
Functional and Probiotic Characterization of Newly Isolated Strains from Infant Feces and Breast Milk

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification
2.2. Hemolytic Activity
2.3. Antibiotic Susceptibility
2.4. Antimicrobial Activity
2.5. Hydrophobicity
2.6. Auto-Aggregation
2.7. Adherence Assay
2.8. Scavenging of DPPH Radical
2.9. Resistance to Gastrointestinal Tract (GIT) Condition
2.10. Production of Exopolysachaccharides
2.11. Statistical Analysis
3. Results
3.1. Identification and Safety Assessment
3.2. Hydrophobicity of Cell Surface and Aggregation Potential
3.3. Adherence Assay
3.4. Antioxidant Activity of the Tested Isolates
3.5. Bile Salt Tolerance
3.6. Production of Exopolysaccharides
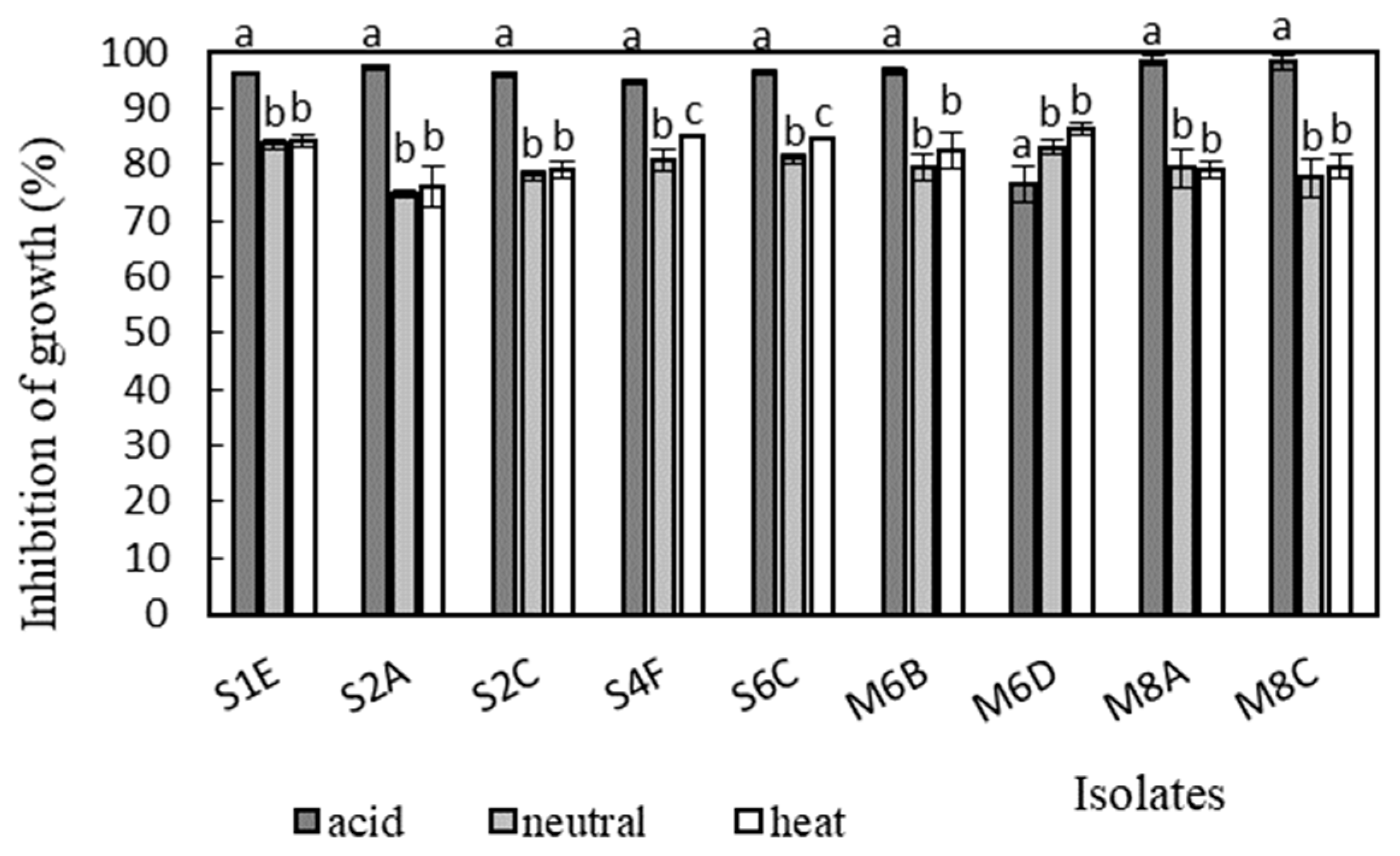
3.7. Antimicrobial Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asan-Ozusaglam, M.; Gunyakti, A. A new probiotic candidate bacterium from human milk: Limosilactobacillus vaginalis MA-10. Acta Aliment. 2021, 50, 13–21. [Google Scholar] [CrossRef]
- Damaceno, Q.S.; Gallotti, B.; Reis, I.M.; Totte, Y.C.; Assis, G.B.; Figueiredo, H.C.; Silva, T.F.; Azevedo, V.; Nicoli, J.R.; Martins, F.S. Isolation and identification of potential probiotic bacteria from human milk. Probiotics Antimicrob. Proteins 2021, 15, 491–501. [Google Scholar] [CrossRef]
- de Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Singh, T.P.; Natraj, B.H. Next-generation probiotics: A promising approach towards designing personalized medicine. Crit. Rev. Microbiol. 2021, 47, 479–498. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Rubayet Ul Alam, A.S.M.; Jahid, I.K. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef] [PubMed]
- Sionek, B.; Szydłowska, A.; Zielińska, D.; Neffe-Skocińska, K.; Kołożyn-Krajewska, D. Beneficial bacteria isolated from food in relation to the next generation of probiotics. Microorganisms 2023, 11, 1714. [Google Scholar] [CrossRef]
- Shin, M.; Ban, O.H.; Jung, Y.H.; Yang, J.; Kim, Y. Genomic characterization and probiotic potential of Lactobacillus casei IDCC 3451 isolated from infant faeces. Lett. Appl. Microbiol. 2021, 72, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.S.; Kim, Y.; Jeong, Y.; Kim, J.E.; Paek, N.S.; Kang, C.H. Antioxidant and probiotic properties of Lactobacilli and bifidobacteria of human origins. Biotechnol. Bioprocess Eng. 2020, 25, 421–430. [Google Scholar] [CrossRef]
