produced by Bacillus amyloliquefaciens DC-4 Screened from douchi, a Traditional Chinese Soybean Food. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 134, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, X.; Chen, L.; Xu, X.; Li, J. Characterization of a Nattokinase from the Newly Isolated Bile Salt-Resistant Bacillus mojavensis LY-06. Foods 2022, 11, 2403. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.; Kim, J.-H.; Kwak, M.-S.; Sung, M.-H.; Jeong, D.-W. Functional Annotation Genome Unravels Potential Probiotic Bacillus velezensis Strain KMU01 from Traditional Korean Fermented Kimchi. Foods 2021, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Wei, X.; Qiu, Y.; Chen, Y.; Chen, J.; Wen, Z.; Chen, S. High-Level Expression of Nattokinase in Bacillus licheniformis by Manipulating Signal Peptide and Signal Peptidase. J. Appl. Microbiol. 2016, 121, 704–712. [Google Scholar] [CrossRef]
- Devi, C.S.; Mohanasrinivasan, V.; Sharma, P.; Das, D.; Vaishnavi, B.; Naine, S.J. Production, Purification and Stability Studies on Nattokinase: A Therapeutic Protein Extracted from Mutant Pseudomonas aeruginosa CMSS Isolated from Bovine Milk. Int. J. Pept. Res. Ther. 2016, 22, 263–269. [Google Scholar] [CrossRef]
- Wang, C.T.; Ji, B.P.; Li, B.; Nout, R.; Li, P.L.; Ji, H.; Chen, L.F. Purification and Characterization of a Fibrinolytic Enzyme of Bacillus subtilis DC33, Isolated from Chinese Traditional Douchi. J. Ind. Microbiol. Biotechnol. 2006, 33, 750–758. [Google Scholar] [CrossRef]
- Wei, X.; Luo, M.; Xie, Y.; Yang, L.; Li, H.; Xu, L.; Liu, H. Strain Screening, Fermentation, Separation, and Encapsulation for Production of Nattokinase Functional Food. Appl. Biochem. Biotechnol. 2012, 168, 1753–1764. [Google Scholar] [CrossRef]
- Kim, W.; Choi, K.; Kim, Y.; Park, H.; Choi, J.; Lee, Y.; Oh, H.; Kwon, I.; Lee, S. Purification and Characterization of a Fibrinolytic Enzyme Produced from Bacillus sp. Strain CK 11-4 Screened from Chungkook-Jang. Appl. Environ. Microbiol. 1996, 62, 2482–2488. [Google Scholar] [CrossRef]
- Kim, S.-H.; Choi, N.-S. Purification and Characterization of Subtilisin DJ-4 Secreted by Bacillus sp. Strain DJ-4 Screened from Doen-Jang. Biosci. Biotechnol. Biochem. 2000, 64, 1722–1725. [Google Scholar] [CrossRef]
- Mahajan, P.M.; Nayak, S.; Lele, S.S. Fibrinolytic Enzyme from Newly Isolated Marine Bacterium Bacillus subtilis ICTF-1: Media Optimization, Purification and Characterization. J. Biosci. Bioeng. 2012, 113, 307–314. [Google Scholar] [CrossRef]
- Vaithilingam, M.; Chandrasekaran, S.D.; Gupta, S.; Paul, D.; Sahu, P.; Selvaraj, J.N.; Babu, V. Extraction of Nattokinase Enzyme from Bacillus cereus Isolated from Rust. Natl. Acad. Sci. Lett. 2016, 39, 263–267. [Google Scholar] [CrossRef]
- Wang, S.-L.; Yeh, P.-Y. Production of a Surfactant- and Solvent-stable Alkaliphilic Protease by Bioconversion of Shrimp Shell Wastes Fermented by Bacillus subtilis TKU007. Process Biochem. 2006, 41, 1545–1552. [Google Scholar] [CrossRef]
- Kumar, D.J.M.; Rakshitha, R.; Vidhya, M.A.; Jennifer, P.S.; Prasad, S.; Kumar, M.R.; Kalaichelvan, P.T. Production, Optimization and Characterization of Fibrinolytic Enzyme by Bacillus subtilis RJAS19. Pak. J. Biol. Sci. 2014, 17, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A Novel Fibrinolytic Enzyme (Nattokinase) in the Vegetable Cheese Natto; a Typical and Popular Soybean Food in the Japanese Diet. Experientia 1987, 43, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liangqi, C.; Xiyu, T.; Jinyao, L. Biotechnology, Bioengineering and Applications of Bacillus Nattokinase. Biomolecules 2022, 12, 980. [Google Scholar] [CrossRef]
- Yin, L.-J.; Lin, H.-H.; Jiang, S.-T. Bioproperties of Potent Nattokinase from Bacillus subtilis YJ1. J. Agric. Food Chem. 2010, 58, 5737–5742. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamagata, Y.; Ichishima, E. Nucleotide Sequence of the Subtilisin NAT Gene, aprN, of Bacillus subtilis (natto). Biosci. Biotechnol. Biochem. 1992, 56, 1869–1871. [Google Scholar] [CrossRef]
- Yanagisawa, Y.; Chatake, T.; Chiba-Kamoshida, K.; Naito, S.; Ohsugi, T.; Sumi, H.; Yasuda, I.; Morimoto, Y. Purification, Crystallization and Preliminary X-ray Diffraction Experiment of Nattokinase from Bacillus subtilis natto. Acta Cryst. 2010, 66, 1670–1673. [Google Scholar] [CrossRef]
- Bhatt, P.C.; Verma, A.; Al-Abbasi, F.A.; Anwar, F.; Kumar, V.; Panda, B.P. Development of Surface-engineered PLGA Nanoparticulate-Delivery System of Tet1-conjugated Nattokinase Enzyme for Inhibition of Aβ40 Plaques in Alzheimer’s Disease. Int. J. Nanomedicine 2017, 2017, 8749–8768. [Google Scholar] [CrossRef]
- Carter, P.; Wells, J.A. Dissecting the Catalytic Triad of a Serine Protease. Nature 1988, 332, 564–568. [Google Scholar] [CrossRef]
- Unrean, P.; Nguyen, N.H.A. Metabolic Pathway Analysis and Kinetic Studies for Production of Nattokinase in Bacillus subtilis. Bioprocess. Biosyst. Eng. 2013, 36, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Guangbo, Y.; Min, S.; Wei, S.; Lixin, M.; Chao, Z.; Yaping, W.; Zunxi, H. Heterologous Expression of Nattokinase from B. subtilis Natto Using Pichia pastoris GS115 and Assessment of Its Thrombolytic activity. BMC Biotechnol. 2021, 21, 49. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, H.; Li, W.; Zhang, C.; Liu, Y.; Xu, F.; Chen, J.; Duan, L.; Zhang, F. Nattokinase-Heparin Exhibits Beneficial Efficacy and Safety—An Optimal Strategy for CKD Patients on Hemodialysis. Glycoconj. J. 2019, 36, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.-Y.; Kim, T.-S.; Cai, J.; Kim, J.; Kim, Y.; Shin, K.; Kim, K.S.; Park, S.K.; Lee, S.-P.; Choi, E.-K.; et al. Nattokinase Improves Blood Flow by Inhibiting Platelet Aggregation and Thrombus Formation. Lab. Anim. Res. 2013, 29, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Ohnishi, K.; Takaoka, S.; Ogasawara, K.; Fukuyama, R.; Nakamuta, H. Antihypertensive Effects of Continuous Oral Administration of Nattokinase and Its Fragments in Spontaneously Hypertensive Rats. Biol. Pharm. Bull. 2011, 34, 1696–1701. [Google Scholar] [CrossRef]
- Querfurth, H.W.; LaFerla, F.M. Alzheimer’s Disease. N. Engl. J. Med. 2010, 362, 329–344. [Google Scholar] [CrossRef]
- Paramanick, D.; Singh, V.D.; Singh, V.K. Neuroprotective Effect of Phytoconstituents via Nanotechnology for Treatment of Alzheimer Diseases. J. Control Release 2022, 351, 638–655. [Google Scholar] [CrossRef]
- Naik, S.; Katariya, R.; Shelke, S.; Patravale, V.; Umekar, M.; Kotagale, N.; Taksande, B. Nattokinase Prevents β-Amyloid Peptide (Aβ1-42) Induced Neuropsychiatric Complications, Neuroinflammation and BDNF Signalling Disruption in Mice. Eur. J. Pharmacol. 2023, 952, 175821. [Google Scholar] [CrossRef]
- Rahman, M.M.; Lendel, C. Extracellular Protein Components of Amyloid Plaques and Their Roles in Alzheimer’s Disease Pathology. Mol. Neurodegener. 2021, 16, 59. [Google Scholar] [CrossRef]
- Fadl, N.N.; Ahmed, H.H.; Booles, H.F.; Sayed, A.H. Serrapeptase and Nattokinase Intervention for Relieving Alzheimer’s Disease Pathophysiology in Rat Model. Hum. Exp. Toxicol. 2013, 32, 721–735. [Google Scholar] [CrossRef]
- Sheng, M.; Sabatini, B.L.; Südhof, T.C. Synapses and Alzheimer’s Disease. Cold Spring Harb. Perspect. Biol. 2012, 4, a005777. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.-L.; Lee, K.-T.; Wang, J.-H.; Lee, L.Y.L.; Chen, R.P.Y. Amyloid-Degrading Ability of Nattokinase from Bacillus subtilis Natto. J. Agric. Food Chem. 2009, 57, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Ni, A.; Li, H.; Wang, R.; Sun, R.; Zhang, Y. Degradation of Amyloid β-Peptides Catalyzed by Nattokinase in vivo and in vitro. Food Sci. Hum. Wellness 2023, 12, 1905–1916. [Google Scholar] [CrossRef]
- Al-Shabrawey, M.; Elsherbiny, M.; Nussbaum, J.; Othman, A.; Megyerdi, S.; Tawfik, A. Targeting Neovascularization in Ischemic Retinopathy: Recent Advances. Expert. Rev. Ophthalmol. 2013, 8, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Ng, T.K.; Chen, W.; Sun, X.; Huang, D.; Zheng, D.; Yi, J.; Xu, Y.; Zhuang, X.; Chen, S. Nattokinase Attenuates Retinal Neovascularization Via Modulation of Nrf2/HO-1 and Glial Activation. Investig. Ophthalmol. Vis. Sci. 2021, 62, 25. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 System in Development, Oxidative Stress Response and Diseases: An Evolutionarily Conserved Mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Huang, Z.; Chu, W.K.; Ng, T.K.; Chen, S.; Liang, J.; Chen, C.-B.; Xu, Y.; Xie, B.; Ke, S.; Liu, Q.; et al. Protective Effects of Nattokinase Against Microvasculopathy and Neuroinflammation in Diabetic Retinopathy. J. Diabetes 2023, 15, 866–880. [Google Scholar] [CrossRef]