- FAO/WHO. Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations/World Health Organization: London, UK; London, ON, Canada, 2002. [Google Scholar]
- Asan-Ozusaglam, M.; Gunyakti, A. Lactobacillus fermentum strains from human breast milk with probiotic properties and cholesterol-lowering effects. Food Sci. Biotechnol. 2019, 28, 501–509. [Google Scholar] [CrossRef]
- Gunyakti, A.; Asan-Ozusaglam, M. Investigation of the potential use of Lactobacillus gasseri originated from human breast milk as food additive. LWT 2018, 93, 613–619. [Google Scholar] [CrossRef]
- Rodrigues da Cunha, L.; Ferreira, C.L.F.; Durmaz, E.; Goh, Y.J.; Sanozky-Dawes, R.; Klaenhammer, T. Characterization of Lactobacillus gasseri isolates from a breastfed infant. Gut Microbes 2012, 3, 15–24. [Google Scholar] [CrossRef]
- Rastogi, S.; Mittal, V.; Singh, A. In vitro assessment of antioxidant and antimicrobial potential of Lactobacillus gasseri strains isolated from human milk and infant faeces. J. Pure Appl. Microbiol. 2020, 14, 1305–1315. [Google Scholar] [CrossRef]
- Verdenelli, M.C.; Ghelfi, F.; Silvi, S.; Orpianesi, C.; Cecchini, C.; Cresci, A. Probiotic properties of Lactobacillus rhamnosus and Lactobacillus paracasei isolated from human faeces. Eur. J. Nutr. 2009, 48, 355–363. [Google Scholar] [CrossRef]
- Cheng, L.; Akkerman, R.; Kong, C.; Walvoort, M.T.C.; de Vos, P. More than sugar in the milk: Human milk oligosaccharides as essential bioactive molecules in breast milk and current insight in beneficial effects. Crit. Rev. Food Sci. Nutr. 2021, 61, 1184–1200. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Mehwish, H.M.; Siddiq, M.; Haobin, Z.; Zhu, J.; Yan, L.; Shao, D.; Xu, X.; Shi, J. Identification, characterization, and probiotic potential of Lactobacillus rhamnosus isolated from human milk. Lebensm. Wiss. Technol. 2017, 84, 271–280. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Liu, J.; Liu, Y.; Wang, Y.; Xiao, Y.; Gao, B.; Zhu, D. In Vitro probiotic characterization of Lactobacillus strains from fermented tangerine vinegar and their cholesterol degradation activity. Food Biosci. 2021, 39, 100843. [Google Scholar] [CrossRef]