- Chung, E.J.; Roh, M.I.; Kwon, O.W.; Koh, H.J. Effects of Macular Ischemia on the Outcome of Intravitreal Bevacizumab Therapy for Diabetic Macular Edema. Retina 2008, 28, 957–963. [Google Scholar] [CrossRef]
- Suwa, T.; Kobayashi, M.; Nam, J.-M.; Harada, H. Tumor Microenvironment and Radioresistance. Exp. Mol. Med. 2021, 53, 1029–1035. [Google Scholar] [CrossRef]
- Emmanuel, N.K.; Antonios, K.; Athanasios, S.; Dimitrios, S.; Aikaterini, M.; Nikolaos, G.; Michail, D.; Kyveli, A.; Georgios, T.; Athanasios, P.; et al. Role of Oncogenes and Tumor-Suppressor Genes in Carcinogenesis: A Review. Anticancer Res. 2020, 40, 6009. [Google Scholar] [CrossRef]
- Kou, F.; Wu, L.; Ren, X.; Yang, L. Chromosome Abnormalities: New Insights into Their Clinical Significance in Cancer. Mol. Ther. Oncolytics 2020, 17, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.R.; Buha Djordjevic, A. Heavy Metal and Pesticide Exposure: A Mixture of Potential Toxicity and Carcinogenicity. Curr. Opin. Toxicol. 2020, 19, 72–79. [Google Scholar] [CrossRef]
- Zhang, Y.; Pei, P.; Zhou, H.; Xie, Y.; Yang, S.; Shen, W.; Hu, L.; Zhang, Y.; Liu, T.; Yang, K. Nattokinase-Mediated Regulation of Tumor Physical Microenvironment to Enhance Chemotherapy, Radiotherapy, and CAR-T Therapy of Solid Tumor. ACS Nano 2023, 17, 7475–7486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wei, P.; Gong, A.; Chiu, W.-T.; Lee, H.-T.; Colman, H.; Huang, H.; Xue, J.; Liu, M.; Wang, Y.; et al. FoxM1 Promotes β-Catenin Nuclear Localization and Controls Wnt Target-Gene Expression and Glioma Tumorigenesis. Cancer Cell 2011, 20, 427–442. [Google Scholar] [CrossRef]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 01370. [Google Scholar] [CrossRef]
- Zhang, B.; Chai, J.; He, L.; Dusanbieke, M.; Gong, A. Nattokinase Produced by Natto Fermentation with Bacillus subtilis Inhibits Breast Cancer Growth. Int. J. Clin. Exp. Med. 2019, 12, 13380–13387. [Google Scholar]
- Yan, Y.; Wang, Y.; Qian, J.; Wu, S.; Ji, Y.; Liu, Y.; Zeng, J.; Gong, A. Nattokinase Crude Extract Inhibits Hepatocellular Carcinoma Growth in Mice. J. Microbiol. Biotechnol. 2019, 29, 1281–1287. [Google Scholar] [CrossRef]
- Yao, Y.S.; Li, T.D.; Zeng, Z.H. Mechanisms Underlying Direct Actions of Hyperlipidemia on Myocardium: An Updated Review. Lipids Health Dis. 2020, 19, 23. [Google Scholar] [CrossRef]
- Rauf, A.; Akram, M.; Anwar, H.; Daniyal, M.; Munir, N.; Bawazeer, S.; Bawazeer, S.; Rebezov, M.; Bouyahya, A.; Shariati, M.A.; et al. Therapeutic Potential of Herbal Medicine for the Management of Hyperlipidemia: Latest Updates. Environ. Sci. Pollut. Res. 2022, 29, 40281–40301. [Google Scholar] [CrossRef]
- Iwai, K.; Nakaya, N.; Kawasaki, Y.; Matsue, H. Antioxidative Functions of Natto, A Kind of Fermented Soybeans: Effect on LDL Oxidation and Lipid Metabolism in Cholesterol-Fed Rats. J. Agric. Food Chem. 2002, 50, 3597–3601. [Google Scholar] [CrossRef]
- Yang, N.C.; Chou, C.W.; Chen, C.Y.; Hwang, K.L.; Yang, Y.C. Combined Nattokinase with Red Yeast Rice but not Nattokinase Alone has Potent Effects on Blood Lipids in Human Subjects with Hyperlipidemia. Asia Pac. J. Clin. Nutr. 2009, 18, 310–317. [Google Scholar] [PubMed]
- Lu, Y.; Ding, H.; Jiang, X.; Zhang, H.; Ma, A.; Hu, Y.; Li, Z. The Effects of the Extract from Peanut Meal Fermented with Bacillus natto and Monascus on Lipid Metabolism and Intestinal Barrier Function of Hyperlipidemic in Mice. J. Sci. Food Agric. 2021, 101, 2561–2569. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kondo, K.; Matsumoto, Y.; Zhao, B.-Q.; Otsuguro, K.; Maeda, T.; Tsukamoto, Y.; Urano, T.; Umemura, K. Dietary Supplementation of Fermented Soybean, Natto, Suppresses Intimal Thickening and Modulates the Lysis of Mural Thrombi after Endothelial Injury in Rat Femoral Artery. Life Sci. 2003, 73, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, J.; Zhang, F.; Li, Y.; Wang, R.; Zheng, Q.; Zhang, X.; Zeng, J.; Xu, F.; Lin, Y. Effective Management of Atherosclerosis Progress and Hyperlipidemia with Nattokinase: A Clinical Study with 1,062 Participants. Front. Cardiovasc. Med. 2022, 9, 964977. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.N.; Chen, H.J.; Li, Y.; McGowan, G.W.; Lin, Y.G. A Clinical Study on the Effect of Nattokinase on Carotid Artery Atherosclerosis and Hyperlipidaemia. Natl. Med. J. China 2017, 97, 2038–2042. [Google Scholar] [CrossRef]
- Arribas, B.; Rodríguez-Cabezas, M.E.; Camuesco, D.; Comalada, M.; Bailón, E.; Utrilla, P.; Nieto, A.; Concha, A.; Zarzuelo, A.; Gálvez, J. A Probiotic Strain of Escherichia coli, Nissle 1917, Given Orally Exerts Local and Systemic Anti-Inflammatory Effects in Lipopolysaccharide-Induced Sepsis in Mice. Br. J. Pharmacol. 2009, 157, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Zhang, J.; Yang, Y.; Xia, Y.; Liu, L.; Liu, L.; Wang, Q.; Gao, X. Nattokinase Enhances the Preventive Effects of Escherichia coli Nissle 1917 on Dextran Sulfate Sodium-Induced Colitis in Mice. World J. Microbiol. Biotechnol. 2022, 39, 8. [Google Scholar] [CrossRef]
- Zhou, L.; Hao, N.; Li, X.; Chen, J.; Yang, R.; Song, C.; Sun, Y.; Zhang, Q. Nattokinase Mitigated Dextran Sulfate Sodium-induced Chronic Colitis by Regulating Microbiota and Suppressing Tryptophan Metabolism via Inhibiting IDO-1. J. Funct. Foods 2020, 75, 104251. [Google Scholar] [CrossRef]
- He, A.; Wang, Z.; Wu, X.; Sun, W.; Yang, K.; Feng, W.; Wang, Y.; Song, H. Incidence of Post-Stroke Cognitive Impairment in Patients with First-Ever Ischemic Stroke: A Multicenter Cross-Sectional Study in China. Lancet Reg. Health–West. Pac. 2023, 33, 100687. [Google Scholar] [CrossRef]
- Islam, M.R.; Valaris, S.; Young, M.F.; Haley, E.B.; Luo, R.; Bond, S.F.; Mazuera, S.; Kitchen, R.R.; Caldarone, B.J.; Bettio, L.E.B.; et al. Exercise Hormone Irisin is a Critical Regulator of Cognitive Function. Nat. Metab. 2021, 3, 1058–1070. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Q.; Xu, P.; Chen, J.; Duan, L.; Xu, F.; Zhang, F. Nattokinase Promotes Post-stroke Neurogenesis and Cognition Recovery via Increasing Circulating Irisin. J. Agric. Food Chem. 2023, 71, 11418–11428. [Google Scholar] [CrossRef]
- Astrup, T.; Müllertz, S. The Fibrin Plate Method for Estimating Fibrinolytic Activity. Arch. Biochem. Biophys. 1952, 40, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Song, J.Y.; Kim, K.M.; Kim, M.K.; Lee, I.Y.; Kim, S.B.; Kim, H.S.; Han, N.S.; Lee, B.H.; Kim, B.S. Production of Nattokinase by Batch and Fed-Batch Culture of Bacillus subtilis. N. Biotechnol. 2010, 27, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Du, M.; Zheng, D.; Kong, F.; Zu, G.; Feng, Y. Purification and Characterization of Nattokinase from Bacillus subtilis Natto B-12. J. Agric. Food Chem. 2009, 57, 9722–9729. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Mahanty, B.; Daverey, A.; Dutta, K. Nattokinase Production from Bacillus subtilis Using Cheese Whey: Effect of Nitrogen Supplementation and Dynamic Modelling. J. Water Process. Eng. 2020, 38, 101533. [Google Scholar] [CrossRef]
- Weng, M.; Deng, X.; Bao, W.; Zhu, L.; Wu, J.; Cai, Y.; Jia, Y.; Zheng, Z.; Zou, G. Improving the Activity of the Subtilisin Nattokinase by Site-Directed Mutagenesis and Molecular Dynamics Simulation. Biochem. Biophys. Res. Commun. 2015, 465, 580–586. [Google Scholar] [CrossRef]
- Yuki, Y.; Nakagawa, T.; Fujita, M.; Asada, A.; Nakanishi, K.; Kato, K. A Sandwich Enzyme-linked Immunosorbent Assay for Nattokinase. Biosci. Biotechnol. Biochem. 1994, 58, 366–370. [Google Scholar] [CrossRef]
- Chen, J.; Wei, Q.; Jiang, F.; Liu, Z.; Lai, Y.; He, J.; Li, B. A New Bacillus Subtilis Strain and Its Use in Preparing Medicine for Treating Thrombosis. U.S. Patent 11/913,002, 14 August 2008. [Google Scholar]