- Sharma, C.; Gulati, S.; Thakur, N.; Singh, B.P.; Gupta, S.; Kaur, S.; Mishra, S.K.; Puniya, A.K.; Gill, J.P.S.; Panwar, H. Antibiotic sensitivity pattern of indigenous lactobacilli isolated from curd and human milk samples. 3 Biotech 2017, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Kostelac, D.; Gerić, M.; Gajski, G.; Markov, K.; Domijan, A.M.; Čanak, I.; Jakopović, Ž.; Svetec, I.K.; Žunar, B.; Frece, J. Lactic acid bacteria isolated from equid milk and their extracellular metabolites show great probiotic properties and anti-inflammatory potential. Int. Dairy J. 2021, 112, 104828. [Google Scholar] [CrossRef]
- Krausová, G.; Hyršlová, I.; Hynštová, I. In Vitro evaluation of adhesion capacity, hydrophobicity, and auto-aggregation of newly isolated potential probiotic strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef]
- Musilova, S.; Modrackova, N.; Doskocil, I.; Svejstil, R.; Rada, V. Influence of human milk oligosaccharides on adherence of bifidobacteria and clostridia to cell lines. Acta Microbiol. Immunol. Hung. 2017, 64, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Hyrslova, I.; Kana, A.; Kantorova, V.; Krausova, G.; Mrvikova, I.; Doskocil, I. Selenium accumulation and biotransformation in Streptococcus, Lactococcus, and Enterococcus strains. J. Funct. Foods 2022, 92, 105056. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- EFSA. Technical guidance—Update of the criteria used in the assessment of bacterial resistance to antibiotics of human or veterinary importance. Eur. Food Saf. Authority J. 2008, 6, 732. [Google Scholar]
- Morandi, S.; Silvetti, T.; Brasca, M. Biotechnological and safety characterization of Enterococcus lactis, a recently described species of dairy origin. Antonie Leeuwenhoek 2013, 103, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Liasi, S.; Azmi, T.; Hassan, M.; Shuhaimi, M.; Rosfarizan, M.; Ariff, A. Antimicrobial activity and antibiotic sensitivity of three isolates of lactic acid bacteria from fermented fish product, Budu. Malays. J. Microbiol. 2009, 5, 33–37. [Google Scholar]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-wide assessment of antibiotic resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar] [CrossRef]
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial activity and antibiotic susceptibility of Lactobacillus and Bifidobacterium spp. intended for use as starter and probiotic cultures. Biotechnol. Biotechnol. Equip. 2015, 29, 84–91. [Google Scholar] [CrossRef]
- Suhonen, A. Antibiotic Susceptibility of Lactic Acid Bacteria. 2019. Available online: http://urn.fi/URN:NBN:fi:hulib-201905081914 (accessed on 20 September 2023).