- Yang, Y. Research on breeding of Bacillus subtilis natto by ultraviolet mutation. In Proceedings of the Biophysical Society of GuangDong Province Academic Forum: Precise Photons and Life Health (PPLH 2022), Guangzhou, China, 9–11 December 2022; SPIE: Bellingham, WA, USA, 11 December 2022; Volume 12603, pp. 21–25. [Google Scholar]
- Wang, Y.; Wang, J.; Zhang, X.; Tong, Y.; Yang, R. Genomic and Transcriptomic Analysis of Bacillus subtilis JNFE1126 with Higher Nattokinase Production Through Ultraviolet Combined 60Co-γ Ray Mutagenesis. LWT 2021, 147, 111652. [Google Scholar] [CrossRef]
- Wang, S.H.; Zhang, C.; Yang, Y.L.; Diao, M.; Bai, M.F. Screening of a High Fibrinolytic Enzyme Producing Strain and Characterization of the Fibrinolytic Enzyme Produced from Bacillus subtilis LD-8547. World J. Microbiol. Biotechnol. 2008, 24, 475–482. [Google Scholar] [CrossRef]
- Gu, Z.; Ning, C.; Liu, Z.; Liang, Q.; Fu, X.; Tian, M.; Zhu, C.; Mou, H. High-Efficiency Heterologous Expression of Nattokinase Based on a Combinatorial Strategy. Process Biochem. 2023, 133, 65–74. [Google Scholar] [CrossRef]
- Wu, S.-M.; Feng, C.; Zhong, J.; Huan, L.-D. Enhanced Production of Recombinant Nattokinase in Bacillus subtilis by Promoter Optimization. World J. Microbiol. Biotechnol. 2011, 27, 99–106. [Google Scholar] [CrossRef]
- Li, C.; Du, Z.; Qi, S.; Zhang, X.; Wang, M.; Zhou, Y.; Lu, H.; Gu, X.; Tian, H. Food-grade expression of nattokinase in Lactobacillus delbrueckii subsp. bulgaricus and its thrombolytic activity in vitro. Biotechnol. Lett. 2020, 42, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Guo, P.-C.; Jiang, W.-L.; Fan, X.-M.; Luo, X.-Y.; Li, H.-H. Expression of Nattokinase in Escherichia coli and Renaturation of its inclusion body. J. Biotechnol. 2016, 231, 65–71. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Xiong, S.; Zhang, J.; Cai, L.; Yang, Y. Expression and Purification of Recombinant Nattokinase in Spodoptera frugiperda cells. Biotechnol. Lett. 2007, 29, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, L.; Liu, J.; Li, H.; Wang, Y.; Hasi, A. Transient Expression of Optimized and Synthesized Nattokinase Gene in Melon (Cucumis melo L.) Fruit by Agroinfiltration. Plant Biotechnol. 2015, 32, 175–180. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, H.; Han, L.; Cui, W.; Zhou, L.; Zhou, Z. Improvement of the acid resistance, catalytic efficiency, and thermostability of nattokinase by multisite-directed mutagenesis. Biotechnol. Bioeng. 2019, 116, 1833–1843. [Google Scholar] [CrossRef]
- Li, T.; Zhan, C.; Guo, G.; Liu, Z.; Hao, N.; Ouyang, P. Tofu Processing Wastewater as a Low-Cost Substrate for High Activity Nattokinase Production Using Bacillus subtilis. BMC Biotechnol. 2021, 21, 57. [Google Scholar] [CrossRef]
- Ju, S.; Cao, Z.; Wong, C.; Liu, Y.; Foda, M.F.; Zhang, Z.; Li, J. Isolation and Optimal Fermentation Condition of the Bacillus subtilis Subsp. natto Strain WTC016 for Nattokinase Production. Fermentation 2019, 5, 92. [Google Scholar] [CrossRef]
- Keziah, S.M.; Devi, C.S. Fibrinolytic and ACE Inhibitory Activity of Nattokinase Extracted from Bacillus subtilis VITMS 2: A Strain Isolated from Fermented Milk of Vigna unguiculata. Protein J. 2021, 40, 876–890. [Google Scholar] [CrossRef]
- Xiao, Z.; Shen, J.; Li, Y.; Wang, Z.; Zhao, Y.; Chen, Y.; Zhao, J.-y. High and Economical Nattokinase Production with Acetoin as a Useful Byproduct from Soybean Milk and Glucose. Probiotics Antimicrob. Proteins 2022, 14, 792–803. [Google Scholar] [CrossRef]
- Mahajan, P.M.; Gokhale, S.V.; Lele, S.S. Production of Nattokinase Using Bacillus natto NRRL 3666: Media Optimization, Scale Up, and Kinetic Modeling. Food Sci. Biotechnol. 2010, 19, 1593–1603. [Google Scholar] [CrossRef]
- Pan, S.; Chen, G.; Zeng, J.; Cao, X.; Zheng, X.; Zeng, W.; Liang, Z. Fibrinolytic Enzyme Production from Low-Cost Substrates by Marine Bacillus subtilis: Process Optimization and Kinetic Modeling. Biochem. Eng. J. 2019, 141, 268–277. [Google Scholar] [CrossRef]
- Garg, R.; Thorat, B.N. Nattokinase Purification by Three Phase Partitioning and Impact of T-butanol on Freeze Drying. Sep. Purif. Technol. 2014, 131, 19–26. [Google Scholar] [CrossRef]