- Elkins, C.A.; Mullis, L.B. Bile-mediated aminoglycoside sensitivity in Lactobacillus species likely results from increased membrane permeability attributable to cholic acid. Appl. Environ. Microbiol. 2004, 70, 7200–7209. [Google Scholar] [CrossRef]
- Kõll, P.; Mändar, R.; Smidt, I.; Hütt, P.; Truusalu, K.; Mikelsaar, R.H.; Shchepetova, J.; Krogh-Andersen, K.; Marcotte, H.; Hammarström, L.; et al. Screening and evaluation of human intestinal lactobacilli for the development of novel gastrointestinal probiotics. Curr. Microbiol. 2010, 61, 560–566. [Google Scholar] [CrossRef]
- Vesterlund, S.; Vankerckhoven, V.; Saxelin, M.; Goossens, H.; Salminen, S.; Ouwehand, A.C. Safety assessment of Lactobacillus strains: Presence of putative risk factors in faecal, blood and probiotic isolates. Int. J. Food Microbiol. 2007, 116, 325–331. [Google Scholar] [CrossRef]
- Jeong, Y.; Kim, H.; Lee, J.Y.; Won, G.; Choi, S.I.; Kim, G.H.; Kang, C.H. The antioxidant, anti-diabetic, and anti-adipogenesis potential and probiotic properties of lactic acid bacteria isolated from human and fermented foods. Fermentation 2021, 7, 123. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Sieo, C.C.; Kalavathy, R.; Liang, J.B.; Alitheen, N.B.; Faseleh Jahromi, M.; Ho, Y.W. Probiotic potential of Lactobacillus strains with antimicrobial activity against some human pathogenic strains. BioMed. Res. Int. 2014, 2014, 927268. [Google Scholar] [CrossRef]
- Reis, N.A.; Saraiva, M.A.F.; Duarte, E.A.A.; de Carvalho, E.A.; Vieira, B.B.; Evangelista-Barreto, N.S. Probiotic properties of lactic acid bacteria isolated from human milk. J. Appl. Microbiol. 2016, 121, 811–820. [Google Scholar] [CrossRef]
- Ali, M.S.; Lee, E.B.; Lim, S.K.; Suk, K.; Park, S.C. Isolation and identification of Limosilactobacillus reuteri PSC102 and evaluation of its potential probiotic, antioxidant, and antibacterial properties. Antioxidants 2023, 12, 238. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Qiao, L.; Liu, R.; Yao, H.; Gao, C. Potential probiotic properties of lactic acid bacteria isolated from the intestinal mucosa of healthy piglets. Ann. Microbiol. 2017, 67, 239–253. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum strains isolated from mozzarella cheese: Probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Evaluation of lactic acid bacteria from kefir, molasses and olive brine as possible probiotics based on physiological properties. Ann. Microbiol. 2008, 58, 661–670. [Google Scholar] [CrossRef]
- MacKenzie, D.A.; Jeffers, F.; Parker, M.L.; Vibert-Vallet, A.; Bongaerts, R.J.; Roos, S.; Walter, J.; Juge, N. Strain-specific diversity of mucus-binding proteins in the adhesion and aggregation properties of Lactobacillus reuteri. Microbiology 2010, 156, 3368–3378. [Google Scholar] [CrossRef] [PubMed]
- Chaffanel, F.; Charron-Bourgoin, F.; Soligot, C.; Kebouchi, M.; Bertin, S.; Payot, S.; Le Roux, Y.; Leblond-Bourget, N. Surface proteins involved in the adhesion of Streptococcus salivarius to human intestinal epithelial cells. Appl. Microbiol. Biotechnol. 2018, 102, 2851–2865. [Google Scholar] [CrossRef]
- González, L.; Sandoval, H.; Sacristán, N.; Castro, J.M.; Fresno, J.M.; Tornadijo, M.E. Identification of lactic acid bacteria isolated from Genestoso cheese throughout ripening and study of their antimicrobial activity. Food Control 2007, 18, 716–722. [Google Scholar] [CrossRef]
- Šušković, J.; Kos, B.; Beganović, J.; Leboš Pavunc, A.; Habjanič, K.; Matošić, S. Antimicrobial activity–the most important property of probiotic and starter lactic acid bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Davoodabadi, A.; Dallal, M.M.S.; Foroushani, A.R.; Douraghi, M.; Harati, F.A.; Amin Harati, F. Antibacterial activity of Lactobacillus spp. isolated from the feces of healthy infants against enteropathogenic bacteria. Anaerobe 2015, 34, 53–58. [Google Scholar] [CrossRef]
- Frece, J.; Markov, K.; Kovačević, D. Determination of indigenous microbial populations, mycotoxins and characterization of potential starter cultures in Slavonian kulen. Meso Prvi Hrvatski Cas. Mesu 2010, 12, 92–99. [Google Scholar]
- Zhang, Q. Lactic Acid Bacteria and Bacteriocins. Lactic Acid Bacteria; Springer: Berlin/Heidelberg, Germany, 2019; pp. 61–91. [Google Scholar]
- Ratsep, M.; Naaber, P.; Koljalg, S.; Smidt, I.; Shkut, E.; Sepp, E. Effect of Lactobacillus plantarum strains on clinical isolates of Clostridium difficile in vitro. J. Probiotics Health 2014, 2, 1–5. [Google Scholar] [CrossRef]
- Mahmoudi, I.; Telmoudi, A.; Hassouna, M. Beneficial effects of probiotic Lactobacillus plantarum isolated from cow, goat and sheep raw milks. Acta Sci. Microbiol. 2018, 1, 17–20. [Google Scholar] [CrossRef]
- Kim, H.S.; Chae, H.S.; Jeong, S.G.; Ham, J.S.; Im, S.K.; Ahn, C.N.; Lee, J.M. In Vitro antioxidative properties of lactobacilli. Asian-Australas. J. Anim. Sci. 2006, 19, 262–265. [Google Scholar] [CrossRef]
- Pieniz, S.; Andreazza, R.; Anghinoni, T.; Camargo, F.; Brandelli, A. Probiotic potential, antimicrobial and antioxidant activities of Enterococcus durans strain LAB18s. Food Control 2014, 37, 251–256. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y. Probiotic and anti-inflammatory potential of Lactobacillus rhamnosus 4B15 and Lactobacillus gasseri 4M13 isolated from infant feces. PLoS ONE 2018, 13, e0192021. [Google Scholar] [CrossRef]
- Badel, S.; Bernardi, T.; Michaud, P. New perspectives for lactobacilli exopolysaccharides. Biotechnol. Adv. 2011, 29, 54–66. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent developments in the biosynthesis and applications of heteropolysaccharides from lactic acid bacteria. Int. Dairy J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Farag, M.M.S.; Moghannem, S.A.M.; Shehabeldine, A.M.; Azab, M.S. Antitumor effect of exopolysaccharide produced by Bacillus mycoides. Microb. Pathog. 2020, 140, 103947. [Google Scholar] [CrossRef] [PubMed]