- Avhad, D.N.; Niphadkar, S.S.; Rathod, V.K. Ultrasound Assisted Three Phase Partitioning of a Fibrinolytic Enzyme. Ultrason. Sonochem 2014, 21, 628–633. [Google Scholar] [CrossRef]
- Hu, H.-B.; Yao, S.-J.; Mei, L.-H.; Zhu, Z.-Q.; Hur, B.-K. Partial Purification of Nattokinase from Bacillus subtilis by Expanded Bed Adsorption. Biotechnol. Lett. 2000, 22, 1383–1387. [Google Scholar] [CrossRef]
- Yang, C.; Xing, J.; Guan, Y.; Liu, H. Superparamagnetic Poly(methyl methacrylate) Beads for Nattokinase Purification from Fermentation Broth. Appl. Microbiol. Biotechnol. 2006, 72, 616–622. [Google Scholar] [CrossRef]
- Liu, C.-H.; Wu, W.-C.; Lai, H.-Y. Adsorption of Nattokinase by Amino Acid-Conjugated Magnetic Nanoadsorbents. Sep. Sci. Technol. 2013, 48, 923–930. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, L.; Zeng, X. Research Progress on the Utilisation of Embedding Technology and Suitable Delivery Systems for Improving the Bioavailability of Nattokinase: A Review. Food Struct. 2021, 30, 100219. [Google Scholar] [CrossRef]
- Dong, X.-Y.; Kong, F.-P.; Yuan, G.-Y.; Wei, F.; Jiang, M.-L.; Li, G.-M.; Wang, Z.; Zhao, Y.-D.; Chen, H. Optimisation of Preparation Conditions and Properties of Phytosterol Liposome-Encapsulating Nattokinase. Nat. Prod. Res. 2012, 26, 548–556. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, J.; Liu, C.; Li, J.; Li, Z.; Zhao, J.; Liu, H. Synthesis of Sustained Release/Controlled Release Nanoparticles Carrying Nattokinase and Their Application in Thrombolysis. Pharmazie 2021, 76, 145–149. [Google Scholar] [CrossRef]
- Kapoor, R.; Harde, H.; Jain, S.; Panda, A.K.; Panda, B.P. Downstream Processing, Formulation Development and Antithrombotic Evaluation of Microbial Nattokinase. J. Biomed. Nanotechnol. 2015, 11, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lyu, X.; Tong, Y.; Wang, J.; Ye, J.; Yang, R. Chitosan/Casein Based Microparticles with a Bilayer Shell–core Structure for Oral Delivery of Nattokinase. Food Funct. 2020, 11, 10799–10816. [Google Scholar] [CrossRef] [PubMed]
- Wen, N.; Lü, S.; Xu, X.; Ning, P.; Wang, Z.; Zhang, Z.; Gao, C.; Liu, Y.; Liu, M. A Polysaccharide-Based Micelle-Hydrogel Synergistic Therapy System for Diabetes and Vascular Diabetes Complications Treatment. Mater. Sci. Eng. C 2019, 100, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-s.; Zhou, M.-j.; Hu, L.-x.; Huang, K.; Zhao, S.-g.; Qian, S.-h.; Wang, Z.; Huang, X.-l.; Huang, J.-B.; Wang, J.; et al. Preparation and Property Studies of Enteric-Soluble Oral Nattokinase Microcapsules with Halloysite Nanotubes. Int. J. Polym. Mater. 2023, 1–13. [Google Scholar] [CrossRef]
- Wu, C.; Gao, C.; Lü, S.; Xu, X.; Wen, N.; Zhang, S.; Liu, M. Construction of Polylysine Dendrimer Nanocomposites Carrying Nattokinase and Their Application in Thrombolysis. J. Biomed. Mater. Res. Part. A 2018, 106, 440–449. [Google Scholar] [CrossRef]
- Ye, W.; Wang, N.; Hu, K.; Zhang, L.; Liu, A.; Pan, C.; Gong, T.; Liu, T.; Ding, H. Bio-inspired Microcapsule for Targeted Antithrombotic Drug Delivery. RSC Adv. 2018, 8, 27253–27259. [Google Scholar] [CrossRef]
- Deepak, V.; Ram Kumar Pandian, S.b.; Kalishwaralal, K.; Gurunathan, S. Purification, Immobilization, and Characterization of Nattokinase on PHB Nanoparticles. Bioresour. Technol. 2009, 100, 6644–6646. [Google Scholar] [CrossRef]
- Li, D.; Hu, M.; Hou, L.; Gao, Y.; Tian, Z.; Wen, W.; Fan, B.; Li, S.; Wang, F. The Structural and Functional Properties of Soybean Protein-Polyglutamic Acid Complex Effected the Stability of W/O/W Emulsion Encapsulated Nattokinase. Food Chem. 2023, 414, 135724. [Google Scholar] [CrossRef]
Figure 1.
Traditional Japanese fermented food—natto. (A) Edible natto. (B) The three-dimensional structure of nattokinase. (C) Commercially available nattokinase products.
Figure 1.
Traditional Japanese fermented food—natto. (A) Edible natto. (B) The three-dimensional structure of nattokinase. (C) Commercially available nattokinase products.
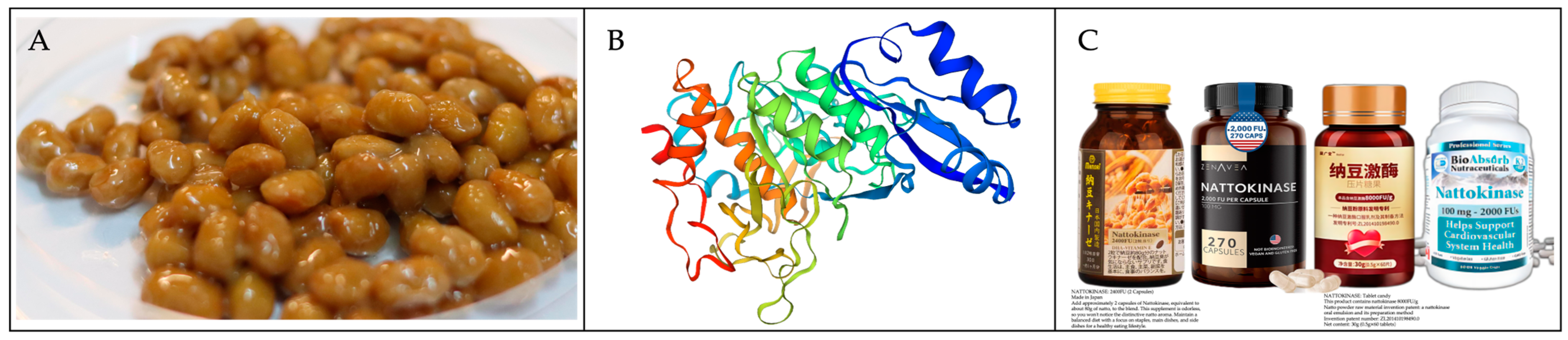
Figure 2.
The medicinal value of nattokinase.