- Oleksy-Sobczak, M.; Klewicka, E.; Piekarska-Radzik, L. Exopolysaccharides production by Lactobacillus rhamnosus strains—Optimization of synthesis and extraction conditions. Lebensm. Wiss. Technol. 2020, 122, 109055. [Google Scholar] [CrossRef]
- Rendueles, O.; Kaplan, J.B.; Ghigo, J.M. Antibiofilm polysaccharides. Environ. Microbiol. 2013, 15, 334–346. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Origin | No. Accession | Gram Staining | Cell Morphology | |
|---|---|---|---|---|---|
| Lacticaseibacillus rhamnosus 2 | S1E | Infant feces | OR636880 | + | rod |
| Limosilactobacillus reuteri 1 | S2A | Infant feces | OR636881 | + | rod |
| Limosilactobacillus reuteri 2 | S2C | Infant feces | OR636882 | + | rod |
| Lacticaseibacillus rhamnosus 1 | S4F | Infant feces | OR636883 | + | rod |
| Lacticaseibacillus rhamnosus 1 | M6B | Breast milk | OR636884 | + | rod |
| Lactobacillus gasseri 1 | M6D | Breast milk | OR636885 | + | rod |
| Lacticaseibacillus rhamnosus 3 | S6C | Infant feces | OR636886 | + | rod |
| Lacticaseibacillus rhamnosus 2 | M8A | Breast milk | OR636887 | + | rod |
| Lactobacillus gasseri 1 | M8C | Breast milk | OR636888 | + | rod |
| Isolates | CN | E | CLI | S | Q/D | K | C | VA | AMP | TE | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lacticaseibacillus rhamnosus 2 | S1E | I | S | S | R | S | R | S | R | I | I |
| Limosilactobacillus reuteri 1 | S2A | I | S | S | R | I | R | I | R | I | I |
| Limosilactobacillus reuteri 2 | S2C | I | S | S | R | S | R | S | R | I | I |
| Lacticaseibacillus rhamnosus 1 | S4F | R | S | S | R | S | R | S | R | I | I |
| Lacticaseibacillus rhamnosus 1 | M6B | I | S | I | R | I | R | S | R | I | I |
| Lactobacillus gasseri 1 | M6D | I | S | S | R | S | R | S | R | I | I |
| Lacticaseibacillus rhamnosus 3 | S6C | R | S | S | R | S | R | S | R | I | I |
| Lacticaseibacillus rhamnosus 2 | M8A | R | S | S | R | S | R | S | R | S | S |
| Lactobacillus gasseri 1 | M8C | I | S | S | R | S | R | S | R | I | S |
| Enzyme | Isolates | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| S1E | S2A | S2C | S4F | M6B | M6D | S6C | M8A | M8C | |
| Alkaline phosphatase | − | − | − | − | − | − | − | − | − |
| Esterase (C4) | + | + | + | + | + | + | + | + | + |
| Lipase esterase (C8) | + | + | + | + | + | + | + | + | + |
| Lipase (C14) | − | − | − | − | − | − | − | − | − |
| Leucin arylamidase | + | + | + | + | + | + | + | + | + |
| Valin arylamidase | + | − | + | + | + | + | + | + | + |
| Cystine arylamidase | + | − | + | + | + | + | + | − | + |
| Trypsin | − | − | − | − | − | − | − | − | − |
| α-chymotrypsin | − | − | − | − | − | − | − | − | − |
| Acid phosphatase | + | + | + | + | + | + | + | + | + |
| Naphtol-AS-BI-fosfohydrolase | + | + | + | + | + | + | + | + | + |
| α-galaktosidase | + | + | + | + | + | + | + | + | + |
| β-galaktosidase | + | + | + | + | + | + | + | + | + |
| β-glukuronidase | − | − | − | − | − | − | − | − | − |