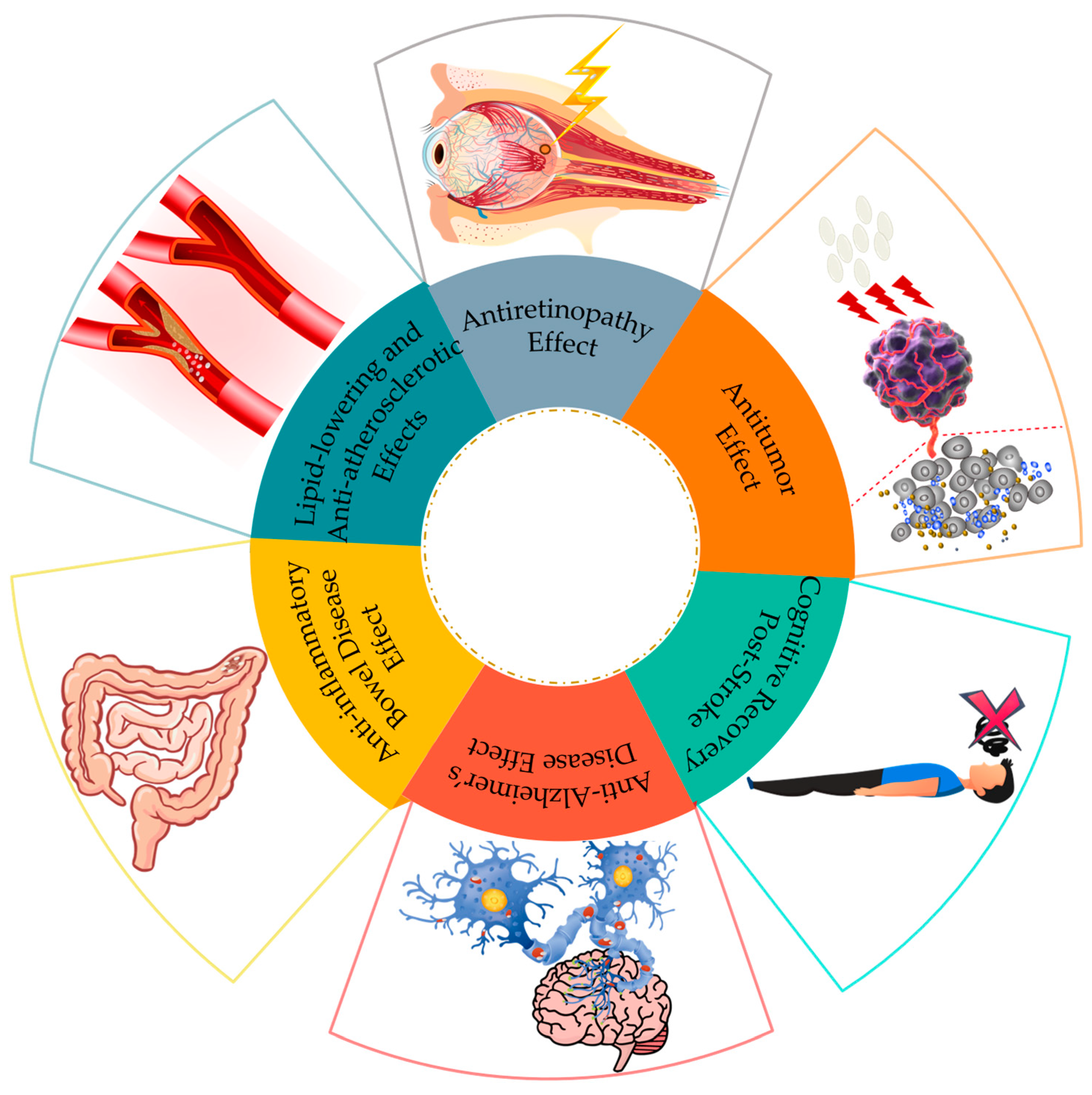
Figure 3.
Approaches to enhance the yield and stability of nattokinase.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Nattokinase activity detection method.
| Method | Principle | Pros | Cons | Ref. |
|---|---|---|---|---|
| Fibrin plate method | Nattokinase degrades the artificial thrombus to form a dissolving circle | Measures multiple samples at the same time | Detection accuracy is greatly affected by time, plate thickness, and product purity | [72] |
| Fibrin degradation method | Fibrin degradation products have an absorption peak at 275 nm | High accuracy | Poor stability and high concentration requirements for sample dilutions, as well as cumbersome operation and long determination time | [73] |
| CLT method | Timing the hydrolysis of fibrin by nattokinase for comparison with the standards | Rapid reaction, suitable for the determination of crude enzymes | Not suitable for simultaneous measurement of multiple samples and products with high sensitivity | [74] |
| Folin phenol method | Nattokinase hydrolyzes casein to produce tyrosine, which reacts with a Folin phenol reagent for color development | Multiple samples can be measured simultaneously at low cost. | Low specificity, and a determination of all protease activities | [75] |
| TAME method | Nattokinase cleaves arginine carboxy-terminal peptide and undergoes a chromogenic reaction | High reaction sensitivity and simple operation | Impurities in the crude enzyme solution has a greater impact on the results | [6] |
| Tetrapeptide substrate method | Hydrolysis of Suc-Ala-Pro-Phe-pNA with a chromogenic reaction | Simple and highly sensitive method for the rapid determination of enzyme activities | The assay does not directly reflect the fibrinolytic activity of nattokinase | [76] |
| ELISA method | Enzyme-linked reaction to determine nattokinase enzyme activity by the combination of marker enzymes and catalase | High specificity and sensitivity | Complex operation and high testing cost | [77] |
| HPLC method | Protein analytical columns for the detection of nattokinase | Fast analysis, good selectivity, and high sensitivity. | Not suitable for detecting crude enzymes, and there will be a great deal of stray peaks | [78] |
Table 2.
Expression of the nattokinase gene in different species.
| Recombinant Expression Vectors | Hosts | Characteristics |
|---|---|---|
| pP43NMK-NKF | B. subtilis WB800 | Screening high-expression signal peptides for nattokinase and optimizing the signal peptide’s start codon from GTG to ATG. Establishing a dual promoter system and employing a combinatorial strategy to enhance nattokinase production [82]. |
| pHY10 | B. subtilis DB104 | Replacing the -10 box of the aprN promoter with the consensus sequence (TATAAT) of the σA-dependent promoter [83]. |
| pMGthyA-ppNK | Lactobacillus delbrueckii subsp. bulgaricus | Employing a chromosomal–plasmid complementation expression system, with the thyA gene serving as a complementary marker, to ensure that the product expression remains unhindered by antibiotic resistance in the system. The expression vector and host strain are both of food-grade quality [84]. |
| pHY-SP-NK | B. licheniformis△0F-3 | Constructing an expression vector within a gene-deficient host strain, whereby the aprE signal peptide is incorporated and the type I signal peptidase gene sipV is overexpressed [14]. |
| pQE30-m-NK | E. coli BL21 (DE3) | A substantial amount of inactive insoluble protein was detected in the inclusion bodies [85]. |
| pHBM905BDM-NK-Bs | Pichia pastoris GS115 | The promoter of the vector pHBM905BDM significantly enhances protein expression levels. The expression level of nattokinase reaches its maximum, when there are five copies of the aprN gene [32]. |
| Bacmid-egfp-NK | Spodoptera frugiperda cells | The Bac-to-Bac system was employed to express nattokinase that was fused with a His tag in insect cells while retaining its fibrinolytic activity [86]. |
| pPZP35SNi | Cucumis melo L. | Transient expression of the sNKi gene in melon fruit using the E8 promoter-involved expression system resulted in higher nattokinase activity [87]. |
Table 3.
Optimization of liquid fermentation conditions for nattokinase.
| Strain | Nitrogen Source | Carbon Source | Mineral Salt | Loading Volume | Temperature | pH | Enzyme Activity | Ref. |
|---|---|---|---|---|---|---|---|---|
| B. subtilis 13932 | 93.669% Tofu process wastewater | 3% Glucose | 0.1% MgSO4·7H2O 0.08% CaCl2 | 70 mL/500 mL Flask | 37 °C | 7.0 | 7209 ± 195 IU/mL (fibrin plate method) | [89] |
| B. subtilis MTCC 2616 | Cheese whey 10 g/L Yeast extract | Cheese whey | - | 50 mL/250 mL Flask | 30 °C | - | 833.43 U/mL (Folin phenol method) | [75] |
| B. subtilis Subsp. Natto Strain WTC016 | 1% Peptone 0.5% Yeast extract | - | 1% NaCl | 60 mL/250 mL Flask | 30 °C | 7.0 | 3284 ± 58 IU/mL (fibrin plate method) | [90] |
| B. subtilis JNFE0126 | 3% Kidney bean flour | 2% Sucrose | 0.2% NaH2PO4 0.4% K2HPO4 0.4% NaCl 0.02% CaCl2 | - Flask | 40 °C | 7.0 | 3511 U/mL (fibrin plate method) | [10] |
| B. subtilis VITMS 2 | 2% Soya bean waste 1.5% Eggshell power | 2% Cane molasses | 10 mM CaCl2 10 mM MgSO4 10 mM Na2HPO4 10 mM K2HPO4 | - Flask | 30 °C | 7.0 | 2748.76 FU/mL (modified fibrin degradation method) | [91] |
| B. subtilis NDF | 18% Soybean milk | 10.5% Glucose | - | 4 L/6 L Batch | 30 °C | 7.0 | 10,220 IU/mL (fibrin plate method) | [92] |
| B. subtilis | 5% Peptone 0.3% NH4Cl | 1% Glucose | 0.1% K2HPO4 0.2% MgSO4·7H2O Trace elements | 2 L/5 L Batch | 37 °C | 7.0 | 3400 U/mL (modified fibrin degradation method) | [73] |
| Bacillus natto NRRL 3666 | 1% Soy peptone 0.325% Yeast extract | 2% Glucose | 0.0407% K2HPO4·3H2O 0.044% MgSO4·7H2O 0.05% CaCl2·2H2O | 2.5 L/5 L Batch | 37 °C | 6.3 | 1932 U/mL (tetrapeptide substrate method) | [93] |
| B. subtilis D21-8 | 2.32% Soybean meal | 2.41% Cassava starch | 0.16% CaCl2 | 7.5 L/10 L Batch | 34 °C | 5.0–5.2 | 3203 U/mL (fibrin plate method) | [94] |
Table 4.
Encapsulation of nattokinase by different materials.
| Embedding Material | Property | Pros | Ref. | |
|---|---|---|---|---|
| Lipids, polysaccharides, proteins | Lecithin, phytosterol, and mannite | 65.25% 1 | Biocompatibility, fast release, and easy absorption | [101] |
| Sodium alginate, carboxymethyl chitosan, and Fe3O4 | 65.24% 2 | Sustained release and magnetic targeting | [102] | |
| Chitosan, tripolyphosphate, and glutaraldehyde | >90% 1 | Inexpensive raw materials and simple manufacture | [103] | |
| Casein and chitosan | 73.8% 1 | Safer and better tolerated | [104] | |
| Chitosan/dialdehyde starch derivatives micelle-hydrogel | 89.4% 2 | Easy self-assembly | [105] | |
| Artificial synthetic polymer | Eudragit L100-55-halloysite nanotubes | 31.6% 1 | Sustained release, and a reduction in the side effects caused by excessive thrombolysis | [106] |
| Polylysine dendrimer | 117% 3 | Biocompatibility and biodegradability | [107] | |
| Polystyrene and polydopamine | 75% 4 | Targets activated platelets with high affinity | [108] | |
| Polyhydroxybutyrate | 120% 3 | Increased enzyme stability | [109] | |
| W/O/W emulsions | Soybean isolate protein and polyglutamic acid | 97.19% 1 | Compact structure and good stability | [110] |
1 Encapsulation efficiency. 2 Release efficiency. 3 Relative enzyme activity. 4 Load efficiency.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fang, M.; Yuan, B.; Wang, M.; Liu, J.; Wang, Z. Nattokinase: Insights into Biological Activity, Therapeutic Applications, and the Influence of Microbial Fermentation. Fermentation 2023, 9, 950. https://doi.org/10.3390/fermentation9110950
AMA Style
Fang M, Yuan B, Wang M, Liu J, Wang Z. Nattokinase: Insights into Biological Activity, Therapeutic Applications, and the Influence of Microbial Fermentation. Fermentation. 2023; 9(11):950. https://doi.org/10.3390/fermentation9110950
Chicago/Turabian StyleFang, Mudannan, Beichen Yuan, Meng Wang, Junfeng Liu, and Zheng Wang. 2023. "Nattokinase: Insights into Biological Activity, Therapeutic Applications, and the Influence of Microbial Fermentation" Fermentation 9, no. 11: 950. https://doi.org/10.3390/fermentation9110950
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.