| α-glukosidase | + | + | + | + | − | + | − | + | + |
| β-glukosidase | + | + | + | + | + | + | + | + | + |
| N-acetyl-β-glukosaminidase | − | − | − | − | − | − | − | − | − |
| α-mannosidase | − | − | − | − | − | − | − | − | − |
| α-fucosidase | − | − | − | − | − | − | − | − | − |
| Isolates | Hydrophobicity (%) | DPPH (%) | Adherence (%) | Bile Salt Tolerance (%) | ||
|---|---|---|---|---|---|---|
| 1.5 g/L | 3.0 g/L | |||||
| Lacticaseibacillus rhamnosus 2 | S1E | 46.1 ± 7.5 a | 34.9 ± 4.1 a | 0.5 ± 0.2 a | 89.8 ± 1.5 a | 89.6 ± 1.0 a |
| Limosilactobacillus reuteri 1 | S2A | 15.4 ± 5.3 bd | 39.3 ± 9.2 a | 3.8 ± 0.6 b | 71.7 ± 3.0 b | 41.5 ± 2.0 b |
| Limosilactobacillus reuteri 2 | S2C | 9.8 ± 4.6 b | 40.4 ± 4.3 a | 3.9 ± 1.0 b | 67.2 ± 3.5 b | 32.0 ± 0.5 c |
| Lacticaseibacillus rhamnosus 1 | S4F | 31.3 ± 5.2 eh | 69.3 ± 2.5 b | 0.3 ± 0.0 a | 87.8 ± 0.5 a | 85.4 ± 1.5 a |
| Lacticaseibacillus rhamnosus 1 | M6B | 12.3 ± 3.8 eg | 52.1 ± 4.7c | 0.4 ± 0.1 a | 78.9 ± 1.5 c | 49.1 ± 3.0 d |
| Lactobacillus gasseri 1 | M6D | 31.2 ± 5.8 dgh | 42.6 ± 4.6 a | 0.4 ± 0.02 a | 78.6 ± 1.0 c | 64.5 ± 3.0 e |
| Lacticaseibacillus rhamnosus 3 | S6C | 27.8 ± 2.3 b | 69.3 ± 2.1 c | 0.4 ± 0.01 a | 84.6 ± 7.5 a | 69.9 ± 1.5 e |
| Lacticaseibacillus rhamnosus 2 | M8A | 50.8 ± 5.5 af | 20.4 ± 1.6 d | 0.8 ± 0.2 a | 95.5 ± 4.0 d | 48.4 ± 4.0 d |
| Lactobacillus gasseri 1 | M8C | 57.1 ± 8.9 cf | 18.4 ± 7.6 d | 6.0 ± 0.3 c | 72.2 ± 4.0 c | 69.0 ± 1.5 e |
| Isolates | Production of Exopolysaccharides | |||
|---|---|---|---|---|
| Glucose | Lactose | Sucrose | ||
| Lacticaseibacillus rhamnosus 2 | S1E | + | + | + |
| Limosilactobacillus reuteri 1 | S2A | + | + | + |
| Limosilactobacillus reuteri 2 | S2C | + | + | + |
| Lacticaseibacillus rhamnosus 1 | S4F | + | + | + |
| Lacticaseibacillus rhamnosus 1 | M6B | + | + | + |
| Lactobacillus gasseri 1 | M6D | + | + | + |
| Lacticaseibacillus rhamnosus 3 | S6C | + | + | + |
| Lacticaseibacillus rhamnosus 2 | M8A | + | + | + |
| Lactobacillus gasseri 1 | M8C | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyrslova, I.; Drab, V.; Cihlar, J.; Krausova, G.; Mrvikova, I.; Kana, A.; Stetina, J.; Musilova, S. Functional and Probiotic Characterization of Newly Isolated Strains from Infant Feces and Breast Milk. Fermentation 2023, 9, 960. https://doi.org/10.3390/fermentation9110960
Hyrslova I, Drab V, Cihlar J, Krausova G, Mrvikova I, Kana A, Stetina J, Musilova S. Functional and Probiotic Characterization of Newly Isolated Strains from Infant Feces and Breast Milk. Fermentation. 2023; 9(11):960. https://doi.org/10.3390/fermentation9110960
Chicago/Turabian StyleHyrslova, Ivana, Vladimir Drab, Jaromir Cihlar, Gabriela Krausova, Iva Mrvikova, Antonin Kana, Jiri Stetina, and Sarka Musilova. 2023. "Functional and Probiotic Characterization of Newly Isolated Strains from Infant Feces and Breast Milk" Fermentation 9, no. 11: 960. https://doi.org/10.3390/fermentation9110960